Соло-мужчина
| Соло-мужчина Временной диапазон: поздний плейстоцен | |
|---|---|
 | |
| Отливка «Черепа XI» в Зале происхождения человека , Вашингтон, округ Колумбия. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Симииформы |
| Семья: | Гоминиды |
| Подсемейство: | Люди |
| Племя: | Люди |
| Род: | гомо |
| Разновидность: | |
| Подвиды: | † Его. солончак |
| Трехчленное имя | |
| † Человек прямоходящий солонский Оппеноорт, 1932 год. | |
| Синонимы [ нужна ссылка ] | |
| |
Соло-человек ( Homo erectus soloensis ) — подвид , H. erectus обитавший вдоль реки Соло на острове Ява , Индонезия, примерно 117 000–108 000 лет назад в позднем плейстоцене . Эта популяция является последней известной находкой этого вида. Он известен по 14 черепам, двум голеням и части таза, раскопанным недалеко от деревни Нгандонг, а также, возможно, трем черепам из Самбунгмакана и черепу из Нгави в зависимости от классификации. Впервые раскопки на территории Нгандонга проводились с 1931 по 1933 год под руководством Виллема Фредерика Флоруса Оппенорта, Карела тер Хаара и Густава Генриха Ральфа фон Кенигсвальда , но дальнейшее исследование было прервано Великой депрессией , Второй мировой войной и индонезийской войной за независимость. . В соответствии с историческими расовыми концепциями , индонезийский подвид H. erectus первоначально классифицировался как прямой предок австралийских аборигенов , но сейчас считается, что у Человека-Соло нет живых потомков, поскольку останки появились намного раньше современной иммиграции людей в этот район, которая началась примерно 55 000 лет назад. до 50 000 лет назад.
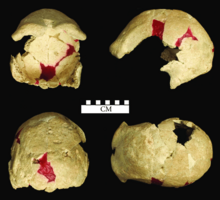
Череп Человека-Соло, вид сверху, имеет овальную форму, с густыми бровями, надутыми скулами и заметной полоской костей, охватывающей спину. Объем мозга был довольно большим: от 1013 до 1251 кубических сантиметров (от 61,8 до 76,3 кубических дюймов) по сравнению со средним размером 1270 см3. 3 (78 куб. дюймов) для современных современных мужчин и 1130 см. 3 (69 у.е. дюймов) для современных современных женщин. Один потенциально женский экземпляр мог иметь рост 158 см (5 футов 2 дюйма) и весить 51 кг (112 фунтов); самцы, вероятно, были намного крупнее самок. Соло-человек во многом был похож на яванского человека ( H.e. erectus ), ранее населявшего Яву, но был гораздо менее архаичным.
Человек-одиночка, вероятно, населял открытую лесную местность, гораздо более прохладную, чем современная Ява, наряду со слонами , тиграми , диким рогатым скотом , водяными буйволами , тапирами и бегемотами , а также другими представителями мегафауны. Они изготавливали простые отщепы и чопперы (ручные каменные орудия ), а возможно, копья или гарпуны из костей, кинжалы из жал скатов, а также болы или молотки из андезита . Возможно, они произошли от человека с Явы или, по крайней мере, были тесно связаны с ним. Образцы Нгандонга, вероятно, погибли во время извержения вулкана. Этот вид, вероятно, вымер с захватом тропических лесов и утратой предпочтительной среды обитания, начиная с 125 000 лет назад. Черепа получили повреждения, но неясно, явилось ли это результатом нападения, каннибализма , извержения вулкана или процесса окаменения.
История исследований
[ редактировать ]Несмотря на то, что английский натуралист Чарльз Дарвин выдвинул гипотезу в своей книге «Происхождение человека» 1871 года , многие натуралисты-эволюционисты конца XIX века постулировали, что Азия, а не Африка, была местом рождения человечества, поскольку она находится на полпути между Европой и Америкой, обеспечивая оптимальные пути расселения по всему миру. мир ( теория выхода из Азии ). Среди них был немецкий натуралист Эрнст Геккель , который утверждал, что первый человеческий вид (который он назвал « Homo primigenius ») развился на ныне опровергнутом гипотетическом континенте « Лемурия » на территории нынешней Юго-Восточной Азии из рода , который он назвал « питекантроп» ( Pitecanthropus ). «человек-обезьяна»). «Лемурия» предположительно затонула под Индийским океаном , поэтому не удалось найти никаких окаменелостей, подтверждающих это. Тем не менее, модель Геккеля вдохновила голландского учёного Эжена Дюбуа присоединиться к Королевской нидерландской Ост-Индской армии (KNIL) и заняться поисками своего « недостающего звена » на Индонезийском архипелаге . На Яве он нашел тюбетейку и бедренную кость ( Яванский человек ), датируемые поздним плиоценом или ранним плейстоценом . Участок Тринил вдоль реки Соло , который он назвал « P. » erectus (используя гипотетическое название рода Геккеля) в 1893 году. Он безуспешно пытался убедить европейское научное сообщество в том, что он нашел прямоходящего человека-обезьяну. Они в основном отвергли его выводы, назвав его уродливой нечеловеческой обезьяной. [1] : 588
Тем не менее «обезьяночеловек Явы» вызвал академический интерес, и, чтобы найти новые останки, Прусская академия наук в Берлине поручила немецкому зоологу Эмилю Селенке продолжить раскопки Тринила. После его смерти в 1907 году раскопки проводила его жена и коллега-зоолог Маргарет Ленор Селенка . Среди членов был голландский геолог Виллем Фредерик Флорус Оппенорт. Годовая экспедиция оказалась безрезультатной, но Геологическая служба Явы продолжала спонсировать раскопки вдоль реки Соло. Примерно два десятилетия спустя Исследование профинансировало несколько экспедиций по обновлению карт острова. Оппеноорт был назначен главой Java-картографической программы в 1930 году. Одной из их задач было четкое различение третичных и четвертичных отложений, среди соответствующих участков пласт, датируемый плейстоценом, открытый голландским геологом Карелом тер Хааром в 1931 году ниже по течению от Тринила. участок, недалеко от деревни Нгандонг. [2]

были обнаружены 12 фрагментов черепа (включая хорошо сохранившиеся тюбетейки), а также две правые большеберцовые кости С 1931 по 1933 год под руководством Оппенорта, Тер Хаара и немецко-голландского геолога Густава (берцовые кости), одна из которых была практически целой. Генрих Ральф фон Кенигсвальд . [3] : 2–3 В середине раскопок Оппеноорт ушел из Исследования и вернулся в Нидерланды, его заменил польский геолог Юзеф Звежицкий в 1933 году. В то же время из-за Великой депрессии фокус Исследования сместился на экономически значимую геологию, а именно нефтяные месторождения. , и раскопки Нгандонга полностью прекратились. В 1934 году тер Хаар опубликовал важные обзоры операций в Нгандонге до того, как заразился туберкулезом . Он вернулся в Нидерланды и умер два года спустя. Фон Кенигсвальд, который был нанят главным образом для изучения яванских млекопитающих, был уволен в 1934 году. После активного лоббирования со стороны Звежицкого в «Обзоре» и после получения финансирования от Института науки Карнеги фон Кенигсвальд восстановил свою должность в 1937 году, но был слишком занят на объекте Сангиран для продолжения исследований в Нгандонге. [3] : 23–26
В 1935 году останки Человека-Соло были перевезены в Батавию (сегодня Джакарта , Ява, Индонезия) на попечение профессора местного университета Виллема Альфонса Мейсберга в надежде, что он возьмет на себя изучение образцов. Прежде чем у него появилась такая возможность, окаменелости были перевезены в Бандунг , Западная Ява, в 1942 году из-за японской оккупации Голландской Ост-Индии . [3] : 23 Японские войска интернировали фон Кенигсвальда на 32 месяца. По окончании войны его освободили, но разразилась индонезийская война за независимость . Еврейско-немецкий антрополог Франц Вайденрайх (который бежал из Китая перед японским вторжением в 1941 году) договорился с Фондом Рокфеллера и Фондом викингов о доставке фон Кенигсвальда, его жены Луитгарде и яванских человеческих останков (включая Человека-Соло) в Нью-Йорк. Фон Кенигсвальд и Вайденрайх изучали материал в Американском музее естественной истории. [4] до смерти Вайденрайха в 1948 году (оставив после себя монографию о Человеке-одиночке, посмертно опубликованную в 1951 году). [5] В свою книгу « Встреча с доисторическими людьми » 1956 года фон Кенигсвальд включил 14-страничный отчет о проекте Нгандонг с несколькими неопубликованными результатами. Останки Соло-человека были отправлены на хранение в Утрехтский университет , Нидерланды. В 1967 году фон Кенигсвальд передал материал Теуку Якобу для его докторского исследования. Джейкоб руководил раскопками Нгандонга с 1976 по 1978 год и обнаружил еще два образца черепа и фрагмент таза . В 1978 году фон Кенигсвальд вернул материал в Индонезию, а останки Соло-человека были перевезены в Университет Гаджа Мада , особый регион Джокьякарта (юго-центральная Ява). [3] : 26












Образцы: [5] : 217
- Череп I, почти целая тюбетейка, вероятно, принадлежавшая пожилой женщине;
- Череп II, лобная кость , вероятно, принадлежавшая ребенку от трех до семи лет;
- Череп III, деформированная тюбетейка, вероятно, принадлежавшая пожилому человеку;
- Череп IV, тюбетейка, вероятно, принадлежавшая женщине средних лет;
- Череп V, вероятная мужская тюбетейка, на что указывает его большая длина - 221 мм (8,7 дюйма);
- Череп VI, почти полная тюбетейка, вероятно, принадлежащая взрослой женщине;
- Череп VII, фрагмент правой теменной кости, вероятно, принадлежавший молодому человеку, возможно, женщине;
- Череп VIII, обе теменные кости (отдельные), возможно, принадлежат молодому мужчине;
- Череп IX, тюбетейка без основания, вероятно, принадлежит пожилому человеку (небольшой размер соответствует женщине, но тяжесть соответствует мужчине);
- Череп X — разбитая тюбетейка, вероятно, принадлежавшая крепкой пожилой женщине;
- Череп XI, почти полная тюбетейка;
- Большеберцовая кость А, несколько фрагментов стержня диаметром 101 мм (4,0 дюйма) в середине стержня, вероятно, принадлежащие взрослому мужчине;
- Большеберцовая кость B, почти целая правая большеберцовая кость длиной 365 мм (14,4 дюйма) и диаметром посередине 86 мм (3,4 дюйма), вероятно, принадлежащая взрослой женщине;
- Яйцо 15, частичная тюбетейка; [3] : 3
- Нгандонг 16, левый теменный фрагмент; [3] : 3 и
- размером 4 × 6 см (1,6 × 2,4 дюйма) Нгандонг 17, левая вертлужная впадина (на тазу, который является частью тазобедренного сустава ). [3] : 3
Возраст и тафономия
[ редактировать ]Местоположение этих окаменелостей на террасе Соло на момент открытия было плохо задокументировано. Оппеноорт, тер Хаар и фон Кенигсвальд находились на объекте только 24 дня из 27 месяцев работы, поскольку им нужно было наблюдать за другими объектами третичного периода для исследования. Они оставили своих помощников-геологов — Самси и Пануджу — наблюдать за раскопками; их записи теперь утеряны. Карта сайта Обзора оставалась неопубликованной до 2010 года (более 75 лет спустя) и сейчас используется ограниченно, поэтому тафономия и геологический возраст Человека-Соло были спорными вопросами. [3] : 5 Сообщается, что все 14 образцов были обнаружены в верхней части слоя II (из шести слоев), который представляет собой пласт толщиной 46 см (18 дюймов) с гравийным песком и вулканокластическим гиперстеновым андезитом . Предполагается, что они были отложены примерно в то же время, вероятно, в ныне высохшем рукаве реки Соло, примерно в 20 м (66 футов) над современной рекой. Участок находится на высоте около 40 м (130 футов) над уровнем моря . [3] : 15–18

Вулканокластическая порода указывает на то, что отложение произошло вскоре после извержения вулкана . Из-за огромного объема окаменелостей люди и животные могли сконцентрироваться в больших количествах в долине вверх по течению от этого места из-за извержения или сильной засухи. Пепел отравил бы растительность или, по крайней мере, препятствовал бы ее росту, что привело бы к голоду и смерти среди травоядных и людей, накапливая массу туш, разлагающихся в течение нескольких месяцев. Отсутствие повреждений, нанесенных плотоядными животными, может указывать на то, что достаточное питание было возможно без необходимости пережевывать кости. Когда дождей наступил сезон , лахары, текущие с вулкана по речным руслам, унесли трупы на участок Нгандонг, где они и другие обломки образовали затор из-за сужения русла там. [6] [7] Окаменелости H. erectus из Самбунгмакана, также расположенного вдоль реки Соло, возможно, были отложены в то же время. [3]
Попытки знакомств:
- В 1932 году, основываясь на высоте этого места над современной рекой, Оппеноорт предположил, что Соло Человек датируется эемским межледниковьем , которое в то время было примерно ограничено 150–100 тысячами лет назад, начиная с перехода среднего и позднего плейстоцена . [2] Более поздние биохронологические исследования (с использованием останков животных для определения возраста) в течение следующих нескольких лет, проведенные Опппеноортом в 1932 году, фон Кенигсвальдом в 1934 году и тер Хааром в 1936 году, подтвердили дату позднего плейстоцена. [8]
- Останки Соло-Человека были впервые радиометрически датированы в 1988 году, а затем в 1989 году с использованием уран-ториевого датирования , от 200 до 30 тысяч лет назад, что представляет собой широкий диапазон ошибок. [9]
- В 1996 году зубы Человека-Соло были датированы с помощью датирования электронного спинового резонанса урана и (ЭПР) и масс-спектрометрии изотопного соотношения тория 53,3–27 тысячами лет назад; это означало бы, что человек-одиночка пережил континентальный H. erectus как минимум на 250 000 лет и был современником современных людей в Юго-Восточной Азии. [8] которые иммигрировали примерно 55–50 тысяч лет назад. [10]
- В 2008 году гамма-спектроскопия трех черепов показала, что они подверглись выщелачиванию урана, а останки Человека-Соло были датированы примерно 70–40 тысячами лет назад. Это по-прежнему делает возможным, что Соло-Человек был современником современных людей. [9]
- В 2011 году аргон-аргоновое датирование пемзы роговой обманки дало максимальный возраст 546 ± 12 тысяч лет назад, а ЭПР и уран-ториевое датирование кости млекопитающего ниже по течению на стоянке Джигар I - минимальный возраст от 143 до 77 тысяч лет назад. . Этот продолжительный интервал позволил бы предположить, что Одиночный Человек был современником континентального H. erectus , задолго до того, как современные люди рассеялись по континенту. [11]
- В 2020 году была опубликована первая полная хронология стоянки Нгандонг, согласно которой река Соло прошла через это место 500 000 лет назад; терраса Соло отложилась более 316–31 тыс. лет назад; терраса Нгандонг 141–92 тыс. лет назад; и костный пласт H. erectus 117–108 тысяч лет назад. Это означало бы, что Одиночный Человек действительно является последней известной популяцией H. erectus и не взаимодействовал с современными людьми. [7]
Классификация
[ редактировать ]
Расовая классификация аборигенов Австралии из-за прочности черепа по сравнению с черепом других современных популяций исторически была сложным вопросом для европейской науки с тех пор, как Иоганн Фридрих Блюменбах (основатель физической антропологии ) представил эту тему в 1795 году в своей работе. его De Generis Humani Varietate Nativa («О естественной истории человечества»). Следуя концепции эволюции Дарвина, английский антрополог Томас Генри Хаксли в 1863 году предположил наличие отношений между предками и потомками между европейскими неандертальцами и австралийскими аборигенами, что было развито более поздними расовыми антропологами до открытия индонезийских архаичных людей. [12]
В 1932 году Оппеноорт предварительно провел параллели между черепом человека-одиночки и черепом родезийского человека из Африки, неандертальцев и современных австралийских аборигенов. [2] В то время считалось, что люди произошли из Центральной Азии, что отстаивали в первую очередь американский палеонтолог Генри Фэйрфилд Осборн и его протеже Уильям Диллер Мэтью . Они считали Азию «матерью континентов», а поднятие Гималаев и Тибета и последующее высыхание региона вынудили предков человека стать наземными и двуногими . Они утверждали, что популяции, отступившие в тропики, а именно яванский человек Дюбуа и « негроидная раса », существенно регрессировали ( теория вырождения ). Они также отвергли Рэймонда Дарта южноафриканского ребенка Таунга ( Australopithecus africanus ) как предка человека, отдав предпочтение мистификации Пилтдаунского человека из Британии. [1] : 599–602 Сначала Оппеноорт полагал, что материал Нгандонга представляет собой азиатский тип неандертальца, который был более тесно связан с родезийским человеком (также считавшимся неандертальским типом), и дал ему родовое обозначение как « Javanthropus Soloensis ». Дюбуа считал Человека-Соло более или менее идентичным Человеку -Ваджаку из Восточной Явы (теперь классифицированному как современный человек), поэтому Оппеноорт впоследствии начал использовать имя « Homo (Javanthropus) Soloensis ». [5] Оппеноорт выдвинул гипотезу, что Яванский человек эволюционировал в Индонезии и был предшественником современных австралийских аборигенов, а Человек-одиночка был переходным ископаемым . Он считал родезийского человека членом этой же группы. Что касается китайского пекинского человека (ныне H. e. pekinensis ), то он считал, что он распространился на запад и дал начало неандертальцам. [2]
Таким образом, древний яванский человек, одиночный человек и родезийский человек обычно группировались в линию «питекантропоид- австралоид ». «Австралоиды» включают австралийских аборигенов и меланезийцев . Это было расширением мультирегионального происхождения современных людей, отстаиваемых Вайденрайхом и американским расовым антропологом Карлтоном С. Куном , которые считали, что все современные расы и этнические группы (которые до середины 20-го века были классифицированы в отдельные подвиды или даже виды) развивались независимо. от местного архаичного человеческого вида ( полигенизм ). Австралийские аборигены считались самой примитивной расой из ныне живущих. [12] В 1950-х годах немецкий биолог-эволюционист Эрнст Майр вошел в область палеоантропологии и, исследуя «ошеломляющее разнообразие названий», решил определить только три вида Homo : « H. transvaalensis » ( австралопитеки ), H. erectus (включая Человек-одиночка и несколько предполагаемых африканских и азиатских таксонов) и Homo sapiens (включая всех, кто моложе H. erectus , например, современных людей и неандертальцев). Майр определил их как последовательную линию, в которой каждый вид развивается в следующий ( хроновид ). Хотя позже Майр изменил свое мнение об австралопитеках (признав австралопитеков ), и с тех пор несколько видов были названы или получили некоторое признание, его более консервативный взгляд на архаичное человеческое разнообразие получил широкое распространение в последующие десятилетия. [13] Хотя Майр не стал подробно останавливаться на подвиде H. erectus , последующие авторы начали формально включать в него виды со всех частей Старого Света. Соло-человек был помещен Вайденрайхом в «группу неандертальцев/неандертальцев/неандертальоидов» в 1940-х годах, которую он зарезервировал для экземпляров, очевидно, переходных между H. erectus и H. sapiens . Эту группу также можно отнести к ныне несуществующему роду Palaeoanthropus . Одиночный человек был впервые классифицирован как подвид H. erectus американским физическим антропологом Карлтоном Куном в его книге 1962 года «Происхождение рас» . [14]
Утверждение о том, что аборигены Австралии произошли от азиатского H. erectus, получило дальнейшее развитие в 1960-х и 1970-х годах, когда некоторые из самых старых известных (современных) окаменелостей человека были обнаружены в Австралии, в первую очередь под руководством австралийского антрополога Алана Торна . Он отметил, что некоторые популяции были заметно более устойчивыми, чем другие, поэтому он предположил, что Австралия была колонизирована двумя волнами («дигибридная модель»): первая волна была очень устойчивой и происходила от близлежащего H. erectus , а вторая волна была более изящной ( менее крепкие) и происходят от анатомически современных аборигенов Восточной Азии (которые, в свою очередь, произошли от китайского H. erectus ). Впоследствии было обнаружено, что некоторые из наиболее крепких экземпляров моложе изящных. В 1980-х годах, когда африканские виды, такие как A. africanus, стали широко признаваться предками человека, а раса стала менее значимой в антропологии, теория «Из Африки» опровергла модели «За пределами Азии» и мультирегиональные модели. В результате многорегиональная модель была переработана: локальные популяции архаичных людей скрещивались и внесли, по крайней мере, некоторый предок в современные популяции в своих соответствующих регионах, что также известно как модель ассимиляции. Человек-одиночка вписывается в это, гибридизовавшись с вполне современными предками австралийских аборигенов, путешествующих на юг через Юго-Восточную Азию. Модель ассимиляции не получила повсеместной поддержки. В 2006 году австралийский палеоантрополог Стив Уэбб вместо этого предположил, что Человек-одиночка был первым человеческим видом, достигшим Австралии, а более устойчивые современные австралийские экземпляры представляют собой гибридные популяции. [12] : 3
Дата существования Человека-Одинока — 117–108 тысяч лет назад, предшествовавшая расселению современного человека по Юго-Восточной Азии (и, в конечном итоге, по Австралии), противоречит этому выводу. Столь древняя дата не оставляет Соло-Мэна живых потомков. [7] Аналогичным образом, геномное исследование 2021 года, в котором изучались геномы более 400 современных людей (из которых 200 прибыли с островов Юго-Восточной Азии ), не обнаружило никаких доказательств какой-либо «суперархаичной» (т. е. H. erectus ) интрогрессии. [15] Обычно считается, что Соло-человек произошел от яванского человека ( H.e. erectus , типичного для популяций Сангиран/Тринил), а три черепа из Самбунгмакана и череп из Нгави были отнесены к H.e. soloensis или какой-то промежуточной стадии между H. e. erectus и H. e. солоенсис . Во многом неясно, был ли поток генов с континента. Альтернативная гипотеза, впервые предложенная Джейкобом в 1973 году, заключается в том, что популяции Сангиран/Тринил и Нгандонг/Нгави/Самбунгмакан были сестринскими группами , которые развивались параллельно друг другу. [16] Если альтернатива верна, это могло бы служить основанием для выделения вида как « H. soloensis », но определения видов и подвидов, особенно в палеоантропологии , составлены плохо. [17]
Анатомия
[ редактировать ]
Идентификация взрослого или подростка была основана на закрытии черепных швов , предполагая, что они закрывались со скоростью, аналогичной скорости современного человека (хотя у H. erectus они могли закрываться в более раннем возрасте ). Что характерно для H. erectus , у человека-одиночки череп чрезвычайно толстый, от двух до трех раз больше, чем у современных людей. Особи мужского и женского пола различались, исходя из предположения, что самцы более крепкие, чем самки, хотя и самцы, и самки исключительно крепкие по сравнению с другими азиатскими H. erectus . Черепа взрослых в среднем имеют размеры 202 × 152 мм (8,0 × 6,0 дюйма) в длину, умноженную на ширину, и пропорционально аналогичны черепам пекинского человека, но имеют гораздо большую окружность. Череп V — самый длинный — 221 мм (8,7 дюйма). [5] : 236–239 Для сравнения, размеры современных человеческих черепов в среднем составляют 176 × 145 мм (6,9 × 5,7 дюйма) для мужчин и 171 × 140 мм (6,7 × 5,5 дюйма) для женщин. [18]
Останки Соло-человека характеризуются более производными чертами, чем более архаичный яванский H. erectus , в первую очередь большим размером мозга, приподнятым сводом черепа , уменьшенным заглазничным сужением и менее развитыми надбровными дугами. Они все еще очень напоминают более раннего H. erectus . [16] небольшой сагиттальный киль Как и у пекинского человека, по средней линии черепа проходил . По сравнению с другими азиатскими H. erectus лоб пропорционально низкий, а также имеет небольшой угол наклона. Надбровные дуги не образуют сплошную полосу, как у пекинского человека, а изгибаются вниз в средней точке, образуя переносицу. Брови довольно густые, особенно на боковых концах (ближний край лица). Как и у пекинского человека, лобные пазухи расположены между глазами, а не доходят до области бровей. По сравнению с неандертальцами и современными людьми область, височная мышца, которую покрывала довольно плоская. Надбровные дуги переходят в заметно утолщенные скулы. Череп фенозиготный: шляпка черепа пропорционально узкая по сравнению со скулами, так что последние все еще видны, если смотреть на череп сверху вниз. Плоская часть височной кости треугольная, как у пекинского человека, подвисочный гребень довольно острый. [5] : 240–242 Как и у более раннего Javan H. erectus , нижняя и верхняя височные линии (на теменной кости) расходятся к задней части черепа. [19] : 143
В задней части черепа имеется острый и толстый затылочный тор (выступающая костная перекладина), который отмечает четкое разделение между затылочной и затылочной плоскостями . Затылочный тор больше всего выступает в той части, которая соответствует наружному затылочному бугорку у современного человека. Основание височной кости больше соответствует яванскому и пекинскому человеку, чем неандертальцам и современным людям. В отличие от неандертальцев и современных людей, у корня крыловидной кости имеется четко выраженная костная пирамидальная структура . Сосцевидная часть височной кости у основания черепа заметно выступает вперед. ( Затылочные мыщелки которые соединяют череп с позвоночником ) пропорционально малы по сравнению с большим затылочным отверстием (где спинной мозг переходит в череп). Крупные костные выступы неправильной формы лежат непосредственно за затылочными мыщелками. [5] : 246–249
Объемы мозга шести особей из Нгандонга, для которых можно рассчитать этот показатель, колеблются от 1013 до 1251 см3. 3 . Череп Нгави I имеет размер 1000 см. 3 ; и три черепа Самбунгмакана (соответственно) 1035; 917; и 1006 см 3 . Это составляет в среднем более 1000 см. 3 . [19] : 136 Для сравнения, средний рост современного человека составляет 1270 см. 3 для мужчин и 1130 см 3 для женщин со стандартным отклонением примерно 115 и 100 см. 3 . [20] Китайский H. erectus (от 780 до 250 тысяч лет назад) в среднем составляет примерно 1028 см. 3 и Javan H. erectus (исключая Нгандонг) около 933 см. 3 . В целом азиатские H. erectus имеют большой мозг, его средний размер составляет около 1000 см. 3 . [21] Основание черепной коробки и, следовательно, мозга, похоже, было скорее плоским, чем изогнутым. у Турецкое седло основания черепа, рядом с гипофизом , намного больше, чем у современных людей, что Вайденрайх в 1951 году осторожно приписывал увеличенной железе, вызвавшей необычайное утолщение костей. [5] : 285
Из двух известных большеберцовых костей большеберцовая кость A гораздо более прочная, чем голень B, и в целом соответствует большеберцовым костям неандертальцев. [5] Как и у других H. erectus , голени толстые и тяжелые. Судя по реконструированной длине 380 мм (15 дюймов), большеберцовая кость B могла принадлежать человеку ростом 158 см (5 футов 2 дюйма) и весом 51 кг (112 фунтов). Предполагается, что большеберцовая кость А принадлежала более крупному человеку. Азиатский H. erectus , для которого берутся оценки высоты (довольно небольшой размер выборки), обычно находится в диапазоне 150–160 см (4 фута 11 дюймов – 5 футов 3 дюйма), при этом индонезийский H. erectus в тропических условиях обычно оценивается по более высокий конец, а континентальные экземпляры в более холодных широтах — нижний конец. Единственный фрагмент таза из Нгандонга официально еще не описан. [19] : 151–152





Культура
[ редактировать ]Палеосреда обитания
[ редактировать ]На видовом уровне фауна Нгандонга в целом похожа на более древнюю фауну Кедунг Брубус примерно 800–700 тысяч лет назад, во время массовой иммиграции крупных видов млекопитающих на Яву, включая азиатских слонов и стегодонов . Другая фауна Нгандонга включает тигра Panthera tigris soloensis , малайского тапира , бегемота Hexaprotodon , самбарского оленя , водяного буйвола , корову Bos palaesondaicus , свиней и макак, питающихся крабами . Они соответствуют открытой лесной среде. [22] Присутствие обыкновенного журавля на близлежащем месте Ватуаланг того времени может указывать на гораздо более прохладные условия, чем сегодня. [23] Самые засушливые условия, вероятно, соответствовали ледниковому максимуму примерно 135 000 лет назад, обнажив Зондский шельф и соединив основные индонезийские острова с континентом. К 125 000 лет назад климат стал намного влажнее, что сделало Яву островом и позволило расширить тропические леса. Это вызвало смену фауны Нгандонг фауной Пунунг, которая представляет собой современный комплекс животных Явы, хотя более типичная фауна Пунунга, а именно орангутанги и гиббоны , вероятно, не могла проникнуть на остров до тех пор, пока не была воссоединена с континентом через 80 000 лет. назад. [22] H. erectus , специалист по биомам лесов и саванн, вероятно, вымер с потерей последнего рефугиума с открытой средой обитания. [24]
Он. soloensis была последней популяцией в долгой истории заселения острова Ява H. erectus , начавшейся от 1,51 до 0,93 миллиона лет назад на стоянке Сангиран, продолжавшейся от 540 до 430 тысяч лет назад на стоянке Тринил и, наконец, от 117 до 108 тысяч лет назад. лет назад в Нгандонге. Если дата для Человека-одиночки верна, то они представляют собой конечную популяцию H. erectus , которая укрывалась в последних убежищах открытой среды обитания в Восточной Азии до захвата тропических лесов. До иммиграции современных людей Юго-Восточная Азия позднего плейстоцена также была домом для H. floresiensis, эндемичного для острова Флорес , Индонезия, и H. luzonensis, эндемичного для острова Лусон , Филиппины. Генетический анализ современных популяций Юго-Восточной Азии указывает на широкое распространение денисовцев ( вид, который в настоящее время можно узнать только по их генетической подписи) по Юго-Восточной Азии, после чего они скрещивались с иммигрировавшими современными людьми 45,7 и 29,8 тысяч лет назад. [7] Геномное исследование 2021 года показывает, что, за исключением денисовцев, современные люди никогда не скрещивались ни с одним из этих эндемичных видов человека, за исключением случаев, когда потомство было нежизнеспособным или гибридные линии с тех пор вымерли. [15]
Судя по огромному количеству экземпляров, захороненных в Нгандонге в то же время, до извержения вулкана, которое привело к их погребению, могла существовать значительная популяция H. e Soloensis , но численность популяции трудно точно определить. Участок Нгандонг находился на некотором расстоянии от северного побережья острова, но неясно, где находились южная береговая линия и устье реки Соло. [3]
Технология
[ редактировать ]
В 1936 году, изучая фотографии, сделанные голландским археологом Питером Винсентом ван Штайном Калленфельсом , Оппеноорт заметил несколько сломанных костных останков животных, в первую очередь повреждения большого черепа тигра и некоторых оленьих рогов , которые он считал доказательством технологии костей . некоторых оленьих рогов имелся резной птичий череп, который Он предположил, что на конце можно было использовать в качестве топора. В 1951 году Вайденрайх выразил свой скептицизм, поскольку кости неизменно были повреждены рекой, а возможно, крокодилами и другими естественными процессами, утверждая вместо этого, что ни одна из костей не содержит достоверных доказательств человеческой модификации. Оппеноорт далее предположил, что длинный кусок кости, вырезанный с волнистым узором с обеих сторон, использовался в качестве гарпуна , похожего на гарпуны, изготовленные в мадленский период в Европе, но Вайденрайх интерпретировал его как наконечник копья. Вайденрайх обратил внимание на аномальные жала скатов, обитающих внутри страны в Нгандонге, которые, как он предположил, были собраны Соло Мэном для использования в качестве кинжалов или наконечников стрел, как у некоторых недавних народов южной части Тихого океана. [5] : 216–218 Неясно, может ли эта очевидная технология костей быть связана с Человеком-одиночкой или более поздней современной человеческой деятельностью. [23] хотя Тринил Х. э. Популяция прямоходящих , похоже, работала с таким материалом, изготавливая скребки из панцирей псевдодонов и, возможно, вскрывая их зубами акулы . [25]
Оппеноорт также обнаружил идеально круглый андезитовый каменный шар из Нгандонга, обычное явление в долине Соло, диаметром от 67 до 92 мм (от 2,6 до 3,6 дюйма). Кроме того, подобные шары были обнаружены в современных и более молодых стоянках европейского мустьерского и африканского среднего каменного века , столь же древних, как и африканские ашельские стоянки (особенно Олоргесайли , Кения). [5] На Яве они были найдены в Ватуаланге (современном Нгандонге) и Сангиране. [26] Традиционно их интерпретировали как болы (связанные попарно или по трое и бросаемые в качестве охотничьего оружия), а также индивидуально брошенные снаряды, дубинки или инструменты для обработки растений или разрушения костей. В 1993 году американские археологи Кэти Шик и Николас Тот продемонстрировали, что сферическую форму можно воспроизвести, просто если использовать камень в качестве молотка в течение длительного периода. [27]
В 1938 году фон Кенигсвальд вернулся на стоянку Нгандонг вместе с археологами Хельмутом де Терра , Халламом Л. Мовиусом и Пьером Тейяром де Шарденом, чтобы собрать каменные ядра и отщепы (то есть каменные орудия ). Из-за износа, вызванного рекой, трудно с уверенностью определить, что некоторые из этих камней являются настоящими орудиями труда. Они маленькие и простые, обычно меньше 5 см (2 дюйма) и сделаны чаще всего из халцедона (но также кремня и яшмы ), выброшенных рекой. Несколько вулканических пород и фрагментов дерева, судя по всему, были превращены в сверхмощные рубящие инструменты. [28] В 1973 году на стоянке Самбунгмакан неподалеку был обнаружен односторонний чоппер (а также отщеп) из андезита. [23] [26] Из-за того, что инструментов было обнаружено мало, невозможно отнести Solo Man к какой-либо отдельной отрасли . [23] [26] Как и во многих других стоянках Юго-Восточной Азии, предшествовавших современным людям, на стоянке Нгандонг отсутствуют сложные чопперы, ручные топоры или какие-либо другие сложные рубящие инструменты, характерные для ашельских стоянок Западной Евразии и Африки. В 1944 году Мовиус предположил, что это произошло из-за большого технологического разрыва между западным и восточным H. erectus (« линия Мовиуса »), вызванного серьезным различием в среде обитания (открытая территория по сравнению с тропическими лесами), как обычно интерпретируются рубящие инструменты. как свидетельство охоты на крупную дичь, которая, по его мнению, была возможна только тогда, когда люди расселились на открытые равнины. [28]

Хотя строгая «линия Мовиуса» больше не получила должного подтверждения с открытием некоторых технологий ручных топоров в Восточной Азии среднего плейстоцена, ручные топоры все еще заметно редки и грубы в Восточной Азии по сравнению с западными современниками. Это было объяснено следующим образом: ашельские популяции появились в Африке после расселения человека по Восточной Азии (но для этого потребовалось бы, чтобы две популяции оставались разделенными в течение почти двух миллионов лет); В Восточной Азии было сырье более низкого качества, а именно кварц и кварцит (но в некоторых местностях Китая из этих материалов производились ручные топоры, и Восточная Азия не полностью лишена полезных ископаемых более высокого качества); Восточноазиатский H. erectus использовал биоразлагаемый бамбук вместо камня для рубящих инструментов (но это сложно проверить); или в Восточной Азии была более низкая плотность населения, и в целом после нее осталось мало инструментов (хотя демографию трудно приблизительно оценить по летописи окаменелостей). [29]
Возможный каннибализм
[ редактировать ]В 1951 году Вайденрайх и фон Кенигсвальд отметили серьезные травмы у Черепов IV и VI, которые, по их мнению, были вызваны режущим и тупым инструментом соответственно. На них имеются следы воспаления и заживления, поэтому люди, вероятно, пережили ссору. Вайденрайх и фон Кенигсвальд отметили, что были найдены только черепа без даже зубов, что весьма необычно. Таким образом, они интерпретировали, по крайней мере, Черепа IV и VI как жертвы «неудачного нападения», а другие черепа, у которых основание было сломано, «результат более успешных попыток убить жертв», предполагая, что это было сделано другими людьми, чтобы получить доступ и поглотить мозг. Они не были уверены, что это сделал соседний H. e. солоенсис , или «более развитыми людьми, которые доказали бы свою «превосходную» культуру, убивая своих более примитивных собратьев». Последний сценарий уже был предложен для пекинского человека (имеющего столь же заметную патологию) французским палеонтологом. Марселлин Буль в 1937 году. Тем не менее, Вайденрайх и фон Кенигсвальд признали, что некоторые травмы могли быть связаны с извержением вулкана. Фон Кенигсвальд предположил, что существуют только тюбетейки, потому что Человек-Соло превращал черепа в чаши из черепов , но Вайденрайх отнесся к этому скептически, поскольку зазубренные края Черепов I, V и X не очень подходят для этой цели. [5] : 238–239
Каннибализм и ритуальная охота за головами также были предложены для стоянок Тринил, Сангиран и Модджокерто (все на Яве) на основании очевидного отсутствия каких-либо останков, кроме тюбетейки. Это было подкреплено исторической практикой охоты за головами и каннибализма в некоторых современных индонезийских, австралийских и полинезийских группах, которые в то время считались потомками этих популяций H. erectus . В 1972 году Джейкоб альтернативно предположил, что, поскольку основание черепа слабее, чем шапка, и поскольку останки были перевезены через реку с большими камнями и валунами, это было чисто естественным явлением. Что касается отсутствия остальной части скелета, то если фактором было хищничество тигров, тигры обычно оставляют только голову, поскольку на ней меньше всего мяса. Кроме того, материал Нгандонга, особенно Черепа I и IX, был поврежден во время раскопок, очистки и подготовки. [30]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б Сяо-Пей, Ю. (2014). «Эволюционный азиатоцентризм, пекинский человек и истоки китаецентрического этнонационализма». Журнал истории биологии . 47 (4): 585–625. дои : 10.1007/s10739-014-9381-4 . ПМИД 24771020 . S2CID 23308894 .
- ^ Jump up to: а б с д Оппеноорт, WFF (1932). «Человек-одиночка — новый ископаемый череп». Научный американец . 147 (3): 154–155. Бибкод : 1932SciAm.147..154O . doi : 10.1038/scientificamerican0932-154 . JSTOR 24966028 .
- ^ Jump up to: а б с д и ж г час я дж к Хаффман, Огайо; де Вос, Дж.; Берхаут, AW; Азиз, Ф. (2010). «Переоценка происхождения человека прямоходящего из Нгандонга (Ява) 1931–1933 годов, подтверждение происхождения костного пласта, о котором сообщили первооткрыватели » Палеоантропология . 2010 : 1–60. дои : 10.4207/PA.2010.ART34 . Архивировано из оригинала 0 октября 2021 г. Получено 1 января 2021 г.
- ^ Тобиас, ПП (1976). «Жизнь и времена Ральфа фон Кенигсвальда: выдающийся палеонтолог». Журнал эволюции человека . 5 (5): 406–410. дои : 10.1016/0047-2484(76)90082-8 .
- ^ Jump up to: а б с д и ж г час я дж к Вайденрайх, Ф .; фон Кенигсвальд, GHR (1951). «Морфология одиночного человека» . Антропологические документы AMNH . 43 . HDL : 2246/297 . Архивировано из оригинала 30 марта 2021 г. Проверено 4 апреля 2021 г.
- ^ Хаффман, Огайо; де Вос, Дж.; Бальзо, А.; Берхаут, AW; Войт, Б. (2010). «Массовая смерть и лахары в тафономии костного пласта Нгандонг Homo erectus и вулканизм в летописях гомининов восточной Явы» . Тезисы докладов собраний Общества палеоантропологов 2010 г., Палеоантропология : A14. Архивировано из оригинала 29 июня 2021 г. Проверено 29 июня 2021 г.
- ^ Jump up to: а б с д Ризал, Ю.; Вестэуэй, Исландия; Заим, Ю.; Берга; Беттис III, Э.А.; Морвуд, MJ; Хаффман, Огайо; Грин, Р.; Жоаннес-Бойо, Р.; Бейли, РМ; Сидарто; Вестэуэй, MC ; С роговицей, И.; Мур, штат Вашингтон; Стори, М.; Азиз, Ф.; Суминто; Чжао, Дж.; Более того; Сипола, Мэн; Ларик, Р.; Зонневельд, Ж.-П.; Скотт, Р.; Путт, С.; Чочон, РЛ (2020). «Последнее появление Homo erectus в Нгандонге, Ява, 117 000–108 000 лет назад» Природа 577 (7790): 381–385. дои : 10.1038/ s41586-019-1863-2 ISSN 0028-0836 . ПМИД 31853068 . S2CID 209410644 .
- ^ Jump up to: а б Свишер, III, CC; Ринк, WJ; Антон, Южная Каролина; Шварц, HP; Кертис, Г.Х.; Видиасморо, А.С. (1996). «Последний Homo erectus с Явы: потенциальная современность Homo sapiens в Юго-Восточной Азии». Наука . 274 (5294): 1870–1874. Бибкод : 1996Sci...274.1870S . дои : 10.1126/science.274.5294.1870 . JSTOR 2891688 . ПМИД 8943192 . S2CID 22452375 .
- ^ Jump up to: а б Ёкояма, Ю.; Фальгер, К.; Шема, Ф.; Джейкоб, Т. (2008). «Гамма-спектрометрическое датирование черепов позднего человека прямоходящего из Нгандонга и Самбунгмакана, Центральная Ява, Индонезия». Журнал эволюции человека . 55 (2): 274–277. дои : 10.1016/jhevol.2008.01.006 . ПМИД 18479734 .
- ^ О'Коннелл, Дж. Ф.; Аллен, Дж.; Уильямс, MAJ; Уильямс, АН; Терни, CSM; Спунер, Северная Каролина; Камминга, Дж.; Браун, Г.; Купер, А. (2018). «Когда Homo sapiens впервые достиг Юго-Восточной Азии и Сахула?» . Труды Национальной академии наук . 115 (34): 8482–8490. Бибкод : 2018PNAS..115.8482O . дои : 10.1073/pnas.1808385115 . ПМК 6112744 . ПМИД 30082377 .
- ^ Индриати, Э.; Свишер III, CC; Лепре, К.; Куинн, РЛ; Суриянто, РА; Хаскарьо, Арканзас; Грин, Р.; Фейбель, CS; Побинер, Б.Л.; Обер, М.; Лиз, В.; Энтони, Южная Каролина (2011). «Эпоха 20-метровой одиночной речной террасы, Ява, Индонезия и выживание человека прямоходящего в Азии » ПЛОС Один . 6 (6): e21562. Бибкод : 2011PLoSO...621562I . дои : 10.1371/journal.pone.0021562 . ПМК 3126814 . ПМИД 21738710 .
- ^ Jump up to: а б с д и Курно, Д. (2011). «150-летняя загадка: устойчивость черепа и ее влияние на происхождение австралийских аборигенов» . Международный журнал эволюционной биологии . 2011 : 2–3. дои : 10.4061/2011/632484 . ПМК 3039414 . ПМИД 21350636 .
- ^ Шварц, Дж. Х .; Таттерсолл, И. (2010). «Ископаемые свидетельства происхождения Homo sapiens » . Американский журнал физической антропологии . 143 (С51): 96–98, 101–103. дои : 10.1002/ajpa.21443 . ПМИД 21086529 .
- ^ Хауэллс, WW (1981). «5. Homo erectus в происхождении человека: идеи и проблемы». Homo erectus: Документы в честь Дэвидсона Блэка . Университет Торонто Пресс. п. 68. ИСБН 9781487585372 . JSTOR 10.3138/j.ctvcj2jdw.11 .
- ^ Jump up to: а б Тейшейра, JC; Джейкобс, Г.С.; Стрингер, К. ; Тьюк, Дж.; Худжашов Г.; Пурномо, Джорджия; Судойо, Х.; Кокс, член парламента; Тоблер, Р.; Терни, CSM; Купер, А.; Хелген, КМ (2021). «Широко распространенное денисовское происхождение на островах Юго-Восточной Азии, но нет свидетельств существенной примеси суперархаичных гомининов» . Экология и эволюция природы . 5 (5): 616–624. Бибкод : 2021NatEE...5..616T . дои : 10.1038/s41559-021-01408-0 . ПМИД 33753899 . S2CID 232323599 .
- ^ Jump up to: а б Кайфу, Ю.; Азиз, Ф.; Индриати, Э.; Джейкоб, Т.; Курниаван, И.; Баба, Х. (2008). «Краниальная морфология яванского человека прямоходящего : новые доказательства непрерывной эволюции, специализации и окончательного вымирания». Журнал эволюции человека . 55 (4): 578. doi : 10.1016/j.jhevol.2008.05.002 . ПМИД 18635247 .
- ^ Зейтун, В.; Детройт, Ф.; Гримо-Эрве, Д.; Видианто, Х. (2010). «Одинокий человек, о котором идет речь: сходящиеся взгляды на разделение индонезийского Homo erectus на две категории» (PDF) . Четвертичный интернационал . 223–224: 288. Бибкод : 2010QuInt.223..281Z . дои : 10.1016/j.quaint.2010.01.018 . S2CID 84764668 . Архивировано (PDF) из оригинала 29 апреля 2021 г. Проверено 22 мая 2021 г.
- ^ Ли, Х.; Руан, Дж.; Се, З.; Ван, Х.; Лю, В. (2007). «Исследование важнейших геометрических характеристик черепов живых людей с использованием методов анализа медицинских изображений». Международный журнал безопасности транспортных средств . 2 (4): 345. doi : 10.1504/IJVS.2007.016747 .
- ^ Jump up to: а б с Антон, СК (2003). «Естественная история человека прямоходящего †» . Американский журнал физической антропологии . 122 (37): 136–152. дои : 10.1002/ajpa.10399 . ПМИД 14666536 .
- ^ Аллен, Дж. С.; Дамасио, Х.; Грабовски, Т.Дж. (2002). «Нормальные нейроанатомические изменения в мозге человека: объемное МРТ-исследование». Американский журнал физической антропологии . 118 (4): 351. дои : 10.1002/ajpa.10092 . ПМИД 12124914 . S2CID 21705705 .
- ^ Антон, Южная Каролина; Табоада, ХГ; Миддлтон, ER; Дождевая вода, CW; Тейлор, AB; Тернер, TR; Тернквист, Дж. Э.; Вайнштейн, К.Дж.; Уильямс, Ю.А. (2016). «Морфологическая изменчивость Homo erectus и истоки пластичности развития» . Философские труды Королевского общества Б. 371 (1698): 20150236. doi : 10.1098/rstb.2015.0236 . ПМЦ 4920293 . ПМИД 27298467 .
- ^ Jump up to: а б Ван ден Берг, Грузия; де Вос, Дж.; Сондаар, ПЮ (2001). «Позднечетвертичная палеогеография эволюции млекопитающих на Индонезийском архипелаге». Палеогеография, Палеоклиматология, Палеоэкология . 171 (3–4): 387–392. Бибкод : 2001PPP...171..385В . дои : 10.1016/s0031-0182(01)00255-3 .
- ^ Jump up to: а б с д Беллвуд, П. (1997). « Человек прямоходящий в Сундаланде». Предыстория Индо-Малазийского архипелага: исправленное издание . Гонолулу: Издательство Гавайского университета . стр. 65–66. ISBN 978-1-921313-11-0 . JSTOR j.ctt24hf81.5 .
- ^ Луис, Дж.; Робертс, П. (2020). «Экологические факторы вымирания мегафауны и гомининов в Юго-Восточной Азии». Природа . 586 (7829): 402–406. Бибкод : 2020Natur.586..402L . дои : 10.1038/s41586-020-2810-y . hdl : 10072/402368 . ПМИД 33029012 . S2CID 222217295 .
- ^ Йоорденс, JCA; д'Эррико, Ф.; Весселинг, ФП; Манро, С.; де Вос, Дж.; Валлинга, Дж.; Анкьергаард, К.; Рейманн, Т.; Вейбранс, младший; Койпер, К.Ф.; Мучер, HJ; Кокеньо, Х.; Прие, В.; Йоостен, И.; ван Ос, Б.; Шульп, А.С.; Пануэль, М.; ван дер Хаас, В.; Люстенхауэр, В.; Реймер, JJG; Робрукс, В. (2015). « Человек прямоходящий в Триниле на Яве использовал ракушки для изготовления инструментов и гравировки». Природа . 518 (7538): 228–231. Бибкод : 2015Natur.518..228J . дои : 10.1038/nature13962 . ПМИД 25470048 . S2CID 4461751 .
- ^ Jump up to: а б с Семах, Ф.; Симанджунтак, Т.; Дизон, Э.; Гайяр, К.; Семах, А.-М. (2014). «Островная Юго-Восточная Азия в нижнем палеолите». Смит, К. (ред.). Энциклопедия глобальной археологии . Springer Science+Business Media . стр. 3909, 3914. doi : 10.1007/978-1-4419-0465-2_1907 . ISBN 978-1-4419-0466-9 . S2CID 239106990 .
- ^ Шик, К.Д. ; Тот, Н.П. (1994). «Создавая наше будущее: изготовление ранних каменных орудий». Заставить молчащие камни говорить: эволюция человека и зарождение технологий . Саймон и Шустер . стр. 132–133. ISBN 978-0-671-87538-1 .
- ^ Jump up to: а б Бартстра, Г.-Ж.; Соегондхо, С.; ван дер Вейк, А. (1988). «Человек Нгандонг: возраст и артефакты». Журнал эволюции человека . 17 (3): 332–335. дои : 10.1016/0047-2484(88)90074-7 .
- ^ Лисетт, С.Дж.; Бэ, CJ (2010). «Спор о линии Мовиуса: состояние дебатов». Мировая археология . 42 (4): 526–531. дои : 10.1080/00438243.2010.517667 . JSTOR 20799447 . S2CID 2209392 .
- ^ Джейкоб, Т. (1972). «Проблема охоты за головами и поедания мозгов среди мужчин плейстоцена в Индонезии». Археология и физическая антропология в Океании . 7 (2): 86–89. JSTOR 40386169 .