Гаплогруппа F-M89
| Гаплогруппа F-M89 | |
|---|---|
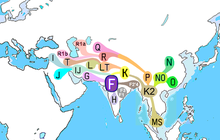 | |
| Возможное время происхождения | 57 500–62 500; (Рагхаван, 2014); [ 1 ] 45 000–55 700 лет назад (Карафет, 2008); [ 2 ] 43 000–56 800 лет назад (Хаммер и Зегура, 2002). [ 3 ] |
| Возможное место происхождения | Западная Азия или Южная Азия [ 4 ] [ 5 ] [ 6 ] [ 7 ] или Юго-Восточная Азия [ 8 ] |
| Предок | CF |
| Потомки | Первичные: F1 , F2 , F3 , GHIJK . |
| Определение мутаций | M89/PF2746, L132.1, M213/P137/страница38, M235/страница80, P14, P133, P134, P135, P136, P138, P139, P140, P141, P142, P145, P146, P148, P149, P151, P157, П158, П159, П160, П161, П163, П166, П187, П316 |
Гаплогруппа F , также известная как F-M89 и ранее как гаплогруппа FT , является очень распространенной гаплогруппой Y-хромосомы . Клада и ее субклады составляют более 90% отцовских линий за пределами Африки.
Подавляющее большинство отдельных мужчин с F-M89 относятся к ее прямому потомку гаплогруппы GHIJK (F1329/M3658/PF2622/YSC0001299). [ 9 ] Помимо GHIJK, гаплогруппа F имеет три других субклада-потомка: F1 (P91/P104), F2 (M427/M428) и F3 (M481). Эти три вместе с F* (M89*) составляют парагруппу F(xGHIJK) . В основном они встречаются в Южной Азии, Юго-Восточной Азии и некоторых частях Восточной Азии.
Ветви гаплогруппы GHIJK впоследствии разделились на двух прямых потомков: G (M201/PF2957) и HIJK (F929/M578/PF3494/S6397). HIJK, в свою очередь, делится на H (L901/M2939) и IJK (F-L15). К потомкам гаплогруппы IJK относятся клады I , J , K и, в конечном итоге, несколько основных гаплогрупп, произошедших от гаплогруппы K, а именно: M , N , O , P , Q , R , S , L и T. гаплогруппы
Происхождение
[ редактировать ]Подсчитано, что SNP M89 появился 38 700–55 700 лет назад и, скорее всего, возник в Западной Азии. [ 2 ] [ 10 ] или Юго-Восточная Азия [ 11 ] Предыдущие исследования также показали, что F-M89, скорее всего, впервые появился на Аравийском полуострове , в Леванте или Северной Африке примерно 43 800–56 800 лет назад. [ 3 ] Было также высказано предположение, что возможное место первого распространения и роста распространенности этой линии, по-видимому, находилось на Индийском субконтиненте или где-то рядом с ним, а большинство субкладов и гаплогрупп-потомков, по-видимому, распространились за пределы Персидского залива. и/или соседние части Ближнего Востока и Юго-Восточной Азии .
Некоторые линии, происходящие от гаплогруппы F-M89, по-видимому, мигрировали обратно в Африку из Западной Азии в доисторические времена. Например, субклады F-M89 были обнаружены в древних образцах ДНК из Судана, которые были связаны как с меройским , так и с постмеройским периодом. [1] Архивировано 25 августа 2022 г. в захоронениях Wayback Machine .
Распределение
[ редактировать ]Подавляющее большинство ныне живущих особей, несущих F-M89, принадлежат к субкладам GHIJK. Для сравнения, случаи парагруппы F(xG,H,I,J,K) – то есть либо базального F* (M89), либо первичных субкладов F1 (P91; P104), F2 (M427; M428) и F3 ( M481) – относительно редки во всем мире.
F(xG,H,I,J,K)
[ редактировать ]Отсутствие точных испытаний с высоким разрешением в прошлом затрудняет отдельное обсуждение F*, F1, F2* и F3*. ISOGG заявляет, что F(xG,H,I,J,K) недостаточно изучен, встречается «нечасто» в современных популяциях и достигает максимума в Южной Азии , особенно в Шри-Ланке . [ 9 ] Похоже, что он также уже давно присутствует в Юго-Восточной Азии . Однако вероятность ошибочной идентификации считается относительно высокой, и некоторые случаи могут фактически принадлежать к ошибочно идентифицированным субкладам гаплогруппы GHIJK. Так было, например, с субкладом гаплогруппы H2 (P96), который первоначально назывался «F3», то есть – имя, которое с тех пор было присвоено F-M481.
F(xF1,F2,F3) зарегистрирован у 10% мужчин в Шри-Ланке, у 5,2% мужчин по всей Индии (включая до 10% мужчин в Южной Индии), у 5% в Пакистане, а также у более низких уровней среди мужчин. народ таманг (Непал) и в Иране .
мужчины, происходящие из Индонезии, Сообщалось также, что являются носителями F(xG,H,I,J,K) – особенно F-M89* – в относительно значительных количествах. Сообщается, что уровень заболеваемости составляет 4-5% в Сулавеси и Лембата . Одно исследование, в котором не проводился всесторонний скрининг других субкладов F-M89 (включая некоторые субклады GHIJK), показало, что индонезийские мужчины с SNP P14/PF2704 (который эквивалентен M89) составляют 1,8% мужчин в Западном Тиморе . 1,5% Флореса, 5,4% Лембаты, 2,3% Сулавеси и 0,2% Суматры . [ 12 ] [ 13 ] F1 (P91), F2 (M427) и F3 (M481; ранее F5) очень редки и практически эксклюзивны для регионов/этнических меньшинств в Шри-Ланке, Индии, Непале, Южном Китае , Таиланде , Бирме и Вьетнаме .
В Центральной Азии примеры F(xG,H,I,J,K) были зарегистрированы у лиц из Туркменистана и Узбекистана . [ 14 ]
Кутанан и др. (2020) обнаружили F*-M89 в 50,0% (8/16) образца Красного Лаху , 47,1% (8/17) образца Черного Лаху и 6,7% (1/15) образца Лису в провинции Мае Хонг Сон , Таиланд. все эти говорящие на лолоишском Было обнаружено, что языке члены F*-M89 на северо-западе Таиланда довольно тесно связаны по отцовской линии, причем TMRCA их Y-ДНК оценивается на 584 года раньше настоящего. Однако вышеупомянутые Y-хромосомы лишь отдаленно связаны с экземплярами F*-M89, наблюдаемыми в выборках других популяций Таиланда, в том числе 5,6% (1/18) выборки Фуан из Центрального Таиланда , 11,8% (2/17 ) образца Соа из Северо-Восточного Таиланда и 29% (2/7) образца Саек из Северо-Восточного Таиланда . По оценкам, TMRCA кластера Лолоиш из Северного Таиланда и Y-ДНК особи Фуан из Центрального Таиланда возникли на 12 675 лет раньше настоящего. По оценкам, TMRCA скопления F *-M89 на северо-востоке Таиланда возник 6492 года назад. TMRCA всех этих особей F*-M89 из Таиланда, по оценкам, возник 16 006 лет назад. [ 15 ]
Есть также свидетельства палеолитической обратной миграции F(xG,H,I,J,K) на запад из Южной Азии в Иран , Аравию и Северо-Восточную Африку . [ 16 ] [ 17 ] а также субклады гаплогруппы К в Юго-Восточную Европу . [ 18 ]
Неолитическая миграция в Европу из Юго-Западной Азии первой волны европейских фермеров была выдвинута в качестве источника F и G2a, обнаруженных в европейских неолитических останках, датируемых примерно 4000 годом до нашей эры. Эти останки, по мнению Herrerra et al. (2012) показали «большое генетическое сходство» с «людьми с современного Ближнего Востока», чем с современными европейцами. [ 19 ] F(xG,H,I,J,K), возможно, был обнаружен в останках бронзового века из Европы, а именно в людях, известных как DEB 20 и DEB 38 , которые жили около 7000–7210 лет назад и были найдены в Деренбурге Мееренштиг II. сайт в Германии. [ 20 ]
Три менее надежных случая, которые не были проверены для всех субкладов GHIJK, были обнаружены среди неолитических останков в Европе. I0411 ( Troc 4 ), живший 7195–7080 лет назад, был найден в пещере Эльс-Трокс , недалеко от Бисаурри (современная Испания) – тогда как гаплогруппы G, I1 (I-M450; I-S247) J, L1b2, Q1b1, Q1a2a , R1a1a1 (R-L449), R1b1a2b1a (R-M35) и T были исключены, [ 20 ] I2a1b1 и R1b1a2 обнаружены и в других останках с того же памятника ( Трок 5 и Трок 2 ). Аналогичным образом, три набора останков из Венгрии не были проверены на наличие всех субкладов GHIJK: BAM 17 , BAM 26 (оба из Альсоньека-Баташека , около 7850–7675 лет назад) и TOLM 3 (7030–7230 лет назад, найдены в Тольна-Мезсе ). . [ 20 ] (Человек, известный ученым как « Оазис 1 », живший около 37 800 лет назад в Восточной Европе, первоначально был классифицирован как принадлежащий либо к парагруппе F(xGHIJK), либо к K. группе [ 18 ] Однако последующие исследования показали, что Oase 1 принадлежал K2a* . [ 21 ] )
- встречается выше в Корее , на Филиппинах и в Индонезии.
Некоторые случаи, зарегистрированные среди современных популяций европейцев, коренных американцев и жителей островов Тихого океана, могут быть связаны с миграцией и примесью F(xG,H,I,J,K) в результате контакта с Южной и/или Юго-Восточной Азией во время раннее Новое время (16–19 века). [ 9 ] К таким примерам относятся:
- низкие уровни в Полинезии ; [ 22 ]
- выше среди семинолов и борука ; коренных американцев [ 23 ]
- редкие случаи в Нидерландах ; [ 2 ]
- два случая в Португалии . [ 9 ]
Ф* (М89*)
[ редактировать ]Базальный F-M89* был зарегистрирован у 5,2% мужчин в Индии. [ 10 ] Региональная разбивка была предоставлена Chiaroni et al. (2009 г.): 10% в Южной Индии ; 8% в Центральной Индии ; около 1,0% в Северной Индии и Западной Индии , а также 5% в Пакистане ; 10% в Шри-Ланке ; среди тамангов Непала 4 % ; 2% на Борнео и Яве ; 4-5% в Сулавеси и Лембате в Юго-Восточной Азии . [ 12 ]
В Иране , что 2,3% мужчин бандари из провинции Хормозган являются носителями базального F-M89. было обнаружено [ 16 ]
Гаплогоруп F* был обнаружен только у 11,67% протестированных юньнаньских ханьцев. [ 24 ] Сиань (1/34), [ 24 ]
Гаплогруппа F-M89 также наблюдалась в Северо-Восточной Африке среди двух людей христианского периода , которые были раскопаны на Нила Четвертом пороге и на острове Мероэ . [ 25 ] Останки из пещеры Бачо-Киро доисторического человека F6-620 / AA7-738 из начального верхнего палеолита , датированные периодом от 45 930 до 42 580 калиброванных лет до настоящего времени, несут также базальную линию гаплогруппы Y-хромосомы F-M89. [ 26 ]
Ф1 (Р91)
[ редактировать ]Этот субклад определяется SNP P91. Наиболее распространено в Шри-Ланке . [ 2 ] [ 27 ]
Ф2 (М427)
[ редактировать ]Y-хромосомы F2 были зарегистрированы среди меньшинств из приграничных территорий Южного Китая ( Юньнань и Гуйчжоу ), Таиланда , Бирмы и Вьетнама , а именно И и Куконг или Лаху Ши («Желтый Лаху»), подгруппы Лаху . [ 28 ]
F3 (M481)
[ редактировать ]Недавно определенный и редкий субклад F3 (M481; ранее F5) был обнаружен в Индии и Непале, среди народа тхару и в Андхра-Прадеше . [ 29 ] F-M481 не следует путать с гаплогруппой H2 (L279, L281, L284, L285, L286, M282, P96), которую ранее ошибочно классифицировали как F-M89 как «F3».
Гаплогруппа GHIJK
[ редактировать ]Базальный GHIJK никогда не был обнаружен ни у живых самцов, ни у древних останков.
Субклады, включая некоторые основные гаплогруппы, широко распространены в современном населении Центральной Азии , Кавказа , Ближнего Востока , Южной Азии , Европы , Юго-Восточной Азии , островов Тихого океана и коренных американцев .
Филогенетика
[ редактировать ]В филогенетике Y-хромосомы субклады являются ветвями гаплогрупп. Эти субклады также определяются однонуклеотидными полиморфизмами (SNP) или полиморфизмами уникальных событий (UEP).
Филогенетические деревья
[ редактировать ]Существует несколько подтвержденных и предложенных филогенетических деревьев гаплогруппы F-M89. Научно признанным является вариант Консорциума Y-хромосомы (YCC), опубликованный в Карафете, 2008 г. [ 2 ] и впоследствии обновлялся. Черновик дерева, показывающего новые научные достижения, предоставлен Томасом Краном из Центра геномных исследований в Хьюстоне, штат Техас . Международное общество генетической генеалогии (ISOGG) также предоставляет любительское дерево.
Черновик дерева Центра геномных исследований
[ редактировать ]Черновик дерева гаплогруппы F-M89 Центра геномных исследований выглядит следующим образом. [ 30 ] (Показаны только первые три уровня субкладов.)
- F-M89 P14, M89, M213, P133, P134, P135, P136, P138, P139, P140, P141, P142, P145, P146, P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P316, L132.1, L313, L498
- Ф-П91 П91, П104
- Ф-М427 М427, М428
- Ф-П96 П96, М282, Л279, Л281, Л284, Л285, Л286
- Ф-Л280 Л280
- G-M201 M201, P257, L116, L154, L204, L240, L269, L402, L605, L769, L770, L836, L837, L1258, U2, U3, U6, U7, U12, U17, U20, U21, U23, U33
- H-M69 M69, M370, СТРАНИЦЫ00049
- ИЖК L15, L16
Дерево YCC/ISOGG
[ редактировать ]Это официальное научное дерево, созданное Консорциумом Y-хромосомы (YCC). Последнее крупное обновление было в 2008 году. [ 2 ] Последующие обновления были ежеквартальными и двухгодичными. Текущая версия является версией обновления 2010 года. [ нужна ссылка ]
- CF
- Ф (L132.1, M89/PF2746).
- Ф1 (Р91, Р104)
- Ф2 (М427, М428)
- F3 (M481)
- Макрогаплогруппа GHIJK (F1329/M3658/PF2622/YSC0001299).
- Ф (L132.1, M89/PF2746).
Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. Позже группа гражданских ученых, интересующихся популяционной генетикой и генетической генеалогией, сформировала рабочую группу для создания любительского дерева, стремясь быть прежде всего своевременным. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ф-М89 | 2 | МЫ | 1Р | 20 | ЕС10 | Н4 | Б | Ф* | Ф | Ф | Ф | Ф | Ф | Ф | Ф | Ф | Ф | Ф |
| F-APT/H-APT | 15 | МЫ | 1Р | 20 | ЕС10 | Н4 | Б | Ф1 | Н2 | Н2 | Н2 | Н2 | Н2 | Н2 | Н2 | Н2 | Н2 | Н2 |
См. также
[ редактировать ]Генетика
[ редактировать ]- Генетика и археогенетика Южной Азии
- Генетическая история Ближнего Востока
- Таблица перевода гаплогрупп Y-хромосомы
- Генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- Молекулярная филогенетика
- Парагруппа
- Субклад
- Y-хромосома Аарон
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК у населения Южной Азии
Магистральное дерево
[ редактировать ]Ссылки
[ редактировать ]- ^ Расчетное время отделения F от C = 70 000–75 000 BP; расчетное время, когда G отделится от HIJK = 45 000–50 000 Рагхаван, М.; и др. (2014). «Геном верхнепалеолита Сибири свидетельствует о двойном происхождении коренных американцев» . Природа . 505 (7481): 87–91. Бибкод : 2014Natur.505...87R . дои : 10.1038/nature12736 . ПМК 4105016 . ПМИД 24256729 .
- ^ Jump up to: а б с д и ж Карафет, Татьяна; и др. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Jump up to: а б Хаммер, МФ; Зегура, СЛ (2002). «Дерево гаплогрупп Y-хромосомы человека: номенклатура и филогеография его основных подразделений». Ежегодный обзор антропологии . 31 : 303–321. дои : 10.1146/annurev.anthro.31.040402.085413 .
- ^ Кивисилд и др. 2003, Генетическое наследие первых поселенцев сохраняется как в индийских племенных, так и в кастовых популяциях.
- ^ Сенгупта и др. 2005, Полярность и временность распределения Y-хромосомы с высоким разрешением в Индии Выявляют как местное, так и экзогенное расширение и выявляют незначительное генетическое влияние скотоводов Центральной Азии
- ^ Сангамитра Саху и др. 2006, Предыстория индийских Y-хромосом: оценка сценариев демического распространения
- ^ Арункумар и др. 2012 г.
- ^ Халласт, Таблетки; Грин, Анастасия; Балановский Олег; Сюэ, Он был; Тайлер-Смит, Крис (14 июля 2020 г.). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/ s00439-020-02204-9 ISSN 1432-1203 . ПМЦ 7864842 . ПМИД 32666166 .
- ^ Jump up to: а б с д ISOGG, 2015, Гаплогруппа F Y-ДНК и ее субклады - 2015 (8 сентября 2015 г.).
- ^ Jump up to: а б Сенгупта, Сангамитра; и др. (2006). «Полярность и временной характер распределения Y-хромосомы с высоким разрешением в Индии определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ Халласт, Таблетки; Грин, Анастасия; Балановский Олег; Сюэ, Он был; Тайлер-Смит, Крис (14 июля 2020 г.). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/ s00439-020-02204-9 ISSN 1432-1203 . ПМЦ 7864842 . ПМИД 32666166 .
- ^ Jump up to: а б Кьярони, Жак; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л. (1 декабря 2009 г.). «Разнообразие Y-хромосомы, человеческая экспансия, дрейф и культурная эволюция» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20174–9. Бибкод : 2009PNAS..10620174C . дои : 10.1073/pnas.0910803106 . ПМЦ 2787129 . ПМИД 19920170 .
- ^ Тумонггор, МК; Карафет, ТМ; Дауни, С; Лансинг, Дж. С.; Норквест, П; Судойо, Х; Хаммер, МФ; Кокс, член парламента (2014). «Изоляция, контакты и социальное поведение сформировали генетическое разнообразие в Западном Тиморе» . Журнал генетики человека . 59 (9): 494–503. дои : 10.1038/jhg.2014.62 . ПМЦ 4521296 . ПМИД 25078354 .
- ^ Балареск и др. (2015), Кластеры происхождения Y-хромосомы и мужской дифференциальный репродуктивный успех: молодые линии преобладают в азиатских пасторальных кочевых популяциях , Дополнительная таблица 2
- ^ Вибху Кутанан, Расми Шуконгдеж, Метави Шрикуммул и др. (2020), «Культурные различия влияют на отцовские и материнские генетические линии хмонг-миенских и китайско-тибетских групп из Таиланда». Европейский журнал генетики человека . https://doi.org/10.1038/s41431-020-0693-x
- ^ Jump up to: а б Груни, В; Батталья, В; Хушияр Кашани, Б; и др. (2012). «Древние миграционные события на Ближнем Востоке: новые подсказки по вариации Y-хромосомы современных иранцев» . ПЛОС ОДИН . 7 (7): е41252. Бибкод : 2012PLoSO...741252G . дои : 10.1371/journal.pone.0041252 . ПМЦ 3399854 . ПМИД 22815981 .
- ^ Юсиф, Хишам; Эльтайеб, Мунтасер. «Генетические закономерности вариаций Y-хромосомы и митохондриальной ДНК, имеющие значение для заселения Судана» (PDF) . Хартумский университет . Проверено 17 июля 2016 г.
- ^ Jump up to: а б Цяомэй Фу; Матея Хайдиняк; Оана Теодора Молдован; Сильвиу Константин; Свапан Маллик; Понтус Скоглунд; Ник Паттерсон; Надин Роланд; Иосиф Лазаридис; Биргит Никель; Бенс Виола; Кей Прюфер; Маттиас Мейер; Джанет Келсо; Дэвид Райх; Сванте Пяабо (13 августа 2015 г.). «Ранний современный человек из Румынии с недавним предком-неандертальцем» . Природа . 524 (7564): 216–219. Бибкод : 2015Natur.524..216F . дои : 10.1038/nature14558 . ПМЦ 4537386 . ПМИД 26098372 .
- ^ Эррера, К.Дж.; Лоури, РК; Хадден, Л; и др. (март 2012 г.). «Неолитические отцовские сигналы указывают на то, что Армянское нагорье было заселено земледельцами» . Евро. Дж. Хум. Жене . 20 (3): 313–20. дои : 10.1038/ejhg.2011.192 . ПМК 3286660 . ПМИД 22085901 .
- ^ Jump up to: а б с Жан Манко, 2016 г., ДНК европейского неолита (1 марта 2016 г.).
- ^ Г. Дэвид Позник и др., 2016, «Периодические всплески демографии мужского пола, выведенные на основе 1244 последовательностей Y-хромосомы по всему миру» Nature Genetics , no. 48, стр. 593–599.
- ^ «Меланезийское и азиатское происхождение полинезийцев: градиенты мтДНК и Y-хромосомы через Тихий океан» . Архивировано из оригинала 13 мая 2011 г.
- ^ Филип Эдвард Мелтон, 2008, Генетическая история и доколумбовая диаспора населения, говорящего на языке чибчан: молекулярно-генетические данные ; Анн-Арбор, Мичиган; ПроКвест, с. 29.
- ^ Jump up to: а б Сун-Хи Ким 2011, Высокие частоты линий гаплогруппы Y-хромосомы O2b-SRY465 в Корее: генетический взгляд на заселение Кореи
- ^ Мохамед, Хишам Юсиф Хасан. «Генетические закономерности вариаций Y-хромосомы и митохондриальной ДНК, имеющие значение для заселения Судана» . Хартумский университет. п. 76. Архивировано из оригинала (PDF) 10 ноября 2020 года . Проверено 22 августа 2016 г.
- ^ Хайдиньяк М., Мафессони Ф., Сков Л. и др. « Первоначальные люди верхнего палеолита в Европе имели недавнее неандертальское происхождение ». Nature 592, 253–257 (2021). https://doi.org/10.1038/s41586-021-03335-3
- ^ «PhyloTree y — Минимальное дерево y» .
- ^ Блэк, МЛ; Уайз, Калифорния; Ван, В.; Биттлс, АХ (июнь 2006 г.). «Объединение генетики и истории населения в изучении этнического разнообразия в Китайской Народной Республике» . Биология человека . 78 (3): 277–293. дои : 10.1353/hub.2006.0041 . ПМИД 17216801 . S2CID 42002729 .
- ^ Форнарино, С; Пала, М; Батталья, В; и др. (2009). «Митохондриальное и Y-хромосомное разнообразие Таруса (Непал): резервуар генетических вариаций» . БМК Эвол. Биол . 9 :154. дои : 10.1186/1471-2148-9-154 . ПМК 2720951 . ПМИД 19573232 .
- ^ «Архивная копия» . Архивировано из оригинала 26 мая 2013 г. Проверено 6 января 2013 г.
{{cite web}}: CS1 maint: архивная копия в заголовке ( ссылка )
Внешние ссылки
[ редактировать ]- «ГЕНО 2.0: Великое путешествие, когда-либо рассказанное» . Нэшнл Географик . Архивировано из оригинала 26 июля 2011 г.
- «Дерево гаплогрупп Y-ДНК 2013, версия: 8.72» . Международное общество генетической генеалогии . 25 августа 2013 г.
- Макдональд, доктор юридических наук (2005). «Карты гаплогрупп мира» (PDF) . Архивировано из оригинала (PDF) 28 июля 2004 г. Проверено 6 февраля 2007 г.