Ранние миграции человека
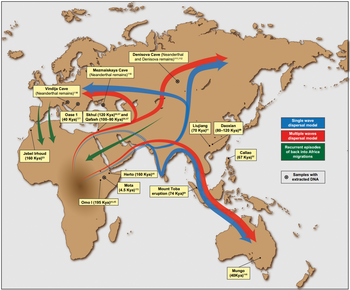
Ранние миграции человека — это самые ранние миграции и расселения архаичных и современных людей по континентам. Считается, что они начались примерно 2 миллиона лет назад с Африки из ранним распространением человека прямоходящего . За этой первоначальной миграцией последовали другие архаичные люди, включая H. heidelbergensis , который жил около 500 000 лет назад и был вероятным предком денисовцев и неандертальцев, а также современных людей. Ранние гоминиды, вероятно, пересекали сухопутные мосты , которые сейчас затонули.
В Африке Homo sapiens рассеялся примерно во время своего видообразования , примерно 300 000 лет назад. [примечание 1] Недавняя парадигма африканского происхождения предполагает, что анатомически современные люди за пределами Африки произошли от популяции Homo sapiens, мигрировавшей из Восточной Африки примерно 70–50 000 лет назад и распространившейся вдоль южного побережья Азии и в Океанию примерно 50 000 лет назад. Современные люди распространились по Европе около 40 000 лет назад.
Окаменелости ранних евразийских Homo sapiens были найдены в Израиле и Греции, их возраст составляет 194 000–177 000 и 210 000 лет соответственно. Эти окаменелости, похоже, представляют собой неудачные попытки расселения ранних Homo sapiens , которых, вероятно, заменили местные популяции неандертальцев.
Известно, что мигрирующие современные человеческие популяции скрещивались с более ранними местными популяциями, так что современные человеческие популяции в небольшой части (менее 10%) произошли от региональных разновидностей архаичных людей. [примечание 2]
После последнего ледникового максимума население Северной Евразии мигрировало в Америку около 20 000 лет назад. Арктическая Канада и Гренландия были достигнуты палеоэскимосской экспансией около 4000 лет назад. Наконец, Полинезия была заселена в течение последних 2000 лет во время последней волны австронезийской экспансии .
Ранние люди (до Homo sapiens )
[ редактировать ]Самые ранние люди произошли от предков австралопитеков около 3 миллионов лет назад, скорее всего, в районе Кенийской рифтовой долины , где древнейшие известные каменные орудия были найдены . Каменные орудия, недавно обнаруженные на стоянке Шанчен в Китае и датированные 2,12 миллиона лет назад, считаются самым ранним известным свидетельством существования гомининов за пределами Африки, превосходящим Дманиси в Грузии на 300 000 лет. [6]
Мужчина встал
[ редактировать ]Между 2 и менее миллиона лет назад Homo распространился по Восточной Африке и Южной Африке ( Homo ergaster ), но еще не в Западной Африке. Около 1,8 миллиона лет назад Homo erectus мигрировал из Африки через Левантийский коридор и Африканский Рог в Евразию . Предполагается, что эта миграция связана с работой сахарского насоса около 1,9 миллиона лет назад. [ нужна ссылка ] Homo erectus расселился по большей части Старого Света , достигнув Юго -Восточной Азии . Его распространение прослеживается по олдованской каменной индустрии, которая 1,3 миллиона лет назад простиралась на север до 40-й параллели ( Сяочанлян ).
Ключевыми местами этой ранней миграции из Африки являются Риват в Пакистане (около 2 млн лет назад? [7] ), Убейдия в Леванте (1,5 млн лет) и Дманиси на Кавказе (1,81 ± 0,03 млн лет, p = 0,05). [8] ).
Китай демонстрирует свидетельства существования Homo erectus, жившего 2,12 млн лет назад в Гунванлине, уезд Ланьтянь. [9] Два резца человека прямоходящего были найдены недалеко от Юаньмоу на юге Китая и датированы 1,7 млн лет назад, а череп из Ланьтяня был датирован 1,63 млн лет назад. Артефакты из Маджуангоу III и Шаншазуй в бассейне Нихевань на севере Китая датируются 1,6–1,7 млн лет назад. [9] [10] Археологический памятник Сиходу ( 西侯渡 ) в провинции Шаньси является самым ранним зарегистрированным использованием огня человеком прямоходящим , датируемым 1,27 миллиона лет назад. [11]
Юго-Восточная Азия ( Ява ) была достигнута около 1,7 миллиона лет назад ( Мегантроп ). Западная Европа была впервые заселена около 1,2 миллиона лет назад ( Атапуэрка ). [12]
Роберт Г. Беднарик предположил, что Homo erectus, возможно, строил плоты и плавал по океанам, и эта теория вызвала некоторые споры. [13]
После H. erectus
[ редактировать ]
Через миллион лет после своего расселения H. erectus начал выделяться на новые виды. H. erectus является хроновидом и никогда не вымерал, поэтому его «позднее выживание» является вопросом таксономической конвенции. Считается, что поздние формы H. erectus сохранились примерно от 0,5 миллиона лет назад до 143 000 лет назад, самое позднее. [примечание 3] с производными формами, классифицированными как H. antecessor в Европе около 800 000 лет назад и H. heidelbergensis в Африке около 600 000 лет назад. H. heidelbergensis, в свою очередь, распространился по Восточной Африке ( H. rhodesiensis ) и Евразии, где дал начало неандертальцам и денисовцам .
H. heidelbergensis , неандертальцы и денисовцы распространились на север за пределы 50-й параллели ( Яма Эртем, Самшитовая роща, 500 тыс. лет назад, Парк наследия Суонскомб, 400 тыс. лет назад, Денисова пещера , 50 тыс. лет назад). Было высказано предположение, что поздние неандертальцы, возможно, даже достигли границы Арктики примерно к ок. 32 000 лет назад, когда они были вытеснены из прежних мест обитания H. sapiens , по данным раскопок 2011 года на стоянке Бызовая на Урале ( Республика Коми , 65 ° 01' с.ш. 57 ° 25' в.д. / 65,02 ° с.ш. 57,42 ° в.д. ). [15]

Предполагается, что к этому времени по всей Африке распространились и другие архаичные виды человека, хотя летопись окаменелостей скудна. Их присутствие предполагается на основании следов смешения с современным человеком, обнаруженных в геноме африканских популяций. [5] [16] [17] [18] Homo naledi , обнаруженный в Южной Африке в 2013 году и предположительно датируемый примерно 300 000 лет назад, может представлять собой ископаемые свидетельства существования такого архаичного человеческого вида. [19]
Неандертальцы распространились по Ближнему Востоку и Европе, а денисовцы, по-видимому, распространились по Центральной и Восточной Азии, а также по Юго-Восточной Азии и Океании. Есть свидетельства того, что денисовцы скрещивались с неандертальцами в Центральной Азии, где их ареалы пересекались. [20] Свидетельства существования неандертальцев были также обнаружены довольно поздно, 33 000 лет назад, на 65-й широте стоянки Бызовая в Уральских горах . Это находится далеко за пределами любой другой известной среды обитания, в период высокого ледяного покрова, и, возможно, отражает рефугиум, находящийся на грани исчезновения.
Мудрый человек
[ редактировать ]Распространение по Африке
[ редактировать ]Считается, что Homo sapiens появился в Африке около 300 000 лет назад, что частично основано на термолюминесцентном датировании артефактов и останков из Джебель-Ирхуда , Марокко, опубликованном в 2017 году. [примечание 4] [22] Флорисбадский череп из Флорисбада, Южная Африка, датируемый примерно 259 000 лет назад, также был классифицирован как ранний Homo sapiens . [23] [24] [25] [26] Ранее останки Омо , раскопанные между 1967 и 1974 годами в Национальном парке Омо и в Эфиопии датированные 200 000 лет назад, долгое время считались старейшими известными окаменелостями Homo sapiens . [27]
В сентябре 2019 года ученые сообщили о компьютеризированном определении на основе 260 компьютерных томографий виртуальной формы черепа последнего общего человеческого предка анатомически современных людей, представителя самых ранних современных людей, и предположили, что современные люди возникли между 260 000 и 350 000 лет назад. назад в результате слияния населения Восточной и Южной Африки . [28] [29]
В июле 2019 года антропологи сообщили об открытии останков H. sapiens возрастом 210 000 лет и останков H. neanderthalensis возрастом 170 000 лет в пещере Апидима на юге Греции , что более чем на 150 000 лет старше, чем предыдущие H. sapiens в Европе. находки [30] [31] [32] [33]
С момента своего появления ранние современные люди распространились на Западную Евразию, Центральную, Западную и Южную Африку. Хотя ранняя экспансия в Евразию, похоже, не имела продолжения, [34] [20] Экспансия в Южную и Центральную Африку привела к глубочайшим временным расхождениям в ныне живущих человеческих популяциях. Ранняя современная человеческая экспансия в Африке к югу от Сахары, по-видимому, способствовала исчезновению позднеашельской ( форсмитской ) промышленности примерно 130 000 лет назад, хотя очень позднее сосуществование архаичных и ранних современных людей, вплоть до 12 000 лет назад, было В частности, выступал за Западную Африку. [35]
Предки современного кой-сана распространились на Южную Африку около 150 000 лет назад, возможно, еще около 260 000 лет назад. [примечание 5] так что к началу » MIS 5 « мегазасухи , 130 000 лет назад, в Африке существовало два наследственных кластера населения, носителей гаплогруппы L0 мт-ДНК на юге Африки, предков кой-сан, и носителей гаплогруппы L1- 6 в центральной/восточной Африке, предок всех остальных. Между 120 и 75 тыс. лет назад произошла значительная обратная миграция носителей L0 в Восточную Африку. [примечание 6]
Экспансия в Центральную Африку предков центральноафриканских популяций собирателей (африканских пигмеев), скорее всего, произошла до 130 000 лет назад и, конечно, до 60 000 лет назад. [37] [38] [39] [40] [примечание 7]
Ситуацию в Западной Африке трудно интерпретировать из-за скудности ископаемых свидетельств. Homo sapiens , по-видимому, достиг зоны западного Сахеля 130 000 лет назад, в то время как тропические места Западной Африки, связанные с H. sapiens, известны только после 130 000 лет назад. В отличие от других регионов Африки, архаичные стоянки среднего каменного века , по-видимому, сохраняются до очень позднего периода, вплоть до границы голоцена (12 000 лет назад), что указывает на возможность позднего выживания архаичных людей и поздней гибридизации с H. sapiens в Западной Африке. [35]
Раннее расселение в Северной Африке
[ редактировать ]
Популяции Homo sapiens мигрировали в Левант и Европу. [ сомнительно – обсудить ] между 130 000 и 115 000 лет назад и, возможно, в более ранних волнах, уже 185 000 лет назад. [примечание 8]
Фрагмент челюстной кости с восемью зубами, найденный в пещере Мислия, датируется примерно 185 000 лет назад. Слои, датированные периодом от 250 000 до 140 000 лет назад в той же пещере, содержали инструменты леваллуазского типа , что позволило бы установить дату первой миграции еще раньше, если бы эти инструменты можно было связать с находками современных человеческих челюстных костей. [42] [43]
Эти ранние миграции, похоже, не привели к длительной колонизации и отступили примерно 80 000 лет назад. [20] Существует вероятность того, что эта первая волна экспансии достигла Китая (или даже Северной Америки). [ сомнительно – обсудить ] [44] ) еще 125 000 лет назад, но вымерли бы, не оставив следа в геноме современного человека. [20]

Есть некоторые свидетельства того, что современные люди покинули Африку по крайней мере 125 000 лет назад, используя два разных маршрута: через долину Нила , Синайский полуостров и Левант ( пещера Кафзе : 120 000–100 000 лет назад); и второй маршрут через современный Баб-эль-Мандебский пролив в Красном море (в то время с гораздо более низким уровнем моря и более узкой протяженностью), пересекающий Аравийский полуостров. [45] [46] и поселились в таких местах, как современные Объединенные Арабские Эмираты (125 000 лет назад). [47] и Оман (106 000 лет назад), [48] и, возможно, достигнув Индийского субконтинента ( Джвалапурам : 75 000 лет назад). Хотя в этих трех местах до сих пор не обнаружено человеческих останков, очевидное сходство между каменными орудиями, найденными в Джебель-Фая , орудиями из Джвалапурама и некоторыми из Африки, позволяет предположить, что их создатели все были современными людьми. [49] Эти находки могут дать некоторую поддержку утверждению о том, что современные люди из Африки прибыли в южный Китай около 100 000 лет назад ( Пещера Жирен , Жирендун , город Чунцзуо : 100 000 лет назад; [примечание 9] и Люцзянский гоминид ( уезд Люцзян ): датировка спорная: 139 000–111 000 лет назад. [54] ). Результаты датирования зубов Лунадонга ( бассейн Бубин , Гуанси , южный Китай ), которые включают правый верхний второй моляр и левый нижний второй моляр, показывают, что возраст коренных зубов может достигать 126 000 лет. [55] [56]
Поскольку эти предыдущие выходы из Африки не оставили следов в результатах генетического анализа на основе Y-хромосомы и мтДНК, похоже, что те современные люди не выжили в больших количествах и были ассимилированы нашими основными предками. Объяснением их исчезновения (или небольшого генетического отпечатка) может быть извержение Тоба (74 000 лет назад), хотя некоторые утверждают, что оно почти не затронуло человеческую популяцию. [57]
Прибрежная миграция
[ редактировать ]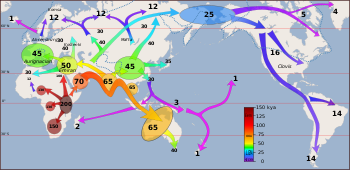
Так называемое « недавнее расселение » современного человека произошло около 70–50 000 лет назад. [58] [59] [60] Именно эта миграционная волна привела к прочному распространению современного человека по всему миру.
Небольшая группа населения Восточной Африки, несущая митохондриальную гаплогруппу L3 и насчитывающая, возможно, менее 1000 человек. [61] [62] пересек пролив Красного моря в Баб-эль-Мандебе , на территории современного Йемена , примерно 75 000 лет назад. [63] Недавний обзор также показал поддержку северного маршрута через Синайский полуостров и Левант . [20] Их потомки распространились по прибрежному маршруту вокруг Аравии и Персии в Южную Азию еще 55 000 лет назад. Другие исследования подтверждают миграцию из Африки примерно 65 000–50 000 лет назад. [58] [64] [60] Прибрежная миграция примерно 70 000–50 000 лет назад связана с митохондриальными гаплогруппами M и N , обе производными от L3.
По пути H. sapiens скрещивался с неандертальцами и денисовцами, [65] ДНК денисовца составляет 0,2% ДНК материковой Азии и коренных американцев. [66]
Соседняя Океания
[ редактировать ]Миграции продолжались вдоль азиатского побережья в Юго-Восточную Азию и Океанию, колонизируя Австралию примерно 65 000–50 000 лет назад. [67] [68] [69] Достигнув Австралии, H. sapiens впервые расширил свою среду обитания за пределы ареала H. erectus . Денисовское происхождение разделяют меланезийцы , австралийские аборигены и более мелкие разбросанные группы людей в Юго-Восточной Азии, такие как маманва , народ негрито на Филиппинах , что позволяет предположить, что скрещивание имело место в Восточной Азии, где жили денисовцы. [70] [71] [72] Денисовцы, возможно, пересекли линию Уоллеса , и Уоллеса служила их последним убежищем . [73] [74] Homo erectus пересек пролив Ломбок и достиг Флореса, но так и не добрался до Австралии. [75]

В это время уровень моря был намного ниже, и большая часть Приморской Юго-Восточной Азии образовала единый массив суши, известный как Зонд . Миграция продолжалась на юго-восток по прибрежному маршруту к проливам между Зондой и Сахулом , континентальным массивом современной Австралии и Новой Гвинеи . Проломы на линии Вебера имеют ширину до 90 км. [76] поэтому миграция в Австралию и Новую Гвинею потребовала бы навыков мореплавания. Миграция также продолжалась вдоль побережья, в конечном итоге повернув на северо-восток в сторону Китая и, наконец, достигнув Японии, а затем повернуть вглубь страны. Об этом свидетельствует структура митохондриальных гаплогрупп, произошедших от гаплогруппы М , и в Y-хромосомы гаплогруппе С .
Секвенирование генома одного аборигена из старого образца волос в Западной Австралии показало, что этот человек произошел от людей, которые мигрировали в Восточную Азию между 62 000 и 75 000 лет назад. Это подтверждает теорию единой миграции в Австралию и Новую Гвинею до прибытия современных азиатов (между 25 000 и 38 000 лет назад) и их более поздней миграции в Северную Америку. [77] Считается, что эта миграция произошла около 50 000 лет назад, до того, как Австралия и Новая Гвинея были разделены повышением уровня моря примерно 8 000 лет назад. [78] [79] Это подтверждается датой 50 000–60 000 лет назад для самых старых свидетельств поселения в Австралии. [67] [80] около 40 000 лет назад для древнейших человеческих останков, [67] самые ранние человеческие артефакты, которым не менее 65 000 лет. [81] и исчезновение австралийской мегафауны людьми между 46 000 и 15 000 лет назад, о чем утверждает Тим Флэннери, [82] что похоже на то, что произошло в Америке. Продолжающееся использование орудий каменного века в Австралии вызывает много споров. [83]
Распространение по Евразии
[ редактировать ]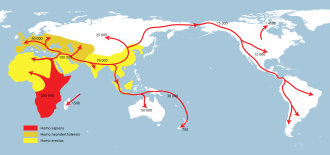
Homo erectus наибольшей степени (желтый),
Homo neanderthalensis в наибольшей степени (охра) и
Человек разумный (красный).
Население, завезенное в Южную Азию в результате прибрежной миграции, по-видимому, оставалось там в течение некоторого времени, примерно от 60 000 до 50 000 лет назад, прежде чем распространиться дальше по Евразии. Это расселение ранних людей в начале верхнего палеолита привело к возникновению основных групп населения Старого Света и Америки .
На западе верхнепалеолитические популяции, связанные с митохондриальной гаплогруппой R и ее производными, распространились по всей Азии и Европе с обратной миграцией M1 в Северную Африку и на Африканский Рог несколько тысячелетий назад. [ сомнительно – обсудить ]
Присутствие в Европе несомненно после 40 000 лет назад, возможно, уже 43 000 лет назад. [84] быстро вытеснили популяцию неандертальцев. Современные европейцы имеют неандертальское происхождение , но вполне вероятно, что существенное скрещивание с неандертальцами прекратилось до 47 000 лет назад, то есть имело место до того, как современные люди проникли в Европу. [85]
свидетельствуют Исследования митохондриальной ДНК о том, что современные люди прошли по крайней мере через одно генетическое узкое место , в котором разнообразие генома резко сократилось. Генри Харпендинг предположил, что люди распространились из географически ограниченной области около 100 000 лет назад, пройдя через географическое узкое место, а затем резко увеличившись среди географически рассредоточенных популяций около 50 000 лет назад, начиная сначала в Африке, а затем распространившись в других местах. [86] Климатологические и геологические данные свидетельствуют о наличии узкого места. Взрыв Тоба , крупнейшего извержения вулкана четвертичного периода , возможно, создал 1000-летний холодный период, потенциально сократив человеческое население до нескольких тропических рефугиумов. По оценкам, выжило всего лишь 15 000 человек. В таких обстоятельствах генетический дрейф и эффекты основателя могли быть максимальными. Большее разнообразие африканских геномов может отражать масштабы африканских рефугиумов во время инцидента в Тобе. [87] Однако недавний обзор подчеркивает, что гипотеза единого источника неафриканского населения менее согласуется с анализом древней ДНК, чем гипотеза множественных источников с генетическим смешением по всей Евразии. [20]
Европа
[ редактировать ]
Недавняя экспансия анатомически современных людей достигла Европы около 40 000 лет назад из Центральной Азии и Ближнего Востока в результате культурной адаптации к охоте на крупную дичь подледниковой степной фауны. [88] Неандертальцы присутствовали как на Ближнем Востоке, так и в Европе, а прибывшие популяции анатомически современных людей (также известных как « кроманьонцы » или европейские ранние современные люди ) скрещивались с популяциями неандертальцев в ограниченной степени. Популяции современных людей и неандертальцев пересекались в различных регионах, таких как Пиренейский полуостров и Ближний Восток. Скрещивание, возможно, способствовало появлению генов неандертальцев в палеолите и, в конечном итоге, в современных евразийцах и океанийцах.
Важным отличием Европы от других частей обитаемого мира была северная широта. Археологические данные свидетельствуют о том, что люди, будь то неандертальцы или кроманьонцы, достигли мест в арктической части России 40 000 лет назад. [89]
Кроманьонцы считаются первыми анатомически современными людьми в Европе. Они вошли в Евразию через горы Загрос (около современного Ирана и восточной Турции ) около 50 000 лет назад, при этом одна группа быстро заселила прибрежные районы вокруг Индийского океана , а другая мигрировала на север, в степи Центральной Азии . [90] Останки современного человека, датируемые 45 000–47 000 лет, были найдены в Германии . [91] а в Италии обнаружены находки 43 000–45 000 лет назад. [92] и Британия, [93] а также в европейской части Российской Арктики 40 000 лет назад. [89] [94]
Люди колонизировали окружающую среду к западу от Урала, особенно охотясь на северных оленей. [95] но столкнулись с проблемами адаптации; Зимние температуры в среднем составляли от -20 до -30 ° C (от -4 до -22 ° F), при нехватке топлива и укрытия. Они путешествовали пешком и в качестве пропитания полагались на охоту на очень подвижные стада. Эти проблемы были преодолены за счет технологических инноваций: пошив одежды из шкур пушных зверей; строительство убежищ с очагами, использующими кости в качестве топлива; и рытье «ледяных погребов» в вечной мерзлоте для хранения мяса и костей. [95] [96]
Однако, согласно недавним исследованиям, считается, что экологический кризис, возникший в результате извержения в c. 38 000 г. до н.э. извержение супервулкана на Флегрейских полях недалеко от Неаполя, в результате которого большая часть Восточной Европы была покрыта пеплом, уничтожило как последнюю неандертальскую, так и первую популяцию Homo Sapiens раннего верхнего палеолита. [97] [98] Сегодняшние современные европейцы не несут в себе никаких следов геномов первых европейцев Homo Sapiens, а только тех, которые появились после экологического кризиса 38 000 г. до н.э. [99] Современные люди затем вновь заселили Европу с востока после извержения и ледникового периода, который произошел с 38 000 по 36 000 лет до нашей эры. [100]
Последовательность митохондриальной ДНК двух кроманьонцев из пещеры Пагличи в Италии, датированная возрастом 23 000 и 24 000 лет (Пагличи 52 и 12), идентифицировала мтДНК как гаплогруппу N , типичную для последней группы. [101]
(YBP = Годы до настоящего времени )



Считается, что расширение современного человеческого населения началось 45 000 лет назад, и для колонизации Европы могло потребоваться 15 000–20 000 лет. [103] [104]
В это время неандертальцы медленно вытеснялись. Поскольку оккупация Европы заняла так много времени, похоже, что люди и неандертальцы могли постоянно конкурировать за территорию. У неандертальцев был больший мозг и в целом они были крупнее, с более крепким или тяжелым телосложением, что позволяет предположить, что они были физически сильнее, чем современные Homo sapiens . Прожив в Европе 200 000 лет, они лучше приспособились к холодной погоде. Анатомически современные люди, известные как кроманьонцы , с широко распространенными торговыми сетями, превосходными технологиями и телом, вероятно, лучше приспособленным к бегу, в конечном итоге полностью вытеснили неандертальцев, последним прибежищем которых был Пиренейский полуостров . Неандертальцы исчезли около 40 000 лет назад. [105]
Судя по степени неравновесия по сцеплению, было подсчитано, что последний поток генов неандертальцев к ранним предкам европейцев произошел 47 000–65 000 лет назад . В сочетании с археологическими и ископаемыми свидетельствами предполагается, что скрещивание произошло где-то в Западной Евразии, возможно, на Ближнем Востоке. [85] Исследования показывают более высокую примесь неандертальцев у жителей Восточной Азии, чем у европейцев. [106] [107] Группы Северной Африки имеют такой же избыток производных аллелей, что и неандертальцы, как и неафриканские популяции, тогда как группы Африки к югу от Сахары являются единственными современными человеческими популяциями без существенной примеси неандертальцев. [примечание 10] Сцепленный с неандертальцами гаплотип B006 гена дистрофина также был обнаружен среди групп кочевых скотоводов в Сахеле и Африканском Роге, которые связаны с северными популяциями. Следовательно, присутствие этого гаплотипа B006 на северном и северо-восточном периметре Африки к югу от Сахары объясняется потоком генов из неафриканской точки происхождения. [примечание 11]
Восточная, Юго-Восточная и Северная Азия
[ редактировать ]
« Человек Тяньюань », человек, живший в Китае ок. 40 000 лет назад была обнаружена значительная примесь неандертальцев. Исследование древней ДНК человека Тяньюань, проведенное в 2017 году, показало, что этот человек связан с современным населением Азии и коренных американцев. [111] Исследование 2013 года выявило интрогрессию неандертальцами 18 генов в области хромосомы 3p21.31 (область HYAL) жителей Восточной Азии. Интрогрессивные гаплотипы были положительно отобраны только в популяциях Восточной Азии, их рост неуклонно рос с 45 000 лет назад до внезапного увеличения скорости роста примерно 5 000–3 500 лет назад. Они встречаются с очень высокой частотой среди популяций Восточной Азии в отличие от других популяций Евразии (например, популяций Европы и Южной Азии). Результаты также позволяют предположить, что эта интрогрессия неандертальцев произошла в пределах наследственной популяции, разделяющей аборигенов Восточной Азии и коренных американцев. [112]
В исследовании 2016 года представлен анализ популяционной генетики айнов северной Японии как ключ к реконструкции раннего заселения Восточной Азии. Было обнаружено, что айны представляют собой более базальную ветвь, чем современное земледельческое население Восточной Азии, что позволяет предположить древнюю (донеолитическую) связь с жителями северо-восточной Сибири. [113] Исследование 2013 года связало несколько фенотипических черт, связанных с монголоидами, с единственной мутацией гена EDAR , датируемой ок. 35 000 лет назад. [примечание 12] [примечание 13]
Митохондриальные гаплогруппы A , B и G возникли около 50 000 лет назад, и их носители впоследствии колонизировали Сибирь , Корею и Японию примерно 35 000 лет назад . Части этих популяций мигрировали в Северную Америку во время последнего ледникового максимума .
В обзорном документе Мелинды А. Янг (2022 г.) резюмировано и сделано заключение о том, что особая «базальная восточноазиатская популяция», называемая « линиями Восточной и Юго-Восточной Азии » (ESEA); который является предком современных аборигенов Восточной Азии, Юго-Восточной Азии , полинезийцев и сибиряков , зародился в материковой части Юго-Восточной Азии примерно в 50 000 году до нашей эры и распространился посредством многочисленных миграционных волн на юг и на север соответственно. Эта линия ESEA дала начало различным подлиниям, а также является предком хоабиньских охотников-собирателей , возраст которой ~ 40 000 лет, из Юго-Восточной Азии и линии Тяньюань найденной в Северном Китае , но уже дифференцированной и отличной от родственных европейских и австралазийских линий. Встречаются и в других регионах доисторической Евразии. Линия ESEA отделилась от более ранней восточно-евразийской или «восточно-неафриканской» (ENA) метапопуляции, которая также способствовала формированию древних предков южных индейцев (AASI), а также австралазийцев. [117]
Последний ледниковый максимум
[ редактировать ]Евразия
[ редактировать ]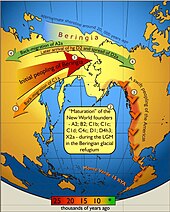
Около 20 000 лет назад, примерно через 5 000 лет после вымирания неандертальцев, Последний ледниковый максимум заставил жителей северного полушария мигрировать в несколько убежищ ( рефугиумов ) до конца этого периода. Предполагается, что образовавшееся население проживало в таких убежищах во время LGM, чтобы в конечном итоге вновь оккупировать Европу, где архаичные исторические популяции считаются их потомками. Позднее состав европейского населения был изменен в результате дальнейших миграций, особенно неолитической экспансии с Ближнего Востока, а еще позже - энеолитических перемещений населения, связанных с индоевропейской экспансией . Палеолитическая стоянка на реке Яна в Сибири, на 71° с.ш., находится значительно выше Полярного круга и датируется 27 000 радиоуглеродными годами раньше настоящего времени, во времена ледникового периода. Этот сайт показывает, что люди адаптировались к этой суровой, высокоширотной среде позднего плейстоцена гораздо раньше, чем считалось ранее. [118]
Америка
[ редактировать ]Палеоиндейцы произошли из Центральной Азии , перейдя Берингийский сухопутный мост между Восточной Сибирью и современной Аляской. [119] Люди жили по всей Америке к концу последнего ледникового периода , или, точнее, того, что известно как поздний ледниковый максимум . [119] [120] [121] [122] Подробности миграции палеоиндейцев на американский континент и по его территории, включая даты и маршруты путешествия, являются предметом продолжающихся исследований и обсуждений. [123]
По общепринятым оценкам, люди достигли Северной Америки где-то между 15 000 и 20 000 лет назад. [124] [125] [126] [127] Традиционная теория состоит в том, что эти ранние мигранты переселились, когда уровень моря значительно снизился из-за четвертичного оледенения . [120] [123] преследуя стада ныне вымершей плейстоценовой мегафауны по свободным ото льда коридорам , простиравшимся между ледниковыми щитами Лаврентиды и Кордильеров . [128] Другой предложенный маршрут заключается в том, что либо пешком, либо на примитивных лодках они мигрировали вдоль побережья Тихого океана в Южную Америку до Чили . [129] Любые археологические свидетельства заселения побережья во время последнего ледникового периода теперь были бы скрыты повышением уровня моря , которое с тех пор достигло ста метров. [130] Недавнее обнаружение коренных австралазийских генетических маркеров в Амазонии подтверждает, что некоторые мигранты действительно имели прибрежный маршрут и последующую изоляцию. [131]
Голоценовые миграции
[ редактировать ]
начался Считается, что голоцен 12 000 лет назад, после окончания последнего ледникового максимума . Во время климатического оптимума голоцена , начавшегося около 9000 лет назад, человеческие популяции, географически ограниченные рефугиумами, начали мигрировать. К этому времени большая часть земного шара была заселена H. sapiens ; однако большие территории, которые раньше были покрыты ледниками , теперь были вновь заселены.
происходит переход от мезолита к неолиту В этот период во всем умеренном поясе . Неолит впоследствии уступает место бронзовому веку в культурах Старого Света и постепенному появлению исторических свидетельств на Ближнем Востоке и в Китае, начавшемуся около 4000 лет назад.
Считается, что крупномасштабные миграции от мезолита к эпохе неолита привели к досовременному распространению основных языковых семей мира, таких как нигер-конголезская , нило-сахарская , афро-азиатская , уральская , китайско-тибетская или индоевропейская. -Европейский тип. Спекулятивная ностратическая теория постулирует происхождение основных языковых семей Евразии (за исключением сино-тибетской) от единого праязыка, на котором говорили в начале периода голоцена.
Евразия
[ редактировать ]Данные, опубликованные в 2014 году на основе анализа генома древних человеческих останков, позволяют предположить, что современное коренное население Европы в значительной степени происходит от трех различных линий: « западные охотники-собиратели », производные от кроманьонского населения Европы, ранние европейские фермеры, завезенные в Европу из Ближний Восток во время неолитической революции и древние жители Северной Евразии , распространившиеся в Европу в контексте индоевропейской экспансии . [133] Древний североевразийский компонент был занесен в Западную Европу людьми, связанными с ямной культурой . [134] Дополнительная родословная ANE обнаружена в европейских популяциях в результате палеолитических взаимодействий с восточными охотниками-собирателями . [135]
Африка к югу от Сахары
[ редактировать ]Западно-евразийские обратные миграции начались в раннем голоцене или уже раньше, в период палеолита (30-15 тыс. лет назад), за которыми последовали донеолитические и неолитические миграционные события с Ближнего Востока , в основном затронувшие Северную Африку, Африканский Рог и более широкие территории. регионы зоны Сахеля и Восточной Африки. [136]

Считается, что нилотские народы произошли от более раннего недифференцированного восточно-суданского единства, существовавшего в 3-м тысячелетии до нашей эры. Развитие протонилотов как группы могло быть связано с приручением ими домашнего скота . Восточно-суданское единство должно было возникнуть еще значительно раньше, возможно, около 5-го тысячелетия до нашей эры (тогда как предполагаемое нило-сахарское единство датируется верхним палеолитом около 15 тыс. лет назад). Первоначальное место обитания первых носителей нилотского языка предположительно находилось к востоку от Нила, на территории современного Южного Судана . Протонилоты III тысячелетия до нашей эры были скотоводами , в то время как их соседи, прото- центральные суданские народы, были в основном земледельцами. [137]
Африке . Считается, что тип Нигер-Конго возник около 6000 лет назад в Западной или Центральной Его расширение могло быть связано с расширением сельского хозяйства Сахеля в период африканского неолита после высыхания Сахары в ок. 3900 год до нашей эры . [138] Экспансия банту распространила языки банту в Центральную, Восточную и Южную Африку, частично вытеснив коренное население этих регионов, включая африканских пигмеев , народ хадза и народ сан . Примерно 3000 лет назад он достиг Южной Африки.около 1700 лет назад. [139]
Некоторые данные (в том числе исследование Басби и др., проведенное в 2016 году) предполагают примесь древних и недавних миграций из Евразии в некоторые части Африки к югу от Сахары. [140] Другое исследование (Рамзи и др., 2018) также демонстрирует доказательства того, что древние евразийцы мигрировали в Африку и что евразийская примесь среди современных африканцев к югу от Сахары колеблется от 0% до 50%, в зависимости от региона и, как правило, выше на Африканском Роге и в некоторых частях Африки. в зоне Сахеля и в меньшей степени встречается в некоторых частях Западной и Южной Африки (исключая недавних иммигрантов). [141]
Индо-Тихоокеанский регион
[ редактировать ]
Первые морские миграции людей совершили австронезийские народы. [ сомнительно – обсудить ] происходящая из Тайваня, известная как « австронезийская экспансия ». [142] Используя передовые технологии парусного спорта, такие как катамараны , лодки с выносными опорами и паруса с крабовыми когтями , они построили первые морские корабли и быстро колонизировали острова Юго-Восточной Азии примерно в 3000–1500 годах до нашей эры. С Филиппин и Восточной Индонезии они колонизировали Микронезию в период с 2200 по 1000 год до нашей эры. [142] [143]
Ветвь австронезийцев достигла острова Меланезия между 1600 и 1000 годами до нашей эры, основав культуру лапита (названную в честь археологического памятника в Лапите, Новая Каледония , где впервые была обнаружена их характерная керамика). Они являются прямыми предками современных полинезийцев . Они отправились в отдаленную Океанию, достигнув Вануату , Новой Каледонии и Фиджи к 1200 г. до н. э., а также до Самоа и Тонги примерно к 900–800 гг. до н. э. Это была самая дальняя степень распространения культуры лапита. В течение примерно 1500 лет они постепенно утратили технологию гончарного дела (вероятно, из-за отсутствия залежей глины на островах), заменив ее резными деревянными и бамбуковыми сосудами. Обратные миграции из культуры лапита также объединили острова Юго-Восточной Азии в 1500 г. до н.э. и Микронезию примерно в 200 г. до н.э. Лишь в 700 году нашей эры они начали путешествовать дальше в Тихий океан, когда они колонизировали острова Кука , острова Общества и Маркизские острова . Оттуда они продолжили колонизацию Гавайи к 900 г. н.э., Рапа-Нуи к 1000 г. н.э. и Новая Зеландия к 1200 г. н.э. [143] [144] [145]
В Индийском океане австронезийцы с Борнео также колонизировали Мадагаскар и Коморские острова примерно к 500 году нашей эры. Австронезийцы остаются доминирующей этнолингвистической группой на островах Индо-Тихоокеанского региона и первыми создали морскую торговую сеть, простирающуюся на запад до Восточной Африки и Аравийского полуострова . Они ассимилировали раннего плейстоцена и раннего голоцена наземные миграции людей через Сундаленд, как папуасы и негритосы на островах Юго-Восточной Азии. [142] [143] Австронезийская экспансия была последним и наиболее масштабным событием миграции людей в эпоху неолита . [146]
Карибский бассейн
[ редактировать ]Карибский бассейн был одним из последних мест в Америке, заселенных людьми. Самые старые останки известны с Больших Антильских островов (Куба и Эспаньола) и датируются периодом между 4000 и 3500 годами до нашей эры, а сравнение технологий орудий позволяет предположить, что эти народы переселились через пролив Юкатан из Центральной Америки. Все данные свидетельствуют о том, что более поздние мигранты, начиная с 2000 г. до н.э. и далее, происходили из Южной Америки через регион Ориноко. [147] Среди потомков этих мигрантов — предки народов таино и калинаго (островной кариб). [148]
Арктика
[ редактировать ]
Самые ранние обитатели центральной и восточной Арктики Северной Америки относятся к арктической традиции малых инструментов (AST) и существовали ок. 2500 г. до н.э. AST состоял из нескольких палео-эскимосских культур, включая культуры Независимости и додорсетскую культуру. [149] [150]
Инуиты постепенно являются потомками культуры Туле , которая возникла на западе Аляски около 1000 г. н.э. и вытеснила культуру Дорсет. [151] [152]
См. также
[ редактировать ]- Список первых населенных пунктов
- Средний палеолит
- Четвертичное вымирание
- Хронология эволюции человека
- Хронология морской миграции и исследований
Примечания
[ редактировать ]- ^ На основании Шлебуша и др., «Древние геномы Южной Африки оценивают расхождение современного человека от 350 000 до 260 000 лет назад», [1] Рис. 3 ( время дивергенции H. sapiens ) и Стрингер (2012), [2] (архаичная примесь).
- ^ Архаичная примесь из различных источников известна из Европы и Азии (неандертальцы), Юго-Восточной Азии и Меланезии (денисовцы), а также из Западной и Южной Африки. Доля примеси варьируется в зависимости от региона, но во всех случаях сообщается о значении ниже 10%: в Евразии в основном оценивается в 1–4% (с высокой оценкой 3,4–7,3% по Lohse (2014). [3] ) у меланезийцев оценивается в 4–6% (Reich et al. (2010)). [4] Примесь неизвестного архаичного гоминина в популяциях охотников-собирателей Африки к югу от Сахары оценивалась в 2% (Hammer et al. (2011)). [5]
- ^ Homo erectus Soloensis , найденный на Яве , считается последним известным экземпляром H. erectus . Исследование 2011 года, которое раньше датировалось 50 000–40 000 лет назад, отодвинуло дату исчезновения H. e. soloensis самое позднее 143 000 лет назад, но, скорее всего, не позднее 550 000 лет назад. [14]
- ^ «Здесь мы сообщаем о возрасте, определенном методом термолюминесцентного датирования, нагретых огнем кремневых артефактов, полученных в результате новых раскопок на стоянке среднего каменного века в Джебель-Ирхуде, Марокко, которые напрямую связаны с недавно обнаруженными останками H. sapiens. Средний возраст относит эти артефакты и окаменелости среднего каменного века к 315–34 тысячам лет назад. Подтверждение получено с помощью пересчитанной серии урана с датой электронного спинового резонанса 286–32 тысячи лет назад для зуба нижней челюсти гоминина Ирхуда 3». [21]
- ^ Расчетное время разделения, указанное в цитируемом источнике (в тысячах лет): Человек-неандерталец: 530–690, Глубокий человек [H. sapiens]: 250–360, НКСП-СКСП: 150–190, Из Африки (ОА): 70–120. [1]
- ^ «Примерно 130 тыс. лет назад в Африке сосуществовали две отдельные группы анатомически современных людей: в целом, предки многих современных популяций хое и сан на юге и вторая центрально-восточная африканская группа, которая включает предков большинства современных популяций Расселение населения по всему миру в раннее современное время коррелирует с изменениями климата, особенно с тропическими африканскими «мегазасухами» MIS 5 (морская изотопная стадия 5, 135–75 тыс. лет назад), которые парадоксальным образом могли способствовать распространению в центральной и восточной Африке, что в конечном итоге спровоцировало расселение. В Африке людей, несущих гаплогруппу L3 ~60 тыс. лет назад, внутри гаплогруппы L0 можно различить две миграции с юга на восток. Одна, между 120 и 75 тыс. лет назад, представляет собой первое однозначное расселение современного человека на большие расстояния, обнаруженное с помощью мтДНК, и могло способствовать расселению. несколько маркеров современности. Второй, отмеченный L0d, в течение последних 20 тыс. лет назад, возможно, был ответственен за распространение южных языков с щелкающими согласными в Восточную Африку, вопреки мнению, что эти восточные примеры представляют собой реликты древнего, очень древнего языка. более широкое распространение». [36]
- ^ «Мы изучили историю ветвления пигмеев-охотников-собирателей и сельскохозяйственных популяций из Африки и оценили время разделения и поток генов между этими популяциями. Выявленная модель включала раннее расхождение предков пигмеев-охотников-собирателей и земледельческих популяций ~ 60 000 лет назад. , за которым последовал раскол предков пигмеев на западную и восточную группы пигмеев – 20 000 лет назад». [41]
- ^ Предполагается, что присутствие раннего современного человека за пределами Африки датируется еще 177 000 лет назад. [34]
- ↑ Авторы Лю (2010), похоже, признают, что этот человек недавно совершил африканское восхождение, но с примесью азиатского архаичного человека. [50] См. также Деннелл (2010). [51] Краткие комментарии на [52] и [53]
- ^ «Мы обнаружили, что население Северной Африки имеет значительный избыток производных аллелей, общих с неандертальцами, по сравнению с африканцами к югу от Сахары. Этот избыток аналогичен тому, который обнаружен у неафриканских людей, и этот факт можно интерпретировать как признак Примесь неандертальцев. Более того, генетический сигнал неандертальцев выше в популяциях с местным донеолитическим происхождением из Северной Африки. Таким образом, обнаруженная древняя примесь не связана с недавними миграциями из стран Ближнего Востока или Европы. повлияло событие смешения с неандертальцами». [108]
- ^ «Из 1420 хромосом к югу от Сахары только одна копия B006 наблюдалась в Эфиопии и пять в Буркина-Фасо, одна среди римайбе и четыре среди фулани и туарегов, кочевников-скотоводов, известных своими контактами с северным населением (дополнительная таблица S1, Дополнительный материал онлайн). B006 встречается только на северных и северо-восточных окраинах Африки к югу от Сахары, таким образом, вероятно, является результатом потока генов из неафриканского источника». [109]
- ^ Признаки, на которые влияет мутация, - это потовые железы, зубы, толщина волосяного стержня и ткань молочной железы. [114] [115]
- ^ Генетика Восточной Азии показывает ряд концентрированных аллелей, свидетельствующих о давлении отбора. В частности, это касается генов EDAR , ADH1B , ABCC1 и ALDH2 . Восточноазиатские типы ADH1B связаны с одомашниванием риса и, таким образом, могли возникнуть после ок. 11 000 лет назад. [116]
Ссылки
[ редактировать ]- ^ Jump up to: а б Шлебуш; и др. (3 ноября 2017 г.). «Древние геномы Южной Африки оценивают дивергенцию современного человека от 350 000 до 260 000 лет назад» . Наука . 358 (6363): 652–655. Бибкод : 2017Sci...358..652S . дои : 10.1126/science.aao6266 . ПМИД 28971970 .
- ^ Стрингер, К. (2012). «Что делает современного человека» . Природа . 485 (7396): 33–35. Бибкод : 2012Natur.485...33S . дои : 10.1038/485033а . ПМИД 22552077 . S2CID 4420496 .
- ^ Лозе, К; Франц, LAF (2014). «Примесь неандертальцев в Евразии подтверждена анализом максимального правдоподобия трех геномов» . Генетика . 196 (4): 1241–1251. дои : 10.1534/genetics.114.162396 . ПМЦ 3982695 . ПМИД 24532731 .
- ^ Райх, Д; Зеленый, RE; Кирхер, М; Краузе, Дж; Паттерсон, Н.; Дюран, EY; Виола, Б; Бриггс, AW; Стензель, Ю; Джонсон, Польша; Маричич, Т; Хорошо, Дж. М.; Маркес-Боне, Т; Алкан, К; Фу, Кью; Маллик, С; Ли, Х; Мейер, М; Эйхлер, Э.Э.; Стоункинг, М; Ричардс, М; Таламо, С; Шунков, М.В.; Деревянко А.П.; Хаблин, Джей-Джей; Келсо, Дж; Слаткин М; Паабо, С (2010). «Генетическая история архаической группы гомининов из Денисовой пещеры в Сибири» . Природа . 468 (7327): 1053–1060. Бибкод : 2010Natur.468.1053R . дои : 10.1038/nature09710 . ПМК 4306417 . ПМИД 21179161 .
- ^ Jump up to: а б Хаммер, МФ; Вернер, А.Е.; Мендес, Флорида; Уоткинс, Дж. К.; Уолл, Джей Ди (2011). «Генетические доказательства архаической примеси в Африке» . Труды Национальной академии наук . 108 (37): 15123–15128. Бибкод : 2011PNAS..10815123H . дои : 10.1073/pnas.1109300108 . ПМК 3174671 . ПМИД 21896735 .
- ^ Чжу, Чжаоюй, Робин; Ву, И; Ян, Шися; Хоу, Ямей; Хан, Цзянвэй, Тинпин (2018). Китайского Лёссового плато примерно 2,1 миллиона лет назад». Nature . 559 (7715): 608–612. Bibcode : 2018Natur.559..608Z . doi : /s41586-018-0299-4 . PMID 29995848. 10.1038 S2CID 49670311 .
- ^ Флигл, Дж. Г.; Ши, Джей-Джей; Грин, FE; Баден, Алабама; Лики, Р.Э., ред. (2010). Из Африки I: Первая колонизация Евразии гомининами . Палеобиология и палеоантропология позвоночных. Спрингер. дои : 10.1007/978-90-481-9036-2 . ISBN 978-9048190355 .
- ^ Гарсия, Т.; Феро, Ж.; Фальгер, К.; де Ламли, Х.; Перрену, К.; Лордкипанидзе, Д. (2010). «Самые ранние человеческие останки в Евразии: новая датировка 40Ar/39Ar уровней гоминидов в Дманиси, Грузия». Четвертичная геохронология . 5 (4): 443–451. Бибкод : 2010QuGeo...5..443G . дои : 10.1016/j.quageo.2009.09.012 .
- ^ Jump up to: а б Чжу, Чжаоюй, Робин Деннелл, Вэйвэнь Хуан, И Ву, Шифан Цю, Шися Ян, Чжиго Рао и др. 2018. «Заселение Китайского Лёссового плато гомининами примерно 2,1 миллиона лет назад». Nature 559 (7715): 608. –612. два : 10.1038/s41586-018-0299-4 .
- ^ Р. Чжу и др. (2004), Новые свидетельства самого раннего присутствия человека в высоких северных широтах Северо-Восточной Азии .
- ^ Чжу, Рисян; Ань, Чжишэн; Потт, Ричард; Хоффман, Кеннет А. (июнь 2003 г.). «Магнитостратиграфическое датирование древних людей в Китае» (PDF) . Обзоры наук о Земле . 61 (3–4): 191–361. Бибкод : 2003ESRv...61..191A . дои : 10.1016/S0012-8252(02)00110-1 . Архивировано из оригинала (PDF) 24 июля 2011 года.
- ^ Хопкин М. (26 марта 2008 г.). «Ископаемая находка — самая старая европейская находка» . Природа . Новости природы. дои : 10.1038/news.2008.691 .
- ^ Беднарик Р.Г. (2003). «Мореплавание в плейстоцене». Кембриджский археологический журнал . 13 (1): 41–66. дои : 10.1017/S0959774303000039 . S2CID 162678917 .
Краткое изложение ScienceNews. Архивировано 22 апреля 2008 г. в Wayback Machine. - ^ Индриати, Э; Свишер CC III; проказа С; Куинн Р.Л.; Суриянто РА (2011). «Эпоха 20-метровой одиночной речной террасы, Ява, Индонезия и выживание человека прямоходящего в Азии » ПЛОС ОДИН 6 (6): e21562. Бибкод : 2011PLoSO...621562I . дои : 10.1371/journal.pone.0021562 . ПМК 3126814 . ПМИД 21738710 .
- ^ Каллауэй, Юэн (12 мая 2011 г.). «Арктическое убежище для неандертальцев?» . Nature.com. Архивировано из оригинала 28 июля 2019 года . Проверено 8 мая 2019 г.
- ^ Сюй, Д.; и др. (2017). «Архаическая интрогрессия гомининов в Африке способствует функциональной генетической вариации слюны MUC7» . Молекулярная биология и эволюция . 34 (10): 2704–2715. дои : 10.1093/molbev/msx206 . ПМК 5850612 . ПМИД 28957509 .
- ^ Каллауэй, Э. (26 июля 2012 г.). «Геномы охотников-собирателей — кладезь генетического разнообразия» . Природа . дои : 10.1038/nature.2012.11076 . S2CID 87081207 .
- ^ Лашанс, Дж.; Верно, Б.; Элберс, CC; Ферверда, Б.; Фромант, А.; Бодо, Дж. М.; и др. (2012). «Эволюционная история и адаптация на основе полногеномных последовательностей разнообразных африканских охотников-собирателей» . Клетка . 150 (3): 457–469. дои : 10.1016/j.cell.2012.07.009 . ПМК 3426505 . ПМИД 22840920 .
- ^ Диркс, Пол ХГМ; Робертс, Эрик М.; Гильберт-Вольф, Ханна; Крамерс, Ян Д.; Хоукс, Джон; Доссето, Энтони; Дюваль, Мэтью; Эллиотт, ВМФ; Эванс, Мэри; Грин, Райнер; Хеллстром, Джон; Херрис, Энди И.Р.; Жоаннес-Бойо, Фокс; Махубела, Тебого В.; Плачек, Криста Дж.; Роббинс, Джесси; Спэндлер, Карл; Виерсма, Йелле; Вудхед, Джон; Бергер, Ли Р. (9 мая 2017 г.). «Возраст Homo naledi и связанных с ним отложений» (PDF ) электронная жизнь 6 . doi : 10.7554/eLife.24231 . ПМЦ 5423772 . ПМИД 28483040 .
- ^ Jump up to: а б с д и ж Лопес, Сайоа; ван Дорп, Люси; Хеллентал, Гаррет (2016). «Расселение людей из Африки: длительные дебаты» . Эволюционная биоинформатика . 11с2 (Приложение 2): 57–68. дои : 10.4137/EBO.S33489 . ISSN 1176-9343 . ПМЦ 4844272 . ПМИД 27127403 .
- ^ Рихтер, Дэвид; и др. (8 июня 2017 г.). «Возраст окаменелостей гомининов из Джебель-Ирхуда, Марокко, и истоки среднего каменного века». Природа . 546 (7657): 293–296. Бибкод : 2017Natur.546..293R . дои : 10.1038/nature22335 . ПМИД 28593967 . S2CID 205255853 .
- ^ Смит Т.М., Тафоро П., Рид Дж. и др. (апрель 2007 г.). «Самые ранние свидетельства современной истории жизни человека у раннего Homo Sapiens в Северной Африке» . Труды Национальной академии наук Соединенных Штатов Америки . 104 (15): 6128–6133. Бибкод : 2007PNAS..104.6128S . дои : 10.1073/pnas.0700747104 . ПМК 1828706 . ПМИД 17372199 .
- ^ Стрингер, К. (2016). «Происхождение и эволюция человека разумного» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 371 (1698): 20150237. doi : 10.1098/rstb.2015.0237 . ПМЦ 4920294 . ПМИД 27298468 .
- ^ Образец, Ян (7 июня 2017 г.). «Самые древние кости Homo sapiens , когда-либо найденные, потрясают основы человеческой истории» . Хранитель . Проверено 7 июня 2017 г.
- ^ Юблин, Жан-Жак; Бен-Нсер, Абделуахед; Бейли, Шара Э.; Фридлайн, Сара Э.; Нойбауэр, Саймон; Скиннер, Мэтью М.; Бергманн, Мать; Ле Кабек, Аделина; Беназир, Стивен; Харвати, Екатерина; Гунц, Филипп (2017). «Новые окаменелости из Джебель-Ирхуда, Марокко, и панафриканское происхождение Homo sapiens » (PDF) . Природа 546 (7657): 289–292. Бибкод : 2017Nature.546..289H . дои : 10.1038/nature22336 . ПМИД 28593953 . S2CID 256771372 .
- ^ Скерри, Элеонора М.Л.; Томас, Марк Г.; Маника, Андреа; Гунц, Филипп; Сток, Джей Т.; Стрингер, Крис; Гроув, Мэтт; Гроукатт, Хью С.; Тиммерманн, Аксель ; Райтмайр, Дж. Филип; д'Эррико, Франческо (2018). «Эволюционировал ли наш вид в разделенных популяциях по всей Африке, и почему это имеет значение?» . Тенденции в экологии и эволюции . 33 (8): 582–594. Бибкод : 2018TEcoE..33..582S . дои : 10.1016/j.tree.2018.05.005 . ПМК 6092560 . ПМИД 30007846 .
- ^ Макдугалл, И.; Браун, Х.; Флигл, Г. (февраль 2005 г.). «Стратиграфическое размещение и возраст современных людей из Кибиша, Эфиопия» (PDF) . Природа . 433 (7027): 733–736. Бибкод : 2005Natur.433..733M . дои : 10.1038/nature03258 . ISSN 0028-0836 . ПМИД 15716951 . S2CID 1454595 .
- ^ Циммер, Карл (10 сентября 2019 г.). «Ученые нашли череп предка человечества на компьютере. Сравнивая окаменелости и компьютерную томографию, исследователи говорят, что реконструировали череп последнего общего предка современных людей» . Нью-Йорк Таймс . Проверено 31 декабря 2019 г.
- ^ Мунье, Орельен; Лар, Марта (2019). «Расшифровка разнообразия гомининов Африки позднего среднего плейстоцена и происхождения нашего вида» . Природные коммуникации . 10 (1): 3406. Бибкод : 2019NatCo..10.3406M . дои : 10.1038/s41467-019-11213-w . ПМЦ 6736881 . ПМИД 31506422 .
- ^ Циммер, Карл (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить историю предыстории человечества» . Нью-Йорк Таймс . Проверено 31 декабря 2019 г.
- ^ Персонал (10 июля 2019 г.). « Самые старые останки за пределами Африки обнуляют часы миграции людей» . Физика.орг . Проверено 10 июля 2019 г.
- ^ Делсон, Эрик (10 июля 2019 г.). «Раннее расселение современных людей из Африки в Грецию. Анализ двух окаменелостей из греческой пещеры пролил свет на ранних гоминидов в Евразии. Одно ископаемое — самый ранний известный образец Homo sapiens, найденный за пределами Африки; другое — неандерталец, живший 40 000 лет спустя» . Природа . 571 (7766): 487–488. дои : 10.1038/d41586-019-02075-9 . ПМИД 31337897 .
- ^ Харвати, Катерина; и др. (10 июля 2019 г.). «Окаменелости пещеры Апидима представляют собой самые ранние свидетельства существования Homo sapiens в Евразии» . Природа . 571 (7766): 500–504. дои : 10.1038/s41586-019-1376-z . ПМИД 31292546 . S2CID 195873640 .
- ^ Jump up to: а б Гершковитц и др. (26 января 2018 г.). «Самые ранние современные люди за пределами Африки» . Наука . 359 (6374): 456–459. Бибкод : 2018Sci...359..456H . дои : 10.1126/science.aap8369 . hdl : 10072/372670 . ПМИД 29371468 .
- ^ Jump up to: а б Шерри, Элеонора (2017). «Археология каменного века Западной Африки» . Африканская история . doi : 10.1093/acrefore/9780190277734.013.137 . ISBN 978-0190277734 .
- ^ Рито, Т; Ричардс, МБ; Фернандес, В; Альшамали, Ф; Черный, В; Перейра, Л; Соарес, П. (2013). «Первое расселение современного человека по Африке» . ПЛОС ОДИН . 8 (11): е80031. Бибкод : 2013PLoSO...880031R . дои : 10.1371/journal.pone.0080031 . ПМЦ 3827445 . ПМИД 24236171 .
- ^ Джарвис Дж.П., Шейнфельдт Л.Б., Сой С., Ламберт С., Омберг Л., Ферверда Б. и др. (2012). «Модели происхождения, признаки естественного отбора и генетическая связь с ростом западноафриканских пигмеев» . ПЛОС Генетика . 8 (4): e1002641. дои : 10.1371/journal.pgen.1002641 . ПМК 3343053 . ПМИД 22570615 .
- ^ Лопес Эрраес Д., Боше М., Тан К., Теунерт С., Пугач И., Ли Дж. и др. (2009). «Генетическая изменчивость и недавний положительный отбор в человеческих популяциях по всему миру: данные почти 1 миллиона SNP» . ПЛОС ОДИН . 4 (11): е7888. Бибкод : 2009PLoSO...4.7888L . дои : 10.1371/journal.pone.0007888 . ПМЦ 2775638 . ПМИД 19924308 .
- ^ Тишкофф С.А., Рид Ф.А., Фридлендер Ф.Р., Эрет С., Ранчиаро А., Фромент А., Хирбо Дж.Б., Авомойи А.А., Бодо Дж.М., Думбо О., Ибрагим М., Джума А.Т., Котце М.Дж., Лема Г., Мур Дж.Х., Мортенсен Х., Ньямбо Т.Б. , Омар С.А., Пауэлл К., Преториус Г.С., Смит М.В., Тера М.А., Вамбебе С., Вебер Дж.Л., Уильямс С.М. (2009). «Генетическая структура и история африканцев и афроамериканцев» . Наука . 324 (5930): 1035–1044. Бибкод : 2009Sci...324.1035T . дои : 10.1126/science.1172257 . ПМЦ 2947357 . ПМИД 19407144 . ( Дополнительные данные )
- ^ Кинтана-Мурси и др. (2008). «Материнские следы глубокого общего происхождения и асимметричного потока генов между пигмеями-охотниками-собирателями и фермерами, говорящими на языке банту» . ПНАС . 105 (5): 1596–1601. Бибкод : 2008PNAS..105.1596Q . дои : 10.1073/pnas.0711467105 . ПМК 2234190 . ПМИД 18216239 . инжир. 3 .
- ^ Патен Э, Лаваль Дж, Баррейро ЛБ, Салас А, Семино О, Сантакьяра-Бенерекетти С, Кидд К.К., Кидд Дж.Р., Ван дер Вин Л., Хомберт Дж.М., Гессен А., Фромант А., Бахучет С., Хейер Е., Кинтана-Мурси Л. (2009). «Выводы о демографической истории африканских фермеров и пигмеев-охотников-собирателей с использованием набора данных многолокусного повторного секвенирования» . ПЛОС Генетика . 5 (4): e1000448. дои : 10.1371/journal.pgen.1000448 . ПМЦ 2661362 . ПМИД 19360089 .
- ^ «Ученые обнаружили древнейшие из известных окаменелостей современного человека за пределами Африки: анализ окаменелостей показывает, что Homo sapiens покинул Африку как минимум на 50 000 лет раньше, чем считалось ранее» . ScienceDaily . Проверено 27 января 2018 г.
- ^ Гош, Паллаб (2018). «Современные люди покинули Африку гораздо раньше» . Новости Би-би-си . Проверено 27 января 2018 г.
- ^ Холен, Стивен Р. (2017). «Археологический памятник возрастом 130 000 лет в южной Калифорнии, США». Природа . 544 (7651): 479–483. Бибкод : 2017Natur.544..479H . дои : 10.1038/nature22065 . ПМИД 28447646 . S2CID 205255425 .
- ^ «Следы человека возрастом 120 000 лет найдены в Саудовской Аравии» . физ.орг . Проверено 9 октября 2020 г.
- ^ Стюарт, Мэтью; Кларк-Уилсон, Ричард; Бриз, Пол С.; Янулис, Клинт; Кэнди, Ян; Армитидж, Саймон Дж.; Райвс, Дэвид Б.; Луис, Жюльен; Дюваль, Матье; Прайс, Гилберт Дж.; Катбертсон, Патрик; Берналь, Марко А.; Дрейк, Ник А.; Альшарех, Абдулла М.; Захрани, Бадр; Аль-Омари, Абдулазиз; Робертс, Патрик; Гроукатт, Хью С.; Петралья, Майкл Д. (1 сентября 2020 г.). «Следы человека дают представление о последней межледниковой экологии во внутренних районах Аравии» . Достижения науки . 6 (38): eaba8940. Бибкод : 2020SciA....6.8940S . дои : 10.1126/sciadv.aba8940 . ISSN 2375-2548 . ПМЦ 7500939 . ПМИД 32948582 .
- ^ Лоулер, Эндрю (2011). «Выехали ли современные люди из Африки через Аравию?». Наука . 331 (6016): 387. Бибкод : 2011Sci...331..387L . дои : 10.1126/science.331.6016.387 . ПМИД 21273459 .
- ^ «След «каменных хлебных крошек» раскрывает личность одной из первых групп людей, покинувших Африку» . ScienceDaily. 1 декабря 2011 года . Проверено 8 мая 2019 г.
- ^ Бауэр, Брюс (27 января 2011 г.). «Намеки на более ранний выход человека из Африки» . Новости науки . Проверено 8 мая 2019 г.
- ^ Лю, Ву; и др. (2010). «Человеческие останки из Жирендуна, Южный Китай, и появление современного человека в Восточной Азии» . Труды Национальной академии наук . 107 (45): 19201–19206. Бибкод : 2010PNAS..10719201L . дои : 10.1073/pnas.1014386107 . ПМК 2984215 . ПМИД 20974952 .
- ^ Деннелл, Робин (2010). «Две интерпретации нижней челюсти Жирендуна, описанные Лю и его коллегами». Природа . 468 (7323): 512–513. дои : 10.1038/468512а . ПМИД 21107416 . S2CID 205060486 .
- ^ Современные люди появились намного раньше, чем считалось ранее, предполагают окаменелости из Китая, ScienceDaily (25 октября 2010 г.) https://www.sciencedaily.com/releases/2010/10/101025172924.htm
- ^ Найден самый старый современный человек за пределами Африки
- ^ Шена, Гуаньцзюнь; и др. (2002). «Датирование серии U стоянки гоминидов Люцзян в Гуанси, Южный Китай». Журнал эволюции человека . 43 (6): 817–829. Бибкод : 2002JHumE..43..817S . дои : 10.1006/jhev.2002.0601 . ПМИД 12473485 .
- ^ «Окаменелости Лунадонга подтверждают теорию более раннего расселения современного человека» . Гавайский университет в Маноа. 18 сентября 2014 года . Проверено 8 мая 2019 г.
- ^ Каллауэй, Юэн (2015). «Зубы из Китая свидетельствуют о раннем пути человека из Африки» . Природа . дои : 10.1038/nature.2015.18566 . S2CID 181399291 . Проверено 23 октября 2015 г.
- ^ Балтер, Майкл (2010). «Два мнения о влиянии Тобы». Наука . 327 (5970): 1187–1188. Бибкод : 2010Sci...327.1187B . дои : 10.1126/science.327.5970.1187-a . ПМИД 20203021 .
- ^ Jump up to: а б Пост С., Рено Г., Миттник М., Друкер Д.Г., Ружье Х., Купийяр С. и др. (2016). «Митохондриальные геномы плейстоцена предполагают единое крупное расселение неафриканцев и смену населения в конце ледникового периода в Европе». Современная биология . 26 (6): 827–833. Бибкод : 2016CBio...26..827P . дои : 10.1016/j.cub.2016.01.037 . hdl : 2440/114930 . ПМИД 26853362 . S2CID 140098861 .
- ^ Кармин М., Сааг Л., Висенте М., Уилсон Сейрес М.А., Ярве М., Талас У.Г. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосомы совпадает с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Jump up to: а б Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арсьеро Э., Хуанмин Ю., Томас М.Г., Сюэ Ю., Тайлер-Смит С. (июнь 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- ^ Животовский; Розенберг, Н.А.; Фельдман, М.В.; и др. (2003). «Особенности эволюции и расширения современного человека, выявленные на основе полногеномных микросателлитных маркеров» . Американский журнал генетики человека . 72 (5): 1171–1186. дои : 10.1086/375120 . ПМК 1180270 . ПМИД 12690579 .
- ^ Стикс, Гэри (2008). «История миграции людей: исследование ДНК прослеживает происхождение человека на разных континентах» . Научный американец . Проверено 14 июня 2011 г.
- ^ Армитидж, Саймон Дж.; Джасим, Сабах А.; Маркс, Энтони Э.; Паркер, Адриан Г.; Усик, Виталий И.; Урпманн, Ханс-Петер (2011). «Намеки на более ранний исход человека из Африки» . Наука . 331 (6016): 453–456. Бибкод : 2011Sci...331..453A . дои : 10.1126/science.1199113 . ПМИД 21273486 . S2CID 20296624 . Проверено 1 мая 2011 г.
- ^ Вай С., Сарно С., Лари М., Луизелли Д., Манци Г., Галлинаро М., Матаич С., Хюбнер А., Моди А., Пилли Е., Тафури М.А., Карамелли Д., ди Лерния С. (март 2019 г.). «Предковая митохондриальная линия N из неолитической «зеленой» Сахары» . Научные отчеты . 9 (1): 3530. Бибкод : 2019НатСР...9.3530В . дои : 10.1038/s41598-019-39802-1 . ПМК 6401177 . ПМИД 30837540 .
- ^ Деннелл, Робин ; Петралья, Майкл Д. (2012). «Распространение Homo sapiens по южной Азии: насколько рано, как часто и насколько сложно?». Четвертичные научные обзоры . 47 : 15–22. Бибкод : 2012QSRv...47...15D . doi : 10.1016/j.quascirev.2012.05.002 .
- ^ Прюфер, Кей; и др. (2013). «Полная последовательность генома неандертальца с Горного Алтая» . Природа . 505 (7481): 43–49. Бибкод : 2014Natur.505...43P . дои : 10.1038/nature12886 . ПМК 4031459 . ПМИД 24352235 .
- ^ Jump up to: а б с Боулер, Джеймс М.; и др. (2003). «Новый век человеческой деятельности и изменение климата на озере Мунго, Австралия». Природа . 421 (6925): 837–840. Бибкод : 2003Natur.421..837B . дои : 10.1038/nature01383 . ПМИД 12594511 . S2CID 4365526 .
- ^ Вуд Р. (2 сентября 2017 г.). «Комментарии к хронологии Маджедбебе». Австралийская археология . 83 (3): 172–174. дои : 10.1080/03122417.2017.1408545 . ISSN 0312-2417 . S2CID 148777016 .
- ^ О'Коннелл Дж. Ф., Аллен Дж., Уильямс М.А., Уильямс А.Н., Терни К.С., Спунер Н.А. и др. (август 2018 г.). «Homo sapiens впервые достигает Юго-Восточной Азии и Сахула?» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (34): 8482–8490. Бибкод : 2018PNAS..115.8482O . дои : 10.1073/pnas.1808385115 . ПМК 6112744 . ПМИД 30082377 .
- ^ Каллауэй, Юэн (22 сентября 2011 г.), «Первое секвенирование генома аборигенов» , Nature , doi : 10.1038/news.2011.551
- ^ Рейх; и др. (2011), «Примесь Денисовой и первое расселение современного человека в Юго-Восточную Азию и Океанию», Американский журнал генетики человека , 89 (4): 516–528, doi : 10.1016/j.ajhg.2011.09.005 , PMC 3188841 , PMID 21944045
- ^ Чой, Чарльз (22 сентября 2011 г.). «Ныне вымерший родственник занимался сексом с людьми повсюду» . ЖиваяНаука .
- ^ Купер А.; Стрингер CB (2013). «Пересекли ли денисовцы линию Уоллеса». Наука . 342 (6156): 321–333. Бибкод : 2013Sci...342..321C . дои : 10.1126/science.1244869 . ПМИД 24136958 . S2CID 206551893 .
- ^ Саллех, Анна (18 октября 2013 г.). «Люди встречались с древними денисовскими родственниками за пределами линии Уоллеса» . Азбука науки.
- ^ Первые моряки - Архив журнала археологии . Archive.archaeology.org. Проверено 16 ноября 2013 г.
- ^ «Карты уровня моря плейстоцена» . Fieldmuseum.org. Архивировано из оригинала 17 марта 2009 года . Проверено 23 сентября 2010 г.
- ^ Расмуссен, М; и др. (октябрь 2011 г.). «Геном австралийских аборигенов показывает отдельные расселения людей в Азии» . Наука . 334 (6052): 94–98. Бибкод : 2011Sci...334...94R . дои : 10.1126/science.1211177 . ПМЦ 3991479 . ПМИД 21940856 .
- ^ Худжашов Г., Кивисилд Т., Андерхилл П.А. и др. (май 2007 г.). «Выявление доисторического поселения Австралии с помощью Y-хромосомы и анализа мтДНК» . Proc Natl Acad Sci США . 104 (21): 8726–8730. Бибкод : 2007PNAS..104.8726H . дои : 10.1073/pnas.0702928104 . ПМК 1885570 . ПМИД 17496137 .
- ^ Уэйд, Николас (8 мая 2007 г.). «Анализ ДНК: ключ к разгадке единой австралийской миграции» . Нью-Йорк Таймс . Австралия . Проверено 31 декабря 2019 г.
- ^ Кларксон, Крис; Смит, Майк; Марвик, Бен; Фуллагар, Ричард; Уоллис, Линли А.; Фолкнер, Патрик; Манне, Тиина; Хейс, Элспет; Робертс, Ричард Г.; Джейкобс, Зенобия; Кара, Ксавьер; Лоу, Келси М.; Мэтьюз, Жаклин; Флорин, С. Анна (июнь 2015 г.). «Археология, хронология и стратиграфия Маджедбебе (Малакунанджа II): место раннего заселения» . Журнал эволюции человека . 83 : 46–64. Бибкод : 2015JHumE..83...46C . дои : 10.1016/jhevol.2015.03.014 . hdl : 1773/33254 . ПМИД 25957653 . S2CID 19824757 .
- ^ Кларксон, Крис; Джейкобс, Зенобия; Марвик, Бен; Фуллагар, Ричард; Уоллис, Линли; Смит, Майк; Робертс, Ричард Г.; Хейс, Элспет; Лоу, Келси; Кара, Ксавьер; Флорин, С. Анна; Макнил, Джессика; Кокс, Делит; Арнольд, Ли Дж.; Хуа, Цюань; Хантли, Джиллиан; Брэнд, Хелен Э.А.; Манне, Тиина; Фэйрберн, Эндрю; Шульмейстер, Джеймс; Лайл, Линдси; Салинас, Макия; Пейдж, Мара; Коннелл, Кейт; Пак, Гаёнг; Норман, Касих; Мерфи, Тесса; Пардо, Колин (19 июля 2017 г.). «Человеческая оккупация северной Австралии 65 000 лет назад» (PDF) . Природа . 547 (7663): 306–310. Бибкод : 2017Natur.547..306C . дои : 10.1038/nature22968 . hdl : 2440/107043 . ПМИД 28726833 . S2CID 205257212 .
- ^ Фланнери, Тим (2002), «Пожиратели будущего: экологическая история австралийских земель и людей» (Grove Press)
- ^ Мелларс, Пол (11 августа 2006 г.). «Путешествие на Восток: новые генетические и археологические перспективы современной человеческой колонизации Евразии». Наука . 313 (5788): 796–800. Бибкод : 2006Sci...313..796M . дои : 10.1126/science.1128402 . ПМИД 16902130 . S2CID 24631308 .
- ^ «Ископаемые зубы привели людей в Европу раньше, чем предполагалось» . Нью-Йорк Таймс . 2 ноября 2011 года . Проверено 31 декабря 2019 г.
- ^ Jump up to: а б Шанкарараман, С.; Паттерсон, Н.; Ли, Х.; Паабо, С.; Райх, Д; Эйки, Дж. М. (2012). «Дата скрещивания неандертальцев и современных людей» . ПЛОС Генетика . 8 (10): e1002947. arXiv : 1208.2238 . Бибкод : 2012arXiv1208.2238S . дои : 10.1371/journal.pgen.1002947 . ПМЦ 3464203 . ПМИД 23055938 .
- ^ Харпендинг, Генри; Кокран, Грегори (2009). 10 000-летний взрыв: как цивилизация ускорила эволюцию человека . Основные книги. п. 214. ИСБН 978-0465002214 . Проверено 1 декабря 2015 г.
- ^ Эмброуз, Стэнли (1998). «Узкие места человеческого населения позднего плейстоцена, вулканическая зима и дифференциация современных людей». Журнал эволюции человека . 34 (6): 623–651. Бибкод : 1998JHumE..34..623A . дои : 10.1006/jhev.1998.0219 . ПМИД 9650103 . S2CID 33122717 .
- ^ Оппенгеймер, Стивен «Из Эдема: Заселение мира» (Робинсон; издание New Ed (1 марта 2012 г.))
- ^ Jump up to: а б Павлов П; Свендсен, Дж.И.; Индрелид, С. (2001). «Присутствие человека в Европейской Арктике почти 40 000 лет назад». Природа . 413 (6851): 64–67. Бибкод : 2001Natur.413...64P . дои : 10.1038/35092552 . ПМИД 11544525 . S2CID 1986562 .
- ^ «Атлас человеческого пути: 45–40 000» . Генографический проект . Национальное географическое общество. 1996–2010 гг. Архивировано из оригинала 14 января 2013 года . Проверено 8 мая 2019 г.
- ^ Смит, Джефф М.; Рубенс, Карен; Завала, Елена Ирина; Синет-Матиот, Вирджиния; Фьюласс, Хелен; Педерзани, Сара; Жауэн, Клервия; Милопотамитаки, Доротея; Бриттон, Кейт; Ружье, Хелен; Штальшмидт, Марайке; Мейер, Матиас; Меллер, Харальд; Дитль, Хольгер; Оршидт, Джордж; Краузе, Джон; Шулер, Тим; Макферрон, Шеннон П.; Вайс, Марсель; Юблин, Жан-Жак; Велкер, Фридо (31 января 2024 г.). «Экология, существование и диета примерно 45 000-летнего человека разумного в Ильзенхёле в Ранисе, Германия » Экология и эволюция природы . 8 (3): 564–577. Бибкод : 2024NatEE...8..564S . дои : 10.1038/ s41559-023-02303-6 ПМЦ 10927544 . ПМИД 38297138 .
- ^ Бенацци, С.; Дука, К.; Форнаи, К.; Бауэр, CC; Куллмер, О.; Свобода, Дж. Ф.; Пап, И.; Маллени, Ф.; Бэйл, П.; Кокерель, М.; Кондеми, С.; Рончителли, А.; Харвати, К.; Вебер, GW (2011). «Раннее расселение современных людей в Европе и последствия для поведения неандертальцев». Природа . 479 (7374): 525–528. Бибкод : 2011Natur.479..525B . дои : 10.1038/nature10617 . ПМИД 22048311 . S2CID 205226924 .
- ^ Хайэм, Т.; Комптон, Т.; Стрингер, К.; Якоби, Р.; Шапиро, Б.; Тринкаус, Э.; Чендлер, Б.; Грёнинг, Ф.; Коллинз, К.; Хиллсон, С.; о'Хиггинс, П.; Фицджеральд, К.; Фэган, М. (2011). «Самые ранние свидетельства существования анатомически современных людей в северо-западной Европе». Природа . 479 (7374): 521–524. Бибкод : 2011Natur.479..521H . дои : 10.1038/nature10484 . ПМИД 22048314 . S2CID 4374023 .
- ^ «Мамонтовая Курья: загадочная стоянка палеолита возрастом около 40 000 лет в Российской Арктике» (PDF) . Архивировано из оригинала (PDF) 14 августа 2015 года . Проверено 17 июня 2015 г.
- ^ Jump up to: а б Хоффекер, Дж. (2006). Предыстория Севера: населенные пункты высоких широт . Нью-Джерси: Издательство Университета Рутгерса. п. 101 .
- ^ Хоффекер, Джон Ф. (2002). Пустынные ландшафты: поселения ледникового периода в Восточной Европе . Нью-Брансуик: Издательство Университета Рутгерса. стр. 158–162, 217–233.
- ^ Кэтрин Э. Фицсиммонс и др., Кампанское извержение игнимбрита: новые данные о рассеянии вулканического пепла и его потенциальном влиянии на эволюцию человека, 2013 https://doi.org/10.1371/journal.pone.0065839
- ^ Джаччо, Б. и др., Высокоточное датирование кампанского игнимбрита (Y-5) с помощью 14C и 40Ar/39Ar согласовывает временные рамки климатических и культурных процессов на уровне 40 тыс. лет назад. Sci Rep 7, 45940 (2017). https://doi.org/10.1038/srep45940
- ^ Хайдиньяк, М. и др., Первые люди верхнего палеолита в Европе имели недавнее неандертальское происхождение. Природа 592, 253–257 (2021). https://doi.org/10.1038/s41586-021-03335-3
- ^ Беннетт, Э.А. и др., Последовательности генома современных людей возрастом от 36 000 до 37 000 лет на Буран-Кая III в Крыму. Нат Эколь Эвол (2023). https://doi.org/10.1038/s41559-023-02211-9
- ^ Карамелли, Д; Лалуэса-Фокс, Ц; Вернези, К; Лари, М; Казоли, А; Маллени, Ф; Кьярелли, Б; Дюпанлуп, я; Бертранпети, Дж; Барбуджани, Дж; Берторель, Дж. (май 2003 г.). «Доказательства генетического разрыва между неандертальцами и анатомически современными европейцами возрастом 24 000 лет» . ПНАС . 100 (11): 6593–6597. Бибкод : 2003PNAS..100.6593C . дои : 10.1073/pnas.1130343100 . ISSN 0027-8424 . ПМК 164492 . ПМИД 12743370 .
- ^ Куррат М., Экскофье Л. (2004). «Современные люди не смешивались с неандертальцами во время их распространения в Европу» . ПЛОС Биология . 2 (12): е421. дои : 10.1371/journal.pbio.0020421 . ПМЦ 532389 . ПМИД 15562317 .
- ^ Мака-Мейер Н., Гонсалес А.М., Ларруга Х.М., Флорес С., Кабрера В.М. (2001). «Основные геномные митохондриальные линии определяют раннюю экспансию человека» . БМК Генет . 2 (1): 13. дои : 10.1186/1471-2156-2-13 . ПМК 55343 . ПМИД 11553319 .
- ^ Куррат М., Экскофье Л. (декабрь 2004 г.). «Современные люди не смешивались с неандертальцами во время их экспансии в Европу» . ПЛОС Биология . 2 (12): е421. дои : 10.1371/journal.pbio.0020421 . ПМЦ 532389 . ПМИД 15562317 .
- ^ Хайдиньяк и др. Первые люди верхнего палеолита в Европе имели недавнее неандертальское происхождение. Природа 592, 253–257 (2021). https://doi.org/10.1038/s41586-021-03335-3
- ^ Мейер, М.; Кирхер, М.; Гансоге, Монтана; Ли, Х.; Расимо, Ф.; Маллик, С.; и др. (2012). «Последовательность генома с высоким уровнем покрытия архаичного денисовского человека» . Наука . 338 (6104): 222–226. Бибкод : 2012Sci...338..222M . дои : 10.1126/science.1224344 . ПМК 3617501 . ПМИД 22936568 .
- ^ Уолл, Джей Ди; Ян, Массачусетс; Джей, Ф.; Ким, СК; Дюран, EY; Стевисон, Лос-Анджелес; и др. (2013). «Более высокий уровень неандертальского происхождения у жителей Восточной Азии, чем у европейцев» . Генетика . 194 (1): 199–209. дои : 10.1534/genetics.112.148213 . ПМЦ 3632468 . ПМИД 23410836 .
- ^ Санчес-Кинто, Ф.; Владелец магазина, Л.Р.; Сивит, С.; Аренас, Дж.; Авила-Аркос, MC; Бустаманте, CD; и др. (2012). «Население Северной Африки несет в себе признак смешения с неандертальцами» . ПЛОС ОДИН . 7 (10): е47765. Бибкод : 2012PLoSO...747765S . дои : 10.1371/journal.pone.0047765 . ПМЦ 3474783 . ПМИД 23082212 .
- ^ Йотова, Ваня; и др. (2011). «Х-сцепленный гаплотип неандертальского происхождения присутствует среди всех неафриканских популяций» . Молекулярная биология и эволюция . 28 (7): 1957–1962. дои : 10.1093/molbev/msr024 . ПМИД 21266489 .
- ^ Балтер, М. (25 октября 2013 г.). «Древняя ДНК связывает коренных американцев с Европой» . Наука . 342 (6157): 409–410. Бибкод : 2013Sci...342..409B . дои : 10.1126/science.342.6157.409 . ПМИД 24159019 .
- ^ Ян; и др. (2017). «40 000-летний человек из Азии дает представление о ранней структуре населения Евразии» . Современная биология . 27 (20): 3202–3208. Бибкод : 2017CBio...27E3202Y . дои : 10.1016/j.cub.2017.09.030 . ПМК 6592271 . ПМИД 29033327 .
- ^ Дин, К.; Ху, Ю.; Сюй, С.; Ван, Дж.; Джин, Л. (2014) [Онлайн, 2013]. «Неандертальская интрогрессия на хромосоме 3p21.31 находилась под действием положительного естественного отбора у жителей Восточной Азии» . Молекулярная биология и эволюция . 31 (3): 683–695. дои : 10.1093/molbev/mst260 . ПМИД 24336922 . .
- ^ Чон; и др. (январь 2016 г.). «Глубокая история популяций Восточной Азии, раскрытая посредством генетического анализа айнов» . Генетика . 202 (1): 261–272. дои : 10.1534/genetics.115.178673 . ПМК 4701090 . ПМИД 26500257 .
- ^ Камберов, Яна Г (14 февраля 2013 г.). «Моделирование недавней эволюции человека у мышей путем экспрессии выбранного варианта EDAR» . Клетка . 152 (4): 691–702. дои : 10.1016/j.cell.2013.01.016 . ПМК 3575602 . ПМИД 23415220 .
- ^ Уэйд, Николас (14 февраля 2013 г.). «Восточноазиатские физические черты, связанные с 35 000-летней мутацией» . Нью-Йорк Таймс . Проверено 31 декабря 2019 г.
- ^ Пэн, Ю (2010). «Полиморфизм ADH1B ARG47His в популяциях Восточной Азии и расширение одомашнивания риса в истории» . Эволюционная биология BMC . 10 (15): 15. Бибкод : 2010BMCEE..10...15P . дои : 10.1186/1471-2148-10-15 . ПМЦ 2823730 . ПМИД 20089146 .
- ^ Ян, Мелинда А. (6 января 2022 г.). «Генетическая история миграции, диверсификации и примеси в Азии» . Популяционная генетика и геномика человека . 2 (1): 1–32. дои : 10.47248/hpgg2202010001 . ISSN 2770-5005 .
...Напротив, жители материковой части Восточной и Юго-Восточной Азии и другие жители островов Тихого океана (например, говорящие на австронезийском языке) тесно связаны друг с другом [9,15,16] и здесь обозначены как принадлежащие к линии Восточной и Юго-Восточной Азии (ESEA) ( Графа 2). …линия ESEA разделилась, по крайней мере, на три отдельные предки: предки Тяньюань, которые можно найти 40 000–33 000 лет назад в северной части Восточной Азии, предки, обнаруженные сегодня у современных популяций Восточной Азии, Юго-Восточной Азии и Сибири, но чье происхождение неизвестно, а происхождение Хоабинхов обнаружено 8 000–4 000 лет назад в Юго-Восточной Азии, но происхождение которых в верхнем палеолите неизвестно.
- ^ Питулько В.В.; Никольский, Пенсильвания; Гиря, EY; Басилян А.Е.; Тумской, В.Е.; Кулаков С.А.; Астахов С.Н.; Павлова, Э.Ю.; Анисимов, М.А. (2004). «Участок Яны RHS: Люди в Арктике до последнего ледникового максимума» Наука 303 (5654): 52–56. Бибкод : 2004Sci...303... 52P дои : 10.1126/science.1085219 . ПМИД 14704419 . S2CID 206507352 .
- ^ Jump up to: а б Уэллс, Спенсер; Прочтите, Марк (2002). Путешествие человека – генетическая одиссея . Случайный дом. стр. 138–140. ISBN 978-0812971460 .
- ^ Jump up to: а б Фитцхью, доктора. Уильям; Годдард, Айвз; Оусли, Стив; Оусли, Дуг; Стэнфорд, Деннис. «Палеоамериканский» . Отделение антропологии Смитсоновского института. Архивировано из оригинала 5 января 2009 года . Проверено 15 января 2009 г.
- ^ «Поиск ДНК первых американцев связывает группы Амазонки с коренными австралийцами» . Смитсоновский институт . Проверено 8 мая 2019 г.
- ^ Бонатто, СЛ; Сальзано, FM (1997). «Единичная и ранняя миграция населения Большой Америки, подтвержденная данными о последовательностях митохондриальной ДНК» . Труды Национальной академии наук . 94 (5): 1866–1871. Бибкод : 1997PNAS...94.1866B . дои : 10.1073/pnas.94.5.1866 . ПМК 20009 . ПМИД 9050871 .
- ^ Jump up to: а б «Атлас человеческого пути-генографический проект» . Национальное географическое общество. 1996–2008 гг. Архивировано из оригинала 1 мая 2011 года . Проверено 27 января 2017 г.
- ^ Спенсер Уэллс (2006). Глубокое происхождение: внутри генографического проекта . Национальные географические книги. стр. 222–. ISBN 978-0792262152 . OCLC 1031966951 .
- ^ Джон Х. Релетфорд (17 января 2017 г.). 50 великих мифов об эволюции человека: понимание заблуждений о нашем происхождении . Джон Уайли и сыновья. стр. 192–. ISBN 978-0470673911 . OCLC 1238190784 .
- ^ Х. Джеймс Биркс, изд. (10 июня 2010 г.). Антропология XXI века: Справочник . Публикации SAGE. ISBN 978-1452266305 . OCLC 1102541304 .
- ^ Джон Э. Кица; Ребекка Хорн (3 ноября 2016 г.). Устойчивые культуры: коренные народы Америки противостоят европейской колонизации 1500–1800 (2-е изд.). Рутледж. ISBN 978-1315509877 .
- ^ «Население Америки: генетическое происхождение влияет на здоровье» . Научный американец . Проверено 8 мая 2019 г.
- ^ Фладмарк, КР (1979). «Альтернативные миграционные коридоры для древних людей в Северной Америке». Американская древность . 44 (1): 55–69. дои : 10.2307/279189 . JSTOR 279189 . S2CID 162243347 .
- ^ «68 ответов на «Море поднимется до уровня последнего ледникового периода»» . Центр исследования климатических систем Колумбийского университета . 26 января 2009 года . Проверено 17 ноября 2009 г.
- ^ Геггель, Лаура (2 апреля 2021 г.). «Первые американцы имели гены коренных австралийцев» . www.livscience.com . Проверено 10 декабря 2022 г.
- ^ Су, Бинг; Ма, Ранлин З.; Чжан, Сяомин; Пэн, Йи; Чжун, Хуа; Ци, Сюэбин; Ши, Хун (20 июня 2013 г.). «Генетические доказательства восточноазиатского происхождения и палеолитической миграции на север гаплогруппы N Y-хромосомы» . ПЛОС ОДИН . 8 (6): е66102. Бибкод : 2013PLoSO...866102S . дои : 10.1371/journal.pone.0066102 . ISSN 1932-6203 . ПМЦ 3688714 . ПМИД 23840409 .
- ^ Гиббонс, Энн (4 сентября 2014 г.). «Трёхчастное происхождение европейцев» . Наука . Американская ассоциация содействия развитию науки. Архивировано из оригинала 11 октября 2014 года . Проверено 30 октября 2014 г.
- ^ Карри, Эндрю (август 2019 г.). «Первые европейцы были не такими, как вы думаете» . Нэшнл Географик . Архивировано из оригинала 19 марта 2021 года.
- ^ Гюнтер, Торстен; Мальмстрем, Хелена; Свенссон, Эмма М.; Омрак, Айча; и др. (9 января 2018 г.). «Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам» . ПЛОС Биология . 16 (1): e2003703. дои : 10.1371/journal.pbio.2003703 . ПМК 5760011 . ПМИД 29315301 .
- ^ Jump up to: а б Висенте, Марио; Шлебуш, Карина М (1 июня 2020 г.). «История населения Африки: взгляд на древнюю ДНК» . Текущее мнение в области генетики и развития . Генетика происхождения человека. 62 : 8–15. дои : 10.1016/j.где.2020.05.008 . ISSN 0959-437X . ПМИД 32563853 . S2CID 219974966 .
- ^ Джон Десмонд Кларк, От охотников к фермерам: причины и последствия производства продуктов питания в Африке , University of California Press, 1984, стр. 31
- ^ Игорь Копытофф, Африканский рубеж: воспроизводство традиционных африканских обществ (1989), 9–10 (цитируется по Корням языка игбо и (до)-истории, заархивировано 17 июля 2019 г. в Wayback Machine , A Mighty Tree , 2011) .
- ^ Вансина, Дж. (1995). «Новые лингвистические свидетельства и« экспансия банту » ». Журнал африканской истории . 36 (2): 173–195. дои : 10.1017/S0021853700034101 . JSTOR 182309 . S2CID 162117464 .
- ^ Басби, Джордж Би Джей; Бэнд, Гэвин; Си Ле, Куанг; Джаллоу, Муминато; Бугама, Эдит; Мангано, Валентина Д; Аменга-Этего, Лукас Н; Энимил, Энтони; Апинджо, Тобиас (2016). «Примесь в Африке к югу от Сахары и внутри нее» . электронная жизнь . 5 . дои : 10.7554/eLife.15266 . ISSN 2050-084X . ПМЦ 4915815 . ПМИД 27324836 .
- ^ Рамзи, Мишель; Хейзелхерст, Скотт; Сенгупта, Дхрити; Арон, Шон; Чоудхури, Ананьо (1 августа 2018 г.). «Генетическое разнообразие Африки дает новое понимание истории эволюции и местных адаптаций» . Молекулярная генетика человека . 27 (С2): Р209–Р218. дои : 10.1093/hmg/ddy161 . ISSN 0964-6906 . ПМК 6061870 . ПМИД 29741686 .
- ^ Jump up to: а б с Мичем, Уильям (1984–1985). «О невероятности австронезийского происхождения в Южном Китае» (PDF) . Азиатская перспектива . 26 : 89–106. Архивировано из оригинала (PDF) 9 ноября 2019 года . Проверено 9 ноября 2019 г.
- ^ Jump up to: а б с Беллвуд, Питер (1991). «Распространение австронезийцев и происхождение языков». Научный американец . 265 (1): 88–93. Бибкод : 1991SciAm.265a..88B . doi : 10.1038/scientificamerican0791-88 . JSTOR 24936983 .
- ^ Хит, Хелен; Саммерхейс, Гленн Р.; Хун, Сяо-чунь (2017). «Введите керамическую матрицу: определение природы раннего австронезийского поселения в долине Кагаян, Филиппины» . В Пайпер, Филип Дж.; Мацумара, Хирофуми; Булбек, Дэвид (ред.). Новые перспективы в предыстории Юго-Восточной Азии и Тихого океана . Южная земля. Том. 45. АНУ Пресс. ISBN 978-1760460952 .
- ^ Карсон, Майк Т.; Хун, Сяо-чунь; Саммерхейс, Гленн; Беллвуд, Питер (январь 2013 г.). «Гончарная тропа из Юго-Восточной Азии в отдаленную Океанию». Журнал островной и прибрежной археологии . 8 (1): 17–36. дои : 10.1080/15564894.2012.726941 . hdl : 1885/72437 . S2CID 128641903 .
- ^ Лансинг, Стив. «Изменил ли эффект бабочки историю Тихого океана?» . Стокгольмский центр устойчивости . Стокгольмский университет. Архивировано из оригинала 9 ноября 2019 года . Проверено 9 ноября 2019 г.
- ^ Лоулер, Эндрю (23 декабря 2020 г.). «Захватчики почти уничтожили первых жителей Карибского бассейна задолго до прихода испанцев, как показывает ДНК» . Нэшнл Географик . Архивировано из оригинала 23 декабря 2020 года.
- ^ Фэган, Б.М. (2007). Люди земли: Введение в мировую предысторию. Река Аппер-Сэддл, Нью-Джерси: Пирсон Прентис Холл
- ^ Хоффекер, Джон Ф. (2005). Предыстория севера: заселение человеком высоких широт . Издательство Университета Рутгерса. п. 130. ИСБН 978-0813534695 .
- ^ Гиббон, стр. 28–31.
- ^ Ригби, Брюс. «101. Исторический парк Кауммаарвит, Справочник Нунавута» (PDF) . Архивировано из оригинала (PDF) 29 мая 2006 года . Проверено 2 октября 2009 г.
- ^ Вуд, Шеннон Рэй (апрель 1992 г.). «Износ зубов и половое разделение труда среди инуитов» (PDF) . Кафедра археологии Университета Саскачевана . Университет Саймона Фрейзера . Проверено 2 октября 2009 г.
Дальнейшее чтение
[ редактировать ]- Деметр Ф., Шакелфорд Л.Л., Бэкон А.М., Дюрнер П., Вестэуэй К., Саявонгкхамди Т., Брага Дж., Сичантонгтип П., Кхамдалавонг П., Понче Дж.Л. и др. (2012). «Анатомически современный человек Юго-Восточной Азии (Лаос)» . Труды Национальной академии наук . 109 (36): 14375–14380. Бибкод : 2012PNAS..10914375D . дои : 10.1073/pnas.1208104109 . ПМЦ 3437904 . ПМИД 22908291 .
- Литт, Томас; Рихтер, Юрген; Шебиц, Франк (2021). Путешествие современного человека из Африки в Европу . Штутгарт, Германия: Издательство Schweizerbart Science. ISBN 978-3510655342 . Проверено 22 декабря 2021 г.
- Райх, Дэвид (2018). Кто мы и как мы сюда попали – древняя ДНК и новая наука о человеческом прошлом . Книги Пантеона . ISBN 978-1101870327 . ; Даймонд, Джаред (20 апреля 2018 г.). «Совершенно новая версия истории нашего происхождения» . Нью-Йорк Таймс . Проверено 23 апреля 2018 г.
- Вирама, Кришна Р.; Хаммер, Майкл Ф. (4 февраля 2014 г.). «Влияние полногеномного секвенирования на реконструкцию истории человеческой популяции». Обзоры природы Генетика . 15 (3): 149–162. дои : 10.1038/nrg3625 . ПМИД 24492235 . S2CID 19375407 .