гомо-предшественник
| гомо-предшественник Временной диапазон: ранний плейстоцен , | |
|---|---|
 | |
| Окаменелости «Мальчика из Гран-Долины» АТД6-15 ( лобная кость ) АТД6-69 ( верхняя челюсть ) Национальный археологический музей, Мадрид | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Симииформы |
| Семья: | Гоминиды |
| Подсемейство: | Люди |
| Племя: | Люди |
| Род: | гомо |
| Разновидность: | † Х. предшественник |
| Биномиальное имя | |
| † Гомо-предшественник Бермудес де Кастро и др. , 1997 г. | |
Homo antecessor ( лат. «человек-первопроходец») — вымерший вид архаичного человека, зарегистрированный в испанской Сьерра-де-Атапуэрка , продуктивном археологическом объекте, от 1,2 до 0,8 миллиона лет назад во время раннего плейстоцена . Популяции этого вида, возможно, присутствовали в других местах Западной Европы и были одними из первых, кто заселил этот регион мира, отсюда и название. Первые окаменелости были найдены в пещере Гран-Долина этот вид был в 1994 году, а официально описан в 1997 году как последний общий предок современного человека и неандертальцев более традиционного H. heidelbergensis , вытеснив в этом положении . С тех пор H. antecessor был интерпретирован как ответвление линии современного человека, хотя, вероятно, оно ответвилось незадолго до разделения современного человека и неандертальца.
Несмотря на свою древность, лицо неожиданно похоже на лицо современного человека, а не на лицо других архаичных людей, а именно по своей общей плоскостности, а также по изгибу скулы, сливающейся с верхней челюстью, хотя эти элементы известны только по ювенильный экземпляр. Объем мозга мог составлять 1000 куб.см (61 куб.дюйм) или более, но неповрежденной черепной коробки обнаружено не было. Для сравнения, средний рост современного человека составляет 1270 см. 3 для мужчин и 1130 см 3 для женщин. Оценочный рост колеблется в пределах 162,3–186,8 см (5 футов 4 дюйма – 6 футов 2 дюйма). H. antecessor, возможно, был широкогрудым и довольно тяжелым, как неандертальцы, хотя конечности были пропорционально длинными, что чаще встречается у тропических популяций. Коленные чашечки тонкие, со слабо развитыми сухожильными креплениями. Ступни указывают на то, что H. antecessor ходил иначе, чем современные люди.
H. antecessor преимущественно производил простые орудия из гальки и чешуйчатого камня из кварца и кремня , хотя они использовали различные материалы. Эта индустрия имеет некоторое сходство с более сложной ашельской индустрией, характерной для современных африканских, а затем и европейских памятников. Группы, возможно, отправляли охотничьи отряды, которые в основном охотились на оленей в их саванне и смешанных лесах. Многие экземпляры H. antecessor были съедены , возможно, в рамках культурной практики. Нет никаких доказательств того, что они использовали огонь, и они также населяли внутренние районы Иберии только в теплые периоды, в противном случае предположительно отступая к побережью.
Таксономия
[ редактировать ]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
История исследований
[ редактировать ]
останками . Давно известно, что Сьерра-де-Атапуэрка на севере Испании изобилует ископаемыми Гран Долина («большая воронка») была впервые исследована на предмет окаменелостей археологом Франсиско Хорда Серда во время короткой поездки в этот регион в 1966 году, где он обнаружил несколько окаменелостей животных и каменные орудия. Ему не хватало ресурсов и рабочей силы, чтобы продолжать дальше. В 1976 году испанский палеонтолог Тринидад Торрес исследовал Гран-Долину на предмет окаменелостей медведя (он обнаружил останки Урсуса ), но спелеологическая группа Эдельвейса посоветовала ему продолжить исследование в близлежащем Сима-де-лос-Уэсос («костяная яма»). Здесь, в дополнение к множеству окаменелостей медведя, он также обнаружил архаичные окаменелости человека , что побудило к масштабному исследованию Сьерра-де-Атапуэрка, которое сначала возглавил испанский палеонтолог Эмилиано Агирре , но быстро перешло к Хосе Марии Бермудесу де Кастро и Эудальду Карбонеллу. и Хуан Луис Арсуага . Они возобновили раскопки Гран-Долины в 1992 году, а два года спустя обнаружили архаичные человеческие останки, которые в 1997 году они обнаружили. формально описан новый вид Homo как antecessor . [1] Голотип , представляет собой образец ATD6-5, фрагмент правой нижней челюсти сохранивший коренные зубы и обнаруженный с некоторыми изолированными зубами. В своем первоначальном описании Кастро и его коллеги утверждали, что этот вид был первым человеком, колонизировавшим Европу, отсюда и название « предшественник» ( с латыни «исследователь», «пионер» или «ранний поселенец»). [2]
25 м (82 фута) плейстоценовых отложений Гран Долины разделены на одиннадцать единиц , от TD1 до TD11 («тринчера долина» или «карстовая воронка»). H. antecessor был обнаружен в TD6, который впоследствии стал наиболее хорошо изученной единицей этого участка. [3] В первые полевые сезоны 1994–1995 годов бригада раскопок выкопала небольшую испытательную яму (чтобы проверить, требует ли установка дальнейшего исследования) на юго-восточном участке размером 6 м. 2 (65 кв. футов). [3] Человеческие окаменелости были впервые обнаружены Авророй Мартином Нахерой; Слой толщиной 30 см (12 дюймов), в котором они были найдены, в ее честь получил прозвище « Стратум Авроры ». [4] 13 м 2 Треугольная секция (140 кв. футов) в центральной части была раскопана в начале 2000-х годов. [3] Окаменелости человека также были найдены в северной части. Всего было обнаружено около 170 экземпляров H. antecessor . [3] Лучше всего сохранились ATD6-15 и ATD6-69 (возможно, принадлежащие одному и тому же человеку), которые наиболее четко проясняют анатомию лица. [2] Последующие полевые сезоны дали еще около шестидесяти экземпляров. [1] Обнаруженные части скелета H. antecessor включают: элементы лица, ключицы , предплечья, пальцев, коленей, а также несколько позвонков и ребер . [5]
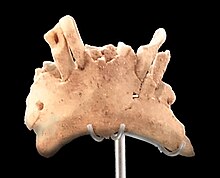
В 2007 году фрагмент нижней челюсти с несколькими зубами, ATE9-1, предварительно отнесенный Карбонеллом к H. antecessor , был обнаружен в близлежащем Sima del Elefante («слоновья яма») в отряде TE9 («trinchera elefante»), принадлежащем 20 – 25-летний человек. На этом месте также были обнаружены каменные отщепы и свидетельства бойни. [6] В 2011 году, после гораздо более глубокого анализа материала Сима дель Элефанте, Кастро и его коллеги не были уверены в классификации видов, решив оставить ее в Homo sp. (не высказывая мнения по поводу определения вида) в ожидании дальнейших открытий. [7]
Комплекс каменных орудий в Гран Долине во многом похож на несколько других современных инструментов в Западной Европе, которые могут представлять собой работу того же вида, хотя это неподтвержденно, поскольку во многих из этих мест не было обнаружено окаменелостей человека. [2] , Англия, было обнаружено пятьдесят следов, датируемых периодом от 1,2 до 800 000 лет назад В 2014 году в Хапписбурге , которые потенциально можно отнести к группе H. antecessor, поскольку это единственный человеческий вид, идентифицированный в то время в Западной Европе. [8]
Классификация
[ редактировать ]Лицо H. antecessor неожиданно похоже на лицо современных людей по сравнению с другими архаичными группами, поэтому в своем первоначальном описании Кастро и коллеги классифицировали его как последнего общего предка современных людей и неандертальцев , вытеснив H. heidelbergensis . в этом качестве [2] [5] [1] В последующие годы анатомия лица стала объектом пристального внимания. [1]
В 2001 году французский палеоантрополог Жан-Жак Юблен предположил, что остатки Гран Долины и современного Тигеннифа остаются из Алжира (обычно классифицируемого как Homo ergaster [=? Homo erectus ], первоначально « Atlantanthropus mauritanicus »). [а] ) представляют одну и ту же популяцию, поскольку четырнадцать из пятнадцати особенностей зубов, перечисленных Кастро и его коллегами для H. antecessor, также были идентифицированы в среднем плейстоцене Северной Африки; это означало бы, что H. antecessor является младшим синонимом Homo mauritanicus , т.е. людей Гран Долины и Тигеннифа следует отнести к последним. [12] В 2007 году Кастро и его коллеги изучили окаменелости и обнаружили, что останки Тигеннифа намного крупнее H. antecessor и стоматологически похожи на другие африканские популяции. Тем не менее, они по-прежнему рекомендовали возродить mauritanicus для размещения всех североафриканских экземпляров раннего плейстоцена как « H. ergaster mauritanicus ». [13]
В 2007 году приматолог Эстебан Сармьенто и его коллеги поставили под сомнение легитимность H. antecessor как отдельного вида, поскольку большая часть анатомии черепа неизвестна; H. heidelbergensis известен примерно из того же времени и региона; и поскольку типовой экземпляр был ребенком (предположительно характерные черты могли исчезнуть с возрастом). Такая реструктуризация лица, как они утверждали, также может быть вызвана региональной климатической адаптацией, а не видообразованием. [14] В 2009 году американский палеоантрополог Ричард Кляйн заявил, что он скептически относится к тому, что H. antecessor был предком H. heidelbergensis , интерпретируя H. antecessor как «ответвление H. ergaster [из Африки], которое исчезло после неудачной попытки колонизировать южную Европу». [15] Точно так же в 2012 году британский физический антрополог Крис Стрингер считал H. antecessor и H. heidelbergensis двумя разными линиями, а не имеющими отношения предка и потомка. [9] В 2013 году антрополог Сара Фрейдлайн и ее коллеги предположили, что современное человекоподобное лицо несколько раз независимо развивалось среди Homo . [16] В 2017 году Кастро и его коллеги признали, что H. antecessor может быть, а может и не быть предком современного человека, хотя если это не так, то он, вероятно, разделился незадолго до разделения современного человека и неандертальца. [17] В 2020 году голландский молекулярный палеоантрополог Фридо Велкер и его коллеги пришли к выводу, что H. antecessor не является предком современного человека, проанализировав древние белки, собранные из зуба ATD6-92. [18]
Возраст и тафономия
[ редактировать ]
Раскопки 2003–2007 годов выявили гораздо более сложную стратиграфию , чем считалось ранее, и TD6 был разделен на три подразделения, охватывающие тринадцать слоев и девять осадочных фаций (тела горных пород, отличающиеся от соседних тел). Присутствие человека зафиксировано в субъединицах 1 и 2, а также в фациях A, D1 и F. Случайно ориентированные разрозненные кости отлагались в фации D1 слоя TD6.2.2 (субъединица 2 TD6, слой 2) и фации F слоев TD6.2.2. и TD6.2.3, но в фации D они, по-видимому, были заметно сгруппированы в северо-западной части. Это может указывать на то, что их затащил в пещеру поток мусора . Что касается фации F, которая содержит наибольшее количество человеческих останков, они могли быть отложены низкоэнергетическим селевым потоком (соответствующим поведению поймы ) от главного входа на северо-запад, а также более сильным селевым потоком от другого входа на юг. . Окаменелости, отложенные речным путем (занесенные потоком воды), также были обнаружены в фации А в слоях TD6.2.2, TD6.2.1 и TD6.1.2, обозначенных известняковый гравий в пределах размера останков. Таким образом, H. antecessor , возможно, не обитал в пещере, хотя, по крайней мере, действовал поблизости. Только 5,6% окаменелостей имеют какие-либо следы выветривания на открытом воздухе, корнях и почве, что может означать, что они были отложены глубоко в пещере относительно вскоре после смерти. [3]
Человеческое заселение, по-видимому, происходило волнами, соответствующими периодам времени, когда существовала теплая и влажная среда обитания саванны (хотя берега рек, вероятно, поддерживали лесные массивы). Эти условия присутствовали только во время перехода от прохладных ледниковых периодов к теплым межледниковым периодам, после потепления климата и до того, как леса смогли расшириться и доминировать над ландшафтом. [20] Попытки датировки останков H. antecessor следующие:
- В 1999 году два зуба копытных из TD6 были датированы с помощью уран-тория возрастом от 794 до 668 тысяч лет назад, а палеомагнитно датированы более чем 780 000 лет назад. [21]
- В 2008 году возраст TE9 Сима-дель-Элефанте был ограничен 1,2–1,1 миллиона лет назад с использованием палеомагнетизма и космогенного датирования . [6]
- В 2013 году TD6 был датирован примерно от 930 до 780 тысяч лет назад с использованием палеомагнетизма, а также датирования уран-тория и электронного спинового резонанса (ЭПР) на большем количестве зубов. [22]
- В 2018 году ЭПР-датирование экземпляра H. antecessor ATD6-92 выявило возраст от 949 до 624 тысяч лет назад, что дополнительно ограничивается палеомагнитными данными до 772 000 лет назад. [23]
- В 2022 году ЭПР и термопереносная оптически стимулированная люминесценция отдельных зерен (SG TT-OSL) датировали открытие Гран Долины примерно 900 000 лет назад, а отложения от TD4 до TD6 - от 890 000 до 770 000 лет назад. Эти три единицы, вероятно, были отложены в течение периода менее 100 000 лет. [24]
, Испания, был обнаружен молочный зуб возрастом 1,4 миллиона лет До 2013 года, когда в Барранко-Леон , Орсе , это были старейшие человеческие окаменелости, известные в Европе. [25] хотя деятельность человека на континенте началась еще 1,6 миллиона лет назад, в Восточной Европе и Испании, о чем свидетельствуют каменные орудия. [26]
Анатомия
[ редактировать ]Череп
[ редактировать ]

Анатомия лица H. antecessor преимущественно известна по 10–11,5-летнему ребенку H. antecessor ATD6-69, поскольку немногие другие образцы лица являются фрагментарными. ATD6-69 поразительно похож на современных людей (а также на архаичных людей среднего плейстоцена Восточной Азии ) в отличие от архаичных людей западноевразийского или африканского среднего плейстоцена, включая неандертальцев. Наиболее примечательными чертами являются совершенно плоское лицо и изогнутый скулово-альвеолярный гребень (костная перемычка, соединяющая щеку с частью верхней челюсти , удерживающей зубы). В 2013 году антрополог Сара Фрейдлайн и ее коллеги статистически определили, что эти особенности не исчезнут с возрастом. H. antecessor предполагает, что лицо современного человека в прошлом эволюционировало и исчезало несколько раз, что вполне вероятно, поскольку на анатомию лица сильно влияет диета и, следовательно, окружающая среда. [16] Носовые кости напоминают кости современного человека. [2] Нижняя челюсть (нижняя челюсть) довольно изящная, в отличие от большинства других архаичных людей. Он имеет несколько архаичных черт, но форма нижнечелюстной вырезки напоминает современную человеческую, а альвеолярная часть (прилегающая к зубам) полностью вертикальна, как у современных людей. Как и у многих неандертальцев, медиальный крыловидный бугорок крупный. В отличие от большинства неандертальцев, у них нет ретромолярного пространства (большого зазора между последним коренным зубом и концом тела нижней челюсти). [17]
Верхние резцы имеют лопатовидную форму (язычная, или языковая, сторона отчетливо вогнутая), что характерно для других евразийских популяций человека, в том числе современных. Клыки видов несут поясную часть (выступ по направлению к основанию) и основной гребень (по направлению к средней линии), как у более производных , но сохраняют бугорки (небольшие выступы) возле кончика и граничащего с резцом, как у более архаичных видов. Коронки верхних премоляров довольно производные, почти симметричные, имеют лингвальный бугорок (со стороны языка), поясную кость и продольные бороздки со стороны щеки. Верхние коренные зубы имеют несколько черт, типичных для неандертальцев. С другой стороны, зубы нижней челюсти довольно архаичны. P 3 (первый нижний премоляр) имеет сильно асимметричную коронку и сложную зубов корневую систему . P 3 меньше, чем P 4, как и у более производных видов, но, как и у других ранних Homo , M 1 (первый нижний коренной зуб) меньше, чем M 2 , а бугорки коронок коренных зубов имеют Y-образную форму. Распределение эмаль неандертальская, с более толстыми слоями на периферии, чем на буграх. [17] На основе двух клыков (ATD6-69 и ATD6-13) толщина эмали и доля зуба, покрытого деснами, различаются в той же степени, что и у самцов и самок современного человека и многих других обезьян, поэтому это может быть связано с половым диморфизмом: у женщин зубы меньшего размера, относительно более толстая эмаль и меньшая доля покрытия десен. [27]
( Теменные кости каждая из которых представляет собой одну сторону задней части верхней части черепа) уплощены и соединяются на вершине по средней линии. Этот «палаточный» профиль также наблюдается у более архаичных африканских H. ergaster и азиатских H. erectus . Как и у H. ergaster , височный шиловидный отросток чуть ниже уха сращен с основанием черепа . Надбровные дуги выступающие. Верхний край чешуйчатой части височных костей (сбоку черепа) выпуклый, как у более производных видов. [17] объем мозга ATD6-15, возможно, принадлежавшего 11-летнему ребенку, мог составлять 1000 куб.см (61 куб.дюйм) или более Согласно измерениям лобной кости, . [б] [2] Для сравнения, средний рост современного человека составляет 1270 см. 3 для мужчин и 1130 см 3 для женщин со стандартным отклонением примерно 115 и 100 см. 3 . [28]
Торс
[ редактировать ]Особенно крупный образец ключицы взрослого человека ATD6-50, предположительно мужской, исходя из абсолютного размера, имел рост 162,3–186,8 см (5 футов 4 дюйма – 6 футов 2 дюйма), в среднем 174,5 см (5 футов 9 дюймов), исходя из о соотношении у современных индийцев длины ключицы и роста. взрослого человека Лучевая кость (кость предплечья), ATD6-43, которая могла быть мужской, исходя из абсолютного размера, или женской, исходя из грациозности, по оценкам, принадлежала человеку ростом 172,5 см (5 футов 8 дюймов) на основе среднего значения уравнений. среди нескольких современных популяций связывают радиальную длину с ростом. Судя по длине плюсневой кости (кости стопы), рост мужчины оценивается в 173 см (5 футов 8 дюймов), а у женщины - 168,9 см (5 футов 6 дюймов). Это все довольно схожие ценности. Для сравнения, средний рост западноевропейских неандертальцев составляет 165,3 см (5 футов 5 дюймов), а у ранних европейских современных людей - 178,4 см (5 футов 10 дюймов). [5] Голеностопный сустав приспособлен выдерживать высокие нагрузки, что может указывать на тяжелое и крепкое телосложение, очень похожее на неандертальцев. [29] Судя по взаимосвязи между длиной человеческого следа и размером тела, двенадцать отпечатков Гапписбурга , которые сохранились достаточно хорошо, чтобы их можно было измерить, соответствуют людям ростом от 93 до 173 см (от 3 футов 1 дюйма до 5 футов 8 дюймов), что может означать некоторое из путевых мастеров были дети. По этой логике, три самых больших следа — соответствующие росту 160 см (5 футов 3 дюйма), 163 см (5 футов 4 дюйма) и 173 см (5 футов 8 дюймов) — варьировались от 48 до 53 кг (от 106 до 117 фунтов) по весу. Оценки роста H. antecessor , H. heidelbergensis и неандертальцев примерно соответствуют друг другу. [8]

два атласа (первый шейный позвонок Известны ), что является исключительным, поскольку у архаичных людей эта кость встречается редко. Они неотличимы от современных людей. Для оси (второй шейный позвонок) угол остистого отростка (выступающий из позвонка) составляет около 19°, что сравнимо с неандертальцами и современным человеком, от H. ergaster расходится с небольшим углом около 8°. Позвоночное отверстие (в котором находится спинной мозг ) более узкое по сравнению с современными людьми. В остальном позвоночник в целом соответствует современному человеку. [5]
Есть одна известная (и неполная) ключица, ATD6-50, которая толще, чем у современного человека. Это может указывать на то, что у H. antecessor были длинные и плоские (платиклеидные) ключицы, как и у других архаичных людей. Это указывало бы на широкую грудь. Проксимальная кривизна (перекручивание кости на стороне, ближайшей к шее) на виде спереди находится на одном уровне с таковой у неандертальцев, но дистальная кривизна (со стороны плеча) гораздо более выражена. Грудина . узкая Акромион . (который простирается над плечевым суставом) мал по сравнению с акромионом современного человека [5] Лопатка H. аналогична всем видам Homo с типичным человеческим строением тела, что указывает на то, что antecessor не был таким искусным альпинистом, как человекообразные обезьяны или виды до эректусов , но был способен эффективно метать снаряды, такие как камни или копья. [30]
Конечности
[ редактировать ]Неполный радиус ATD6-43 оценивался в 257 мм (10,1 дюйма). Оно странно длинное и прямое для человека с далекого севера и напоминает пропорции, наблюдаемые у ранних современных людей и многих людей из тропических популяций. Это можно объяснить сохранением древней тропической формы с длинными конечностями, в отличие от неандертальцев, у которых развились более короткие конечности. Это также может указывать на высокий плечевой индекс (отношение длины лучевой кости к длине плечевой кости ). По сравнению с более поздними видами человека поперечное сечение радиального стержня довольно круглое и изящное по всей длине. Как и у архаичных людей, лучевая шейка (около локтя) длинная, что дает больше рычагов двуглавой мышце плеча . Как и у современных людей и H. heidelbergensis , но в отличие от неандертальцев и более архаичных гомининов, радиальный бугорок (костный выступ, выступающий чуть ниже лучевой шейки) расположен спереди (к передней стороне, когда рука обращена наружу). [5]
Как и у других архаичных людей, бедренная кость имеет развитую вертельную ямку и задний гребень. Эти черты сильно различаются среди современных человеческих популяций. Две известные коленные чашечки , ATD6-22 и ATD6-56, имеют субпрямоугольную форму в отличие от более распространенной подтреугольной, хотя и довольно узкие, как у современных людей. Они довольно маленькие и тонкие, у современных человеческих женщин они находятся на нижнем конце. Вершина коленной чашечки (область, которая не соединяется с другой костью) развита недостаточно, поэтому сухожилие надколенника остается мало прикрепленным . Медиальная (по направлению к средней линии) фасетка и латеральная (по бокам) фасетка коленного сустава примерно одинакового размера у ATD6-56, а медиальная больше у ATD6-22, тогда как у современных ATD6-22 обычно больше латеральная. люди. Боковая фасетка простирается на прямую плоскую область, а не ограничивается определенной выемкой широкой мышцы бедра , что является нечастым состоянием среди любого человеческого вида. [5]
Фаланги H. и плюсневые кости стопы сравнимы с таковыми у более поздних людей, но кость большого пальца ноги довольно прочная, что может быть связано с тем, как antecessor отталкивался от земли. Голеностопная кость (таранная кость) исключительно длинная и высокая, как и фасетка, где она соединяется с ногой (блоком), что может быть связано с тем, как H. antecessor ходил . Длинный блок обусловил короткую шейку таранной кости, которая соединяет головку таранной кости, соединяющуюся с пальцами ног, и тело таранной кости, соединяющееся с ногой. Это в некоторой степени сходно с состоянием, наблюдаемым у неандертальцев, которое обычно объясняется реакцией на тяжелое и крепкое тело, чтобы облегчить, как следствие, более высокую нагрузку на суставной хрящ голеностопного сустава. Это также позволило бы добиться большей гибкости . [29]
Темпы роста
[ редактировать ]
Музей естественной истории, Лондон
В 2010 году Кастро и его коллеги подсчитали, что ATD6-112, представленный постоянными верхними и нижними первыми молярами, умер в возрасте от 5,3 до 6,6 лет, исходя из скорости формирования зубов у шимпанзе (нижняя оценка) и современных людей (верхняя оценка). Коренные зубы почти не изношены, а это означает, что человек умер вскоре после прорезывания зуба, и что первое прорезывание коренных зубов произошло примерно в этом возрасте. Возраст находится в пределах вариаций современного человека, и эта веха развития может быть спорно коррелирована с историей жизни. Если это соотношение верно, у H. antecessor было продолжительное детство, что характерно для современных людей, в которых происходит значительное когнитивное развитие. [31]
Патология
[ редактировать ]На частичном лице ATD6-69 имеется эктопическая М. 3 (верхний левый третий моляр), где он прорезался неправильно, и это вызвало ретенцию М. 2 , где его извержение вообще было заблокировано. Хотя воздействие М. 3 довольно часто встречается у современных людей, в некоторых популяциях достигает пятидесяти процентов, поражение M 2 встречается редко, всего от 0,08 до 2,3%. Удар может привести к вторичным поражениям, таким как кариес , резорбция корня , кератоцисты и зубочелюстные кисты . [32]
На нижней челюсти ATE9-1 наблюдается сильное стирание зубов и стирание коронок зубов, а также резорбция кости у корня, настолько сильная, что обнажаются корневые каналы (чувствительная внутренняя часть) клыков. Травма соответствует заболеванию десен из-за перегрузки зубов, например, использования рта в качестве третьей руки для переноски предметов. Аналогичное состояние было также зарегистрировано в отношении более поздних останков Сима-де-лос-Уэсос, также находящихся на территории Сьерра-де-Атапуэрка. [33]
(костную шпору) размером 4,7 × 15 мм (0,19 × 0,59 дюйма) Левая коленная кость ATD6-56 имеет остеофит по нижнему (нижнему) краю. Остеофиты обычно формируются в ответ на стресс, вызванный остеоартритом , который может возникнуть в результате старости или неправильной нагрузки на сустав вследствие смещения костей или слабости связок . В случае ATD6-56 причиной, вероятно, была неправильная загрузка. К этому состоянию могут привести частые приседания и стояния на коленях, но если правая коленная кость ATD6-22 (не имеющая такой травмы) принадлежит одному и тому же человеку, то вряд ли это является причиной. Если это так, то поражение было вызвано местной травмой, такой как растяжение мягких тканей вокруг сустава из-за активности высокой интенсивности или перелом левого бедра и/или большеберцовой кости (это неподтвержденно, поскольку ни одна кость не связана с этим суставом). индивидуальный). [34]
В правой четвертой плюсневой кости ATD6-124 имеется повреждение длиной и шириной 25,8 мм × 8 мм (1,02 дюйма × 0,31 дюйма) на медиальной (по направлению к средней линии кости) стороне, соответствующее перелому марша . С этим заболеванием чаще всего сталкиваются солдаты, бегуны на длинные дистанции и люди, потенциально имеющие плоскостопие , у которых кости стопы разрушаются при повторяющихся нагрузках высокой интенсивности. Позже у неандертальцев развился гораздо более крепкий нижний скелет, возможно, способный противостоять такому утомительному передвижению по неровной местности. Хотя среди архаичных людей был выявлен только один другой пример этого состояния (в Сима-де-лос-Уэсос), маршевые переломы, вероятно, были для них обычной травмой, учитывая, что заживший перелом не оставляет видимых следов, а также их предполагаемый высокоинтенсивный образ жизни. [35]
Культура
[ редактировать ]Технология
[ редактировать ]
H. antecessor производил простые каменные орудия в Гран-Долине. Эта индустрия встречается повсюду в Испании раннего плейстоцена, особенно в Барранк-де-ла-Боэлья и близлежащей Галерии, и отличается подготовкой и заточкой кернов перед отслаиванием, наличием (сырых) бифасов и некоторой степенью стандартизации типов инструментов. Это имеет некоторое сходство с гораздо более сложной ашельской индустрией, характерной для африканских, а затем и европейских памятников. [19] Самые ранние свидетельства существования типичных ашельских инструментов происходят из Африки 1,75 миллиона лет назад, но типичный ашельский набор инструментов появляется в Западной Европе почти миллион лет спустя. Спорный вопрос, превратились ли эти ранние европейские памятники в европейскую ашельскую индустрию независимо от африканских аналогов, или ашельская индустрия была завезена из Африки и распространилась по Европе. В 2020 году французский антрополог Мари-Элен Монсель утверждала, что появление типичных ашельских бифасов 700 000 лет назад в Европе было слишком внезапным, чтобы быть результатом полностью независимой эволюции от местных технологий, поэтому должно было быть влияние Африки. [36] Ношение каменных орудий TD6 сопровождается неоднократным истиранием плоти, поэтому они, вероятно, использовались в качестве орудий разделки мяса. [19]
ТД6.3
[ редактировать ]В нижней части TD6.3 (подраздел 3 TD6) было обнаружено 84 каменных орудия, преимущественно мелких, немодифицированных кварцитовых гальки с ударными повреждениями, вероятно, полученными в результате ударов по таким предметам, как кость, а не при изготовлении более специализированных орудий. [19]
Хотя 41% комплекса разреза состоит из отщепов, они довольно грубые и крупные - в среднем 38 мм × 30 мм × 11 мм (1,50 дюйма × 1,18 дюйма × 0,43 дюйма) - либо в результате элементарных навыков скалывания (обработки камня), либо в результате трудностей в обработке. такие некачественные материалы. Они использовали униполярный продольный метод, отслаивая только одну сторону ядра , вероятно, чтобы компенсировать отсутствие предварительного планирования, предпочитая раскалывать гальку неправильной формы и, следовательно, более низкого качества. [19]
ТД6.2
[ редактировать ]


Большая часть каменных орудий находилась в нижней (старой) половине TD6.2: 831 каменное орудие. Кнепперы использовали гораздо более разнообразный набор материалов (хотя чаще всего это кремень ), что указывает на то, что они продвигались дальше в поисках лучшего сырья. Сьерра-де-Атапуэрка отличается обилием и разнообразием обнажений полезных ископаемых, подходящих для изготовления каменных орудий, в дополнение к кремню и кварцу , а именно кварциту, песчанику и известняку , которые можно собрать всего в 3 км (1,9 мили) от Гран-Долины. [19]
Они произвели гораздо меньше гальки и потратили больше времени на откалывание отщепов, но они не особенно экономно расходовали материалы, и примерно половина нуклеусов могла бы дать больше отщепов. Они дополнительно модифицировали заготовки неправильной формы, придавая им более удобную форму, прежде чем отслаивать их. Такое предварительное планирование позволило им использовать другие методы: центростремительный метод (отслаивание только краев ядра) и биполярный метод (укладывание ядра на наковальню и ударение по нему молотком ). Всего имеется 62 чешуйки высотой менее 20 мм (0,79 дюйма) и 28 чешуек высотой более 60 мм (2,4 дюйма). Есть три отщепа заметно более высокого качества, тоньше и длиннее остальных, которые, возможно, были изготовлены одним и тем же человеком. Встречаются и ретушированные орудия: насечки, шипы, зубчики , острия, скребки и одинарный чоппер . Эти небольшие ретушированные орудия редки в европейском раннем плейстоцене. [19]
ТД6.1
[ редактировать ]В TD6.1 было найдено 124 каменных орудия, но они плохо сохранились, поскольку это место также использовалось гиенами в качестве уборной, а моча разъедала это место. В слое отсутствуют галька и нуклеусы, а 44 каменных орудия неопределимы. Чешуйки намного меньше: в среднем 28 мм × 27 мм × 11 мм (1,10 дюйма × 1,06 дюйма × 0,43 дюйма), десять из них имеют размер менее 20 мм (0,79 дюйма) и только три превышают 60 мм (2,4 дюйма). [19]
Похоже, они использовали те же методы, что и люди, производившие инструменты TD6.2. Они ретушировали только более крупные отщепы, четырнадцать таких орудий в среднем имели размеры 35 × 26 × 14 мм (1,38 × 1,02 × 0,55 дюйма): один отщеп с ретушью по краям, одна выемка, три шипа, семь зубчатых боковых скребков и одно зубчатое острие. . [19]
Огонь и палеоклимат
[ редактировать ]Из TD6 было собрано лишь несколько частиц древесного угля , который, вероятно, возник в результате пожара за пределами пещеры. Нет никаких свидетельств использования огня или сжигания костей (приготовления пищи) в эпизодах оккупации Гран Долины. В других частях мира надежные свидетельства использования огня не появляются в археологических записях примерно 400 000 лет назад. [37] В 2016 году кости мелких млекопитающих, сгоревшие при пожарах с температурой выше 600 °C (1112 °F), были обнаружены в отложениях возрастом от 780 до 980 тысяч лет в Куэва-Негра на юге Испании, которые потенциально могли принадлежать человеку. Источником такой высокой температуры обычно (хотя и не всегда) является костер, а не естественные лесные пожары. [38]

Вместо того, чтобы использовать огонь, эти ранние европейцы, вероятно, физиологически противостояли холоду, например, придерживаясь диеты с высоким содержанием белка для поддержания повышенного метаболизма. [37] Несмотря на ледниковые циклы , климат, вероятно, был таким же или на несколько градусов теплее по сравнению с сегодняшним: самая холодная средняя температура достигала 2 ° C (36 ° F) где-то в декабре и январе, а самая жаркая в июле и августе - 18 ° C. (64 ° F). Температура замерзания могла достигаться с ноября по март, но присутствие оливковых деревьев и дуба предполагает, что замерзание было нечастым явлением. [14] TE9 также указывает на в целом теплый климат. [6] Следы Хапписбурга пролегали по эстуариев илистым отмелям с открытыми лесами, где преобладали сосны , ели , березы , а в более влажных районах ольха , с участками вереска и лугов; растительность соответствует более прохладному началу или концу межледниковья. [8]
H. antecessor, вероятно, мигрировал с побережья Средиземного моря во внутренние районы Иберии , когда более холодные ледниковые периоды переходили в более теплые межледниковья, и доминировали теплые луга, покидая этот регион в любое другое время. Возможно, во время миграции они следовали за водоемами, в случае Сьерра-де-Атапуэрка, скорее всего, за рекой Эбро . [20]
Еда
[ редактировать ]Были обнаружены окаменелости шестнадцати видов животных. [39] случайно смешанный [4] с материалом H. antecessor в Гран-Долине, включая вымершего кустарникового оленя , вымерший вид лани Dama vallonetensi , вымерший подвид благородного оленя Cervus elaphus acoronatus , вымершего зубра Bison voigstedtensi , вымершего носорога Stephanorhinus etruscus , вымершая лошадь Equus stenonis , вымершая лисица Vulpes praeglacialis , вымерший медведь Ursus dolinensis , вымерший волк Canis mosbachensis , пятнистая гиена , кабан , неопределенные виды мамонта , обезьяны и рыси . На некоторых экземплярах первых восьми видов и обезьянах наблюдаются следы порезов, соответствующие резне, при этом около 13% всех останков Гран Долины имеют некоторые свидетельства человеческой модификации. Олени - наиболее часто забиваемые животные: 106 экземпляров. Судя по всему, жители, когда это было возможно, приносили обратно целые трупы, а из более крупных карьеров — только конечности и черепа. Это указывает на то, что предшественник Gran Dolina H. отправлял охотничьи отряды, которые убивали и вытаскивали добычу, чтобы поделиться со всей группой, а не каждый человек добывал пищу исключительно для себя, что свидетельствует о социальном сотрудничестве и разделении труда. Менее 5% всех останков сохраняют повреждения, нанесенные плотоядными животными: в двух случаях следы зубов перекрывают порезы неопознанного животного, что может указывать на то, что животные иногда питались мусором. H. antecessor . Остатки [39]

На стоянке Сима-дель-Элефанте встречаются лань, кустарниковый олень, носороги, E. stenonis , C. mosbachensis , U. dolinensis , вымершая большая кошка Panthera gombaszoegensis , вымершая рысь Lynx issiodorensis , вымершая лисица Vulpes alopecoides , несколько крысы , землеройки и кролики , а также неопределенные виды макак , кабанов, бизонов и бобров . Крупные млекопитающие чаще всего представлены длинными костями , некоторые из которых расколоты, предположительно для доступа к костному мозгу . Некоторые другие имеют следы ударов и удаления кожи. [6] Они также разделывали черепаху Германа — легкодоступный источник мяса, учитывая, насколько медленно черепахи передвигаются. [40]
Прохладная и влажная горная среда способствовала росту оливковых, мастиковых , буковых , фундуковых и каштановых деревьев, которые H. antecessor, возможно, использовал в качестве источников пищи, хотя они становятся более распространенными в TD7 и TD8 по мере развития межледниковья и изменения окружающей среды. более влажный. В единице H. antecessor TD6 пыльца преимущественно происходит от можжевельника и дуба. Деревья, вероятно, росли вдоль рек и ручьев, тогда как на остальных холмах и хребтах преобладали травы. [14] Особи TD6, по-видимому, также употребляли каркас , который в исторические времена использовался скорее из-за своих лечебных свойств, чем для утоления голода, поскольку в этих ягодах содержится очень мало мяса. [41]
Нет никаких доказательств того, что H. antecessor мог владеть огнем и готовить, и аналогичным образом стертость коренных зубов указывает на более частое потребление более грубых и механически сложных продуктов, чем более поздние европейские виды, таких как сырое, а не приготовленное мясо и подземные органы хранения . [42]
Каннибализм
[ редактировать ]Восемьдесят экземпляров H. antecessor молодых взрослых и детей из Гран-Долины имеют следы порезов и трещины, указывающие на каннибализм . [5] [39] и H. antecessor - второй по распространенности вид, имеющий следы забоя. [39] Человеческие тела использовались эффективно, и это может быть причиной того, что большинство костей раздроблено или иным образом сильно повреждено. Целых черепов нет, элементы лицевой и задней части черепа обычно выстукивают, а прикрепления мышц на лице и основании черепа отсекают. Интенсивная модификация лица, вероятно, была направлена на доступ к мозгу. Вероятно, был поражен темя головы, в результате чего на зубах по линии десен остались шрамы от удара. На нескольких фрагментах черепа наблюдается шелушение. [4]
На ребрах также имеются следы порезов вдоль прикрепления мышц, соответствующие удалению кожи, а у ATD6-39 имеются порезы по всей длине ребра, что может быть связано с выпотрошением. Мышцы затылка были отрезаны, а голова и шея, вероятно, были отделены от тела. Позвонки часто разрезали, очищали и перкуссировали. Мышцы на всех ключицах были отпилены, чтобы отсоединить плечо. Один радиус, ATD6-43, был разрезан и очищен. Бедренная кость была раздроблена, вероятно, для извлечения костного мозга. Руки и ноги по-разному подвергаются перкуссиям, порезам или шелушению, вероятно, в результате расчленения. [4]
В общем, в основном были подготовлены более мясные участки, а остальные выброшены. Это говорит о том, что они забивали людей в пищевых целях, но на лице обычно гораздо больше порезов, чем на лицах животных. Когда это наблюдается у доисторических образцов современного человека, это обычно интерпретируется как свидетельство экзоканнибализма , формы ритуального каннибализма, при котором человек ест кого-то, не входящего в его социальную группу, например, врага из соседнего племени. Но, анализируя доказательства каннибализма H. antecessor в 1999 году, испанский палеонтолог Иоланда Фернандес-Халво и ее коллеги вместо этого приписали относительное количество порезов на лице в образце H. antecessor сильно контрастирующей структуре мышечных прикреплений у людей и типичных предметы добычи животных (то есть, чтобы снять кожу с человеческого лица, просто требовалось больше порезов, или мясники были менее знакомы со снятием кожи с людей). [4]
Тем не менее, в скоплении не было пожилых людей, и оно полностью состояло из молодых людей и подростков. В 2010 году Карбонелл выдвинул гипотезу, что они практиковали экзоканнибализм и охотились на представителей соседних племен. [43] В 2019 году испанский палеоантрополог Хесус Родригес и его коллеги утверждали, что, учитывая высокий уровень смертности среди молодежи в современных группах охотников-собирателей, эту демографию лучше объяснить как потребление соплеменников (уже умерших по естественным причинам, войне или несчастному случаю), возможно, просто чтобы не тратить еду зря. [44]
См. также
[ редактировать ]Примечания
[ редактировать ]- ^ Останки Тигеннифа были классифицированы французским палеонтологом позвоночных Камиллой Арамбург как Atlantanthropus mauritanicus в 1955 году. [10] Это название обычно погружается в H. erectus , но многие авторы предпочитают различать азиатские и африканские популяции на видовом уровне, причем последняя классифицируется как H. ergaster ergaster , который был назван в 1975 году , который был назван в 1975 году. Это несколько проблематично, поскольку несколько африканских экземпляров попали в H. . ergaster уже получили уникальные видовые обозначения, которые должны иметь приоритет , включая H. mauritanicus . [11]
- ^ Ширина лобной части (длина лобной кости) и бистефаническая ширина (длина между каждым стефанионом ) ATD6-15 составляют соответственно 95–100 мм (3,7–3,9 дюйма) и 100 мм (3,9 дюйма), что существенно длиннее, чем измерено у H. erectus образцов KNM ER 3733 , KNM ER 3883 , Sangiran 2 или Trinil 2 , каждый из которых имеет предполагаемый объем мозга менее 1000 куб.см (61 куб.дюйм). [2]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д де Кастро, Ж.-МБ (2015). « Homo antecessor : современное состояние восемнадцать лет спустя». Четвертичный интернационал . 433 : 22–31. дои : 10.1016/j.quaint.2015.03.049 .
... событие видообразования могло произойти в Африке/Западной Евразии, в результате чего возникла новая клада Homo [...] Homo antecessor [...] могла быть боковой ветвью этой клады, расположенной в самом западном регионе Евразийского континента.
- ^ Перейти обратно: а б с д и ж г Бермудес де Кастро, JM; Арсуага, JL; Карбонелл, Э.; Росас, А.; Мартинес, И.; Москера, М. (1997). «Гоминоид из нижнего плейстоцена Атапуэрки, Испания: возможный предок неандертальцев и современных людей». Наука . 276 (5317): 1392–1395. дои : 10.1126/science.276.5317.1392 . ПМИД 9162001 . S2CID 31088294 .
- ^ Перейти обратно: а б с д и Кампанья, И.; Перес-Гонсалес А.; Бенито-Кальво, А.; Роселл, Дж.; Бласко, Р.; де Кастро, JMB; Карбонелл, Э.; Арсуага, JL (2016). Гран Долина-TD6 «Новая интерпретация отложений предшественника Homo посредством седиментологического анализа» . Научные отчеты . 6 (34799): 34799. Бибкод : 2016NatSR...634799C . дои : 10.1038/srep34799 . ПМК 5054435 . ПМИД 27713562 .
- ^ Перейти обратно: а б с д и Фернандес-Халво, Ю.; Диес, Дж. К.; Касерес, И.; Роселл, Дж. (1999). «Человеческий каннибализм в раннем плейстоцене Европы (Гран-Долина, Сьерра-де-Атапуэрка, Бургос, Испания)». Журнал эволюции человека . 37 (34): 591–622. дои : 10.1006/jhev.1999.0324 . ПМИД 10497001 . S2CID 25096156 .
- ^ Перейти обратно: а б с д и ж г час Карретеро, Дж. М.; Лоренцо, К.; Арсуага, Дж. Л. (1999). «Осевой и аппендикулярный скелет Homo antecessor ». Журнал эволюции человека . 37 (3–4): 459–499. дои : 10.1006/jhev.1999.0342 . ПМИД 10496997 .
- ^ Перейти обратно: а б с д Карбонелл, Э. (2008). «Первый человекообразный человек Европы». Природа . 452 (7186): 465–469. Бибкод : 2008Natur.452..465C . дои : 10.1038/nature06815 . hdl : 2027.42/62855 . ПМИД 18368116 . S2CID 4401629 .
- ^ де Кастро, JMB; Мартинон-Торрес, М.; Гомес-Роблес, А.; Прадо-Симон, Л.; Мартин-Франсеза, Л.; Лапреса, М.; Олейничака А.; Карбонелл, Э. (2011). «Нижняя челюсть человека раннего плейстоцена из пещеры Сима-дель-Элефанте (TE) в Сьерра-де-Атапуэрка (Испания): сравнительное морфологическое исследование». Журнал эволюции человека . 61 (1): 12–25. дои : 10.1016/j.jhevol.2011.03.005 . ПМИД 21531443 .
- ^ Перейти обратно: а б с Эштон, Н.; Льюис, СГ; Де Гроот, И.; Даффи, С.М.; Бейтс, М.; Бейтс, Р.; Хоар, П.; Льюис, М.; Парфитт, ЮАР; Пеглар, С.; Уильямс, В.; Стрингер, К. (2014). «Следы гомининов из отложений раннего плейстоцена в Хапписбурге, Великобритания» . ПЛОС Один . 9 (2): e88329. Бибкод : 2014PLoSO...988329A . дои : 10.1371/journal.pone.0088329 . ПМЦ 3917592 . ПМИД 24516637 .
- ^ Перейти обратно: а б Стрингер, К. (2012). «Что делает современного человека» . Природа . 485 (7396): 33–35. Бибкод : 2012Natur.485...33S . дои : 10.1038/485033а . ПМИД 22552077 . S2CID 4420496 .
- ^ Арамбург, К. (1955). «Месторождения Атлантантропа » Тернифина и . Бюллетень Французского доисторического общества (на французском языке). 52 (52): 94–95. дои : 10.3406/bspf.1955.3159 . JSTOR 27914994 .
- ^ Энтони, Южная Каролина; Миддлтон, ER (2014). « Homo ergaster ». Энциклопедия глобальной археологии . Спрингер. п. 3462. дои : 10.1007/978-1-4419-0465-2_688 . ISBN 978-1-4419-0426-3 .
- ^ Хублин, Ж.-Ж. (2001). «Гоминиды среднего плейстоцена Северо-Западной Африки и их влияние на появление Homo sapiens ». В Бархеме, Л.; Робсон-Браун, К. (ред.). Человеческие корни: Африка и Азия в среднем плейстоцене (PDF) . Западная академическая и специальная пресса. стр. 116–118. ISBN 978-0-9535418-4-3 .
- ^ де Кастро, Ж.-МБ; Мария-Торрес, М.; Гомес-Роблес, А.; Прадо, Л.; Сармьенто, С. (2007). «Сравнительный анализ нижних челюстей гомининов Гран Долина-TD6 (Испания) и Тигенниф (Алжир)» . Бюллетени и мемуары Парижского общества антропологии . 19 (3–4): 149–167. дои : 10.4000/bmsap.4623 .
- ^ Перейти обратно: а б с Сармьенто, Э.Э.; Моубрей, К.; Сойер, Дж.Дж.; Милнер, Р.; Дик, В.; Таттерсолл, И. (2007). « Гомо предшественник ». Последний человек: Путеводитель по двадцати двум видам вымерших людей . Издательство Йельского университета . стр. 190–191. ISBN 978-0-300-10047-1 .
- ^ Кляйн, Р. (2009). «Расселение гомининов в Старом Свете». В Скарре, К. (ред.). Человеческое прошлое (2-е изд.). Темза и Гудзон. п. 108. ИСБН 978-0-500-29063-7 .
- ^ Перейти обратно: а б Фридлайн, SE; Гунц, П.; Харвати, К.; Хублин, Ж.-Ж. (2013). «Оценка изменений формы в процессе развития Homo antecessor морфологии лица полувзрослых людей ». Журнал эволюции человека . 65 (4): 404–423. дои : 10.1016/j.jhevol.2013.07.012 . ПМИД 23998458 .
- ^ Перейти обратно: а б с д де Кастро, JMB; Мартинон-Торрес, М.; Арсуага, JL ; Карбонелл, Э. (2017). «Двадцатая годовщина Homo antecessor (1997–2017): обзор». Эволюционная антропология . 26 (4): 157–171. дои : 10.1002/evan.21540 . ПМИД 28815959 . S2CID 11442202 .
- ^ Велкер, Фридо; Рамос-Мадригал, Джасмин; Гутенбруннер, Петра; Маки, Миган; Тивари, Шивани; Раковников Джерси-Кристенсен, Роза; Чива, Кристина; Дикинсон, Марк Р.; Кульвильм, Мартин; де Мануэль, Марк; Гелаберт, о.; Мартинон-Торрес, Мария; Маргвелашвили, Анна; Арсуага, Хуан Луис; Карбонелл, Эудальд; Маркес-Боне, Томас; Пенкман, Кирсти; Сабидо, Эдвард; Кокс, Юрген; Олсен, Джеспер В.; Лордкипанидзе, Давид; Расимо, Фернандо; Лалуэса-Фокс, Чарльз; Бермудес де Кастро, Хосе Мария; Виллерслев, Эске; Каппеллини, Энрико (2020). «Дентальный протеом Homo antecessor » . Природа . 580 (7802): 235–238. Бибкод : 2020Natur.580..235W . дои : 10.1038/s41586-020-2153-8 . ISSN 1476-4687 . ПМЦ 7582224 . ПМИД 32269345 . S2CID 214736611 .
- ^ Перейти обратно: а б с д и ж г час я Москера, М.; Олле, А.; Родригес-Альварес, XP; Карбонелл, Э. (2018). «Проливая свет на ранний плейстоцен TD6 (Гран-Долина, Атапуэрка, Испания): технологическая последовательность и профессиональные выводы» . ПЛОС Один . 13 (1): 0190889. Бибкод : 2018PLoSO..1390889M . дои : 10.1371/journal.pone.0190889 . ПМЦ 5784927 . ПМИД 29370188 .
- ^ Перейти обратно: а б с Блейн, Х.-Л.; Куэнка-Бескос, Г.; Бурджакс, Ф.; Лопес-Гарсия, Х.М.; Лосано-Фернандес, И.; Роселл, Дж. (2013). «Палеосреды раннего плейстоцена во времена поселения предшественников Homo в пещере Гран-Долина (Атапуэрка, Испания)». Журнал четвертичной науки . 28 (3): 311–319. Бибкод : 2013JQS....28..311B . дои : 10.1002/jqs.2622 . S2CID 129402115 .
- ^ Фальгер, К.; Бахейн, Дж.; Ёкояма, Ю.; Арсуага, Дж.; Бермудес де Кастро, Дж.; Карбонелл, Э.; Бишофф, Дж.; Доло, Дж. (1999). «Самые ранние люди в Европе: возраст TD6 Гран Долина, Атапуэрка, Испания» (PDF) . Журнал эволюции человека . 37 (3–4): 343–352. дои : 10.1006/jhev.1999.0326 . ПМИД 10496991 . S2CID 41094440 .
- ^ Парес, Ж.М.; Арнольд, Л.; Дюваль, М.; Демуро, М.; Перес-Гонсалес А.; Бермудес де Кастро, JM; Карбонелл, Э.; Арсуагач, JL (2013). «Переоценка возраста Атапуэрки-TD6 (Испания): новые палеомагнитные результаты» (PDF) . Журнал археологической науки . 40 (12): 4586–4595. Бибкод : 2013JArSc..40.4586P . дои : 10.1016/j.jas.2013.06.013 .
- ^ Дюваль, Мэтью; Грюн, Райнер; Парес, Хосеп М.; Мартин-Френч, Лаура; Кампанья, Исидор; Роселл, Хорди; Шао, Цинфэн; Арсуага, Хуан Луис; Карбонелл, Эудальд; Бермудес де Кастро, Хосе Мария (2018). «Первое прямое ЭПР-датирование зуба гоминина из Атапуэрки Гран Долина TD-6 (Испания) подтверждает древность предшественника Homo » . Четвертичная геохронология . 47 : 120–137. Бибкод : 2018QuGeo..47..120D . дои : 10.1016/j.quageo.2018.05.001 . hdl : 10072/381342 .
- ^ Дюваль, М.; Арнольд, LJ; Демуро, М.; Парес, Ж.М.; Кампанья, И.; Карбонелл, Э.; де Кастро, JMB (2022). «Новые хронологические ограничения для самого нижнего стратиграфического подразделения Атапуэрка-Гран-Долина (Бургос, Северная Испания)». Четвертичная геохронология . 71 : 101292. Бибкод : 2022QuGeo..7101292D . дои : 10.1016/j.quageo.2022.101292 . S2CID 248162093 .
- ^ Торо-Мояно, И.; Мартинес-Наварро, Б.; Августин Дж.; Судей, Дж.; Бермудес; де Кастро, Дж. М.; Мартинон-Торрес, М.; Фахардо, Б.; Дюваль, М.; Фальгер, К.; Омс, О.; Парес, Ж.М.; Анадон, П.; Джулия, Р.; Гарсиа-Агилар, Ж.М.; Муань, AM; Спайкс, депутат парламента; Рос-Монтойя, С.; Палмквист, П. (2013). «Самая старая окаменелость человека в Европе из Орсе (Испания)». Журнал эволюции человека . 65 (1): 1–9. дои : 10.1016/j.jhevol.2013.01.012 . hdl : 10261/84112 . ПМИД 23481345 .
- ^ Мояно, IT; Барский, Д. (2011). «Архаичная промышленность каменных орудий из Барранко-Леон и Фуэнте-Нуэва 3 (Орсе, Испания): свидетельства самого раннего присутствия гомининов в южной Европе». Четвертичный интернационал . 243 (1): 80–91. Бибкод : 2011QuInt.243...80M . дои : 10.1016/j.quaint.2010.12.011 .
- ^ Гарсия-Кампос, К.; Мартинен-Торрес, М.; Модесто-Мата, М.; Мартин-Френч, Л.; де Пинильос, ММ; де Кастро, JMB (2021). «Показатели полового диморфизма у постоянных клыков человека-предшественника ». Журнал антропологических наук . 99 (99): 1–18. дои : 10.4436/JASS.99001 . ПМИД 33707343 . S2CID 232206781 .
- ^ Аллен, Дж. С.; Дамасио, Х.; Грабовски, Т.Дж. (2002). «Нормальные нейроанатомические изменения в мозге человека: объемное МРТ-исследование». Американский журнал физической антропологии . 118 (4): 351. дои : 10.1002/ajpa.10092 . ПМИД 12124914 . S2CID 21705705 .
- ^ Перейти обратно: а б Паблос, А.; Лоренцо; Мартинес, К.; де Кастро, JMB; Мартинон-Торрес, М.; Карбонелл, Э.; Арсуага, JL (2012). «Новые останки стопы с памятника раннего плейстоцена Гран Долина-TD6 (Сьерра-де-Атапуэрка, Бургос, Испания)». Журнал эволюции человека . 63 (4): 610–623. дои : 10.1016/j.jhevol.2012.06.008 . ПМИД 22921478 .
- ^ Гарсиа-Мартинес, Даниэль; Грин, Дэвид Дж.; Бермудес де Кастро, Хосе Мария (2021). «Эволюционное развитие лопаток предшественника Homo (местоположение Гран-Долина, Атапуэрка) предполагает современное развитие человека нижнего плейстоцена» . Научные отчеты . 11 (4102): 4102. Бибкод : 2021НатСР..11.4102Г . doi : 10.1038/s41598-021-83039-w . ПМЦ 7892855 . ПМИД 33602966 .
- ^ де Кастро, Ж.-МБ; Мартинон-Торрес, М.; Прадо, Л.; Гомес-Роблес, А.; Роселл, Дж.; Лопес-Полин, Л.; Арсуага, JL; Карбонелл, Э. (2010). «Новая окаменелость неполовозрелого гоминина из европейского нижнего плейстоцена демонстрирует самые ранние свидетельства современной модели развития зубов человека» . Труды Национальной академии наук . 107 (26): 11739–11744. Бибкод : 2010PNAS..10711739B . дои : 10.1073/pnas.1006772107 . ПМК 2900696 . ПМИД 20547843 .
- ^ Мартин-Френч, Л.; Мартинон-Торрес, М.; де Пинильос, ММ; Бэйл, П.; Фернандес-Колон, П.; Гарсия-Кампос, К.; Модесто-Мата, М.; Карбонелл, Э.; Арсуага, JL; де Кастро, JMB (2020). «Эктопический третий моляр верхней челюсти у Homo antecessor раннего плейстоцена из стоянки Атапуэрка-Гран-Долина (Бургос, Испания)». Американский журнал биологической антропологии . 171 (4): 733–741. дои : 10.1002/ajpa.24010 . ПМИД 31943140 . S2CID 210333472 .
- ^ Лопес-Вальверде, А.; Лопес-Кристиа, М.; Гомес де Диего, Р. (2012). «Самая старая челюсть Европы: свидетельства патологии полости рта» . Британский стоматологический журнал . 212 (5): 243–245. дои : 10.1038/sj.bdj.2012.176 . ПМИД 22402544 . S2CID 22346193 .
- ^ Мартин-Френч, Л.; Мартинон-Торрес, М.; Грасия-Теллес, А.; де Кастро, JMB (2016). «Свидетельства травмы надколенника Homo antecessor возрастом около 1 миллиона лет , Гран Долина-Атапуэрка (Испания)» (PDF) . Аккаунты Палевол . 15 (8): 1011–1016. Бибкод : 2016CRPal..15.1011M . дои : 10.1016/j.crpv.2016.04.014 .
- ^ Мартин-Френч, Л.; Мартинон-Торрес, М.; Грасия-Теллес, А.; де Кастро, Ж. М. Б. (2015). «Свидетельства стрессового перелома плюсневой кости предшественника Homo из участка Гран Долина (Атапуэрка, Испания)». Международный журнал остеоархеологии . 25 (4): 564–573. дои : 10.1002/oa.2310 .
- ^ Монсель, М.-Х.; Сантагата, К.; Перейра, А.; Кочевник, С.; Войнчет, П.; Бахейн, Ж.-Ж.; Дожер, К.; Курчи, А.; Леморини, К.; Харди, Б.; Эрамо, Г.; Берто, К.; Рейналь, Ж.-П.; Арзарелло, М.; Мекоцци, Б.; Яннуччи, А.; Сарделла, Р.; Аллегретта, И.; ДеллУниверсита, Э.; Терцано, Р.; Дугас, П.; Жуаник, Г.; Кеффелек, А.; д'Андреа, А.; Валентини, Р.; Минуччи, Э.; Карпентер, Л.; Пиперно, М. (2020). «Происхождение ранней ашельской экспансии в Европе 700 тыс. лет назад: новые находки в Нотарчирико (Италия)» . Научные отчеты . 10 (1): 13802. Бибкод : 2020НатСР..1013802М . дои : 10.1038/s41598-020-68617-8 . ПМЦ 7429832 . ПМИД 32796860 .
- ^ Перейти обратно: а б Робрукс, В.; Вилла, П. (2011). «О самых ранних свидетельствах привычного использования огня в Европе» . Труды Национальной академии наук . 108 (13): 5209–5214. Бибкод : 2011PNAS..108.5209R . дои : 10.1073/pnas.1018116108 . ПМК 3069174 . ПМИД 21402905 .
- ^ Родос, ЮВ; Уокер, MJ; Лопес-Хименес, А.; Лопес-Мартинес, М.; Хабер-Уриарте, М.; Фернандес-Халво, Ю.; Чазан, М. (2016). «Пожар в раннем палеолите: свидетельства обгоревших костей мелких млекопитающих в Черной пещере в проливе реки Кипар, Мурсия, Испания». Журнал археологической науки: отчеты . 9 : 427–436. Бибкод : 2016JArSR...9..427R . дои : 10.1016/j.jasrep.2016.08.006 .
- ^ Перейти обратно: а б с д Саладье, П.; Уге, Р.; Деез, К.; Родригес-Идальго, А.; Касерес, И.; Вальверду, Дж.; Роселл, Дж.; де Кастро, JMB; Карбонелл, Э. (2011). «Решения о транспортировке туш в стратегиях существования Homo Antecessor ». Журнал эволюции человека . 61 (4): 425–446. дои : 10.1016/j.jhevol.2011.05.012 . ПМИД 21802117 . S2CID 36583909 .
- ^ Бласко, Р.; Блейн, Х.-А.; Роселл, Дж.; Диес, Дж. К.; Уге, Р.; Родригес, Дж.; Арсуага, JL; де Кастро, JMB; Карбонелл, Э. (2011). «Самые ранние свидетельства потребления черепах человеком в европейском раннем плейстоцене из Сима-дель-Элефанте, Сьерра-де-Атапуэрка, Испания». Журнал эволюции человека . 61 (4): 503–509. дои : 10.1016/j.jhevol.2011.06.002 . ПМИД 21807397 .
- ^ Аллюэ, Э.; Касерес, И.; Экспозито, И.; Каналс, А.; Родригес, А.; Роселл, Дж.; де Кастро, JMB; Карбонелл, Э. (2015). « Останки Селтиса из нижнего плейстоцена Гран-Долины, Атапуэрка (Бургос, Испания)». Журнал археологической науки . 53 : 570–577. Бибкод : 2015JArSc..53..570A . дои : 10.1016/j.jas.2014.11.016 .
- ^ Перес-Перес, А.; Лозано, М.; Ромеро, А.; Мартинес, LM; Гэлбани, Дж.; Пинилла, Б.; Эстебаранс-Санчес, Ф.; де Кастро, JMB; Карбонелл, Э.; Арсуага, JL (2017). «Питание первых европейцев из Атапуэрки» . Научные отчеты . 7 : 43319. Бибкод : 2017NatSR...743319P . дои : 10.1038/srep43319 . ПМК 5327419 . ПМИД 28240290 .
- ^ Карбонелл, Э .; Касерес, И.; Лозано, М.; Саладье, П.; Роселл, Дж.; Лоренцо, К.; Уге, Р.; Каналс, А.; де Кастро, Ж.-МБ (2010). «Культурный каннибализм как палеоэкономическая система в европейском нижнем плейстоцене». Современная антропология . 51 (4): 539–549. дои : 10.1086/653807 . S2CID 1311044 .
- ^ Родригес, Дж.; Гильермо, З.-Р.; Ана, М. (2019). «Объясняет ли теория оптимального кормодобывания поведение древнейших людей-каннибалов?». Журнал эволюции человека . 131 : 228–239. дои : 10.1016/j.jhevol.2019.03.010 . ПМИД 31182203 . S2CID 149855232 .
Дальнейшее чтение
[ редактировать ]- де Кастро, Ж.-МБ (2002). Мальчик из Гран Долины [ Мальчик Гран Долина ] (на испанском языке). Критика. ISBN 978-84-8432-317-4 .