Австралопитек седиба
| Австралопитек седиба Временной диапазон: ранний плейстоцен , | |
|---|---|
 | |
| Реконструированный скелет MH1 в Музее естественной истории в Лондоне. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Симииформы |
| Семья: | Гоминиды |
| Подсемейство: | Люди |
| Племя: | Люди |
| Род: | † Австралопитек |
| Разновидность: | † А. ну |
| Биномиальное имя | |
| † Австралопитек седиба | |
Australopithecus sediba — вымерший вид австралопитеков , обнаруженный в пещере Малапа , колыбели человечества , Южная Африка. Он известен по частичному ювенильному скелету, голотипу MH1, и частичному скелету взрослой женщины, паратипу MH2. Они датируются примерно 1,98 миллионами лет назад, в раннем плейстоцене , и сосуществовали с Paranthropusrobustus и Homo ergaster / Homo erectus . Малапа интерпретируется как естественная смертельная ловушка, основание длинной вертикальной шахты, в которую существа могли случайно упасть. Первоначально A. sediba описывался как потенциальный предок человека и, возможно, прародитель Homo , но это оспаривается, и он также мог представлять собой позднее выжившую популяцию или родственный вид A. africanus , ранее населявший эту территорию.
MH1 имеет объем мозга около 350–440 куб. См, как и у других австралопитеков. Лицо MH1 поразительно похоже на Homo , а не на других австралопитеков, с менее выраженными надбровными дугами, скулами и прогнатизмом (насколько лицо выступает вперед), а также есть признаки небольшого подбородка. Однако такие характеристики могут быть обусловлены молодостью и утрачиваться с возрастом. Зубы довольно маленькие для австралопитека. Высота MH1 оценивается в 130 см (4 фута 3 дюйма), что соответствует росту взрослого человека 150–156 см (4 фута 11 дюймов – 5 футов 1 дюйм). По оценкам, MH1 и MH2 имели примерно одинаковый вес - 30–36 кг (66–79 фунтов). Считается, что, как и у других австралопитеков, у A. sediba была узкая верхняя часть груди, напоминающая обезьянью , но широкая и человеческая нижняя часть груди. Как и у других австралопитеков, анатомия рук, по-видимому, предполагает определенную степень лазания и древесного поведения. Таз A. указывает на то, что sediba был способен передвигаться по-человечески, но ступня указывает на своеобразную походку, не продемонстрированную ни у одного другого гоминина, с гиперпронацией лодыжки и, как следствие, с поворотом ноги внутрь при отталкивании. Этот набор адаптаций может представлять собой компромисс между привычными прямохождение и древесность.
A. sediba , по-видимому, питался только лесными растениями C 3 , такими как некоторые травы и осоки , плоды, листья и кора. Это сильно контрастирует с другими ранними гомининами , которые питались смесью растений саванны C 3 и C 4 , но похоже на современных шимпанзе саванны . Ни у одного другого гоминина нет доказательств того, что он ел кору. Такая универсальная диета, возможно, позволила ему занять меньшую территорию обитания, чем шимпанзе саванны. В районе Малапы, возможно, было прохладнее и влажнее, чем сегодня, и здесь были закрытые леса, окруженные более открытыми лугами.
История исследований
[ редактировать ]Образцы
[ редактировать ]





Первой ископаемой находкой была правая ключица MH1 (UW88-1) в пещере Малапа , колыбели человечества , Южная Африка, обнаруженная 9-летним Мэтью Бергером 15 августа 2008 года во время исследования места раскопок, возглавляемого его отцом Саутом. Африканский палеоантрополог Ли Роджерс Бергер . Дальнейшие раскопки позволили получить частичный скелет MH1, дополнительно включающий частичный череп и фрагменты челюстной кости, а также части рук, пальцев, плеч грудной клетки , , позвоночника , таза , ног и ступней. MH1 интерпретируется как молодой самец из-за явно выраженного развития надбровных дуг и клыков корней , выворота угла нижней челюсти и крупных рубцов на костях. [1] Однако антропологи Уильям Кимбел и Йоэль Рак утверждают, что это ненадежные методы определения пола, и предполагают, что MH1 является женским на основании отсутствия передних столбиков (столбиков, спускающихся вдоль носового отверстия до вокруг рта) и слегка выпуклой подносовой кости. пластинка с использованием методов определения пола A. africanus . [2] MH1 получил прозвище «Карабо», что на языке тсвана означает «ответ» , 17-летней Омфеметсе Кипил из школы Святой Марии в Йоханнесбурге на конкурсе имен. Она выбрала это имя, потому что «это ископаемое представляет собой решение для понимания происхождения человечества». [3]
Другой частичный скелет, взрослый MH2, был обнаружен Ли 4 сентября 2008 года с изолированными верхними зубами, частичной челюстной костью, почти полной правой рукой, правой лопаткой и фрагментами плеч, правой руки, позвоночника, ребер, таза и коленный сустав и стопы. Лобковая кость широкая и квадратная, а мышечные рубцы на теле от слабых до умеренных, что позволяет предположить, что MH2 принадлежит женщине. [1]
Присутствие видов, которые развились после 2,36 миллиона лет назад и вымерли около 1,5 миллиона лет назад, указывает на то, что слой A. sediba датируется где-то внутри этого интервала в раннем плейстоцене . Датирование уран-свинцовым , камнем перекрывающим этот слой, дало дату 2,026 ± 0,021 миллиона лет назад. Согласно археомагнитному датированию , отложения имеют нормальную магнитную полярность (в отличие от обратной магнитной полярности в наши дни), и единственный раз, когда это произошло в течение этого интервала, - между 1,95 и 1,78 миллионами лет назад. [4] В 2011 году возраст камня был более точно датирован 1,977 ± 0,002 миллиона лет назад с использованием уран-свинцового датирования. [5]
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
Тафономия
[ редактировать ]Сети пещер вокруг Малапы представляют собой длинные взаимосвязанные пещерные отверстия площадью 500 × 100 м (1640 × 330 футов). Местонахождение в Малапе могло находиться у подножия системы пещер глубиной не более 30 метров (98 футов). Пещера находится на пересечении разлома, заполненного кремнем на северо-северо-востоке и северо-северо-западе , а останки гомининов были обнаружены на участке размером 3,3 × 4,4 × 3,5 м (11 футов × 14 футов × 11 футов) на севере. - северо-западный перелом. Слой был открыт в результате добычи известняка в начале 20 века. Пещера состоит из пяти осадочных фаций A–E водного песчаника , при этом A. sediba обнаружен из фации D, а большее количество остатков гомининов принадлежит фации E. MH1 и MH2 разделены по вертикали не более чем на 40 см (16 дюймов). Фация D представляет собой светлоокрашенный слой толщиной 1,5 метра (4,9 фута), перекрывающий текучий камень. Мелкие пелоиды обычны, но они слиты в большие и неправильные группы, что указывает на то, что они отлагались в заболоченных условиях. Пелоиды могут представлять собой фекалии или почвенные микробы. Состояние сохранности MH1 и MH2 указывает на то, что они были сданы на хранение быстро, перемещались очень мало и были цементируется вскоре после отложения во фреатической среде (в подземном потоке). Нет никаких свидетельств сбора мусора, что указывает на то, что эта территория была недоступна для хищников. [4]
Все это может указывать на то, что пещера Малапа была смертельной ловушкой с незаметными пещерными отверстиями на поверхности. Животные могли быть соблазнены запахом воды, исходящей из шахты, а хищники - запахом мертвых животных, а затем падали насмерть. Из-за сильного селевого потока останки оказались занесены глубже в пещеру вдоль подземного потока, возможно, из-за сильного ливня. В конце концов камера обрушилась и заполнилась грязью. [4]
Классификация
[ редактировать ]В 2010 году Ли и его коллеги официально описали вид Australopithecus sediba с MH1 в качестве голотипа и MH2 в качестве паратипа . Название вида « седиба » на местном языке сесото означает «фонтан» или «родник» . [1] Поскольку A. sediba имел много общих черт с Homo ergaster / H. erectus , особенно в области таза и ног, описатели предположили, что A. sediba был переходным ископаемым между австралопитеками и Homo . [1] Зубные особенности также позволяют предположить некоторое близкое родство между A. sediba и предком Homo . [6] Однако экземпляры были обнаружены в стратиграфическом подразделении , датируемом 1,95–1,78 миллиона лет назад, тогда как самые ранние окаменелости Homo того времени датировались 2,33 миллиона лет назад ( H. habilis из Хадара , Эфиопия). [1] В настоящее время самым старым экземпляром Homo является LD 350-1, датируемый 2,8–2,75 миллиона лет назад из Леди-Герару , Эфиопия. [7] Чтобы устранить несоответствие в датировке, авторы описания также выдвинули гипотезу, что A. sediba произошел от популяции A. africanus (населявшей тот же регион) незадолго до появления гоминидов Малапы, и что Homo отделился от A. sediba некоторое время после этого. [1] Это предполагает 800-тысячелетнюю призрачную родословную между A. africanus и гоминидами Малапы. [2] Было также высказано предположение, что A. sediba , а не H. habilis или H. rudolfensis , был прямым предком H. ergaster / H. erectus (самого раннего неоспоримого представителя рода Homo ), прежде всего потому, что гоминиды Малапы были датированы 1,98 миллиона лет назад в 2011 году, который на тот момент предшествовал самому раннему представителю H. ergaster / H. erectus . [5] Сейчас считается, что A. sediba был современником H. ergaster / H. erectus и Paranthropusrobustus в колыбели человечества. [8]
В качестве альтернативы, A. sediba также может представлять собой поздно выжившую морфу или сестринский вид , A. africanus не связанный с Homo , что означало бы, что гомо -подобные черты развились независимо у A. sediba и Homo ( гомоплазия ). [2] [9] [10] [11] [12] Летопись окаменелостей раннего человека плохо изучена и основана в основном на фрагментарных останках, что делает убедительные анатомические сравнения трудными, а иногда и невозможными. [12] A. africanus , A. afarensis и A. garhi также считались истинными предками Homo , и этот вопрос вызывает много споров. [7] Кроме того, голотип представляет собой подростка, на что ссылаются Кимбел и Рак, утверждая, что некоторые черты лица, подобные гомо, могли быть утрачены с возрастом. [2]
Современная классификация австралопитеков находится в беспорядке. Australopithecus можно считать таксоном , члены которого объединены схожей физиологией, а не тесными отношениями друг с другом по сравнению с другими родами гоминидов, и по большей части неясно, как тот или иной вид связан с другими. [13]
 |
Анатомия
[ редактировать ]Череп
[ редактировать ]

Сохранился только свод черепа МН1 объемом 363 куб.см. По оценкам, объем самой задней части мозга составлял 7–10 см3. Для оценки мозжечка были использованы австралопитеки KNM-ER 23000 ( Paranthropus boisei ) и Sts 19 ( A. africanus ) объёмами 40–50 куб.см, а также KNM-ER 1813 ( H. habilis ), KNM-ER 1805 ( H . habilis ) и KNM-ER 1470 ( H. rudolfensis ) объёмами 55–75 куб.см были использованы для оценки объёма мозжечка MH1 примерно в 50 куб.см. Учитывая все это, MH1 мог иметь объем мозга около 420–440 куб.см. Это характерно для австралопитеков. [1] Используя тенденции, наблюдаемые у современных приматов между размером мозга взрослого и новорожденного, размер мозга новорожденного мог составлять 153–201 куб. См, что аналогично тому, что предположительно наблюдается у других австралопитеков. [14] Конфигурация мозга, по-видимому, в основном напоминала австралопитеков, но орбитофронтальная кора, по-видимому, больше напоминала человеческую. [15]
В целом анатомия черепа A. sediba наиболее похожа на A. africanus . Однако MH1 имеет меньший череп, более широкий свод черепа в поперечном направлении, более вертикально наклоненные стенки теменной кости и более широко расположенные височные линии . Как и у Homo , надбровные дуги менее выражены, скулы менее расширены, лицо не так сильно выступает вперед (меньший прогнатизм ), имеется небольшой подбородок. [1] Однако такие характеристики также обнаружены в некоторых A. africanus черепах из Стеркфонтейна, член 4, что, по мнению Кимбела и Рака, может указывать на то, что эти гомоподобные признаки были утрачены с возрастом. Кроме того, если прогнатизм измеряется с использованием передней носовой ости, а не самого основания носа, прогнатизм при MH1 попадает в диапазон, наблюдаемый у A. africanus . [2] Зубы довольно малы для австралопитека и больше соответствуют зубам раннего человека . Однако, в отличие от Homo , коренные зубы постепенно увеличиваются в размерах по направлению к задней части рта (в отличие от второго моляра, который является самым большим), а бугорки расположены ближе друг к другу. [1]
Форма ветви нижней челюсти (стержня, соединяющего челюсть с черепом) сильно различается у MH1 и MH2. У MH1 выше и шире; передняя и задняя границы почти вертикальны и параллельны, в отличие от непараллельных границ MH2 с вогнутой передней границей; и венечный отросток MH1 наклонен назад с глубокой и асимметричной нижнечелюстной вырезкой , тогда как MH2 имеет неизогнутый венечный отросток с неглубокой нижнечелюстной вырезкой. По сравнению с закономерностями, наблюдаемыми у современных человекообразных обезьян , такие заметные различия превышают то, что можно было бы ожидать, если бы их можно было объяснить половым диморфизмом или ювенильным статусом MH1. Скелет A. sediba мог быть весьма изменчивым видом. [16]
Торс
[ редактировать ]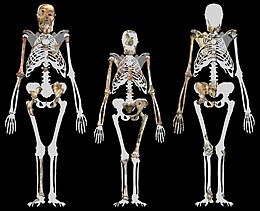
Высота MH1 составляет 130 см (4 фута 3 дюйма). [17]
По оценкам, MH1 и MH2 имели примерно одинаковый размер, около 30–36 кг (66–79 фунтов). Это меньше, чем у многих современных гомининов, но вполне приемлемо для австралопитека. [18] MH1 был ростом около 130 см (4 фута 3 дюйма), но он был подростком и имел примерно такое же развитие скелета, как у 12-летнего человеческого ребенка или 9-летнего шимпанзе. A. sediba , как и более ранние и современные гоминины, по-видимому, имела обезьяноподобную скорость роста, основанную на скорости развития зубов, поэтому MH1 мог достичь примерно 85% своего взрослого размера, если предположить траекторию роста, подобную шимпанзе, или 80%, если предположить, что человекоподобная траектория. Это будет примерно равняться 150 или 156 см (4 фута 11 дюймов или 5 футов 1 дюйм). [17]
MH1 сохраняет 4 шейных , 6 грудных и 2 поясничных позвонка; а MH2 сохраняет 2 шейных, 7 грудных, 2 поясничных и 1 крестцовый позвонки. [19] Лордоз (человеческое искривление) и суставы шейных позвонков, указывающие на положение головы, подобное человеческому. Однако общая анатомия шейных позвонков напоминает обезьянью и указывает на гораздо более жесткую шею. У A. sediba , подобное человеческому отсутствует плечевое сплетение (которое имеется у некоторых A. afarensis ), а плечевое сплетение человека отвечает за иннервацию нервов и мышц рук и кистей, улучшая двигательный контроль. [20] Как и люди, A. sediba , по-видимому, имел гибкий поясничный отдел, состоящий из 5 позвонков - в отличие от 6 статических позвонков у человекообразных обезьян - и демонстрирующий поясничный лордоз (искривление позвоночника человека), соответствующий привычному вертикальному положению. Однако у A. sediba , по-видимому, была очень подвижная нижняя часть спины и выраженный поясничный лордоз. [19] который, возможно, участвовал в противодействии крутящим моментам, направленным внутрь, при ходьбе гиперпронационной походкой, предложенной для A. sediba . [21] MH1 сохраняет 2 верхних грудных, 1 среднегрудное и 3 нижних грудных ребра; и MH2 — 4 последовательных верхне-среднегрудных и 3 нижних грудных ребра, соединенных с позвонками. [19] Это указывает на то, что у A. sediba была суженная верхняя часть груди, напоминающая обезьянью, но анатомия таза, подобная человеческой, может свидетельствовать о том, что у A. sediba была широкая и человеческая нижняя часть груди. Узкая верхняя часть груди препятствовала бы размахиванию руками во время ходьбы, ограничивала бы грудную клетку и препятствовала бы тяжелому дыханию и, следовательно, быстрой ходьбе или бегу на длинные дистанции. Напротив, у A. sediba , по-видимому, была узкая талия, похожая на человеческую, измененное положение наружных косых мышц живота и более широкие подвздошно-реберные мышцы на спине, что повышало эффективность ходьбы, противодействуя боковому сгибанию туловища. [22]

Таз имеет несколько общих черт с ранними Homo и H. ergaster , а также с KNM-ER 3228 из Кооби Фора , Кения, и OH 28 из Олдувайского ущелья , Танзания, которые не отнесены к виду (хотя обычно классифицируются как Homo spp. ) Было больше поддержки вдоль вертлужной впадины и крестца , улучшающее разгибание бедра, расширение места прикрепления подвздошно-бедренной связки , смещающее вес позади центра вращения бедра, больше поддержки вдоль вертлужной впадины и лопатки подвздошной кости , улучшающее попеременный наклон таза , и большее расстояние между вертлужная впадина и седалищный бугр, уменьшающие момент плеча сухожилий подколенных . Возможно, это позволило A. sediba сделать шаг, похожий на человеческий . Тазобедренный сустав, по-видимому, имел более человеческий характер несущей нагрузки, чем образец H. habilis OH 62. [1] A. Родовые пути кажутся sediba более гинекоидными (нормальное состояние человека), чем у других австралопитеков, которые более платипеллоидны, хотя A. sediba не является полностью гинекоидным, что может быть связано с меньшим размером мозга (и, следовательно, головы) новорожденного. . Как и у людей, родовые пути имели увеличенный диаметр в сагиттальном направлении (спереди назад), а лобковая кость загибалась вверх. [14]
Верхние конечности
[ редактировать ]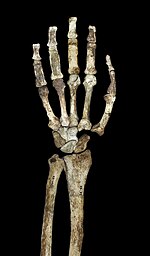
Подобно другим австралопитекам и ранним Homo , A. sediba имел несколько обезьяноподобные пропорции верхней части тела с относительно длинными руками, высоким плечевым индексом (соотношение предплечья к плечевой кости ) 84 и большими суставными поверхностями. Спорный вопрос, является ли обезьяноподобная конфигурация верхних конечностей австралопитеков показателем древесного поведения или просто является базальной чертой, унаследованной от последнего общего предка человекообразных обезьян в отсутствие серьезного селективного давления, направленного на принятие более человеческой анатомии рук. Плечи расправлены, лопатка имеет хорошо развитый подмышечный край , хорошо выражен коноидальный бугорок (важный для прикрепления мышц вокруг плечевого сустава). [1] Мышечные рубцы на ключицах указывают на человеческий диапазон движений. Лопатка наиболее сходна с таковой орангутанов по размерам суставной впадины (образующей плечевой сустав) и ее углу с позвоночником, хотя по форме лопатка наиболее сходна с людьми и шимпанзе . Плечевая кость имеет низкую степень скручивания в отличие от людей и африканских обезьян, что (наряду с короткой ключицей) предполагает, что лопатка располагалась дальше от средней линии, как у Homo , хотя она располагалась выше спины, как у других австралопитеков. [23] Обезьяноподобные качества рук, по-видимому, более выражены у A. sediba , чем у более древнего A. afarensis , и если A. afarensis является предком A. sediba , это может указывать на адаптивный сдвиг в сторону древесного поведения. [24]
В локтевом суставе латеральные и медиальные надмыщелки плечевой кости удлинены, как и у других австралопитеков и африканских обезьян, не являющихся человеком. Плечевая кость также имеет развитый гребень в локтевом суставе для поддержки плечелучевой мышцы, сгибающей предплечье. Как и у африканских обезьян, не являющихся людьми, у них имеется сильная привязанность к бицепсам лучевой кости и трицепсам локтевой кости . меньше Однако механическое преимущество бицепсов и плечевых мышц . [23] Локтевая кость также обеспечивает прочное прикрепление локтевого сгибателя запястья . Олекранонная ямка большая и глубокая, с выступающим блоковым килем , который важен для поддержания устойчивости рук при их выпрямлении. Кости пальцев длинные, прочные и изогнутые, поддерживают сильные мышцы-сгибатели поверхностных пальцев, важные для сгибания пальцев. [1] Иногда это считают свидетельством древесного поведения австралопитеков. Рука также имеет относительно длинный большой палец и короткие пальцы, как у человека , что может свидетельствовать о точном хвате, важном для создания и использования сложных каменных орудий . [25]
Нижние конечности
[ редактировать ]
Как и у других австралопитеков, голеностопные, коленные и тазобедренные суставы указывают на привычное прямохождение . Кости ног очень похожи на кости A. afarensis . Лодыжка в основном похожа на человеческую, возможно, с ахилловым сухожилием , похожим на человеческое . [26]
Таранная кость толстая и больше похожа на таковую у человекообразных обезьян, имеет изогнутую медиально шею и низкий угол скручивания шеи. Спорный вопрос, ли A. sediba имел свод стопы, похожий на человеческий , или ступня была больше похожа на обезьянью. [27] Пяточная кость изогнута под углом 45 градусов и заметно наклонена спереди назад, наиболее сильно в малоберцовом блоке. Крепкий блок малоберцовой кости указывает на сильные малоберцовые мышцы , которые простираются от икры до лодыжки. На стопе отсутствует латеральный подошвенный бугорок (который может участвовать в рассеивании сил, когда пятка касается земли при нормальной походке человека), наблюдаемый у людей и A. afarensis . [1] [26] Изящное тело пяточной кости и крепкая лодыжка (костные выступы с каждой стороны лодыжки) весьма напоминают обезьяну, с менее эффективной передачей силы между пяточной костью и таранной костью и обезьяньей подвижностью в средней части стопы. Состояние A. sediba наиболее похоже на состояние, наблюдаемое у горилл , а стопа, возможно, функционально эквивалентна стопе A. africanus . [26] [28]
Палеобиология
[ редактировать ]Диета
[ редактировать ]Анализ фитолитов (микроскопических остатков растений) из зубного налета обоих образцов и анализ изотопов углерода показывают, что в их рационе почти исключительно присутствуют лесные растения C 3, несмотря на предположительно широкую доступность растений C 4 в их смешанной среде саванны. Такая модель питания также наблюдается у современных шимпанзе саванны и, как предполагается, относится к раннеплиоценовому Ardipithecus ramidus , но она сильно отличается от любого другого раннего гоминина. Всего из двух зубов MH1 было обнаружено 38 фитолитов, из которых 15 соответствуют двудольным , 9 однодольным и остальные 14 неопределенным. Однодольные растения, вероятно, произошли от трав C 3 и осоки, растущих в хорошо поливаемых и тенистых местах, а другие фитолиты были получены из фруктов, листьев, древесины или коры. Хотя кору обычно едят другие приматы из-за высокого содержания в ней белка и сахара, неизвестно, что ни один другой гоминин не употреблял кору регулярно. Анализ микроизноса зубов также показывает, что два гоминина Малапы ели твердую пищу, значения сложности варьируются от H. erectus крепкий P.robustus и [29] Тем не менее, челюсти, по-видимому, не были так хорошо приспособлены для производства высоких штаммов по сравнению с другими ранними гомининами, что может указывать на то, что A. sediba не так сильно зависела от своей способности перерабатывать механически сложную пищу. [30] [31]
Интерпретация A. sediba как универсального травоядного животного лесных растений C 3 согласуется с тем, что он, по крайней мере, частично является древесным. Такая широкая диета, возможно, позволила A. sediba занять гораздо меньшие ареалы обитания, чем современные шимпанзе саванны, которые преимущественно потребляют только фрукты, поскольку A. sediba смогла вернуться к коре и другим устойчивым к разрушению продуктам. [29]
походка
[ редактировать ]
Во время ходьбы у A. sediba могла наблюдаться гиперпронация голеностопного сустава, вызывающая чрезмерный перенос веса внутрь во время фазы опоры. У современных людей-гиперпронаторов стопа сильно перевернута во время фазы переноса, и контакт с землей сначала осуществляется внешним краем стопы, вызывая высокие крутящие моменты, вращающие всю ногу внутрь. Аналогичным образом, крепления прямой мышцы бедра и двуглавой мышцы бедра у A. sediba соответствуют нагрузкам, направленным по средней линии ног, бедер и коленей. Этот способ ходьбы неидеален для современной анатомии человека, а гиперпронаторы подвергаются более высокому риску развития подошвенного фасциита , растяжения голени и большеберцовой кости стрессовых переломов . Чтобы противодействовать этому, A. sediba , возможно, использовал подвижную среднюю часть стопы вместо жесткой, как у человека, что могло предотвратить чрезмерную стрессовую нагрузку на лодыжку. [21]
Гиперпронационная походка и связанный с ней набор адаптаций не были выявлены у других гомининов, и неясно, почему у A. sediba это развилось. [21] Подвижная средняя часть стопы также будет полезна при интенсивном лазании. [1] [21] [26] таким образом, гиперпронация могла быть компромиссом между привычным прямохождением и древесностью. [21]
Рождение
[ редактировать ]
По оценкам, входное отверстие таза самки A. sediba составляло 80,8 мм × 112,4 мм (3,18 дюйма × 4,43 дюйма) в длину и ширину (сагиттальный x поперечный), а поскольку размер головы новорожденного оценивается в 89,2 мм (3,51 дюйма). в) в лучшем случае новорожденный, вероятно, вошел в входное отверстие таза, ориентированное в поперечном направлении, как и другие гоминиды. Средняя плоскость входа в таз сужена минимум до 96,9 мм (3,81 дюйма), поэтому новорожденному, возможно, не потребуется поворачивать во время рождения. Размеры входа в таз рассчитывали с использованием комплексной реконструкции, включающей седалищную кость юных мужчин ; Точно так же родовой канал, возможно, на самом деле был больше, чем рассчитывалось. По оценкам, плечи имели ширину 74,3 мм (2,93 дюйма), поэтому они не препятствовали родам больше, чем голова. Таким образом, новорожденный в точке наибольшего сужения занимал бы около 92,1% родового канала, обеспечивая достаточно места для совершенно неротационного рождения, как это наблюдается у человекообразных обезьян и, возможно, у других австралопитеков (хотя и у полу-человекообразных обезьян). также предлагаются ротационные роды). Хотя можно пройти и без вращения, средняя плоскость расширяется в переднезаднем направлении (спереди назад), и для новорожденного было бы больше места, если бы он вращался так, чтобы самая длинная часть головы совпадала с этим расширением. [32]
У современных людей, по сравнению с этим, роды гораздо более трудоемкие и сложные, требующие полного вращения новорожденного, поскольку большой мозг и, следовательно, размер головы, а также жесткие плечи новорожденного человека значительно затрудняют его прохождение через роды. родовые пути. Используя оценку размера мозга новорожденного A. sediba в 145,8–180,4 куб.см , размер головы новорожденного должен был составлять 73 мм × 89 мм (2,9 дюйма × 3,5 дюйма), как у новорожденного шимпанзе. [32]
Разработка
[ редактировать ]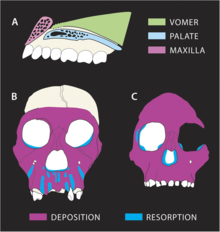
Траектория роста MH1, по-видимому, заметно отличалась от других гомининов. Носочелюстной комплекс (кость от носа до верхней губы) указывает на большую степень резорбции кости, наиболее выраженную у корней передних зубов. Это контрастирует с A. africanus и A. afarensis , которые являются депозитарными, что отражает усиление прогнатизма с возрастом. Для P.robustus также характерна резорбция верхней челюсти, но резорбция при MH1 распространяется вдоль передних зубов до клыковой ямки вблизи скуловых костей, в результате чего лицо становится мезогнатическим (несколько выдающимся) в отличие от плоского лица у P.robustus . Поскольку резорбция происходит так близко к скулам, это может объяснить, почему MH1 не демонстрирует расширяющихся скул, характерных для A. africanus . Прорезывание зубов, вероятно, не повлияло на ремоделирование нижней части лица, поскольку у MH1 уже были все постоянные зубы. Тем не менее, меньший размер щечных зубов мог позволить создать мезогнатическое лицо. Диета A. sediba , по-видимому, заметно отличалась от типичной диеты ранних гоминидов, возможно, она была похожа на диету современных гоминидов. оливковая обезьяна-колобус, питающаяся преимущественно молодыми листьями; эти два вида, по-видимому, имеют схожие модели роста лицевых костей. Это может указывать на расходящиеся модели резорбции и отложения у A. sediba , отражающие различные модели нагрузки на челюсти от других гомининов. Края глазниц MH1 изогнуты, тогда как у A. africanus они изрезаны , что может указывать на отложение кости у A. sediba в регионах, где у A. africanus происходит резорбция кости . [33]
Патология
[ редактировать ]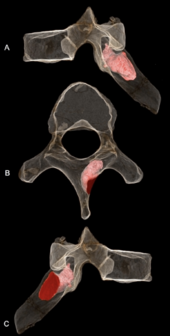
Правая пластинка шестого грудного позвонка MH1 представляет собой проникающую костную опухоль , вероятно, доброкачественную остеоид-остеому . Поражение проникает на глубину 6,7 мм (0,26 дюйма) и ширину 5,9 мм (0,23 дюйма) и на момент смерти все еще было активным. Он не проник в нервный канал , поэтому, вероятно, не вызвал каких-либо неврологических осложнений, и нет никаких признаков сколиоза (аномального искривления позвоночника). Это могло повлиять на движение лопатки и верхнего правого квадранта спины, вызывая острую или хроническую боль, мышечные нарушения или мышечные спазмы . Учитывая, что A. sediba может обладать необходимой способностью лазать, расположение очага поражения вблизи места прикрепления трапециевидных мышц , мышц, выпрямляющих позвоночник , и больших ромбовидных мышц может ограничивать нормальные модели движений. MH1 имеет самый ранний диагностированный случай рака SK 7923 возрастом 1,8–1,6 миллиона лет, у гоминина, по крайней мере, на 200 000 лет, предшествующий фрагменту плюсневой кости представляющему собой остеосаркому из Сварткранса , колыбели человечества. Опухоли встречаются редко в летописи окаменелостей гомининов, вероятно, из-за низкой заболеваемости приматов в целом; Ранние гоминины, вероятно, имели такие же показатели заболеваемости, как и современные приматы. У ювенильного MH1, у которого развивается опухоль кости, согласуется с общей тенденцией опухолей костей, возникающих преимущественно у молодых людей. [34]
У MH1 и MH2 наблюдаются присмертные (приблизительно во время смерти) повреждения костей, соответствующие травме тупым предметом . Это согласуется с интерпретацией этого места как основания высокой шахты, действующей как естественная смертельная ловушка, в которую случайно попали животные. MH1 и MH2, возможно, упали с высоты примерно 5–10 м (16–33 футов) на наклонную кучу гравия, песка и помета летучих мышей , что, вероятно, в некоторой степени смягчило падение. При MH1 присмертные переломы наиболее выражены на челюстных костях и зубах, хотя возможно, что эти травмы возникли не только в результате падения, но и в результате удара падающим предметом. MH2 имеет признаки использования корсета во время травмы, с нагрузкой на предплечье и кисть и ударом в грудь, на правой стороне тела обнаружены присмертные переломы. Это первые случаи смерти в летописи окаменелостей австралопитеков, которые уверенно не связаны с хищничеством или естественными причинами. [35]
Палеоэкология
[ редактировать ]
В 2010 году вместе с гомининами в фациях D и E было обнаружено в общей сложности 209 окаменелостей, не принадлежащих человеческому роду, и таксоны из них были идентифицированы : саблезубая кошка Dinofelis barlowi , леопард , африканская дикая кошка , черноногая кошка , бурая гиена , плащовая лисица , мангусты Atilax mesotes и Mungos , генета , африканская дикая собака , лошадь , свинья , клипспрингер , антилопа Megalotragus , большая антилопа алселафин , родственник запряженного бушбока , родственник большого куду и зайца . [4] [36] Сегодня черноногий кот и капская лисица являются эндемиками южноафриканских лугов, кустарников и кустарников . Точно так же коричневая гиена обитает в сухих открытых местах обитания и никогда не встречалась в закрытых лесах. Динофелис и Атилакс , напротив, обычно являются индикаторами закрытой, влажной среды обитания. Это может указывать на то, что на этой территории есть закрытая среда обитания, а также луга — судя по ареалу обитания капской лисицы, оба они существовали в пределах 20 км. 2 (7,7 квадратных миль) участка. [36]
хищника В копролите фации D обнаружены пыльца и фитолиты деревьев Podocarpus или Afrocarpus , а также обломки древесины неидентифицированных хвойных и двудольных деревьев. Фитолиты трав не обнаружены. В наши дни территория Малапы представляет собой луга, а Подокарпус и Афрокарпус встречаются в 30 км (19 миль) от Афромонтанского леса биома в каньонах на высоте 1500–1900 м (4900–6200 футов) над уровнем моря в Магалисберг горном хребте . , где лесные пожары встречаются реже. Это может указывать на то, что Малапа была более прохладной и влажной территорией, чем сегодня, что позволяло достаточно уменьшить пожары, чтобы позволить таким лесным растениям распространиться так далеко за пределы естественно защищенных территорий. Малапа в раннем плейстоцене, возможно, также находилась на несколько более низкой высоте, чем сегодня, при этом долины и Магалисберг были менее выражены. [37]
Австралопитеки и ранние Homo, вероятно, предпочитали более прохладные условия, чем более поздние Homo , поскольку на момент отложения не было мест обитания австралопитеков на высоте ниже 1000 м (3300 футов) над уровнем моря. Это означало бы, что, как и шимпанзе, они часто населяли районы со средней дневной температурой 25 °C (77 °F), а ночью опускались до 10 или 5 °C (50 или 41 °F). [38] Пещера Малапа в настоящее время находится на высоте 1442 м (4731 фут) над уровнем моря. [4] A. sediba жил рядом с P.robustus и H.ergaster / H. erectus . Поскольку A. africanus вымер примерно в это же время, вполне возможно, что Южная Африка была убежищем для австралопитеков примерно до 2 миллионов лет назад, когда началась значительная климатическая изменчивость и непостоянство, а также потенциально конкуренция с Homo и Paranthropus . [8]
См. также
[ редактировать ]- Африканская археология
- Australopithecus africanus - вымерший гоминид из Южной Африки.
- Homo ergaster - вымершие виды или подвиды архаичного человека.
- Homo gautengensis - название, предложенное для вымершего вида гомининов из Южной Африки.
- Homo habilis - архаичный человеческий вид от 2,8 до 1,65 млн лет назад.
- Homo star - южноафриканский архаичный вид человека.
- Paranthropus boisei - вымерший вид гоминидов Восточной Африки.
- Paranthropusrobustus - вымерший вид гоминидов Южной Африки.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Бергер, ЛР; де Рюитер, диджей; Черчилль, SE; Шмид, П.; Карлсон, К.Дж.; Диркс, PHGM; Кибии, Дж. М. (2010). « Австралопитек седиба : новый вид гомоподобных австралопитеков из Южной Африки». Наука . 328 (5975): 195–204. Бибкод : 2010Sci...328..195B . CiteSeerX 10.1.1.729.7802 . дои : 10.1126/science.1184944 . ПМИД 20378811 . S2CID 14209370 .
- ^ Перейти обратно: а б с д и Кимбел, В.; Рак, Ю. (2017). « Австралопитек седиба и появление человека : сомнительные доказательства черепа ювенильного голотипа MH 1». Журнал эволюции человека . 107 : 94–106. Бибкод : 2017JHumE.107...94K . дои : 10.1016/j.jhevol.2017.03.011 . ПМИД 28526292 .
- ^ Кинг, Дж. (4 июня 2010 г.). « Окаменелость австралопитека седиба, названная 17-летним студентом из Йоханнесбурга» . Центр происхождения. Архивировано из оригинала 25 марта 2012 года . Проверено 9 июля 2011 г.
- ^ Перейти обратно: а б с д и Диркс, PHGM; Кибии, Дж. М.; Кун, Б.Ф.; Штайнингер, К.; Черчилль, SE; Крамерс, доктор медицинских наук; Пикеринг, Р.; Фарбер, Д.Л.; и др. (2010). «Геологическое положение и возраст австралопитека седиба из Южной Африки» (PDF) . Наука . 328 (5975): 205–208. Бибкод : 2010Sci...328..205D . дои : 10.1126/science.1184950 . ПМИД 20378812 . S2CID 206524717 .
- ^ Перейти обратно: а б Пикеринг, Р.; Диркс, PHGM; Джинна, З.; и др. (2011). « Австралопитек седиба, возраст 1,977 млн лет назад, и последствия для происхождения рода Homo » . Наука . 333 (6048): 1421–1423. Бибкод : 2011Sci...333.1421P . дои : 10.1126/science.1203697 . ПМИД 21903808 . S2CID 22633702 .
- ^ Ирландец, доктор юридических наук; Гаутелли-Стейнберг, Д.; Легге, СС; и др. (2013). «Морфология зубов и филогенетическое «место» австралопитека седиба ». Наука . 340 (6129): 1233062. doi : 10.1126/science.1233062 . ПМИД 23580535 . S2CID 206546794 .
- ^ Перейти обратно: а б Вильмоаре, Б.; Кимбел, Вашингтон; Сейюм, К.; и др. (2015). «Ранний человек возрастом 2,8 млн лет назад из Леди-Герару, Афар, Эфиопия» . Наука . 347 (6228): 1352–1355. Бибкод : 2015Sci...347.1352V . дои : 10.1126/science.aaa1343 . ПМИД 25739410 .
- ^ Перейти обратно: а б Херрис, ВОЗДУХ; Мартин, Дж. М.; и др. (2020). «Современность австралопитеков , парантропов и раннего человека прямоходящего в Южной Африке». Наука . 368 (6486): eaaw7293. дои : 10.1126/science.aaw7293 . hdl : 11568/1040368 . ПМИД 32241925 . S2CID 214763272 .
- ^ Балтер, Майкл (2010). «Кандидат в предки человека из Южной Африки вызывает похвалы и споры» (PDF) . Наука . 328 (5975): 154–155. дои : 10.1126/science.328.5975.154 . ПМИД 20378782 .
- ^ Черри, М. (8 апреля 2010 г.). «Претензия на «предка человека» вызывает фурор» . Природа . Новости природы. дои : 10.1038/news.2010.171 .
- ^ Ду, А.; Алемсегед, З. (2019). «Временные данные показывают, что австралопитек седиба вряд ли был предком человека» . Наука . 5 (5): е9038. Бибкод : 2019SciA....5.9038D . дои : 10.1126/sciadv.aav9038 . ПМК 6506247 . ПМИД 31086821 .
- ^ Перейти обратно: а б Спур, Фред (5 октября 2011 г.). «Палеоантропология: Малапа и род Homo » . Природа . дои : 10.1038/478044a .
- ^ МакНалти, КП (2016). «Таксономия и филогения гомининов: что в имени?» . Знания о природном образовании . 7 (1): 2.
- ^ Перейти обратно: а б Кибии, Дж. М.; Черчилль, SE; Шмид, П.; и др. (2011). «Частичный таз австралопитека седиба » . Наука . 333 (6048): 1407–1411. Бибкод : 2011Sci...333.1407K . дои : 10.1126/science.1202521 . ПМИД 21903805 . S2CID 206532267 .
- ^ Карлсон, К.Дж.; Стаут, Д.; Джашашвили Т.; и др. (2011). «Эндокаст MH1, австралопитек седиба ». Наука . 333 (6048): 1402–1407. Бибкод : 2011Sci...333.1402C . дои : 10.1126/science.1203922 . ПМИД 21903804 . S2CID 206533255 .
- ^ Ритцман, ТБ; Терхьюн, CE; Гунц, П.; Робинсон, Калифорния (2018). «Форма ветви нижней челюсти Australopithecus sediba предполагает наличие одного изменчивого вида». Журнал эволюции человека . 100 : 54–64. дои : 10.1016/j.jhevol.2016.09.002 . ПМИД 27765149 .
- ^ Перейти обратно: а б Кэмерон, Н.; Богин, Б.; Болтер, Д.; Бергер, ЛР (2018). «Посткраниальное созревание скелета австралопитека седиба » . Американский журнал физической антропологии . 163 (3): 633–640. дои : 10.1002/ajpa.23234 . ПМИД 28464269 . S2CID 3287309 .
- ^ Холлидей, ТВ; Черчилль, SE; и др. (2018). «Размер тела и пропорции австралопитека седиба » (PDF) . Палеоантропология : 406–422. doi : 10.4207/PA.2018.ART118 (неактивен 31 января 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - ^ Перейти обратно: а б с Уильямс, Южная Каролина; Островский, КР; и др. (2013). «Позвоночник австралопитека седиба ». Наука . 340 (6129): 1232996. doi : 10.1126/science.1232996 . ПМИД 23580532 . S2CID 206546736 .
- ^ Мейер, MR; Уильямс, Южная Каролина; Шмид, П.; Черчилль, SE; Бергер, ЛР (2017). «Шейный отдел австралопитека седиба ». Журнал эволюции человека . 104 : 32–49. Бибкод : 2017JHumE.104...32M . дои : 10.1016/j.jhevol.2017.01.001 . ПМИД 28317555 .
- ^ Перейти обратно: а б с д и ДеСильва, Дж. М.; Холт, КГ; Черчилль, SE; и др. (2013). «Нижняя конечность и механика ходьбы австралопитека седиба ». Наука . 340 (6149): 1232999. doi : 10.1126/science.1232999 . ПМИД 23580534 . S2CID 13288792 .
- ^ Шмид, П.; Черчилль, SE; Налла, С. (2013). «Мозаичная морфология грудной клетки австралопитека седиба ». Наука . 340 (6129): 1234598. doi : 10.1126/science.1234598 . ПМИД 23580537 . S2CID 31073328 .
- ^ Перейти обратно: а б Черчилль, SE; Холлидей, ТВ; Карлсон, К.Дж.; и др. (2013). «Верхняя конечность австралопитека седиба ». Наука . 340 (6129): 1233477. doi : 10.1126/science.1233477 . ПМИД 23580536 . S2CID 206547001 .
- ^ Рейн, ТР; Харрисон, Т.; Карлсон, К.Дж.; Харвати, К. (2016). «Адаптация к подвесному передвижению у австралопитека седиба » . Журнал эволюции человека . 104 : 1–12. дои : 10.1016/j.jhevol.2016.12.005 . ПМИД 28317552 .
- ^ Кивелл Т.Л., Кибии Дж.М., Черчилль С.Е., Шмид П., Бергер Л.Р. (2011). « Рука австралопитека седиба демонстрирует мозаичную эволюцию локомоторных и манипулятивных способностей». Наука . 333 (6048): 1411–1417. Бибкод : 2011Sci...333.1411K . дои : 10.1126/science.1202625 . ПМИД 21903806 . S2CID 11610235 .
- ^ Перейти обратно: а б с д Зипфель Б., ДеСильва Дж.М., Кидд Р.С., Кэрисон К.Дж., Черчилль С.Е., Бергер Л.Р. (2011). «Стопа и лодыжка австралопитека седиба » . Наука . 333 (6048): 1417–1420. Бибкод : 2011Sci...333.1417Z . дои : 10.1126/science.1202703 . ПМИД 21903807 . S2CID 206532338 .
- ^ Пранг, TC (2015). «Поза задней части стопы австралопитека седиба и эволюция продольного свода гоминина» . Научные отчеты . 5 : 17677. Бибкод : 2015NatSR...517677P . дои : 10.1038/srep17677 . ПМЦ 4667273 . ПМИД 26628197 .
- ^ Пранг, TC (2016). «Подтаранный суставной комплекс австралопитека седиба » . Журнал эволюции человека . 90 : 105–119. Бибкод : 2016JHumE..90..105P . дои : 10.1016/j.jhevol.2015.10.009 . ПМИД 26767963 .
- ^ Перейти обратно: а б Генри, Аманда Г.; Унгар, Питер С.; Пасси, Бенджамин Х.; Спонхаймер, Мэтт; Россоу, Ллойд; Бэмфорд, Мэрион; Сандберг, Пол; де Рюитер, Дэррил Дж.; Бергер, Ли (2012). «Питание австралопитека седиба ». Природа . 487 (7405): 90–93. Бибкод : 2012Natur.487...90H . дои : 10.1038/nature11185 . ПМИД 22763449 . S2CID 205229276 .
- ^ Ледогар, Дж.А.; Смит, Алабама; Бенацци, С.; и др. (2016). «Механические доказательства того, что австралопитек седиба был ограничен в способности есть твердую пищу» . Природные коммуникации . 7 (10596): 10596. Бибкод : 2016NatCo...710596L . дои : 10.1038/ncomms10596 . ПМЦ 4748115 . ПМИД 26853550 .
- ^ Дэглинг, диджей; Карлсон, К.Дж.; Таффоро, П.; де Рюитер, диджей; Бергер, ЛР (2016). «Сравнительная биомеханика нижних челюстей австралопитека седиба». Журнал эволюции человека . 100 : 73–86. Бибкод : 2016JHumE.100...73D . дои : 10.1016/j.jhevol.2016.08.006 . ПМИД 27765151 .
- ^ Перейти обратно: а б Лаудичина, Нью-Мексико; Родригес, Ф.; ДеСильва, Дж. М. (2019). «Реконструкция рождения австралопитека седиба » . ПЛОС ОДИН . 14 (9): e0221871. Бибкод : 2019PLoSO..1421871L . дои : 10.1371/journal.pone.0221871 . ПМК 6750590 . ПМИД 31532788 .
- ^ Лакруз, РС; Бромедж, Т.Г.; О'Хиггинс, П.; и др. (2015). «Заметный рост носочелюстного комплекса у Au. sediba » . Научные отчеты . 5 (15175): 15175. Бибкод : 2015NatSR...515175L . дои : 10.1038/srep15175 . ПМК 4606807 . ПМИД 26469387 .
- ^ Рэндольф-Куинни, PS; Уильямс, ЮАР; Стейн, М.; и др. (2016). «Остеогенная опухоль у австралопитека седиба : самые ранние свидетельства неопластического заболевания у гомининов» . Южноафриканский научный журнал . 112 (7–8): 7. doi : 10.17159/sajs.2016/20150470 .
- ^ Л'Аббе, EN; Саймс, ЮАР; Покинс, Джей Ти; Кабо, Луизиана; и др. (2015). «Доказательства смертельных травм скелета у малапских гомининов 1 и 2» . Научные отчеты . 5 (15120): 15120. Бибкод : 2015NatSR...515120L . дои : 10.1038/srep15120 . ПМК 4602312 . ПМИД 26459912 .
- ^ Перейти обратно: а б Кун, Б.Ф.; Верделин, Л.; Хартстон-Роуз, А.; Лакруз, РС; Бергер, ЛР (2011). «Останки карниворана со стоянки гоминидов в Малапе, Южная Африка» . ПЛОС ОДИН . 6 (11): e26940. Бибкод : 2011PLoSO...626940K . дои : 10.1371/journal.pone.0026940 . ПМК 3207828 . ПМИД 22073222 .
- ^ Бэмфорд, М.; и др. (2010). «Ботанические останки копролита из плейстоценовой стоянки гоминидов в Малапе, долина Стеркфонтейн, Южная Африка». Палеонтол. Афр . 45 : 23–28.
- ^ Дэвид-Барретт, Т.; Данбар, RIM (2016). «Еще раз о двуногости и выпадении волос в эволюции человека: влияние высоты и планирования активности» . Журнал эволюции человека . 94 : 72–82. Бибкод : 2016JHumE..94...72D . дои : 10.1016/j.jhevol.2016.02.006 . ПМЦ 4874949 . ПМИД 27178459 .
Дальнейшее чтение
[ редактировать ]- Уильямс, Южная Каролина; Мейер, MR; Налла, С.; и др. (2018). «Позвонки, ребра и грудина австралопитека седиба » . Палеоантропология : 156–233. doi : 10.4207/PA.2018.ART113 (неактивен 31 января 2024 г.).
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на январь 2024 г. ( ссылка ) - де Рюитер, диджей; Черчилль, SE; Бергер, LL (2013). Рид, Кентукки ; Флигл, Дж. Г.; Лики, Р.Э. (ред.). Австралопитек седиба из Малапы, Южная Африка . Палеобиология и палеоантропология позвоночных. Спрингер Нидерланды. стр. 147–160. дои : 10.1007/978-94-007-5919-0_9 . ISBN 978-94-007-5919-0 .
{{cite book}}:|work=игнорируется ( помогите )
Внешние ссылки
[ редактировать ]- Реконструкции Джона Гурча
- Скелеты представляют собой изысканную палеоголоволомку по науке
- Что такое австралопитек седиба ? Джон Д. Хоукс
- Весь каталог сайта гомининов Малапы 2013, часть 1
- Весь каталог сайта гомининов Малапы 2013, часть 2
- Человеческая временная шкала (интерактивная) – Смитсоновский институт

| Базы данных органов управления : Национальные |
|---|