Гаплогруппа L2
| Гаплогруппа L2 | |
|---|---|
| Возможное время происхождения | 80 000–111 100 английских фунтов [1] |
| Возможное место происхождения | Западная Африка [2] или Центральная Африка |
| Предок | L2─6 (L2'3'4'6) |
| Потомки | L2a─d, L2e |
| Определение мутаций | 146, 150, 152, 2416, 8206, 9221, 10115, 13590, 16311!, 16390 [3] |
Гаплогруппа L2 — это гаплогруппа митохондриальной ДНК (мтДНК) человека, широко распространенная в современном мире, особенно в субэкваториальной Африке. Его субклад L2a представляет собой довольно частый и широко распространенный кластер мтДНК на континенте, а также среди кластеров в Америке .
Источник
[ редактировать ]L2 — распространенная линия в Африке. Считается, что он возник между 87 000 и 107 000 лет назад. [4] или ок. 90 000 йен. [1] Его возраст, а также широкое распространение и разнообразие по всему континенту затрудняют с уверенностью проследить его точную точку происхождения в Африке. [5] Несколько гаплотипов L2, наблюдаемых у гвинейцев и других популяций Западной Африки, имели генетические совпадения с Восточной Африкой и Северной Африкой . [6] Происхождение L2b, L2c, L2d и L2e в Западной или Центральной Африке кажется вероятным. [5] Раннее разнообразие L2 можно наблюдать по всему Африканскому континенту, но, как мы видим в разделе «Субклады» ниже, наибольшее разнообразие наблюдается в Западной Африке . Большинство субкладов в основном приурочено к Западной и западно-Центральной Африке. [7]
Согласно исследованию 2015 года, «результаты показывают, что линии Южной Африки группируются с линиями Западной/Центральной Африки в недавнем временном масштабе, тогда как восточные линии кажутся значительно более древними. Три момента распространения из центральноафриканского источника связаны с L2: одна миграция 70–50 тыс. лет назад в Восточную или Южную Африку, послеледниковые перемещения 15–10 тыс. лет назад в Восточную Африку и экспансия банту на юг за последние 5 тыс. лет назад. Дополнительный популяционный и филогеографический анализ L0a не указывает на убедительные доказательства наличия гена мтДНК; поток между восточным и южным населением во время более позднего перемещения, что позволяет предположить низкую степень смешения между восточноафриканским населением и мигрантами банту. Это означает, что, по крайней мере на ранних стадиях, экспансия банту была в основном демической диффузией с небольшим включением местного населения». [8]
Распределение
[ редактировать ]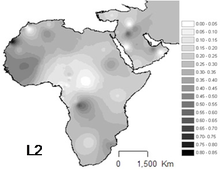
L2 — наиболее распространенная гаплогруппа в Африке, и она наблюдается по всему континенту. Он встречается примерно у трети африканцев и их недавних потомков.
Самая высокая частота встречается среди пигмеев мбути (64%). [9] Также сильное присутствие среди населения Западной Африки в Сенегале (43-54%). [6] Также важно для популяций, не принадлежащих к банту, в Восточной Африке (44%), [10] в Судане и Мозамбике .
Он особенно распространен в Чаде и Канембу (38% выборки), но также относительно часто встречается у кочевников арабов- (33%). [11] [5] и аканцы (~33%) [12]
Субклады
[ редактировать ]
|
L2 имеет пять основных подгаплогрупп: L2a, L2b, L2c, L2d и L2e. Из этих линий наиболее распространенным субкладом является L2a, который встречается в Африке, Леванте и в Америке . [13]
Гаплогруппа L2 наблюдалась среди образцов на островном кладбище в Кулубнарти , Судан , которые датируются раннехристианским периодом (550–800 гг. н.э.). [14]
Гаплогруппа L2a
[ редактировать ]L2a широко распространен в Африке и является наиболее распространенной и широко распространенной африканской гаплогруппой к югу от Сахары, а также довольно часто встречается - 19% в Америке среди потомков континента АфроЕвразия, используя теорию или модель выхода из Африки. (Салас и др., 2002). L2a имеет возможную дату происхождения ок. 48 000 йен. [1]
Он особенно распространен в Чаде (38% выборки; 33% недифференцированного L2 среди арабов Чада , [15] ), а в популяциях, не принадлежащих к банту, в Восточной Африке ( Кения , Уганда и Танзания ) - 38%. [10] Около 33% в Мозамбике [16] и 32% в Гане . [12]
Этот субклад характеризуется мутациями 2789, 7175, 7274, 7771, 11914, 13803, 14566 и 16294. Он составляет 52% от общего количества L2 и является единственным субкладом L2, широко распространенным по всей Африке. [17]
Широкое распространение L2a и разнообразие затрудняют определение географического происхождения. Основная загадка — это почти повсеместная гаплогруппа L2a, которая, возможно, распространилась на восток и запад вдоль коридора Юго-Восточной Африки после последнего ледникового максимума , или истоки этих экспансий могут лежать раньше, в начале позднего каменного века ~ 40 000 г. много лет назад. [5] [17]
В Восточной Африке L2a был обнаружен в 15% в долине Нила ( Нубия) , в 5% у египтян , в 14% у на кушитском языке говорящих , в 15% у семитских людей амхара , в 10% у гураге , в 6% у в 13 % у эфиопов. тыграев-тигринья и йеменцев % 5 [16]
Гаплогруппа L2a также появляется в Северной Африке , с наибольшей частотой у туарегов 20% , фулани (14%). Встречается также среди некоторых арабов Алжира , он встречается в 10% среди марокканских арабов , некоторых марокканских берберов и тунисских берберов. [18] [19]
назначают препарат ставудин У пациентов, которым для лечения ВИЧ , гаплогруппа L2a связана с меньшей вероятностью периферической нейропатии как побочного эффекта. [20]
Гаплогруппа L2a1
[ редактировать ]L2a может быть далее разделена на L2a1, содержащую переход в 16309 (Salas et al. 2002). [21]
Этот субклад с различной частотой наблюдается в Западной Африке среди малинке , волофов и других; среди жителей Северной Африки ; в Сахеле у хауса , фульбе и других; в Центральной Африке у бамилеке , тикара , фали и других; в Южной Африке среди семьи койсан , включая носителей языка хве и банту ; и в Восточной Африке у кикуйю из Кении . [21]
Все клады L2, присутствующие в Эфиопии , в основном произошли от двух субкладов: L2a1 и L2b. L2a1 определяется мутациями в 12693, 15784 и 16309. Большинство эфиопских последовательностей L2a1 имеют общие мутации в nps 16189 и 16309. Однако, тогда как большинство (26 из 33) афроамериканцев имеют общие полные последовательности гаплогруппы L2a, можно разделить на четыре субклада путем замен. в nps L2a1e-3495, L2a1a-3918, L2a1f-5581 и L2a1i-15229. Ни одна из этих последовательностей не наблюдалась в образцах 16309 L2a1 из Эфиопии. (Салас 2002) и др. [21]
Гаплогруппа L2a1 также наблюдалась среди махра (4,6%). [22]
Гаплогруппа L2a1 была обнаружена в древних окаменелостях, связанных с докерамической неолитической культурой в Телль-Халуле , Сирия . [23] Образец, раскопанный на стоянке пасторального неолита Саванны в Люксманде в Танзании, также содержал кладу L2a1. Анализ кластеризации примесей также показал, что этот человек имел значительное происхождение из древнего Леванта, подтверждая родовые связи между создателями пасторального неолита саванны и докерамического неолита. [24]
Гаплогруппа L2a1a
[ редактировать ]Субклад L2a1a определяется заменами в позициях 3918, 5285, 15244 и 15629. Есть два кластера L2a, которые хорошо представлены у жителей юго-восточной Африки, L2a1a и L2a1b, оба определяемые переходами в довольно стабильных положениях HVS-I. Оба они, по-видимому, происходят из Западной Африки или Северо-Западной Африки (на что указывает распределение совпадающих или соседних типов) и претерпели резкое распространение либо в Юго-Восточной Африке, либо в популяции, являющейся предком современных жителей Юго-Восточной Африки. .
Совсем недавние звездные вспышки в субкладах L2a1a и L2a2 предполагают признак экспансии банту, что также было предложено Перейрой и др. (2001).
L2a1a определяется мутацией по номеру 16286. Кандидат в основатели L2a1a датируется 2700 (SE 1200) лет назад. (Перейра и др., 2001). Однако L2a1a, определенный заменой в (np 16286) (Salas et al. 2002), теперь поддерживается маркером кодирующей области (np 3918) (рис. 2A) и был обнаружен в четырех из шести йеменских линий L2a1. . L2a1a наиболее часто встречается в Юго-Восточной Африке (Перейра и др., 2001; Салас и др., 2002). Как частый гаплотип-основатель, так и производные линии (с мутацией 16092), обнаруженные среди йеменцев, имеют точные совпадения с последовательностями Мозамбика (Перейра и др., 2001; Салас и др., 2002). L2a1a также встречается с меньшей частотой в Северо-Западной Африке, среди мор и бамбара в Мали и Мавритании . [25] (Рандо и др., 1998 г.; Мака-Мейер и др., 2003 г.)
L2a1a присутствует у населения США. [13]
Гаплогруппа L2a1a1
[ редактировать ]L2a1a1 определяется маркерами 6152C, 15391T и 16368C. Его нашли в США [13] а также Бразилия, [26] Ливия, Сьерра-Леоне, Ангола, [27] Замбия, [28] и другие страны.
Гаплогруппа L2a1b
[ редактировать ]L2a1b определяется заменами в позициях 16189 и 10143. 16192 также часто встречается в L2a1b и L2a1c; он появляется в Северной Африке в Египте . Он также появляется в Юго-Восточной Африке и поэтому также может быть маркером экспансии банту . [5] Южноафриканский вариант в основном приписывается L2a1b1, являющемуся ветвью L2a1b, он встречается в Южной Африке , Мозамбике , Кении и Кувейте . [13]
Гаплогруппа L2a1c
[ редактировать ]L2a1c часто разделяет мутацию 16189 с L2a1b, но имеет свои собственные маркеры 3010 и 6663. 16192 также часто встречается в L2a1b и L2a1c; он появляется в Юго-Восточной Африке, а также в Восточной Африке. [29] Это предполагает некоторую диверсификацию этой клады in situ.
Позиции T16209C C16301T C16354T на вершине L2a1 определяют небольшой субклад, названный Kivisild et al. L2a1c. (2004, Рисунок 3) (см. также Рисунок 6 в Salas et al. 2002), который в основном встречается в Восточной Африке (например, Судан , Нубия , Эфиопия ), среди Туркана и Западной Африки (например, Канури ).
В бассейне Чада были идентифицированы четыре различных типа L2a1c на одну или две мутационные ступени от восточно- и западноафриканских типов. (Кивисилд и др.) 2004. [29] (цитата на стр.9 или 443) [11] [30]
Было показано, что L2a1c присутствует в Чаде , Габоне , Испании и США. [13]
Гаплогруппа L2a1c1
[ редактировать ]L2a1c1 имеет североафриканское происхождение. [30] Он определяется маркерами 198, 930, 3308, 8604, 16086. Наблюдается у тунисцев, марокканцев, египтян, нубийцев и йеменцев. Филиалы L2a1c1 присутствуют в Нигерии , Сенегале и Замбии . [31]
Гаплогруппа L2a1c2
L2a1c2 является ветвью L2a1c1. Возникает в Северной Африке, в основном наблюдается у североафриканских евреев и других жителей Северной Африки. Филиалы также присутствуют в Латинской Америке и Западной Африке из-за примеси Северной Африки.
Гаплогруппа L2a1d
[ редактировать ]L2a1d определяется мутациями T5196C, T9530C, T11386C, A12612G и C13934T. Его нашли в Объединенных Арабских Эмиратах. [32] и Кувейт . Также L2a1d встречается в Бенине . [13]
Гаплогруппа L2a1d1
[ редактировать ]L2a1d1 — восточноафриканская ветвь. [8] который был выявлен у населения в Сомали [33] и Судан. [34] L2a1d1 также был обнаружен в Эфиопии и Египте . [13]
Гаплогруппа L2a1d2
[ редактировать ]L2a1d2 связан с Африкой к югу от Сахары, включая южную часть Африки ( Мозамбик , [35] Народ мбунда и народ бемба в Замбии . [36] Койсанские народы, включая Тшва Сан [37] ).
Гаплогруппа L2a1e
[ редактировать ]L2a1e определяется мутациями C3495A, G8790A и G12630A. Он был обнаружен в Бразилии , Гренаде и среди афроамериканцев. [13]
Гаплогруппа L2a1e1
[ редактировать ]L2a1e1 был обнаружен в Северной и Южной Америке в Бразилии . [38] Он также встречается в Нигерии , карибских популяциях западноафриканского происхождения ( Ямайка , Доминика и Барбадос ) и среди афроамериканцев.
Гаплогруппа L2a1f
[ редактировать ]L2a1f наблюдается у потомков баконго, проживающих в США и Доминиканской Республике, у хосианцев в Южной Африке, Омане, [39] Замбия и Мадагаскар, он также был обнаружен в Буркина-Фасо и Омане, а также в Северной и Южной Америке .
Гаплогруппа L2a1g
[ редактировать ]L2a1g определяется мутациями A8014G, C14281T, T16131C, C16225T и C16234T. Это наблюдается на юге Африки среди носителей банту, [2] в том числе в Замбии , Мадагаскаре и Южной Африке .
Гаплогруппа L2a1h
[ редактировать ]Было показано, что L2a1h присутствует в Израиле и Кении . [13]
Гаплогруппа L2a1k
[ редактировать ]L2a1k определяется маркерами G6722A и T12903C. Описанный как специфичный для Европы, он ранее назывался субкладом L2a1a и был обнаружен у чехов , словаков , хорватов , сербов и болгар . [40] [41]
Гаплогруппа L2a1l
[ редактировать ]L2a1l определяется мутацией C534T. Он появляется в Алжире , Сьерра-Леоне (среди народа менде ) и Гамбии (среди народа волоф ) и присутствовал в древней Испании. [42]
Гаплогруппа L2a1l1
[ редактировать ]L2a1l1 наблюдается у нуна [43] и Мосси [44] Народы Буркина-Фасо .
Гаплогруппа L2a1l2
[ редактировать ]L2a1l2 присутствует у людей в различных частях Западной Африки, Северной Африки и Западной Европы, включая людей мандинка из Гвинеи-Бисау , людей фула из Гамбии и людей пана из Буркина-Фасо . [42]
Гаплогруппа L2a1l2a
[ редактировать ]L2a1l2a признана « специфичной для ашкенази » гаплогруппой, встречающейся среди евреев-ашкенази, имеющих происхождение из Центральной и Восточной Европы. Он также был обнаружен в небольших количествах у якобы нееврейского населения Польши, где, как предполагается, он произошел от примеси ашкенази. [45] Однако этот гаплотип составляет лишь очень небольшую часть митохондриальных линий ашкенази; различные исследования (включая исследование Бехара) установили, что его частота составляет 1,4–1,6%.
Гаплогруппа L2a1l3
[ редактировать ]L2a1l3 определяется мутациями G14905A и T16357C. Он встречается у народа йоруба в Нигерии и Алжире .
Гаплогруппа L2a1m
[ редактировать ]L2a1m определяется мутацией A13884G. Его нашли в Буркина-Фасо. [46] и Гаити, а также Оман , Йемен , Саудовская Аравия и Израиль . [13]
Гаплогруппа L2a1m1
[ редактировать ]L2a1m1 был обнаружен среди афроамериканцев и доминиканцев .
Гаплогруппа L2a1m1a
[ редактировать ]L2a1m1a был обнаружен среди афроамериканцев и на Сент-Винсенте и Гренадинах .
Гаплогруппа L2a1n
[ редактировать ]L2a1n наблюдается среди народа йоруба в Нигерии и народа моси в Буркина-Фасо , а также был обнаружен в Камеруне и Португалии на Пиренейском полуострове . [13]
Гаплогруппа L2a1o
[ редактировать ]L2a1o определяется мутацией T12438C. Он был обнаружен в Сирии и Буркина-Фасо . [46]
Гаплогруппа L2a1p
[ редактировать ]L2a1p определяется мутациями A9410G, T13818C и C15626T. Встречается в Нигерии , у народа Кассена в Буркина-Фасо и у марокканских евреев . Он также присутствует в Соединенных Штатах Америки . [13]
Гаплогруппа L2a2
[ редактировать ]L2a2 характерен для пигмеев мбути . [9]
Гаплогруппа L2b'c
[ редактировать ]L2b'c, вероятно, развился около 62 000 лет назад. [1]
Гаплогруппа L2b
[ редактировать ]Этот субклад преимущественно встречается в Западной Африке , но распространен и по всей Африке. [47] ветви L2b также включают L2b1a1, который встречается в Лигурии, Италия, и L2b3, который встречается в Галисии (Испания) . [13]
Гаплогруппа L2c
[ редактировать ]L2c наиболее часто встречается в Западной Африке и, возможно, возник там. [17] Особенно присутствует в Сенегале (39%), Кабо-Верде (16%) и Гвинее-Бисау (16%). [6] Также известно, что ответвления L2c можно найти в Европе в таких областях, как Пиренейский полуостров в Андалусии, Испания , Каталония , Испания , а также в континентальной северо- западной Европе в Нидерландах . [13]
Гаплогруппа L2d
[ редактировать ]L2d наиболее часто встречается в Западной Африке, где он, возможно, и возник. [17] Он также встречается в Йемене, Мозамбике и Судане. [16]
Гаплогруппа L2e
[ редактировать ]L2e (бывший L2d2) типичен для Западной Африки . [5] Встречается он и в Тунисе . [48] и среди мандинка из Гвинеи-Бисау и афроамериканцев. [47]
Дерево
[ редактировать ]
Это филогенетическое древо субкладов гаплогруппы L2 основано на статье Манниса ван Овена и Манфреда Кайзера. Обновленное комплексное филогенетическое древо глобальных вариаций митохондриальной ДНК человека. [3] и последующие опубликованные исследования.
- Самый недавний общий предок (MRCA)
- Л1'2'3'4'5'6
- Л2'3'4'6
- Л2
- L2a'b'c'd
- Л2а
- Л2а1
- Л2а1а
- Л2а1а1
- Л2а1а2
- Л2а1а2а
- Л2а1а2а1
- L2a1a2b
- Л2а1а2а
- Л2а1а3
- 16189 (16192)
- L2a1b
- L2a1b1
- L2a1f
- L2a1f1
- L2a1b
- 143
- L2a1c
- L2a1c1
- L2a1c2
- L2a1c3
- L2a1c4
- L2a1d
- L2a1e
- L2a1e1
- L2a1h
- 16189
- L2a1i
- L2a1j
- L2a1k
- 16192
- L2a1l
- Л2а1л1
- Л2а1л1а
- Л2а1л2
- Л2а1л1
- L2a1l
- L2a1c
- Л2а1а
- Л2а2
- Л2а2а
- Л2а2а1
- L2a2b
- L2a2b1
- Л2а2а
- Л2а1
- L2b'c
- Л2б
- Л2б1
- L2b1a
- L2b1a2
- L2b1a3
- L2b1a
- Л2б1
- L2c
- L2c2
- L2c2a
- L2c3
- L2c2
- Л2б
- Л2д
- Л2д1
- L2d1a
- Л2д1
- Л2а
- L2e
- L2a'b'c'd
- Л2
- Л2'3'4'6
- Л1'2'3'4'5'6
См. также
[ редактировать ]- Генеалогический ДНК-тест
- Генетическая генеалогия
- Митохондриальная генетика человека
- Популяционная генетика
- Гаплогруппы митохондриальной ДНК человека
Филогенетическое древо гаплогрупп митохондриальной ДНК (мтДНК) человека | |||||||||||||||||||||||||||||||||||||||
| Митохондриальная Ева ( L ) | |||||||||||||||||||||||||||||||||||||||
| Л0 | Л1–6 | ||||||||||||||||||||||||||||||||||||||
| Л1 | Л2 | Л3 | Л4 | Л5 | Л6 | ||||||||||||||||||||||||||||||||||
| М | Н | ||||||||||||||||||||||||||||||||||||||
| Чехия | Д | И | Г | вопрос | ТО | А | С | Р | я | В | Х | И | |||||||||||||||||||||||||||
| С | С | Б | Ф | Р0 | до JT | П | В | ||||||||||||||||||||||||||||||||
| Х.В. | Джей Ти | К | |||||||||||||||||||||||||||||||||||||
| ЧАС | V | Дж | Т | ||||||||||||||||||||||||||||||||||||
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Соареш, Педро; Лука Эрмини; Ноэль Томсон; Мару Мормина; Тереза Рито; Арне Рёль; Антонио Салас; Стивен Оппенгеймер; Винсент Маколей; Мартин Б. Ричардс (4 июня 2009 г.). «Поправка на очищающий отбор: улучшенные митохондриальные молекулярные часы человека» . Американский журнал генетики человека . 84 (6): 82–93. дои : 10.1016/j.ajhg.2009.05.001 . ПМЦ 2694979 . ПМИД 19500773 .
- ^ Перейти обратно: а б Сильва, Марина (2015). «60 000 лет взаимодействия между Центральной и Восточной Африкой, задокументированные основной африканской митохондриальной гаплогруппой L2» . Научные отчеты . 5 . Природа: 12526. Бибкод : 2015NatSR...512526S . дои : 10.1038/srep12526 . ПМЦ 4515592 . ПМИД 26211407 .
- ^ Перейти обратно: а б ван Овен, Маннис; Манфред Кайзер (13 октября 2008 г.). «Обновленное комплексное филогенетическое древо глобальных вариаций митохондриальной ДНК человека» . Человеческая мутация . 30 (2): E386–E394. дои : 10.1002/humu.20921 . ПМИД 18853457 . S2CID 27566749 .
- ^ Гондер, Мэри Кэтрин; Мортенсен, Холли М.; Рид, Флойд А.; де Соуза, Александра; Тишкофф, Сара А. (2007). «Анализ последовательности генома всей мтДНК древних африканских линий» . Молекулярная биология и эволюция . 24 (3): 757–768. дои : 10.1093/molbev/msl209 . ПМИД 17194802 .
- ^ Перейти обратно: а б с д и ж Салас, Антонио и др., Создание африканского ландшафта мтДНК , Американский журнал генетики человека , том. 71, нет. 5 (2002), стр. 1082–1111.
- ^ Перейти обратно: а б с Роза, Александра; Брем, А; Кивисилд, Т; Метспалу, Э; Виллемс, Р; и др. (2004). «Профиль мтДНК жителей Западной Африки: на пути к лучшему пониманию региона Сенегамбии». Анналы генетики человека . 68 (Часть 4): 340–352. дои : 10.1046/j.1529-8817.2004.00100.x . hdl : 10400.13/3044 . ПМИД 15225159 . S2CID 15391342 .
- ^ Атлас человеческого пути: гаплогруппа L2. Архивировано 6 октября 2011 г. в Wayback Machine. Генографический проект, National Geographic.
- ^ Перейти обратно: а б Сильва, Марина; Альшамали, Фарида; Сильва, Паула; Каррильо, Карла; Мандлате, Флавио; Хесус Тровоада, Мария; Черный, Виктор; Перейра, Луиза; Соареш, Педро (2015). «60 000 лет взаимодействия между Центральной и Восточной Африкой, задокументированные основной африканской митохондриальной гаплогруппой L2» . Научные отчеты . 5 : 12526. Бибкод : 2015NatSR...512526S . дои : 10.1038/srep12526 . ПМЦ 4515592 . ПМИД 26211407 .
- ^ Перейти обратно: а б Кинтана-Мурси и др. 2008. Материнские следы глубокого общего происхождения и асимметричного потока генов между пигмеями-охотниками-собирателями и фермерами, говорящими на языке банту, «Труды Национальной академии наук Соединенных Штатов Америки». 105(5): 1599
- ^ Перейти обратно: а б Сэди Андерсон-Манн 2006, Филогенетический и филогеографический анализ вариаций африканской митохондриальной ДНК. Архивировано 10 сентября 2011 г. в Wayback Machine.
- ^ Перейти обратно: а б Черный, В.; и др. (июль 2007 г.). «Двунаправленный коридор в поясе Сахель-Судан и отличительные особенности популяций бассейна Чада: история, раскрытая геномом митохондриальной ДНК» . Анналы генетики человека . 71 (4): 433–452. дои : 10.1111/j.1469-1809.2006.00339.x . ПМИД 17233755 . S2CID 28105243 .
- ^ Перейти обратно: а б Вирама, Кришна Р. и др. 2010, Небольшая генетическая дифференциация по оценкам однородительских маркеров при наличии значительных языковых вариаций у народов региона Кросс-Ривер в Нигерии.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н «Реконструкция древних связей митохондриальной ДНК между Африкой и Европой», Дополнительные данные S2 М. Сересо и др.
- ^ Сирак, Кендра; Френандес, Даниэль; Новак, Марио; Ван Гервен, Деннис; Пинхаси, Рон (2016). «Реферат Межконгресса IUAES 2016 - Сообщество разделено? Выявление генома(ов) сообщества средневекового Кулубнарти с использованием секвенирования следующего поколения» . Тезисы докладов Межконгресса IUAES 2016 . ИУАЭС: 115.
- ^ Сересо, Мария; и др. (2011). «Новое понимание структуры населения бассейна озера Чад, полученное с помощью высокопроизводительного генотипирования митохондриальной ДНК, кодирующей SNP» . ПЛОС ОДИН . 6 (4): e18682. Бибкод : 2011PLoSO...618682C . CiteSeerX 10.1.1.291.8871 . дои : 10.1371/journal.pone.0018682 . ПМК 3080428 . ПМИД 21533064 .
- ^ Перейти обратно: а б с Тоомас Кивисилд и др., Эфиопское наследие митохондриальной ДНК: отслеживание потока генов через и вокруг ворот слез, Американский журнал генетики человека , том. 75, нет. 5 (ноябрь 2004 г.), стр. 101-1. 752–770.
- ^ Перейти обратно: а б с д Антонио Торрони и др., Эволюционируют ли четыре клады гаплогруппы L2 мтДНК с разной скоростью?, Американский журнал генетики человека , том. 69 (2001), стр. 348–1356.
- ^ Уотсон, Э.; и др. (сентябрь 1997 г.). «Митохондриальные следы человеческой экспансии в Африке» . Американский журнал генетики человека . 61 (3): 691–704. дои : 10.1086/515503 . ПМЦ 1715955 . ПМИД 9326335 .
- ^ Бдительный, Л.; и др. (27 сентября 1991 г.). «Африканские популяции и эволюция митохондриальной ДНК человека». Наука . 253 (5027): 1503–1507. Бибкод : 1991Sci...253.1503V . дои : 10.1126/science.1840702 . ПМИД 1840702 .
- ^ Кампира, Э; Кумвенда, Дж; ван Остерхаут, Джей-Джей; Дандара, К. (август 2013 г.). «Субгаплогруппы митохондриальной ДНК L0a2 и L2a изменяют восприимчивость к периферической нейропатии у взрослых малавийцев, получающих высокоактивную антиретровирусную терапию, содержащую ставудин» . J Приобретенный иммунодефицитный синдром . 63 (5): 647–652. дои : 10.1097/QAI.0b013e3182968ea5 . ПМК 3815091 . ПМИД 23614993 .
- ^ Перейти обратно: а б с Салас А., Ричардс М., Де ла Фе Т., Лареу М.В., Собрино Б., Санчес-Дис П., Маколей В., Карраседо А. Создание африканского ландшафта мтДНК. Ам Джей Хум Жене. Ноябрь 2002 г.;71(5):1082-111. дои: 10.1086/344348. Epub 2002, 22 октября. PMID 12395296; PMCID: PMC385086.
- ^ Нет, Эми. «АНАЛИЗ ГЕНЕТИЧЕСКИХ ДАННЫХ В МЕЖДИСЦИПЛИНАРНЫХ РАМКАХ ДЛЯ ИЗУЧЕНИЯ НЕДАВНЕЙ ЭВОЛЮЦИОННОЙ ИСТОРИИ ЧЕЛОВЕКА И СЛОЖНЫХ ЗАБОЛЕВАНИЙ» (PDF) . Университет Флориды. Архивировано из оригинала (PDF) 13 октября 2020 года . Проверено 2 ноября 2016 г. .
- ^ Манко, Жан (2013). Путешествия предков: Заселение Европы от первых путешественников до викингов . Темза и Гудзон. п. 88. ИСБН 978-0500771822 . Проверено 29 сентября 2017 г.
- ^ Скоглунд; и др. (21 сентября 2017 г.). «Реконструкция структуры доисторического населения Африки» . Клетка . 171 (1): 59–71. дои : 10.1016/j.cell.2017.08.049 . ПМЦ 5679310 . ПМИД 28938123 .
- ^ Гонсалес, AM и др. 2006, Вариации митохондриальной ДНК в Мавритании и Мали и их генетическая связь с другими популяциями Западной Африки.
- ^ GenBank Номер доступа : MN894712.1.
- ^ GenBank Номер доступа : KJ185827.1 .
- ^ GenBank Номер доступа : KJ185589.1 .
- ^ Перейти обратно: а б «Интернет-библиотека Wiley | Научно-исследовательские статьи, журналы, книги и справочные материалы» . Архивировано из оригинала 5 августа 2010 г. Проверено 19 мая 2009 г.
- ^ Перейти обратно: а б Ласкаро, Даниэла; Кастеллана, Стефано; Гаспар, Джузеппе; Ромео, Джованни; Сакконе, Сесилия; Аттимонелли, Марселла (2008). «Компиляция RHNumtS: особенности и биоинформатические подходы к обнаружению и количественной оценке человеческого NumtS» . БМК Геномика . 9 : 267. дои : 10.1186/1471-2164-9-267 . ПМЦ 2447851 . ПМИД 18522722 .
- ^ «L2a1c1 MTree» . YFull.com . Проверено 15 декабря 2023 г.
- ^ GenBank Номер доступа : MF437155.1
- ^ GenBank Инвентарный номер : KR135841.1 .
- ^ GenBank Номер доступа : KR135855.1 .
- ^ GenBank Инвентарный номер : KR135884.1 .
- ^ Номер доступа GenBank : KJ185590.1 GenBank Номер доступа : KJ185686.1 GenBank Номер доступа : KJ185955.1 GenBank Номер доступа : KJ185403.1
- ^ GenBank Номер доступа : KC622248.1.
- ^ GenBank Номер доступа : MN894787.1.
- ^ Сересо, Мария и др. «Реконструкция древних связей митохондриальной ДНК между Африкой и Европой». Геномные исследования, том. 22,5 (2012): 821-6. doi:10.1101/gr.134452.111 https://genome.cshlp.org/content/suppl/2012/03/01/gr.134452.111.DC1/Cerezo_GR_2011_L-Europe_manuscript_Supplemental_Data_S2_final_version.pdf
- ^ Малярчук Борис А.; Мирослава Деренко; Мария Перкова; Томаш Гжибовский; Томас Ванечек; Ян Лазур (сентябрь 2008 г.). «Реконструкция филогении африканских линий митохондриальной ДНК у славян» . Европейский журнал генетики человека . 16 (9): 1091–1096. дои : 10.1038/ejhg.2008.70 . ПМИД 18398433 . S2CID 21522848 .
- ^ Давидович, Слободан; Борис Малярчук; Елена М. Алексич; Мирослава Деренко; Владанка Топалович; Андрей Литвинов; Милена Стеванович; Наташа Ковачевич-Груйчич (март 2015 г.). «Перспектива митохондриальной ДНК сербского генетического разнообразия». Американский журнал биологической антропологии . 156 (3): 449–465. дои : 10.1002/ajpa.22670 . ПМИД 25418795 .
- ^ Перейти обратно: а б Брук, Кевин Алан (2022). Материнские генетические линии евреев-ашкеназов . Пресса академических исследований. п. 78. ИСБН 978-1644699843 .
- ^ GenBank Номер доступа : JQ044956.1 .
- ^ GenBank Номер доступа : JQ044919.1 .
- ^ Мельник-Сикорска, Марта; Дака, Патрисия; Малярчук Борис; Деренко, Мирослава; Сконечна, Катажина; Перкова, Мария; Добош, Тадеуш; Гжибовский, Томаш (14 января 2013 г.). «История славян, выведенная на основе полных последовательностей митохондриального генома» . ПЛОС ОДИН . 8 (1): e54360. дои : 10.1371/journal.pone.0054360 .
- ^ Перейти обратно: а б Дополнительная таблица 3 в книге «60 000 лет взаимодействия между Центральной и Восточной Африкой, задокументированной основной африканской митохондриальной гаплогруппой L2», авторы Марина Сильва, Фарида Альшамали и др., 2015 г.
- ^ Перейти обратно: а б Бехар и др., 2008b, «Рассвет человеческого матрилинейного разнообразия», Am J Hum Genet. 2008 г., 9 мая; 82(5): 1130–1140
- ^ Коста, Марта Д. и др. (Апрель 2009 г.), Данные полного секвенирования мтДНК тунисских долгожителей: проверка ассоциации гаплогрупп и «золотой середины» долголетия. ( [1] ) Механизмы старения и развития . 130(4): 222-226, PMID 19133286, doi:10.1016/j.mad.2008.12.001
Внешние ссылки
[ редактировать ]- Яна Логана Гаплогруппа L2 . Сайт митохондриальной ДНК
- Яна Логана L2bcd . Сайт митохондриальной ДНК
- PhyloTree.org Манниса ван Овена - поддерево мтДНК L
- Распространение гаплогруппы L2 по данным National Geographic.
- Сересо М., Ахилли А., Оливьери А., Перего У.А., Гомес-Карбалла А., БризигеллиФ, Лансиони Х., Лопес-Сото М., Карраседо А., Капелли С., Торрони А., Салас А. [2] Реконструкция места исследования древней мтДНК