Ранний современный человек
Ранний современный человек ( ЭМГ ) или анатомически современный человек ( АМГ ), [1] — это термины, используемые для отличия Homo sapiens (единственного сохранившегося вида Hominina ), анатомически соответствующего диапазону фенотипов, наблюдаемых у современных людей, от вымерших архаичных видов человека . Это различие особенно полезно для времен и регионов, где сосуществовали анатомически современные и архаичные люди, например, в палеолитической Европе . Среди старейших известных останков Homo sapiens — те, что были найдены на археологическом объекте Омо-Кибиш I на юго-западе Эфиопии и датируются примерно 233 000 г. [2] до 196 000 лет назад, [3] стоянка Флорисбад в Южной Африке, датируемая примерно 259 000 лет назад, и стоянка Джебель-Ирхуд в Марокко, датируемая примерно 315 000 лет назад.
Вымершие виды рода Homo включают Homo erectus (существовавшие примерно от 2 до 0,1 миллиона лет назад) и ряд других видов (некоторые авторы считают подвидами или H. sapiens H. erectus ). По оценкам, дивергенция линии, ведущей к H. sapiens от предкового H. erectus (или промежуточного вида, такого как Homo antecessor ), произошла в Африке примерно 500 000 лет назад. Самые ранние ископаемые свидетельства существования ранних современных людей появились в Африке около 300 000 лет назад, а самые ранние генетические расколы среди современных людей, по некоторым данным, датируются примерно тем же временем. [4][5][note 1][8] Sustained archaic human admixture with modern humans is known to have taken place both in Africa and (following the recent Out-Of-Africa expansion) in Eurasia, between about 100,000 and 30,000 years ago.[9]
Name and taxonomy
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — |
| |||||||||||||||||||||||||||||
The binomial name Homo sapiens was coined by Linnaeus, 1758.[10] The Latin noun homō (genitive hominis) means "human being", while the participle sapiēns means "discerning, wise, sensible".
The species was initially thought to have emerged from a predecessor within the genus Homo around 300,000 to 200,000 years ago.[note 2] A problem with the morphological classification of "anatomically modern" was that it would not have included certain extant populations. For this reason, a lineage-based (cladistic) definition of H. sapiens has been suggested, in which H. sapiens would by definition refer to the modern human lineage following the split from the Neanderthal lineage. Such a cladistic definition would extend the age of H. sapiens to over 500,000 years.[note 3]
Estimates for the split between the Homo sapiens line and combined Neanderthal/Denisovan line range from between 503,000 and 565,000 years ago;[15] between 550,000 and 765,000 years ago;[16] and (based on rates of dental evolution) possibly more than 800,000 years ago.[17]
Extant human populations have historically been divided into subspecies, but since around the 1980s all extant groups have tended to be subsumed into a single species, H. sapiens, avoiding division into subspecies altogether.[note 4]
Some sources show Neanderthals (H. neanderthalensis) as a subspecies (H. sapiens neanderthalensis).[21][22] Similarly, the discovered specimens of the H. rhodesiensis species have been classified by some as a subspecies (H. sapiens rhodesiensis), although it remains more common to treat these last two as separate species within the genus Homo rather than as subspecies within H. sapiens.[23]
All humans are considered to be a part of the subspecies H. sapiens sapiens,[24] a designation which has been a matter of debate since a species is usually not given a subspecies category unless there is evidence of multiple distinct subspecies.[24]
Age and speciation process
Derivation from H. erectus
The divergence of the lineage that would lead to H. sapiens out of archaic human varieties derived from H. erectus, is estimated as having taken place over 500,000 years ago (marking the split of the H. sapiens lineage from ancestors shared with other known archaic hominins).[8][5] But the oldest split among modern human populations (such as the Khoisan split from other groups) has been recently dated to between 350,000 and 260,000 years ago,[25][26] and the earliest known examples of H. sapiens fossils also date to about that period, including the Jebel Irhoud remains from Morocco (ca. 300,000 or 350–280,000 years ago),[27] the Florisbad Skull from South Africa (ca. 259,000 years ago), and the Omo remains from Ethiopia (ca. 195,000, or, as more recently dated, ca. 233,000 years ago).[28][2]
An mtDNA study in 2019 proposed an origin of modern humans in Botswana (and a Khoisan split) of around 200,000 years.[29] However, this proposal has been widely criticized by scholars,[30][31][32] with the recent evidence overall (genetic, fossil, and archaeological) supporting an origin for H. sapiens approximately 100,000 years earlier and in a broader region of Africa than the study proposes.[32]
In September 2019, scientists proposed that the earliest H. sapiens (and last common human ancestor to modern humans) arose between 350,000 and 260,000 years ago through a merging of populations in East and South Africa.[33][4]
An alternative suggestion defines H. sapiens cladistically as including the lineage of modern humans since the split from the lineage of Neanderthals, roughly 500,000 to 800,000 years ago.
The time of divergence between archaic H. sapiens and ancestors of Neanderthals and Denisovans caused by a genetic bottleneck of the latter was dated at 744,000 years ago, combined with repeated early admixture events and Denisovans diverging from Neanderthals 300 generations after their split from H. sapiens, as calculated by Rogers et al. (2017).[34]
The derivation of a comparatively homogeneous single species of H. sapiens from more diverse varieties of archaic humans (all of which were descended from the early dispersal of H. erectus some 1.8 million years ago) was debated in terms of two competing models during the 1980s: "recent African origin" postulated the emergence of H. sapiens from a single source population in Africa, which expanded and led to the extinction of all other human varieties, while the "multiregional evolution" model postulated the survival of regional forms of archaic humans, gradually converging into the modern human varieties by the mechanism of clinal variation, via genetic drift, gene flow and selection throughout the Pleistocene.[35]
Since the 2000s, the availability of data from archaeogenetics and population genetics has led to the emergence of a much more detailed picture, intermediate between the two competing scenarios outlined above: The recent Out-of-Africa expansion accounts for the predominant part of modern human ancestry, while there were also significant admixture events with regional archaic humans.[36][37]
Since the 1970s, the Omo remains, originally dated to some 195,000 years ago, have often been taken as the conventional cut-off point for the emergence of "anatomically modern humans". Since the 2000s, the discovery of older remains with comparable characteristics, and the discovery of ongoing hybridization between "modern" and "archaic" populations after the time of the Omo remains, have opened up a renewed debate on the age of H. sapiens in journalistic publications.[38][39][40][41][42] H. s. idaltu, dated to 160,000 years ago, has been postulated as an extinct subspecies of H. sapiens in 2003.[43][24] H. neanderthalensis, which became extinct about 40,000 years ago, was also at one point considered to be a subspecies, H. s. neanderthalensis.[24]
H. heidelbergensis, dated 600,000 to 300,000 years ago, has long been thought to be a likely candidate for the last common ancestor of the Neanderthal and modern human lineages. However, genetic evidence from the Sima de los Huesos fossils published in 2016 seems to suggest that H. heidelbergensis in its entirety should be included in the Neanderthal lineage, as "pre-Neanderthal" or "early Neanderthal", while the divergence time between the Neanderthal and modern lineages has been pushed back to before the emergence of H. heidelbergensis, to close to 800,000 years ago, the approximate time of disappearance of H. antecessor.[44][45]
Early Homo sapiens

The term Middle Paleolithic is intended to cover the time between the first emergence of H. sapiens (roughly 300,000 years ago) and the period held by some to mark the emergence of full behavioral modernity (roughly by 50,000 years ago, corresponding to the start of the Upper Paleolithic).
Many of the early modern human finds, like those of Jebel Irhoud, Omo, Herto, Florisbad, Skhul, and Peștera cu Oase exhibit a mix of archaic and modern traits.[46][47][27] Skhul V, for example, has prominent brow ridges and a projecting face. However, the brain case is quite rounded and distinct from that of the Neanderthals and is similar to the brain case of modern humans. It is uncertain whether the robust traits of some of the early modern humans like Skhul V reflects mixed ancestry or retention of older traits.[48][49]
The "gracile" or lightly built skeleton of anatomically modern humans has been connected to a change in behavior, including increased cooperation and "resource transport".[50][51]
There is evidence that the characteristic human brain development, especially the prefrontal cortex, was due to "an exceptional acceleration of metabolome evolution ... paralleled by a drastic reduction in muscle strength. The observed rapid metabolic changes in brain and muscle, together with the unique human cognitive skills and low muscle performance, might reflect parallel mechanisms in human evolution."[52] The Schöningen spears and their correlation of finds are evidence that complex technological skills already existed 300,000 years ago, and are the first obvious proof of an active (big game) hunt. H. heidelbergensis already had intellectual and cognitive skills like anticipatory planning, thinking and acting that so far have only been attributed to modern man.[53][54]
The ongoing admixture events within anatomically modern human populations make it difficult to estimate the age of the matrilinear and patrilinear most recent common ancestors of modern populations (Mitochondrial Eve and Y-chromosomal Adam). Estimates of the age of Y-chromosomal Adam have been pushed back significantly with the discovery of an ancient Y-chromosomal lineage in 2013, to likely beyond 300,000 years ago.[note 5] There have, however, been no reports of the survival of Y-chromosomal or mitochondrial DNA clearly deriving from archaic humans (which would push back the age of the most recent patrilinear or matrilinear ancestor beyond 500,000 years).[56][57][58]
Fossil teeth found at Qesem Cave (Israel) and dated to between 400,000 and 200,000 years ago have been compared to the dental material from the younger (120,000–80,000 years ago) Skhul and Qafzeh hominins.[note 6]
Dispersal and archaic admixture

Dispersal of early H. sapiens begins soon after its emergence, as evidenced by the North African Jebel Irhoud finds (dated to around 315,000 years ago).[27][60] There is indirect evidence for H. sapiens presence in West Asia around 270,000 years ago.[61]
The Florisbad Skull from Florisbad, South Africa, dated to about 259,000 years ago, has also been classified as representing early H. sapiens.[62][63]Scerri (2018), pp. 582–594[4]
In September 2019, scientists proposed that the earliest H. sapiens (and last common human ancestor to modern humans) arose between 350,000 and 260,000 years ago through a merging of populations in East and South Africa.[33][4]
Among extant populations, the Khoi-San (or "Capoid") hunters-gatherers of Southern Africa may represent the human population with the earliest possible divergence within the group Homo sapiens sapiens. Their separation time has been estimated in a 2017 study to be between 350 and 260,000 years ago, compatible with the estimated age of early H. sapiens. The study states that the deep split-time estimation of 350 to 260 thousand years ago is consistent with the archaeological estimate for the onset of the Middle Stone Age across sub-Saharan Africa and coincides with archaic H. sapiens in southern Africa represented by, for example, the Florisbad skull dating to 259 (± 35) thousand years ago.[6]
H. s. idaltu, found at Middle Awash in Ethiopia, lived about 160,000 years ago,[64] and H. sapiens lived at Omo Kibish in Ethiopia about 233,000-195,000 years ago.[65][2] Two fossils from Guomde, Kenya, dated to at least (and likely more than) 180,000 years ago[62] and (more precisely) to 300–270,000 years ago,[4] have been tentatively assigned to H. sapiens and similarities have been noted between them and the Omo Kibbish remains.[62] Fossil evidence for modern human presence in West Asia is ascertained for 177,000 years ago,[66] and disputed fossil evidence suggests expansion as far as East Asia by 120,000 years ago.[67][68]
In July 2019, anthropologists reported the discovery of 210,000 year old remains of a H. sapiens and 170,000 year old remains of a H. neanderthalensis in Apidima Cave, Peloponnese, Greece, more than 150,000 years older than previous H. sapiens finds in Europe.[69][70][71]
A significant dispersal event, within Africa and to West Asia, is associated with the African megadroughts during MIS 5, beginning 130,000 years ago.[72] A 2011 study located the origin of basal population of contemporary human populations at 130,000 years ago, with the Khoi-San representing an "ancestral population cluster" located in southwestern Africa (near the coastal border of Namibia and Angola).[73]

While early modern human expansion in Sub-Saharan Africa before 130 kya persisted, early expansion to North Africa and Asia appears to have mostly disappeared by the end of MIS5 (75,000 years ago), and is known only from fossil evidence and from archaic admixture. Eurasia was re-populated by early modern humans in the so-called "recent out-of-Africa migration" post-dating MIS5, beginning around 70,000–50,000 years ago.[75] In this expansion, bearers of mt-DNA haplogroup L3 left East Africa, likely reaching Arabia via the Bab-el-Mandeb, and in the Great Coastal Migration spread to South Asia, Maritime South Asia and Oceania between 65,000 and 50,000 years ago,[76][77][78][79] while Europe, East and North Asia were reached by about 45,000 years ago. Some evidence suggests that an early wave of humans may have reached the Americas by about 40,000–25,000 years ago.[citation needed]
Evidence for the overwhelming contribution of this "recent" (L3-derived) expansion to all non-African populations was established based on mitochondrial DNA, combined with evidence based on physical anthropology of archaic specimens, during the 1990s and 2000s,[note 7][81] and has also been supported by Y DNA and autosomal DNA.[82] The assumption of complete replacement has been revised in the 2010s with the discovery of admixture events (introgression) of populations of H. sapiens with populations of archaic humans over the period of between roughly 100,000 and 30,000 years ago, both in Eurasia and in Sub-Saharan Africa. Neanderthal admixture, in the range of 1–4%, is found in all modern populations outside of Africa, including in Europeans, Asians, Papua New Guineans, Australian Aboriginals, Native Americans, and other non-Africans.[83][36] This suggests that interbreeding between Neanderthals and anatomically modern humans took place after the recent "out of Africa" migration, likely between 60,000 and 40,000 years ago.[84][85][86] Recent admixture analyses have added to the complexity, finding that Eastern Neanderthals derive up to 2% of their ancestry from anatomically modern humans who left Africa some 100 kya.[87] The extent of Neanderthal admixture (and introgression of genes acquired by admixture) varies significantly between contemporary racial groups, being absent in Africans, intermediate in Europeans and highest in East Asians. Certain genes related to UV-light adaptation introgressed from Neanderthals have been found to have been selected for in East Asians specifically from 45,000 years ago until around 5,000 years ago.[88] The extent of archaic admixture is of the order of about 1% to 4% in Europeans and East Asians, and highest among Melanesians (the last also having Denisova hominin admixture at 4% to 6% in addition to neanderthal admixture).[36][48] Cumulatively, about 20% of the Neanderthal genome is estimated to remain present spread in contemporary populations.[89]
In September 2019, scientists reported the computerized determination, based on 260 CT scans, of a virtual skull shape of the last common human ancestor to modern humans/H. sapiens, representative of the earliest modern humans, and suggested that modern humans arose between 350,000 and 260,000 years ago through a merging of populations in East and South Africa while North-African fossils may represent a population which introgressed into Neandertals during the LMP.[33][4]
Anatomy

Generally, modern humans are more lightly built (or more "gracile") than the more "robust" archaic humans. Nevertheless, contemporary humans exhibit high variability in many physiological traits, and may exhibit remarkable "robustness". There are still a number of physiological details which can be taken as reliably differentiating the physiology of Neanderthals vs. anatomically modern humans.
Anatomical modernity
The term "anatomically modern humans" (AMH) is used with varying scope depending on context, to distinguish "anatomically modern" Homo sapiens from archaic humans such as Neanderthals and Middle and Lower Paleolithic hominins with transitional features intermediate between H. erectus, Neanderthals and early AMH called archaic Homo sapiens.[90] In a convention popular in the 1990s, Neanderthals were classified as a subspecies of H. sapiens, as H. s. neanderthalensis, while AMH (or European early modern humans, EEMH) was taken to refer to "Cro-Magnon" or H. s. sapiens. Under this nomenclature (Neanderthals considered H. sapiens), the term "anatomically modern Homo sapiens" (AMHS) has also been used to refer to EEMH ("Cro-Magnons").[91] It has since become more common to designate Neanderthals as a separate species, H. neanderthalensis, so that AMH in the European context refers to H. sapiens, but the question is by no means resolved.[note 8]
In this more narrow definition of H. sapiens, the subspecies Homo sapiens idaltu, discovered in 2003, also falls under the umbrella of "anatomically modern".[93] The recognition of H. sapiens idaltu as a valid subspecies of the anatomically modern human lineage would justify the description of contemporary humans with the subspecies name Homo sapiens sapiens.[94] However, biological anthropologist Chris Stringer does not consider idaltu distinct enough within H. sapiens to warrant its own subspecies designation.[95][62]
A further division of AMH into "early" or "robust" vs. "post-glacial" or "gracile" subtypes has since been used for convenience. The emergence of "gracile AMH" is taken to reflect a process towards a smaller and more fine-boned skeleton beginning around 50,000–30,000 years ago.[96]
Braincase anatomy

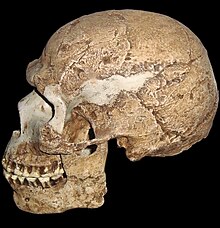
(in Cleveland Museum of Natural History)
Features compared are the braincase shape, forehead, browridge, nasal bone projection, cheek bone angulation, chin and occipital contour
The cranium lacks a pronounced occipital bun in the neck, a bulge that anchored considerable neck muscles in Neanderthals. Modern humans, even the earlier ones, generally have a larger fore-brain than the archaic people, so that the brain sits above rather than behind the eyes. This will usually (though not always) give a higher forehead, and reduced brow ridge. Early modern people and some living people do however have quite pronounced brow ridges, but they differ from those of archaic forms by having both a supraorbital foramen or notch, forming a groove through the ridge above each eye.[97] This splits the ridge into a central part and two distal parts. In current humans, often only the central section of the ridge is preserved (if it is preserved at all). This contrasts with archaic humans, where the brow ridge is pronounced and unbroken.[98]
Modern humans commonly have a steep, even vertical forehead whereas their predecessors had foreheads that sloped strongly backwards.[99] According to Desmond Morris, the vertical forehead in humans plays an important role in human communication through eyebrow movements and forehead skin wrinkling.[100]
Brain size in both Neanderthals and AMH is significantly larger on average (but overlapping in range) than brain size in H. erectus. Neanderthal and AMH brain sizes are in the same range, but there are differences in the relative sizes of individual brain areas, with significantly larger visual systems in Neanderthals than in AMH.[101][note 9]
Jaw anatomy
Compared to archaic people, anatomically modern humans have smaller, differently shaped teeth.[104][105] This results in a smaller, more receded dentary, making the rest of the jaw-line stand out, giving an often quite prominent chin. The central part of the mandible forming the chin carries a triangularly shaped area forming the apex of the chin called the mental trigon, not found in archaic humans.[106] Particularly in living populations, the use of fire and tools requires fewer jaw muscles, giving slender, more gracile jaws. Compared to archaic people, modern humans have smaller, lower faces.
Body skeleton structure
The body skeletons of even the earliest and most robustly built modern humans were less robust than those of Neanderthals (and from what little we know from Denisovans), having essentially modern proportions. Particularly regarding the long bones of the limbs, the distal bones (the radius/ulna and tibia/fibula) are nearly the same size or slightly shorter than the proximal bones (the humerus and femur). In ancient people, particularly Neanderthals, the distal bones were shorter, usually thought to be an adaptation to cold climate.[107] The same adaptation is found in some modern people living in the polar regions.[108]
Height ranges overlap between Neanderthals and AMH, with Neanderthal averages cited as 164 to 168 cm (65 to 66 in) and 152 to 156 cm (60 to 61 in) for males and females, respectively, which is largely identical to pre-industrial average heights for AMH.[note 10] Contemporary national averages range between 158 to 184 cm (62 to 72 in) in males and 147 to 172 cm (58 to 68 in) in females. Neanderthal ranges approximate the contemporary height distribution measured among Malay people, for one.[note 11]
Recent evolution

Following the peopling of Africa some 130,000 years ago, and the recent Out-of-Africa expansion some 70,000 to 50,000 years ago, some sub-populations of H. sapiens had been essentially isolated for tens of thousands of years prior to the early modern Age of Discovery. Combined with archaic admixture this has resulted in significant genetic variation, which in some instances has been shown to be the result of directional selection taking place over the past 15,000 years, i.e., significantly later than possible archaic admixture events.[111]
Some climatic adaptations, such as high-altitude adaptation in humans, are thought to have been acquired by archaic admixture. Introgression of genetic variants acquired by Neanderthal admixture have different distributions in European and East Asians, reflecting differences in recent selective pressures. A 2014 study reported that Neanderthal-derived variants found in East Asian populations showed clustering in functional groups related to immune and haematopoietic pathways, while European populations showed clustering in functional groups related to the lipid catabolic process.[note 12] A 2017 study found correlation of Neanderthal admixture in phenotypic traits in modern European populations.[113]
Physiological or phenotypical changes have been traced to Upper Paleolithic mutations, such as the East Asian variant of the EDAR gene, dated to c. 35,000 years ago.[note 13]
Недавнее расхождение евразийских линий значительно ускорилось во время последнего ледникового максимума (LGM), мезолита и неолита из-за возросшего давления отбора и из-за эффектов основателей , связанных с миграцией . [116] Alleles predictive of light skin have been found in Neanderthals,[117] but the alleles for light skin in Europeans and East Asians, associated with KITLG and ASIP, are (as of 2012[update]), предположительно, были приобретены не в результате архаичной примеси, а в результате недавних мутаций после LGM. [116] Фенотипы, связанные с « белыми » или « кавказскими » популяциями Западной Евразии, возникают во время LGM, примерно 19 000 лет назад. Средняя емкость черепа у современных человеческих популяций колеблется в диапазоне от 1200 до 1450 см. 3 для взрослых мужчин. Больший объем черепа связан с климатическим регионом, наибольшие средние показатели наблюдаются у популяций Сибири и Арктики . [примечание 14] [119] И неандертальцы , и EEMH имели в среднем несколько больший объем черепа, чем современные европейцы, что позволяет предположить ослабление давления отбора в пользу большего объема мозга после окончания LGM. [118]
Примеры еще более поздних адаптаций, связанных с сельским хозяйством и приручением животных , включая восточноазиатские типы ADH1B, связанные с приручением риса , [120] или персистенция лактазы , [121] [122] обусловлены недавним давлением отбора.
Еще более поздняя адаптация была предложена для австронезийских сама-баджау , выведенных под давлением отбора, связанного с существованием за счет фридайвинга на протяжении последней тысячи лет или около того. [123] [124]
Поведенческая современность

поведенческая современность , включающая развитие языка , изобразительного искусства и ранних форм религии Считается, что (и т. д.), возникла до 40 000 лет назад, ознаменовав начало верхнего палеолита (в африканском контексте также известного как поздний каменный век ). [125]
Ведутся серьезные споры относительно того, вели ли самые ранние анатомически современные люди аналогично современным или существующим людям. Поведенческая современность подразумевает полностью развитый язык (требующий способности к абстрактному мышлению ), художественное выражение , ранние формы религиозного поведения , [126] расширение сотрудничества и формирование ранних поселений, а также производство шарнирных орудий из каменных стержней , костей или рога. Термин «верхний палеолит» предназначен для охвата периода с момента быстрого распространения современных людей по Евразии, который совпадает с первым появлением палеолитического искусства, такого как наскальные рисунки , и развитием технологических инноваций, таких как метание копья . Верхний палеолит начинается примерно 50 000–40 000 лет назад и совпадает с исчезновением архаичных людей, таких как неандертальцы .

Термин «поведенческая современность» несколько спорен. Чаще всего его используют для обозначения набора характеристик, обозначающих верхний палеолит, но некоторые ученые используют «поведенческую современность» для обозначения появления H. sapiens около 200 000 лет назад. [127] в то время как другие используют этот термин для обозначения быстрого развития, произошедшего около 50 000 лет назад. [128] [129] [130] Было высказано предположение, что возникновение поведенческой современности было постепенным процессом. [131] [132] [133] [134] [135]
Примеры поведенческой современности

Эквивалент евразийского верхнего палеолита в африканской археологии известен как поздний каменный век , также начавшийся примерно 40 000 лет назад. В то время как наиболее явные доказательства поведенческой современности, обнаруженные в конце 19 века, были получены из Европы, например, фигурки Венеры и другие артефакты ориньякского периода , более поздние археологические исследования показали, что все основные элементы материальной культуры, типичной для современного Сан охотника , Собиратели в Южной Африке также присутствовали, по крайней мере, 40 000 лет назад, включая палки-копатели из аналогичных материалов, используемых сегодня, бусы из скорлупы страусиных яиц , костяные наконечники стрел с индивидуальными отметками производителя, выгравированными и залитыми красной охрой , и аппликаторы яда. [137] Существует также предположение, что «отслаивание под давлением лучше всего объясняет морфологию каменных артефактов, обнаруженных на уровнях среднего каменного века около 75 тыс. лет в пещере Бломбос в Южной Африке. Эта техника использовалась во время окончательного формирования двусторонних точек Стилл-Бэй, сделанных на термообработанный силкрит». [138] Ранее считалось, что как шелушение под давлением, так и термическая обработка материалов произошли гораздо позже в доисторические времена, и оба они указывают на современную поведенческую изощренность в использовании природных материалов. Дальнейшие отчеты об исследованиях пещер вдоль южноафриканского побережья показывают, что «дебаты о том, когда впервые появились культурные и когнитивные характеристики, типичные для современного человека», возможно, подходят к концу, поскольку «передовые технологии со сложными производственными цепочками», которые « часто требуют высокоточной передачи, и поэтому «язык» был обнаружен на участке 5–6 в Южной Африке . Они были датированы примерно 71 000 лет назад. Исследователи предполагают, что их исследование «показывает, что микролитическая технология зародилась в Южной Африке примерно 71 тыс. лет назад, развивалась на протяжении огромного периода времени (около 11 000 лет) и обычно сочеталась со сложной термической обработкой, которая сохранялась почти 100 000 лет. Передовые технологии» в Африке были ранними и долговечными; небольшая выборка раскопанных мест в Африке является лучшим объяснением любого предполагаемого «мерцающего» рисунка». [139] Предполагается, что увеличение сложности поведения связано с более ранним изменением климата на более засушливые условия между 135 000 и 75 000 лет назад. [140] Это могло привести к тому, что группы людей, искавшие убежища от внутренних засух, расселились вдоль прибрежных болот, богатых моллюсками и другими ресурсами. Поскольку уровень моря был низким из-за большого количества воды, связанной ледниками , такие болота могли возникнуть по всему южному побережью Евразии. Использование плотов и лодок вполне могло облегчить исследование прибрежных островов и путешествие вдоль побережья и в конечном итоге позволило распространиться на Новую Гвинею, а затем на Австралию . [141]
Кроме того, в Африке, особенно в Южной, Северной и Восточной Африке, было обнаружено множество других свидетельств абстрактных образов, расширенных стратегий существования и других «современных» моделей поведения, существовавших еще до 50 000 лет назад (некоторые из них появились еще до 100 000 лет назад). Например, пещера Бломбос в Южной Африке известна прямоугольными плитами из охры с выгравированными геометрическими узорами. С помощью нескольких методов датирования было подтверждено, что этому месту около 77 000 и 100 000–75 000 лет. [142] [143] Контейнеры из скорлупы страусиных яиц с выгравированными геометрическими узорами, датируемыми 60 000 лет назад, были найдены в Дипклофе , Южная Африка. [144] В Марокко были найдены бусы и другие личные украшения, возраст которых может достигать 130 000 лет; кроме того, в Пещере очагов в Южной Африке было найдено несколько бус, датируемых значительно раньше 50 000 лет назад. [145] а бусины из ракушек, датируемые примерно 75 000 лет назад, были найдены в пещере Бломбос в Южной Африке. [146] [147] [148] Специализированное метательное оружие также было найдено в различных местах Африки среднего каменного века, в том числе костяные и каменные наконечники стрел в южноафриканских памятниках, таких как пещера Сибуду (вместе с ранней костяной иглой, также найденной в Сибуду), датируемые примерно 72 000–60 000 лет назад. [149] [150] [151] [152] [153] некоторые из них могли быть отравлены ядом, [154] и костяные гарпуны на стоянке Катанда в Центральной Африке, датируемые ок. 90 000 лет назад. [155] Также существуют доказательства систематической термической обработки силикатного камня для повышения его способности к расслаиванию с целью изготовления инструментов, которая началась примерно 164 000 лет назад на южноафриканском участке Пиннакл-Пойнт и стала там обычным явлением для создания микролитических инструментов примерно 72 000 лет назад. назад. [156] [139]
В 2008 году был обнаружен цех по обработке охры, вероятно, для производства красок, датируемый ок. 100 000 лет назад в пещере Бломбос, Южная Африка. Анализ показывает, что в двух раковинах морского ушка образовывалась и хранилась сжиженная смесь, богатая пигментами, и что охра, кость, древесный уголь, точильные камни и молотковые камни также составляли составную часть наборов инструментов. Доказательства сложности задачи включают добычу и объединение сырья из различных источников (подразумевается, что у них был мысленный шаблон процесса, которому они будут следовать), возможное использование пиротехнологии для облегчения извлечения жира из костей, использование вероятного рецепта для производства соединения, и использование контейнеров из ракушек для смешивания и хранения для последующего использования. [157] [158] [159] Современные модели поведения, такие как изготовление бус из ракушек, костяных орудий и стрел, а также использование пигмента охры, очевидны на одном из кенийских памятников 78 000–67 000 лет назад. [160] Свидетельства раннего метательного оружия с каменными наконечниками (характерный инструмент Homo sapiens ), каменные наконечники копий или метательных копий, были обнаружены в 2013 году на эфиопском городище Гадемотта и датируются примерно 279 000 лет назад. [161]
Расширение стратегий существования за пределы охоты на крупную дичь и, как следствие, разнообразие типов инструментов были отмечены как признаки поведенческой современности. Ряд объектов в Южной Африке уже давно начали полагаться на водные ресурсы, от рыбы до моллюсков. Пиннакл-Пойнт , в частности, показывает эксплуатацию морских ресурсов еще 120 000 лет назад, возможно, в ответ на более засушливые условия внутри страны. [162] Например, зависимость от предсказуемых месторождений моллюсков может снизить мобильность и способствовать развитию сложных социальных систем и символического поведения. Пещера Бломбос и Зона 440 в Судане также имеют следы рыбной ловли. Тафономические изменения в скелетах рыб из пещеры Бломбос были интерпретированы как поимка живой рыбы, что явно является преднамеренным поведением человека. [145]
Известно, что люди в Северной Африке ( Назлет-Сабаха , Египет занимались кремня добычей ) еще ≈100 000 лет назад для изготовления каменных орудий . [163] [164]
В 2018 году около 320 000 лет назад на месте Олоргесайли в Кении были обнаружены доказательства раннего возникновения современного поведения, включая: торговлю и транспортировку ресурсов (таких как обсидиан) на большие расстояния, использование пигментов и возможное создание метательных точек. Авторы трех исследований, проведенных на этом сайте в 2018 году, отмечают, что свидетельства такого поведения примерно совпадают с самыми ранними известными ископаемыми останками Homo sapiens из Африки (например, в Джебель-Ирхуде и Флорисбаде), и предполагают, что сложное и современное поведение зародилось в Африке. примерно во времена появления Homo sapiens . [165] [166] [167]
В 2019 году в Адуме, Эфиопия, были обнаружены дополнительные свидетельства существования сложного метательного оружия среднего каменного века в Африке, датированные 100 000–80 000 лет назад, в виде острий, которые, как считается, принадлежали дротикам, доставляемым метателями копья. [168]
Темпы прогресса в Homo sapiens истории
Технологический и культурный прогресс Homo sapiens в последние тысячелетия, по-видимому, был намного быстрее, чем в Homo sapiens ранние периоды . Темпы развития действительно могли ускориться из-за значительного увеличения населения (поэтому больше людей стали думать об инновациях), большего общения и обмена идеями между человеческими популяциями, а также накопления инструментов мышления. Однако может быть также так, что темпы прогресса всегда кажутся людям относительно более высокими в то время, когда они живут, потому что предыдущие достижения не признаются. [169]
Примечания
- ^ На основании Шлебуша и др., «Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад», [6] Рис. 3 ( время дивергенции H. sapiens ) и Стрингер (2012), [7] (архаичная примесь).
- ^ Это вопрос договоренности (а не фактического спора), и универсального консенсуса по терминологии не существует. Некоторые ученые относят к одному и тому же виду людей, живших до 600 000 лет назад. См. Брайант (2003), с. 811. [11] См. также Tattersall (2012), стр. 82 ( ср . К сожалению, этот консенсус в принципе вряд ли многое проясняет на практике. Поскольку нет согласия относительно того, что на самом деле представляют собой «качества человека» [...]). [12]
- ^ Верделин [13] со ссылкой на Либермана и др. [14]
- ^ История заявленных или предполагаемых подвидов H. sapiens сложна и полна противоречий. Единственный широко признанный архаичный подвид. [ нужна ссылка ] — H. sapiens idaltu (2003). Имя Х.с. sapiens происходит от Линнея ( 1758 г. ) и по определению относится к подвиду, типовым экземпляром которого является сам Линней. Однако Линней постулировал четыре других существующих подвида, а именно. Х. с. афер , Х.с. американский , H.s. asiaticus и H. s. Ferus для африканцев, американцев, азиатов и малайцев . Эта классификация оставалась широко распространенной до середины 20 века, иногда наряду с H. s. tasmanianus для австралийцев. См., например, Bailey, 1946; [18] Холл, 1946 год. [19] От разделения современных популяций человека на таксономические подвиды в 1970-е годы постепенно отказались (например, Энциклопедия жизни животных Гржимека). [20] ).
- ^ (95% доверительный интервал 237–581 тысяч лет назад) [55]
- ^ «Хотя ни один из зубов Кесема не демонстрирует набора неандертальских признаков, некоторые черты могут указывать на некоторое сходство с членами эволюционной линии неандертальцев. Однако баланс доказательств предполагает более близкое сходство с зубным материалом Схула / Кафзе, хотя многие из этих сходств, вероятно, представляют собой плезиоморфные черты». [59]
- ^ «Доступные в настоящее время генетические и археологические данные обычно интерпретируются как подтверждающие недавнее единое происхождение современных людей в Восточной Африке». [80]
- ^ Это вопрос общепринятой терминологии, а не фактического разногласия. Паабо (2014) описывает это как дискуссию, которая в принципе неразрешима, «поскольку не существует определения вида, идеально описывающего этот случай». [92]
- ^ Средний объём эндокрана современного человека составляет 1350 см. 3 (82 куб. Дюймов), со значительными различиями между популяциями, средние значения глобальной группы составляют 1085–1580 см. 3 (66,2–96,4 куб. Дюймов). [102] Средний рост неандертальца близок к 1450 см. 3 (88 куб. дюймов) (средний рост мужчин 1600 см) 3 (98 куб. дюймов), средний женский рост 1300 см. 3 (79 куб. Дюймов)), с дальностью действия до 1736 см. 3 (105,9 у.е. дюймов) ( Амуд 1 ). [103]
- ^ «На основе 45 длинных костей максимум от 14 мужчин и 7 женщин, средний рост неандертальцев составляет от 164 до 168 (самцы) или от 152 до 156 см (женщины). Эта высота действительно на 12–14 см ниже, чем высота столба. - Европейцы времен Второй мировой войны, но по сравнению с европейцами примерно 20 000 или 100 лет назад они практически такие же или даже немного выше». [109]
- ^ Малайский, 20–24 (N = м: 749 ж: 893, медиана = м: 166 cm (5 ft 5+1⁄2 in) f:155 cm (5 ft 1 in), SD= m:6.46 cm (2+1⁄2 in) f:6,04 см ( 2 + 1 ⁄ 2 дюйма)) [110]
- ^ «В частности, гены в термине LCP [липидный катаболический процесс] имели наибольший избыток NLS в популяциях европейского происхождения, со средней частотой NLS 20,8 ± 2,6% против 5,9 ± 0,08% по всему геному (двусторонний t-критерий , P<0,0001, n=379 европейцев и n=246 африканцев). Кроме того, среди исследованных популяций людей за пределами Африки избыток NLS [неандертальскоподобных геномных сайтов] в генах LCP наблюдался только у лиц европейского происхождения. : средняя частота NLS у азиатов составляет 6,7±0,7% в генах LCP против 6,2±0,06% по всему геному». [112]
- ^ Признаки, на которые влияет мутация, - это потовые железы, зубы, толщина волос и ткань молочной железы. [114] [115]
- ^ «Мы предлагаем альтернативную гипотезу, которая предполагает, что распространение гоминид в регионы с холодным климатом привело к изменению формы головы. Такое изменение формы способствовало увеличению объема черепа. Биоклиматическое воздействие непосредственно на размер тела (и косвенно на размер мозга) в сочетании с шаровидность черепа, по-видимому, является довольно убедительным объяснением различий между этническими группами». (рисунок в Билсе, стр. 304) [118]
Ссылки
- ^ Нитецкий, Мэтью Х; Нитецкий, Дорис В. (1994). Происхождение анатомически современного человека . Спрингер. ISBN 1489915079 .
- ^ Перейти обратно: а б с Видаль, Селин М.; Лейн, Кристин С .; Асфавроссен, Асрат; и др. (январь 2022 г.). «Возраст древнейшего известного человека разумного из Восточной Африки» . Природа . 601 (7894): 579–583. Бибкод : 2022Природа.601..579В . дои : 10.1038/s41586-021-04275-8 . ПМЦ 8791829 . ПМИД 35022610 .
- ^ Хаммонд, Эшли С.; Ройер, Даниэль Ф.; Флигл, Джон Г. (июль 2017 г.). «Таз Омо-Кибиш I» . Журнал эволюции человека . 108 : 199–219. Бибкод : 2017JHumE.108..199H . дои : 10.1016/j.jhevol.2017.04.004 . ISSN 1095-8606 . ПМИД 28552208 .
- ^ Перейти обратно: а б с д и ж Мунье, Орельен; Лар, Марта (2019). «Расшифровка разнообразия гомининов Африки позднего среднего плейстоцена и происхождения нашего вида» . Природные коммуникации . 10 (1): 3406. Бибкод : 2019NatCo..10.3406M . дои : 10.1038/s41467-019-11213-w . ПМК 6736881 . ПМИД 31506422 .
- ^ Перейти обратно: а б Скерри, Элеонора М.Л.; Томас, Марк Г.; Маника, Андреа; Гунц, Филипп; Сток, Джей Т.; Стрингер, Крис; Гроув, Мэтт; Гроукатт, Хью С.; Тиммерманн, Аксель ; Райтмайр, Дж. Филип; д'Эррико, Франческо (01 августа 2018 г.). «Эволюционировали ли наши виды в разделенных популяциях по всей Африке, и почему это имеет значение?» . Тенденции в экологии и эволюции . 33 (8): 582–594. Бибкод : 2018TEcoE..33..582S . дои : 10.1016/j.tree.2018.05.005 . ISSN 0169-5347 . ПМК 6092560 . ПМИД 30007846 .
- ^ Перейти обратно: а б Шлебуш; и др. (3 ноября 2017 г.). «Древние геномы Южной Африки оценивают дивергенцию современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 .
- ^ Стрингер, К. (2012). «Что делает современного человека» . Природа . 485 (7396): 33–35. Бибкод : 2012Natur.485...33S . дои : 10.1038/485033а . ПМИД 22552077 . S2CID 4420496 .
- ^ Перейти обратно: а б Нойбауэр, Саймон; Юблин, Жан-Жак; Гунц, Филипп (01 января 2018 г.). «Эволюция формы мозга современного человека» . Достижения науки . 4 (1): eaao5961. Бибкод : 2018SciA....4.5961N . дои : 10.1126/sciadv.aao5961 . ISSN 2375-2548 . ПМЦ 5783678 . ПМИД 29376123 .
- ^ Харрод, Джеймс. «Harrod (2014) Таблица дополнительных файлов 1. База данных мифов о языке мтДНК, версия от 17 мая 2019 г.doc» . Родной язык .
- ^ Линней, Карл фон (1758). Система природы. Животный мир (10-е изд.). Правительство Гильельми Энгельмана. стр. 18, 20 Проверено 6 мая 2019 г.
- ^ Брайант, Клифтон Д. (2003). Справочник смерти и умирания . МУДРЕЦ. ISBN 0761925147 .
- ^ Таттерсолл, Ян (2012). Хозяева планеты: В поисках нашего человеческого происхождения . Пресса Святого Мартина. ISBN 978-1137000385 .
- ^ Верделин, Ларс; Сандерс, Уильям Джозеф (2010). Кайнозойские млекопитающие Африки . Университет Калифорнии Пресс. п. 517. ИСБН 978-0520257214 .
- ^ Либерман, Делавэр; МакБрэтни, Б.М.; Кровитц, Г. (2002). «Эволюция и развитие черепной формы у Homo sapiens » . ПНАС . 99 (3): 1134–1139. Бибкод : 2002PNAS...99.1134L . дои : 10.1073/pnas.022440799 . ПМК 122156 . ПМИД 11805284 .
- ^ Хайдиняк, Матея; Фу, Цяомэй; Хюбнер, Александр; Петр, Мартин; и др. (01.03.2018). «Реконструкция генетической истории поздних неандертальцев» . Природа . 555 (7698): 652–656. Бибкод : 2018Natur.555..652H . дои : 10.1038/nature26151 . ISSN 1476-4687 . ПМК 6485383 . ПМИД 29562232 .
- ^ Мейер, Матиас; Арсуага, Хуан-Луис; де Филиппо, Чезаре; Нагель, Сара; и др. (01.03.2016). «Последовательности ядерной ДНК гомининов Sima de los Huesos среднего плейстоцена». Природа . 531 (7595): 504–507. Бибкод : 2016Natur.531..504M . дои : 10.1038/nature17405 . ISSN 1476-4687 . ПМИД 26976447 . S2CID 4467094 .
- ^ Гомес-Роблес, Аида (01 мая 2019 г.). «Темпы эволюции зубов и ее последствия для расхождения неандертальцев и современного человека» . Достижения науки . 5 (5): 1268. Бибкод : 2019SciA....5.1268G . дои : 10.1126/sciadv.aaw1268 . ISSN 2375-2548 . ПМК 6520022 . ПМИД 31106274 .
- ^ Бейли, Джон Венделл (1946). Млекопитающие Вирджинии . п. 356.
- ^ Холл, Э (1946). «Зоологические подвиды человека за столом мира». Журнал маммологии . 27 (4): 358–364. дои : 10.2307/1375342 . JSTOR 1375342 . ПМИД 20247535 .
- ^ Гржимек, Бернхард (1970). Энциклопедия жизни животных Гржимека . Том. 11. Компания Ван Ностранд Рейнхольд. п. 55. ИСБН 978-0442784782 .
- ^ Хаблин, Джей-Джей (2009). «Происхождение неандертальцев» . Труды Национальной академии наук . 106 (38): 16022–16027. Бибкод : 2009PNAS..10616022H . дои : 10.1073/pnas.0904119106 . JSTOR 40485013 . ПМЦ 2752594 . ПМИД 19805257 .
- ^ Харвати, К.; Фрост, СР; МакНалти, КП (2004). «Пересмотр таксономии неандертальцев: значение трехмерных моделей приматов внутри- и межвидовых различий» . Учеб. Натл. акад. наук. США . 101 (5): 1147–1152. Бибкод : 2004PNAS..101.1147H . дои : 10.1073/pnas.0308085100 . ПМК 337021 . ПМИД 14745010 .
- ^ « Король неандертальцев , 1864 год». Энциклопедия эволюции человека Уайли-Блэквелла . Чичестер, Западный Суссекс: Уайли-Блэквелл. 2013. С. 328–331.
- ^ Перейти обратно: а б с д Рафферти, Джон П. «Homo sapiens sapiens» . Британская энциклопедия . Проверено 11 августа 2020 г.
- ^ Шлебуш, Карина М.; Мальмстрем, Хелена; Гюнтер, Торстен; Сьёдин, Пер; Коутиньо, Александра; Эдлунд, Ханна; Мунтерс, Ариэль Р.; Стейн, Марина; Содиалл, Небеса; Ломбард, Марлиз; Якобссон, Маттиас (5 июня 2017 г.). «Древние геномы из южной Африки выдвигают дивергенцию современного человека за пределы 260 000 лет назад». bioRxiv 10.1101/145409 .
- ^ Шлебуш, Карина М.; Мальмстрем, Хелена; Гюнтер, Торстен; Сьёдин, Пер; Коутиньо, Александра; Эдлунд, Ханна; Мунтерс, Ариэль Р.; Висенте, Марио; Стейн, Марина; Судьял, Химла; Ломбард, Марлиз (3 ноября 2017 г.). «Древние геномы Южной Африки оценивают дивергенцию современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ISSN 0036-8075 . ПМИД 28971970 .
- ^ Перейти обратно: а б с Каллауэй, Юэн (7 июня 2017 г.). «Древнейшие ископаемые останки Homo sapiens переписывают историю нашего вида » Природа . дои : 10.1038/nature.2017.22114 . Проверено 11 июня 2017 г.
- ^ Стрингер (2016) , с. 20150237; Образец (2017 г.) ; Хаблин и др. (2017) , стр. 289–292; Шерри (2018) , стр. 582–594
- ^ Чан, Ева, К.Ф. (28 октября 2019 г.). «Происхождение человека в палео-болотных угодьях южной Африки и первые миграции». Природа . 857 (7781): 185–189. Бибкод : 2019Natur.575..185C . дои : 10.1038/s41586-019-1714-1 . ПМИД 31659339 . S2CID 204946938 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Сэмпл, Ян (28 октября 2019 г.). «Исследование показало, что прародина современных людей находится в Ботсване» . Хранитель . ISSN 0261-3077 . Проверено 29 октября 2019 г.
- ^ Вудворд, Эйлин (28 октября 2019 г.). «Новое исследование указывает на прародину всех людей, живущих сегодня» . ScienceAlert.com . Проверено 29 октября 2019 г.
- ^ Перейти обратно: а б Йонг, Эд (28 октября 2019 г.). «Найдена ли родина человечества?» . Атлантика . Проверено 28 октября 2019 г.
- ^ Перейти обратно: а б с Циммер, Карл (10 сентября 2019 г.). «Ученые нашли череп предка человечества – на компьютере – Сравнивая окаменелости и компьютерную томографию, исследователи говорят, что реконструировали череп последнего общего предка современных людей» . Нью-Йорк Таймс . Проверено 10 сентября 2019 г.
- ^ Роджерс, Алан Р.; Болендер, Райан Дж.; Хафф, Чад Д. (12 сентября 2017 г.). «Ранняя история неандертальцев и денисовцев» . Труды Национальной академии наук . 114 (37): 9859–9863. Бибкод : 2017PNAS..114.9859R . дои : 10.1073/pnas.1706426114 . ПМК 5604018 . ПМИД 28784789 .
- ^ Вулпофф, Миннесота; Шпулер, Дж. Н.; Смит, Ф.Х.; Радовчич Дж.; Поуп, Г.; Фрайер, Д.В.; Экхардт, Р.; Кларк, Г. (1988). «Происхождение современного человека». Наука . 241 (4867): 772–774. Бибкод : 1988Sci...241..772W . дои : 10.1126/science.3136545 . ПМИД 3136545 . S2CID 5223638 .
- ^ Перейти обратно: а б с Грин Р.Э., Краузе Дж., Бриггс А.В., Маричич Т., Стензель У., Кирхер М., Паттерсон Н., Ли Х., Чжай В., Фриц М.Х., Хансен Н.Ф., Дюран Э.Ю., Маласпинас А.С., Йенсен Дж.Д., Маркес-Боне Т., Алкан С., Прюфер К., Мейер М., Бурбано Х.А., Гуд Дж.М., Шульц Р., Аксиму-Петри А., Баттхоф А., Хёбер Б., Хёффнер Б., Зигемунд М., Вейманн А., Нусбаум С., Ландер Э.С. и др. (май 2010 г.). «Проект последовательности генома неандертальца» . Наука . 328 (5979): 710–722. Бибкод : 2010Sci...328..710G . дои : 10.1126/science.1188021 . ПМК 5100745 . ПМИД 20448178 .
- ^ Райх Д., Паттерсон Н., Кирхер М., Дельфин Ф., Нандинени М.Р., Пугач И., Ко А.М., Ко Ю.К., Джинам Т.А., Фиппс М.Е., Сайту Н., Вольштейн А., Кайзер М., Паабо С., Стоункинг М. (2011). «Денисова примесь и первые расселения современного человека в Юго-Восточную Азию и Океанию» . Ам Джей Хум Жене . 89 (4): 516–528. дои : 10.1016/j.ajhg.2011.09.005 . ПМК 3188841 . ПМИД 21944045 .
- ^ «Новые подсказки добавляют 40 000 лет к возрасту человеческого вида» . www.nsf.gov . ННФ – Национальный научный фонд.
- ^ «Переоценка возраста древнего человека» . Новости Би-би-си . 16 февраля 2005 года . Проверено 10 апреля 2010 г.
- ^ «Самый древний Homo Sapiens: окаменелости отодвигают появление человека на 195 000 лет назад» . ScienceDaily . 28 февраля 2005 года . Проверено 6 мая 2019 г.
- ^ Алемсегед, З.; Коппенс, Ю.; Гераадс, Д. (2002). «Череп гоминида Homo: Описание и систематика Homo-323-1976-896» (PDF) . Am J Phys Anthropol . 117 (2): 103–112. дои : 10.1002/ajpa.10032 . ПМИД 11815945 . Архивировано (PDF) из оригинала 18 июля 2020 г.
- ^ Стоункинг, Марк; Судьял, Химла (1996). «Эволюция человека и митохондриальный геном». Текущее мнение в области генетики и развития . 6 (6): 731–736. дои : 10.1016/S0959-437X(96)80028-1 . ПМИД 8994844 .
- ^ Эволюция человека: ископаемые свидетельства в 3D , Филип Л. Уокер и Эдвард Х. Хаген, кафедра антропологии Калифорнийского университета, Санта-Барбара. Проверено 5 апреля 2005 г.
- ^ Мейер, Матиас; Арсуага, Хуан-Луис; де Филиппо, Чезаре; Нагель, Сара; Ачиму-Петри, Айнуэр; Никель, Биргит; Мартинес, Игнасио; Грейс, Ана; де Кастро, Хосе Мария Бермудес; Карбонелл, Эудальд; Виола, Бенс; Келсо, Джанет; Прюфер, Кей; Пяабо, Сванте (14 марта 2016 г.). «Последовательности ядерной ДНК гомининов Sima de los Huesos среднего плейстоцена». Природа . 531 (7595): 504–507. Бибкод : 2016Природа.531..504М . дои : 10.1038/nature17405 . ПМИД 26976447 . S2CID 4467094 .
- ^ Каллауэй, Юэн (14 марта 2016 г.). «ДНК древнейшего человека детализирует рассвет неандертальцев» . Природа . 531 (7594): 296–286. Бибкод : 2016Natur.531..296C . дои : 10.1038/531286a . ПМИД 26983523 . S2CID 4459329 .
- ^ Оппенгеймер, С. (2003). Из Эдема: Заселение мира . Робинсон. ISBN 978-1841196978 .
- ^ Тринкаус, Э.; Молдован, О.; Милота, Ш.; Билгар, А.; Сарцина, Л.; Атрея, С.; Бейли, ЮВ; Родриго, Р.; и др. (2003). «Ранний современный человек из Пештера-ку-Оасе, Румыния» . ПНАС . 100 (20): 11231–11236. Бибкод : 2003PNAS..10011231T . дои : 10.1073/pnas.2035108100 . ПМК 208740 . ПМИД 14504393 .
- ^ Перейти обратно: а б Райх, Дэвид; Грин, Ричард Э.; Кирхер, Мартин; Краузе, Йоханнес; Паттерсон, Ник; Дюран, Эрик Ю.; Виола, Бенс; Бриггс, Адриан В.; и др. (2010). «Генетическая история архаической группы гомининов из Денисовой пещеры в Сибири» . Природа . 468 (7327): 1053–1060. Бибкод : 2010Natur.468.1053R . дои : 10.1038/nature09710 . hdl : 10230/25596 . ПМК 4306417 . ПМИД 21179161 .
- ^ Тринкаус, Эрик (октябрь 2005 г.). «Ранние современные люди». Ежегодный обзор антропологии . 34 (1): 207–230. дои : 10.1146/annurev.anthro.34.030905.154913 . S2CID 9039428 .
- ^ Мелдрам, Джефф; Хилтон, Чарльз Э. (2004). От двуногого к страйдеру: появление у современного человека ходьбы, бега и транспорта ресурсов . Springer Science & Business Media. ISBN 978-0306480003 .
- ^ Вонк, Дженнифер; Шекелфорд, Тодд К. (2012). Оксфордский справочник по сравнительной эволюционной психологии . Издательство Оксфордского университета, США. стр. 429–. ISBN 978-0199738182 .
- ^ Божек, Катажина; Вэй, Юнин; Ян, Чжэн; Лю, Силин; Сюн, Цзеи; Сугимото, Масахиро; Томита, Масару; Паабо, Сванте; Пешек, Райк; Шервуд, Чет К.; Хоф, Патрик Р.; Эли, Джон Дж.; Штайнхаузер, Дирк; Вильмицер, Лотар; Бангсбо, Йенс; Ханссон, Ола; Позвони, Жозеп; Джавалиско, Патрик; Хаитович, Филипп (2014). «Исключительное эволюционное расхождение метаболомов мышц и мозга человека соответствует когнитивной и физической уникальности человека» . ПЛОС Биология . 12 (5): e1001871. дои : 10.1371/journal.pbio.1001871 . ПМК 4035273 . ПМИД 24866127 .
- ^ Тиме, Х (2007). «Большой успех Шёнингена: новая картина культуры древних людей». Шёнингенские копья – люди и охота 400 000 лет назад . Конрад Тайсс Верлаг. стр. 224–328. ISBN 978-3896460400 .
- ^ Хейдл, Миннесота (2006). «Обезьяны? Обезьяны? Люди! Познание и язык в верхнем палеолите». В Конарде, штат Нью-Джерси (ред.). Откуда берется человек ? Издательство Попытка. стр. 69–97. ISBN 3893083812 .
- ^ Мендес, Фернандо; Кран, Томас; Шрак, Бонни; Кран, Астрид-Мария; Вирама, Кришна; Вернер, август; Фомине, Форка Лейпей Мэтью; Брэдман, Нил; Томас, Марк (7 марта 2013 г.). «Афроамериканская отцовская линия добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» (PDF) . Американский журнал генетики человека . 92 (3): 454–459. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 . Архивировано из оригинала (PDF) 24 сентября 2019 года . Проверено 21 апреля 2018 г.
- ^ Крингс М., Стоун А., Шмитц Р.В., Крайницки Х., Стоункинг М., Паабо С. (июль 1997 г.). «Последовательности ДНК неандертальцев и происхождение современного человека». Клетка . 90 (1): 19–30. дои : 10.1016/S0092-8674(00)80310-4 . hdl : 11858/00-001M-0000-0025-0960-8 . ПМИД 9230299 . S2CID 13581775 .
- ^ Хилл, Дебора (16 марта 2004 г.). «В генофонде нет неандертальцев» . Наука . Проверено 6 мая 2019 г.
- ^ Серр, Д.; Лангани, А.; Чех, М.; Тешлер-Никола, М .; Паунович, М.; Меннесьер, П.; Хофрейтер, М.; Посснерт, Г.; Паабо, С. (2004). «Нет доказательств вклада неандертальской мтДНК в развитие ранних современных людей» . ПЛОС Биология . 2 (3): 313–317. doi : 10.1371/journal.pbio.0020057 . ПМК 368159 . ПМИД 15024415 .
- ^ Гершковиц, Я; Смит, П; Сариг, Р; Куам, Р; Родригес, Л; Гарсия, Р; Арсуага, JL; Баркай, Р; Суслик, А (2011). «Останки зубов среднего плейстоцена из пещеры Кесем (Израиль)». Американский журнал физической антропологии . 144 (4): 575–592. дои : 10.1002/ajpa.21446 . ПМИД 21404234 . S2CID 3106938 .
- ^ Хаблин и др. 2017 , стр. 289–292.
- ^ Пост, Козимо; и др. (4 июля 2017 г.). «Глубоко расходящийся архаичный митохондриальный геном обеспечивает нижнюю временную границу для потока африканских генов в неандертальцев» . Природные коммуникации . 8 : 16046. Бибкод : 2017NatCo...816046P . дои : 10.1038/ncomms16046 . ПМК 5500885 . ПМИД 28675384 .
- ^ Перейти обратно: а б с д Стрингер 2016 , с. 20150237.
- ^ Образец 2017 года .
- ^ Уайт, Тим Д.; Асфау, Берхане; Дегуста, Дэвид; Гилберт, Генри; Ричардс, Гэри Д.; Сува, генерал; Хауэлл, Кларк Ф. (июнь 2003 г.). «Плейстоценовый Homo sapiens из Среднего Аваша, Эфиопия». Природа . 423 (6941): 742–747. Бибкод : 2003Natur.423..742W . дои : 10.1038/nature01669 . ПМИД 12802332 . S2CID 4432091 .
- ^ «Повторный анализ окаменелостей отодвигает назад происхождение Homo sapiens » . Научный американец . 17 февраля 2005 г. Проверено 6 мая 2019 г.
- ^ Мехта, Анкита (26 января 2018 г.). «Окаменелость челюстной кости возрастом 177 000 лет, обнаруженная в Израиле, является старейшим человеческим останком, найденным за пределами Африки» . Интернэшнл Бизнес Таймс . Проверено 6 мая 2019 г.
- ^ Бэ, Кристофер Дж.; Дука, Катерина; Петралья, Майкл Д. (8 декабря 2017 г.). «О происхождении современного человека: азиатские взгляды» . Наука . 358 (6368): eaai9067. дои : 10.1126/science.aai9067 . ПМИД 29217544 .
- ^ Куо, Лили (10 декабря 2017 г.). «Ранние люди мигрировали из Африки гораздо раньше, чем мы думали» . Кварц . Проверено 6 мая 2019 г.
- ^ Циммер, Карл (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить историю предыстории человечества. Кость, найденная в пещере, является старейшим ископаемым современного человека, когда-либо обнаруженным в Европе. Она намекает на то, что люди начали покидать Африку гораздо раньше, чем когда-то считалось» . Нью-Йорк Таймс . Проверено 11 июля 2019 г.
- ^ Персонал (10 июля 2019 г.). « Самые старые останки за пределами Африки обнуляют часы миграции людей» . Физика.орг . Проверено 10 июля 2019 г.
- ^ Харвати, Катерина; и др. (10 июля 2019 г.). «Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии» . Природа . 571 (7766): 500–504. дои : 10.1038/s41586-019-1376-z . ПМИД 31292546 . S2CID 195873640 .
- ^ Рито, Т.; Ричардс, МБ; Фернандес, В.; Альшамали, Ф.; Черный, В.; Перейра, Л.; Соареш, П. (2013). «Первое расселение современного человека по Африке» . ПЛОС ОДИН . 8 (11): е80031. Бибкод : 2013PLoSO...880031R . дои : 10.1371/journal.pone.0080031 . ПМЦ 3827445 . ПМИД 24236171 .
- ^ Хенн, Бренна; Жиньу, Кристофер Р.; Джобин, Мэтью (2011). «Геномное разнообразие охотников-собирателей предполагает южноафриканское происхождение современного человека» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (13): 5154–5162. Бибкод : 2011PNAS..108.5154H . дои : 10.1073/pnas.1017511108 . ПМК 3069156 . ПМИД 21383195 .
- ^ Перейти обратно: а б с Хайэм, Томас Ф.Г.; Весселинг, Фрэнк П.; Хеджес, Роберт Э.М.; Бергман, Кристофер А.; Дука, Катерина (11 сентября 2013 г.). «Хронология Ксара Акила (Ливан) и последствия колонизации Европы анатомически современными людьми» . ПЛОС ОДИН . 8 (9): е72931. Бибкод : 2013PLoSO...872931D . дои : 10.1371/journal.pone.0072931 . ISSN 1932-6203 . ПМК 3770606 . ПМИД 24039825 .
- ^ Пост и др., 2016 ; Камин и др., 2015 ; Вай и др., 2019 ; Хабер и др., 2019 г.
- ^ Кларксон, Крис; Джейкобс, Зенобия; Пардо, Колин (2017). «Человеческая оккупация северной Австралии 65 000 лет назад» (PDF) . Природа . 547 (7663): 306–310. Бибкод : 2017Natur.547..306C . дои : 10.1038/nature22968 . hdl : 2440/107043 . ПМИД 28726833 . S2CID 205257212 . Архивировано (PDF) из оригинала 28 апреля 2019 г.
- ^ Сен-Флё, Николас (19 июля 2017 г.). «Люди впервые прибыли в Австралию 65 000 лет назад, как показывают исследования» . Нью-Йорк Таймс .
- ^ Вуд Р. (2 сентября 2017 г.). «Комментарии к хронологии Маджедбебе». Австралийская археология . 83 (3): 172–174. дои : 10.1080/03122417.2017.1408545 . ISSN 0312-2417 . S2CID 148777016 .
- ^ О'Коннелл Дж. Ф., Аллен Дж., Уильямс М.А., Уильямс А.Н., Терни К.С., Спунер Н.А. и др. (август 2018 г.). «Homo sapiens впервые достигает Юго-Восточной Азии и Сахула?» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (34): 8482–8490. дои : 10.1073/pnas.1808385115 . ПМК 6112744 . ПМИД 30082377 .
- ^ Лю, Хуа; и др. (2006). «Географически явная генетическая модель всемирной истории поселений» . Американский журнал генетики человека . 79 (2): 230–237. дои : 10.1086/505436 . ПМК 1559480 . ПМИД 16826514 .
- ^ «Возвращение из Африки». Наука . 308 (5724): 921г. 13 мая 2005 г. дои : 10.1126/science.308.5724.921g . S2CID 220100436 .
- ^ Хабер и др., 2019 .
- ^ Шанкарараман, Шрирам; Маллик, Свапан; Паттерсон, Ник; Райх, Дэвид (2016). «Объединенный ландшафт денисовского и неандертальского происхождения у современного человека» . Современная биология . 26 (9): 1241–1247. Бибкод : 2016CBio...26.1241S . дои : 10.1016/j.cub.2016.03.037 . ISSN 0960-9822 . ПМЦ 4864120 . ПМИД 27032491 .
- ^ Санчес-Куинто, форвард; Ботиге, LR; Сивит, С; Аренас, К; Авила-Аркос, MC; Бустаманте, CD; Комас, Д; Лалуэса-Фокс, C (17 октября 2012 г.). «Население Северной Африки несет в себе признак смешения с неандертальцами» . ПЛОС ОДИН . 7 (10): е47765. Бибкод : 2012PLoSO...747765S . дои : 10.1371/journal.pone.0047765 . ПМЦ 3474783 . ПМИД 23082212 .
- ^ Фу, Кью; Ли, Х; Мурджани, П; Джей, Ф; Слепченко С.М.; Бондарев А.А.; Джонсон, Польша; Аксимо-Петри, А; Прюфер, К; де Филиппо, К; Мейер, М; Цвинс, Н.; Саласар-Гарсия, округ Колумбия; Кузьмин Ю.В.; Китс, СГ; Косинцев П.А.; Ражев Д.И.; Ричардс, член парламента; Перистов, Н.В.; Лахманн, М; Дука, К; Хайэм, штат Техас; Слаткин М; Хаблин, Джей-Джей; Райх, Д; Келсо, Дж; Виола, туберкулез; Пяабо, С. (23 октября 2014 г.). «Последовательность генома современного человека возрастом 45 000 лет из Западной Сибири» . Природа . 514 (7523): 445–449. Бибкод : 2014Nature.514..445F . дои : 10.1038/nature13810 . ПМЦ 4753769 . ПМИД 25341783 .
- ^ Брагич, Екатерина (3 февраля 2014 г.). «Забытое возвращение человечества в Африку раскрыто в ДНК» . Новый учёный . Проверено 6 мая 2019 г.
- ^ Кульвильм, Мартин (17 февраля 2016 г.). «Древний поток генов от ранних современных людей к восточным неандертальцам» . Природа . 530 (7591): 429–433. Бибкод : 2016Natur.530..429K . дои : 10.1038/nature16544 . ПМЦ 4933530 . ПМИД 26886800 .
- ^ Дин, К.; Ху, Ю.; Сюй, С.; Ван, Дж.; Джин, Л. (2014) [Онлайн, 2013]. «Неандертальская интрогрессия на хромосоме 3p21.31 находилась под действием положительного естественного отбора у жителей Восточной Азии» . Молекулярная биология и эволюция . 31 (3): 683–695. дои : 10.1093/molbev/mst260 . ПМИД 24336922 .
- ^ Верно, Б.; Эйки, Дж. М. (2014). «Воскрешение сохранившихся линий неандертальцев из геномов современного человека» . Наука . 343 (6174): 1017–1021. Бибкод : 2014Sci...343.1017V . дои : 10.1126/science.1245938 . ПМИД 24476670 . S2CID 23003860 .
- ^ Айяла, Франсиско Хосе; Конде, Камило Хосе Села (2017). Процессы эволюции человека: путешествие от ранних гомининов к неандертальцам и современным людям . Издательство Оксфордского университета. ISBN 978-0198739906 – через Google Книги .
- ^ Шопф, Дж. Уильям (1992). Основные события в истории жизни . Джонс и Бартлетт Обучение. стр. 168–. ISBN 978-0867202687 – через Google Книги .
- ^ Паабо, Сванте (2014). Неандерталец: в поисках утраченных геномов . Нью-Йорк: Основные книги . п. 237.
- ^ Сандерс, Роберт (11 июня 2003 г.). «Окаменелые черепа возрастом 160 000 лет, обнаруженные в Эфиопии, являются древнейшими анатомически современными людьми» . Новости Калифорнийского университета в Беркли . Проверено 7 мая 2019 г.
- ^ Уайт, Тим Д .; Асфау, Б.; ДеГуста, Д.; Гилберт, Х.; Ричардс, Джорджия; Сува, Г.; Хауэлл, ФК (2003). «Плейстоценовый Homo sapiens из Среднего Аваша, Эфиопия». Природа . 423 (6491): 742–747. Бибкод : 2003Natur.423..742W . дои : 10.1038/nature01669 . ПМИД 12802332 . S2CID 4432091 .
- ^ Стрингер, Крис (12 июня 2003 г.). «Эволюция человека: Из Эфиопии». Природа . 423 (6941): 693–695. Бибкод : 2003Natur.423..692S . дои : 10.1038/423692a . ПМИД 12802315 . S2CID 26693109 .
- ^ Хоукс, Дж.; Ван, ET; Кокран, генеральный директор; Харпендинг, ХК; Мойзис, РК (2007). «Недавнее ускорение адаптивной эволюции человека» . Труды Национальной академии наук . 104 (52): 20753–20758. Бибкод : 2007PNAS..10420753H . дои : 10.1073/pnas.0707650104 . ПМК 2410101 . ПМИД 18087044 .
- ^ Бхупендра, П. (апрель 2019 г.). «Анатомия лба» . Ссылки на Medscape . Проверено 6 мая 2019 г.
- ^ «Как идентифицировать современного человека?» . Новости, 2012 . Музей естественной истории, Лондон . Проверено 11 декабря 2013 г.
- ^ Энкарта, Эволюция человека . Архивировано из оригинала 29 октября 2009 года.
{{cite encyclopedia}}:|work=игнорируется ( помогите ) - ^ Моррис, Десмонд (2007). «Бровь» . Обнаженная женщина: исследование женского тела . Макмиллан. ISBN 978-0312338534 .
- ^ Пирс, Эйлунед; Стрингер, Крис; Данбар, RIM (07 мая 2013 г.). «Новое понимание различий в организации мозга между неандертальцами и анатомически современными людьми» . Труды Лондонского королевского общества B: Биологические науки . 280 (1758): 20130168. doi : 10.1098/rspb.2013.0168 . ISSN 0962-8452 . ПМЦ 3619466 . ПМИД 23486442 .
- ^ Смит, CL; Билс, КЛ (1990). «Культурные коррелирует с емкостью черепа». Американский антрополог . 92 : 193–200. дои : 10.1525/aa.1990.92.1.02a00150 . S2CID 162406199 .
- ^ Стрингер, К. (1984). «Эволюция человека и биологическая адаптация в плейстоцене». В Фоли, Р. (ред.). Эволюция гоминидов и экология сообщества . Нью-Йорк: Академическая пресса. ISBN 978-0122619205 .
- ^ Таунсенд Г., Ричардс Л., Хьюз Т. (май 2003 г.). «Молярные межбугорковые размеры: генетический вклад в фенотипические вариации». Журнал стоматологических исследований . 82 (5): 350–355. дои : 10.1177/154405910308200505 . ПМИД 12709500 . S2CID 26123427 .
- ^ Кейт А. (1913). «Проблемы, связанные с зубами ранних форм доисторического человека» . Труды Королевского медицинского общества . 6 (Секта Одонтол): 103–124. дои : 10.1177/003591571300601018 . ПМК 2005996 . ПМИД 19977113 .
- ^ Таттерсолл, Джеффри Х; Шварц, Ян (2003). Летопись окаменелостей человека Краниодентальная морфология рода Homo (Африка и Азия) (том 2) . Вили-Лисс. стр. 327–328. ISBN 978-0471319283 .
- ^ Стегманн, А. Теодор; Черни, Фрэнк Дж.; Холлидей, Трентон В. (2002). «Холодовая адаптация неандертальцев: физиологические и энергетические факторы». Американский журнал биологии человека . 14 (5): 566–583. дои : 10.1002/ajhb.10070 . ПМИД 12203812 . S2CID 2437566 .
- ^ Сток, JT (октябрь 2006 г.). «Посткраниальная устойчивость охотников-собирателей относительно моделей мобильности, климатической адаптации и отбора для экономии тканей». Американский журнал физической антропологии . 131 (2): 194–204. дои : 10.1002/ajpa.20398 . ПМИД 16596600 .
- ^ Хельмут Х (1998). «Рост тела, масса тела и площадь поверхности неандертальцев». Журнал морфологии и антропологии . 82 (1): 1–12. doi : 10.1127/zma/82/1998/1 (неактивен 26 апреля 2024 г.). ПМИД 9850627 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Лим Т.О., Дин Л.М., Заки М. и др. (март 2000 г.). «Распределение массы тела, роста и индекса массы тела в национальной выборке взрослых малайзийцев» (PDF) . Мед. Дж. Малайзия . 55 (1): 108–128. ПМИД 11072496 . Архивировано (PDF) из оригинала 24 февраля 2017 г.
- ^ Уэйд, Н. (7 марта 2006 г.). «Человеческие гены все еще развиваются, рассказывая новую историю» . Нью-Йорк Таймс . Проверено 10 июля 2008 г.
- ^ Храмеева Е; Бозек, К; Он, Л; Ян, З; Цзян, X; Вэй, Ю; Тан, К; Гельфанд, М.С.; Прюфер, К; Келсо, Дж; Паабо, С; Джавалиско, П; Лахманн, М; Хаитович, П (2014). «Неандертальское происхождение стимулирует эволюцию катаболизма липидов у современных европейцев» . Природные коммуникации . 5 (3584): 3584. Бибкод : 2014NatCo...5.3584K . дои : 10.1038/ncomms4584 . ПМЦ 3988804 . ПМИД 24690587 .
- ^ Майкл Даннеманн 1 и Джанет Келсо, «Вклад неандертальцев в фенотипические вариации современных людей», Американский журнал генетики человека 101 , 578–589, 5 октября 2017 г.
- ^ Камберов, Яна Г (14 февраля 2013 г.). «Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR» . Клетка . 152 (4): 691–702. дои : 10.1016/j.cell.2013.01.016 . ПМК 3575602 . ПМИД 23415220 .
- ^ Уэйд, Николас (14 февраля 2013 г.). «Восточноазиатские физические черты, связанные с 35 000-летней мутацией» . Нью-Йорк Таймс . Проверено 6 мая 2019 г.
- ^ Перейти обратно: а б Белеза, Сандра; Сантос, AM; МакЭвой, Б.; Алвес, И.; Мартиньо, К.; Кэмерон, Э.; Шрайвер, доктор медицины; Парра, Э.Дж.; Роча, Дж. (2012). «Сроки осветления пигментации у европейцев» . Молекулярная биология и эволюция . 30 (1): 24–35. дои : 10.1093/molbev/mss207 . ПМЦ 3525146 . ПМИД 22923467 .
- ^ Лалуэса-Фокс; Рёмплер, Х; Карамелли, Д; Штойберт, К; Каталано, Дж; Хьюз, Д; Роланд, Н.; Пилли, Э; Длинный, Л; Кондеми, С; из Расиллы, М; Фортеа, Дж; Росас, А; Стоункинг, М; Шенеберг, Т; Бертранпети, Дж; Хофрейтер, М; и др. (2007). «Аллель рецептора меланокортина-1 предполагает различную пигментацию у неандертальцев». Наука . 318 (5855): 1453–1455. Бибкод : 2007Sci...318.1453L . дои : 10.1126/science.1147417 . ПМИД 17962522 . S2CID 10087710 .
- ^ Перейти обратно: а б Билз, Кеннет Л.; Смит, Кортленд Л; Додд, Стивен М. (1984). «Размер мозга, морфология черепа, климат и машины времени». Современная антропология . 25 (3): 301–330. дои : 10.1086/203138 . S2CID 86147507 .
- ^ Новачевска, Виолетта; Домбровский, Павел; Кузьминский, Лукаш (2011). «Морфологическая адаптация к климату черепов современного человека разумного: важность ширины базисного черепа» . Коллегиум Антропологикум . 35 (3): 625–636. ПМИД 22053534 . Архивировано из оригинала 11 октября 2014 г.
- ^ Пэн, Ю.; и др. (2010). «Полиморфизм ADH1B Arg47His в популяциях Восточной Азии и расширение одомашнивания риса в истории» . Эволюционная биология BMC . 10 (1): 15. Бибкод : 2010BMCEE..10...15P . дои : 10.1186/1471-2148-10-15 . ПМЦ 2823730 . ПМИД 20089146 .
- ^ Сегюрель, Лор; Бон, Селин (2017). «Об эволюции персистенции лактазы у человека». Ежегодный обзор геномики и генетики человека . 18 (1): 297–319. doi : 10.1146/annurev-genom-091416-035340 . ПМИД 28426286 .
- ^ Ингрэм, Кэтрин Дж. Э.; Малкаре, Шарлотта А.; Итан, Юваль; Томас, Марк Г.; Своллоу, Даллас М. (26 ноября 2008 г.). «Переваривание лактозы и эволюционная генетика устойчивости лактазы». Генетика человека . 124 (6): 579–591. дои : 10.1007/s00439-008-0593-6 . ISSN 0340-6717 . ПМИД 19034520 . S2CID 3329285 .
- ^ Илардо, Массачусетс; Мольтке, И.; Корнелиуссен, Т.С.; Ченг, Дж.; Стерн, Эй Джей; Расимо, Ф.; де Баррос Дамгаард, П.; Сикора, М.; Сеген-Орландо, А.; Расмуссен, С.; ван ден Мункхоф, ICL; тер Хорст, Р.; Йоостен, лаборатория; Нетеа, Миннесота; Салингкат, С.; Нильсен, Р.; Виллерслев, Э. (18 апреля 2018 г.). «Физиологическая и генетическая адаптация к нырянию морских кочевников» . Клетка . 173 (3): 569–580.e15. дои : 10.1016/j.cell.2018.03.054 . ПМИД 29677510 .
- ^ Гислен, А; Даке, М; Крегер, Р.Х.; Абрахамссон, М; Нильссон, Делавэр; Ордер, Э.Дж. (2003). «Превосходное подводное зрение у человеческой популяции морских цыган» . Современная биология . 13 (10): 833–836. Бибкод : 2003CBio...13..833G . дои : 10.1016/S0960-9822(03)00290-2 . ПМИД 12747831 . S2CID 18731746 .
- ^ Кляйн, Ричард (1995). «Анатомия, поведение и происхождение современного человека». Журнал мировой предыстории . 9 (2): 167–198. дои : 10.1007/bf02221838 . S2CID 10402296 .
- ^ Фейерман, Джей Р. (2009). Биология религиозного поведения: эволюционные истоки веры и религии . АВС-КЛИО. п. 220. ИСБН 978-0313364303 .
- ^ Соресси М. (2005) Поздняя мустьерская каменная технология. Его значение для темпов возникновения поведенческой современности и взаимосвязи между поведенческой современностью и биологической современностью , стр. 389–417 в книге Л. Бэквелла и Ф. д'Эррико (ред.) От инструментов к символам , Йоханнесбург: Университет Витсуотерсанда. Нажимать. ISBN 1868144178 .
- ^ Сопутствующая энциклопедия археологии (1999). Рутледж. ISBN 0415213304 . Том. 2. п. 763 ( ср ., ... «фактически ограничивается органическими образцами » [ред. органическими соединениями ] «или биогенными карбонатами , возраст которых составляет менее 50 тысяч лет назад (50 000 лет назад)»). См. также: Поздний каменный век и верхний палеолит .
- ^ Мелларс, Пол (2006). «Почему современные человеческие популяции покинули Африку примерно 60 000 лет назад?» . Труды Национальной академии наук . 103 (25): 9381–9386. Бибкод : 2006PNAS..103.9381M . дои : 10.1073/pnas.0510792103 . ПМК 1480416 . ПМИД 16772383 .
- ^ Ши, Джон (2011). «Homo sapiens такой, каким был Homo sapiens». Современная антропология . 52 (1): 1–35. дои : 10.1086/658067 . S2CID 142517998 .
- ^ МакБрирти, Салли; Брукс, Эллисон (2000). «Революция, которой не было: новая интерпретация происхождения современного человеческого поведения». Журнал эволюции человека . 39 (5): 453–563. Бибкод : 2000JHumE..39..453M . дои : 10.1006/jhev.2000.0435 . ПМИД 11102266 . S2CID 42968840 .
- ^ Хеншилвуд, Кристофер; Мэриан, Кертис (2003). «Происхождение современного человеческого поведения: критика моделей и их последствия для испытаний». Современная антропология . 44 (5): 627–651. дои : 10.1086/377665 . ПМИД 14971366 . S2CID 11081605 .
- ^ Мэриан, Кертис; и др. (2007). «Раннее использование человеком морских ресурсов и пигментов в Южной Африке во время среднего плейстоцена» (PDF) . Природа . 449 (7164): 905–908. Бибкод : 2007Natur.449..905M . дои : 10.1038/nature06204 . ПМИД 17943129 . S2CID 4387442 .
- ^ Пауэлл, Адам; и др. (2009). «Демография позднего плейстоцена и внешний вид современного человеческого поведения» (PDF) . Наука . 324 (5932): 1298–1301. Бибкод : 2009Sci...324.1298P . дои : 10.1126/science.1170165 . ПМИД 19498164 . S2CID 206518315 . Архивировано (PDF) из оригинала 29 августа 2017 г.
- ^ Премо, Люк; Кун, Стив (2010). «Моделирование влияния местных вымираний на культурные изменения и разнообразие в палеолите» . ПЛОС ОДИН . 5 (12): e15582. Бибкод : 2010PLoSO...515582P . дои : 10.1371/journal.pone.0015582 . ПМК 3003693 . ПМИД 21179418 .
- ^ Сен-Флер, Николас (12 сентября 2018 г.). «Самый старый известный рисунок, сделанный руками человека, обнаружен в пещере Южной Африки» . Нью-Йорк Таймс . Проверено 15 сентября 2018 г.
- ^ д'Эррико, Ф.; Бэквелл, Л.; Вилла, П.; Дегано, И.; Лусейко, Джей Джей; Бэмфорд, МК; Хайэм, ПФП; Коломбини, член парламента; Бомонт, ПБ (2012). «Ранние свидетельства материальной культуры сан, представленные органическими артефактами из пещеры Бордер, Южная Африка» . Труды Национальной академии наук . 109 (33): 13214–13219. Бибкод : 2012PNAS..10913214D . дои : 10.1073/pnas.1204213109 . ПМК 3421171 . ПМИД 22847420 .
- ^ Мурр, В.; Вилла, П.; Хеншилвуд, CS (2010). «Раннее использование отслаивания под давлением на каменных артефактах в пещере Бломбос, Южная Африка». Наука . 330 (6004): 659–662. Бибкод : 2010Sci...330..659M . дои : 10.1126/science.1195550 . ПМИД 21030655 . S2CID 34833884 .
- ^ Перейти обратно: а б Браун, Кайл С.; Мэриан, Кертис В.; Джейкобс, Зенобия; Шовилл, Бенджамин Дж.; Эстмо, Симен; Фишер, Эрих К.; Бернатчес, Джоселин; Карканас, Панайотис; Мэтьюз, Таласса (2012). «Ранняя и прочная передовая технология, зародившаяся 71 000 лет назад в Южной Африке». Природа . 491 (7425): 590–593. Бибкод : 2012Natur.491..590B . дои : 10.1038/nature11660 . ПМИД 23135405 . S2CID 4323569 .
- ^ Шольц, Калифорния; Джонсон, TC; Коэн, А.С.; Кинг, JW; Пек, Дж. А.; Оверпек, Джей Ти; Талбот, MR; Браун, ET; Калиндекафе, Л.; Амоако, ПЮ; Лайонс, РП; Шанахан, ТМ; Кастанеда, И.С.; Хайль, CW; Форман, СЛ; МакХарг, ЛР; Бенинг, КР; Гомес, Дж.; Пирсон, Дж. (2007). «Мегазасухи в Восточной Африке между 135 и 75 тысячами лет назад и повлияли на происхождение человека раннего Нового времени» . Труды Национальной академии наук . 104 (42): 16416–16421. Бибкод : 2007PNAS..10416416S . дои : 10.1073/pnas.0703874104 . ЧВК 1964544 . ПМИД 17785420 .
- ^ Уэллс, Спенсер (2003). Путешествие человека: генетическая одиссея . Принстон, Нью-Джерси: Издательство Принстонского университета. ISBN 978-0691115320 .
- ^ Хеншилвуд, Кристофер; и др. (2002). «Появление современного человеческого поведения: гравюры среднего каменного века из Южной Африки». Наука . 295 (5558): 1278–1280. Бибкод : 2002Sci...295.1278H . дои : 10.1126/science.1067575 . ПМИД 11786608 . S2CID 31169551 .
- ^ Хеншилвуд, Кристофер С.; д'Эррико, Франческо; Уоттс, Ян (2009). «Гравировка на охре с уровней среднего каменного века в пещере Бломбос, Южная Африка». Журнал эволюции человека . 57 (1): 27–47. Бибкод : 2009JHumE..57...27H . дои : 10.1016/j.jhevol.2009.01.005 . ПМИД 19487016 .
- ^ Тексье, П.Дж.; Порраз, Г; Паркингтон, Дж; Риго, JP; Поггенпол, К; Миллер, К; Триболо, К; Картрайт, К; Куденно, А; Кляйн, Р; Стил, Т; Верна, К. (2010). «Традиция Хоуисонс-Порта гравировать контейнеры из скорлупы страусиных яиц, датируемая 60 000 лет назад, в приюте Дипклоф Рок, Южная Африка» . Труды Национальной академии наук . 107 (14): 6180–6185. Бибкод : 2010PNAS..107.6180T . дои : 10.1073/pnas.0913047107 . ПМК 2851956 . ПМИД 20194764 .
- ^ Перейти обратно: а б МакБрирти, Салли; Брукс, Эллисон (2000). «Революция, которой не было: новая интерпретация происхождения современного человеческого поведения». Журнал эволюции человека . 39 (5): 453–563. Бибкод : 2000JHumE..39..453M . дои : 10.1006/jhev.2000.0435 . ПМИД 11102266 . S2CID 42968840 .
- ^ Хеншилвуд, Кристофер С .; и др. (2004). «Бусы из ракушек среднего каменного века из Южной Африки». Наука . 304 (5669): 404. doi : 10.1126/science.1095905 . ПМИД 15087540 . S2CID 32356688 .
- ^ д'Эррико, Франческо; и др. (2005). «Бусы из ракушек Nassarius kraussianus из пещеры Бломбос: свидетельства символического поведения в среднем каменном веке». Журнал эволюции человека . 48 (1): 3–24. Бибкод : 2005JHumE..48....3D . дои : 10.1016/j.jhevol.2004.09.002 . ПМИД 15656934 .
- ^ Ванхаерен, Мэриан; и др. (2013). «Мыслящие струны: дополнительные доказательства использования личных украшений в среднем каменном веке в пещере Бломбос, Южная Африка». Журнал эволюции человека . 64 (6): 500–517. Бибкод : 2013JHumE..64..500В . дои : 10.1016/j.jhevol.2013.02.001 . ПМИД 23498114 .
- ^ Бэквелл, Л; д'Эррико, Ф; Уодли, Л. (2008). «Костяные орудия среднего каменного века из слоев Ховисонс-Порт, пещера Сибуду, Южная Африка». Журнал археологической науки . 35 (6): 1566–1580. Бибкод : 2008JArSc..35.1566B . дои : 10.1016/j.jas.2007.11.006 .
- ^ Уодли, Лин (2008). «Индустрия Хауисонс-Порт в пещере Сибуду». Серия Гудвина Южноафриканского археологического общества . 10 .
- ^ Ломбард М., Филлипс Л. (2010). «Признаки использования лука и стрел с каменным наконечником 64 000 лет назад в Квазулу-Натале, Южная Африка». Античность . 84 (325): 635–648. дои : 10.1017/S0003598X00100134 . S2CID 162438490 .
- ^ Ломбард М (2011). «Стрелы с кварцевыми наконечниками старше 60 тыс. лет назад: дополнительные свидетельства использования из Сибуду, Ква-Зулу-Натал, Южная Африка». Журнал археологической науки . 38 (8): 1918–1930. Бибкод : 2011JArSc..38.1918L . дои : 10.1016/j.jas.2011.04.001 .
- ^ Бэквелл, Л; Брэдфилд, Дж; Карлсон, К.Дж.; Джашашвили Т; Уодли, Л; д'Эррико, форвард (2018). «Древность технологии изготовления лука и стрел: свидетельства из слоев среднего каменного века в пещере Сибуду» . Журнал археологической науки . 92 (362): 289–303. дои : 10.15184/aqy.2018.11 . hdl : 11336/81248 .
- ^ Ломбард М (2020). «Поперечное сечение кончиков отравленных костяных наконечников стрел из Южной Африки». Журнал археологической науки: отчеты . 33 : 102477. Бибкод : 2020JArSR..33j2477L . дои : 10.1016/j.jasrep.2020.102477 . S2CID 224889105 .
- ^ Йеллен, Дж. Э.; А.С. Брукс; Э. Корнелиссен; М. Дж. Мельман; К. Стюарт (28 апреля 1995 г.). «Средний каменный век работал в костной промышленности из Катанды, Верхней долины Семлики, Заир». Наука . 268 (5210): 553–556. Бибкод : 1995Sci...268..553Y . дои : 10.1126/science.7725100 . ПМИД 7725100 .
- ^ Браун, Кайл С.; Мэриан, Кертис В.; Херрис, Энди И.Р.; Джейкобс, Зенобия; Триболо, Шанталь; Браун, Дэвид; Робертс, Дэвид Л.; Мейер, Майкл С.; Бернатчес, Дж. (14 августа 2009 г.), «Огонь как инженерный инструмент раннего современного человека», Science , 325 (5942): 859–862, Бибкод : 2009Sci...325..859B , doi : 10.1126/science. 1175028 , hdl : 11422/11102 , PMID 19679810 , S2CID 43916405
- ^ Амос, Джонатан (13 октября 2011 г.). «Культурный скачок на заре человечества: обнаружена древняя «фабрика красок»» . Новости Би-би-си . Проверено 13 октября 2011 г.
- ^ Вастаг, Брайан (13 октября 2011 г.). «Южноафриканская пещера дает краски с зарождения человечества» . Вашингтон Пост . Проверено 13 октября 2011 г.
- ^ Хеншилвуд, Кристофер С.; и др. (2011). «100 000-летняя мастерская по обработке охры в пещере Бломбос, Южная Африка». Наука . 334 (6053): 219–222. Бибкод : 2011Sci...334..219H . дои : 10.1126/science.1211535 . ПМИД 21998386 . S2CID 40455940 .
- ^ Шиптон С., д'Эррико Ф., Петралья М. и др. (2018). 78 000-летняя запись инноваций среднего и позднего каменного века в тропическом лесу Восточной Африки. Природные коммуникации
- ^ Сале, Ю.; Хатчингс, ВК; Браун, ДР; Сили, Джей Си; Морган, Ле; Негаш, А.; Атнафу, Б. (2013). Петралья, Майкл Д. (ред.). «Самые ранние снаряды с каменными наконечниками от Эфиопского разлома до > 279 000 лет назад» . ПЛОС ОДИН . 8 (11): e78092. Бибкод : 2013PLoSO...878092S . дои : 10.1371/journal.pone.0078092 . ПМЦ 3827237 . ПМИД 24236011 .
- ^ Мэриан, Кертис; и др. (2007). «Раннее использование человеком морских ресурсов и пигментов в Южной Африке во время среднего плейстоцена» (PDF) . Природа . 449 (7164): 905–908. Бибкод : 2007Natur.449..905M . дои : 10.1038/nature06204 . ПМИД 17943129 . S2CID 4387442 .
- ^ «5 старейших шахт в мире: случайный обзор» . Архивировано из оригинала 5 января 2019 г. Проверено 11 сентября 2019 г.
- ^ Книга рекордов Гиннеса (10 сентября 2015 г.). Книга рекордов Гиннеса 2016 . Книги рекордов Гиннесса. п. 27. ISBN 978-1910561034 .
- ^ Чаттерджи, Риту (15 марта 2018 г.). «Ученые поражены инструментами каменного века, которые они нашли в Кении» . ЭНЕРГЕТИЧЕСКИЙ ЯДЕРНЫЙ РЕАКТОР . Проверено 15 марта 2018 г.
- ^ Йонг, Эд (15 марта 2018 г.). «Культурный скачок на заре человечества. Новые находки из Кении позволяют предположить, что люди использовали торговые сети на большие расстояния, сложные инструменты и символические пигменты еще на заре нашего вида» . Атлантика . Проверено 15 марта 2018 г.
- ^ Брукс А.С., Йеллен Дж.Э., Поттс Р., Беренсмейер А.К., Дейно А.Л., Лесли Д.Э., Амброуз Ш., Фергюсон Дж.Р., д'Эррико Ф., Зипкин А.М., Уиттакер С., Пост Дж., Витч Э.Г., Фёке К., Кларк Дж.Б. (2018). «Перенос камней на большие расстояния и использование пигментов в раннем среднем каменном веке» . Наука . 360 (6384): 90–94. Бибкод : 2018Sci...360...90B . дои : 10.1126/science.aao2646 . ПМИД 29545508 .
- ^ Сале Ю., Брукс А.С. (2018). «Оценка сложных снарядов в раннем позднем плейстоцене в Адуме, Эфиопия» . ПЛОС ОДИН . 14 (5): e0216716. Бибкод : 2019PLoSO..1416716S . дои : 10.1371/journal.pone.0216716 . ПМК 6508696 . ПМИД 31071181 .
- ^ Дуглас, Кейт (24 марта 2012 г.). «Загадки эволюции: почему технологическое развитие было таким медленным?» . Новый учёный . Архивировано из оригинала 13 апреля 2016 года . Проверено 4 мая 2022 г.
{{cite magazine}}: CS1 maint: неподходящий URL ( ссылка )
Источники
- Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арсьеро Э., Хуанмин Ю., Томас М.Г., Сюэ Ю., Тайлер-Смит С. (июнь 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- Юблин, Жан-Жак; Бен-Нсер, Абделуахед; Бейли, Шара Э.; Фридлайн, Сара Э.; Нойбауэр, Саймон; Скиннер, Мэтью М.; Бергманн, Инга; Ле Кабек, Аделина; Бенацци, Стефано; Харвати, Катерина; Гунц, Филипп (2017). «Новые окаменелости из Джебель-Ирхуда, Марокко, и панафриканское происхождение Homo sapiens » (PDF) . Природа . 546 (7657): 289–292. Бибкод : 2017Natur.546..289H . дои : 10.1038/nature22336 . ПМИД 28593953 . S2CID 256771372 .
- Камин М., Сааг Л., Винсенте М. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- Пост К, Рено Г, Миттник М, Друкер Д. Г., Ружье Х, Купийяр С, Валентин Ф, Тевене С, Фуртвенглер А, Виссинг С, Франкен М, Малина М, Болюс М, Лари М, Джильи Е, Капечки Г, Кревекёр И , Боваль С, Флас Д, Жермонпре М, ван дер Плихт Дж, Коттио Р, Жели Б, Рончителли А, Вербергер К, Григореску Д, Свобода Дж, Семаль П, Карамелли Д, Бочеренс Х, Харвати К, Конард Н.Дж., Хаак В. , Пауэлл А., Краузе Дж. (2016). «Митохондриальные геномы плейстоцена предполагают единое крупное расселение неафриканцев и позднеледниковую смену населения в Европе». Современная биология . 26 (6): 827–833. Бибкод : 2016CBio...26..827P . дои : 10.1016/j.cub.2016.01.037 . hdl : 2440/114930 . ПМИД 26853362 . S2CID 140098861 .
- Образец, Ян (7 июня 2017 г.). «Самые древние кости Homo sapiens , когда-либо найденные, потрясают основы человеческой истории» . Хранитель . Проверено 7 июня 2017 г.
- Шерри, МЛ; и др. (2018). «Эволюционировали ли наши виды в разделенных популяциях по всей Африке, и почему это имеет значение?» . Тенденции в экологии и эволюции . 33 (8): 582–594. Бибкод : 2018TEcoE..33..582S . дои : 10.1016/j.tree.2018.05.005 . ПМК 6092560 . ПМИД 30007846 .
- Стрингер, К. (2016). «Происхождение и эволюция человека разумного» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1698): 20150237. doi : 10.1098/rstb.2015.0237 . ПМЦ 4920294 . ПМИД 27298468 .
- Вай С., Сарно С., Лари М., Луизелли Д., Манци Г., Галлинаро М., Матаич С., Хюбнер А., Моди А., Пилли Е., Тафури М.А., Карамелли Д., ди Лерния С. (март 2019 г.). «Предковая митохондриальная линия N из неолитической «зеленой» Сахары» . Лыжный представитель . 9 (1): 3530. Бибкод : 2019НатСР...9.3530В . дои : 10.1038/s41598-019-39802-1 . ПМК 6401177 . ПМИД 30837540 .
Дальнейшее чтение
- Райх, Дэвид (2018). Кто мы и как мы сюда попали – древняя ДНК и новая наука о человеческом прошлом . Книги Пантеона . ISBN 978-1101870327 . [1]
Внешние ссылки
- Хронология человечества (интерактивная) – Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).
 СМИ, связанные с человеком раннего Нового времени, на Викискладе?
СМИ, связанные с человеком раннего Нового времени, на Викискладе?  Данные, относящиеся к раннему современному человеку , в Wikispecies
Данные, относящиеся к раннему современному человеку , в Wikispecies
- ^ Даймонд, Джаред (20 апреля 2018 г.). «Совершенно новая версия истории нашего происхождения» . Нью-Йорк Таймс . Проверено 23 апреля 2018 г.