Восточный охотник-собиратель
Материалы и судебно-криминалистическая реконструкция восточного охотника-собирателя с стоянки острова Южный Олений (ок . 8100 л.н. ) М.М. Герасимова . Национальный музей Карелии. [1] Охотники-собиратели в Европе между 14 и 9 тыс. лет назад, с основным ареалом восточных охотников-собирателей (EHG, ). Отдельные номера соответствуют датам калиброванной выборки. [2] |


В археогенетике восточный охотник-собиратель (EHG) , иногда восточноевропейский охотник-собиратель или восточноевропейский охотник-собиратель , является отдельным предковым компонентом, который представляет мезолитических охотников-собирателей Восточной Европы . [3]
Генетический профиль восточных охотников-собирателей в основном происходит от предков Древней Северной Евразии (ANE), которые были завезены из Сибири , [4] со вторичной и меньшей примесью европейских западных охотников-собирателей (WHG). [5] [6] Тем не менее, взаимосвязь между наследственными компонентами ANE и EHG еще не совсем понятна из-за отсутствия образцов, которые могли бы преодолеть пространственно-временной разрыв. [5]
В эпоху мезолита ЭГГ населяли территорию, простирающуюся от Балтийского моря до Урала и вниз до Понтийско-Каспийской степи . [7] Наряду со скандинавскими охотниками-собирателями (SHG) и западными охотниками-собирателями (WHG), EHG составляли одну из трех основных генетических групп в послеледниковый период ранней голоценовой Европы. [8] Граница между WHG и EHG проходила примерно от нижнего Дуная на север вдоль западных лесов Днепра к западной части Балтийского моря . [9]
Во время неолита и раннего энеолита , вероятно, в 4-м тысячелетии до н. э. ЭХГ в Понтийско-Каспийской степи смешивались с кавказскими охотниками-собирателями (ХГ), в результате чего получалось население, почти наполовину ЭХГ и наполовину ХГГ, образующее генетический кластер, известный как Западный. Степной оленевод (WSH). [10] [11] [12] Предполагается, что популяции WSH, тесно связанные с людьми ямной культуры, начали массовую миграцию , приведшую к распространению индоевропейских языков на значительной части Евразии.
Исследовать
[ редактировать ]

Хаак и др. (2015) идентифицировали ЭГГ как отдельный генетический кластер только у двух самцов. Самец EHG из Самары (датированный примерно 5650-5550 гг. до н.э.) нес Y-гаплогруппу R1b1a1a* и mt-гаплогруппу U5a1d . Другой мужчина EHG, похороненный в Карелии (датированный примерно 5500-5000 гг. до н. э.), нес Y-гаплогруппу R1a1 и mt-гаплогруппу C1g . Авторы исследования также выделили кластер WHG и кластер SHG, промежуточный между WHG и EHG. [а] Они предположили, что EHG имели смешанное происхождение от древних жителей Северной Евразии (ANE) и WHG. [14]

Исследователи предложили различные модели пропорций примесей для ЭТГ из WHG и ANE. [16] [17] Пост и др. (2023) обнаружили, что большинство людей с ЭГГ являются носителями c. 70% предков ANE и c. 30% предков WHG. WHG-подобное происхождение, скорее всего, произошло не непосредственно из кластеров Оберкасселя и Виллабруны, а из родственной, но еще не исследованной эпиграветтской популяции. [2] [18] Высокий вклад древних жителей Северной Евразии также виден в тонкой близости ЭХГ к 40 000-летнему человеку Тяньюань из Северного Китая , что можно объяснить потоком генов из источника, связанного с Тяньюанем, в линию ANE (представленную Мальтой и Афонтова Гора 3), что впоследствии существенно способствовало формированию ЭХГ. [19] По оценкам, ЭХГ (Сиделкино) на 9,4% имеют тяньюаньское происхождение благодаря своему древнему североевразийскому наследию. [20]
По оценкам, формирование предкового компонента EHG произошло 13 000–15 000 лет назад. [18] Связанные с ЭХГ останки принадлежали в первую очередь -хромосомы человека Y R1 с более низкой частотой гаплогрупп J и Q. гаплогруппам Их митохондриальные хромосомы принадлежали преимущественно к гаплогруппам U2 , U4 , U5 , а также C1 и R1b . [2]
ЭХГ, возможно, смешались с «армяноподобным ближневосточным источником», который сформировал ямную культуру еще в эпоху энеолита (5200–4000 гг. До н.э.). [21] Было обнаружено, что люди ямной культуры представляют собой смесь ЭХГ и «ближневосточного населения». В течение 3-го тысячелетия до нашей эры ямные люди приступили к массовой экспансии по всей Европе , что существенно изменило генетический ландшафт континента. Расширение породило такие культуры, как шнуровая керамика , и, возможно, стало источником распространения индоевропейских языков в Европе. [14]
Люди мезолитической кундинской культуры и нарвской культуры восточной Балтики представляли собой смесь WHG и EHG, демонстрируя самое близкое родство с WHG. Было обнаружено, что образцы украинского мезолита и неолита плотно сгруппированы между WHG и EHG, что позволяет предположить генетическую преемственность в Днепровских порогах в течение 4000 лет. Украинские образцы принадлежали исключительно к материнской гаплогруппе U , которая встречается примерно в 80% всех европейских образцов охотников-собирателей. [22]
Люди ямочно -гребенчатой культуры (PCW/CCC) восточной Балтики имеют 65% предков EHG. В этом отличие от более ранних охотников-собирателей в этом районе, которые были более тесно связаны с WHG. Это было продемонстрировано на образце Y-ДНК, выделенном у человека Pit-Comb Ware. Это принадлежало R1a15-YP172 . Четыре извлеченных образца мтДНК представляли собой два образца U5b1d1 , один образец U5a2d и один образец U4a . [23]
Гюнтер и др. (2018) проанализировали 13 ГСП и обнаружили, что все они имеют происхождение от ЭГГ. В целом, у ГСП из западной и северной Скандинавии было больше предков EHG (около 49%), чем у людей из восточной Скандинавии (около 38%). Авторы предположили, что ГСП представляли собой смесь WHG, мигрировавших в Скандинавию с юга, и EHG, которые позже мигрировали в Скандинавию с северо-востока вдоль норвежского побережья. У SHG наблюдалась более высокая частота генетических вариантов, вызывающих светлую кожу ( SLC45A2 и SLC24A5 ) и светлые глаза ( OCA/Herc2 ), чем у WHG и EHG. [24]
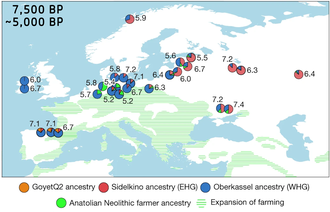

Также было обнаружено, что представители кундинской культуры и нарвской культуры более тесно связаны с WHG, тогда как культура ямно-гребенчатой керамики более тесно связана с EHG. Было обнаружено, что северные и восточные районы восточной Балтики более тесно связаны с ЭГГ, чем южные районы. Исследование отметило, что EHG, как и SHG и балтийские охотники-собиратели, несут высокие частоты производных аллелей SLC24A5 и SLC45A2, которые кодируют светлую кожу . [26]
Мэтисон и др. (2018) проанализировали генетику большого количества скелетов доисторической Восточной Европы. Тридцать семь образцов были из мезолитической и неолитической Украины (9500-6000 гг. до н. э.). Они были классифицированы как промежуточные между ЭГГ и ГВГ. Самцы принадлежали исключительно к гаплотипам R (особенно субкладам R1b1 и R1a ) и гаплотипам I (особенно субкладам I2 ). Митохондриальная ДНК принадлежала почти исключительно U (особенно субкладам U5 и U4 ). [21]
большое количество особей из могильника Звейниеки Проанализировано , которые в основном принадлежали кундинской культуре и нарвской культуре в восточной Прибалтике. На более ранних стадиях эти люди в основном были потомками WHG, но со временем происхождение EHG стало преобладающим. Y-ДНК этого сайта принадлежала почти исключительно гаплотипам гаплогруппы R1b1a1a и I2a1 . МтДНК принадлежала исключительно гаплогруппе U (особенно субкладам U2 , U4 и U5 ). [21]
По оценкам, сорок человек из трех стоянок мезолита Железных ворот на Балканах имели 85% WHG и 15% EHG происхождение. Самцы на этих участках несли исключительно гаплотипы R1b1a и I (в основном субклады I2a ). мтДНК принадлежала в основном U (особенно субкладам U5 и U4 ). [21]
Было обнаружено, что люди культуры Кукутень-Триполье имеют около 20% предков охотников-собирателей, что занимает промежуточное положение между EHG и WHG. [21]
Нарасимшан и др. (2019) ввели новый наследственный компонент — Западно-Сибирский охотник-собиратель (WSHG). WSHG содержали около 20% предков EHG, 73% предков ANE и 6% восточноазиатских предков. [27]
Возможная ассоциация с ранним индоевропейским
[ редактировать ]Некоторые утверждают, что EHG представляет собой возможный источник препротоиндоевропейского языка (см. Также гипотезу отцовского языка ). В отличие от людей ямной культуры (или близкородственных групп), которые связаны с носителями протоиндоевропейского языка, люди богатой EHG днепровско-донецкой культуры не обнаруживают никаких свидетельств кавказских охотников-собирателей (CHG) или ранних европейских земледельцев (EEF). ) родословная. [28] И днепровско-донецкие самцы, и ямные самцы несут одни и те же отцовские гаплогруппы (R1b и I2a), что позволяет предположить, что примесь CHG и EEF среди ямных произошла в результате смешения самцов EHG с самками EEF и CHG. На основании этого Дэвида В. Энтони можно предположить, что на индоевропейских языках изначально говорили ЭХГ, живущие в Восточной Европе. [29]
Другие предположили, что индоевропейская языковая семья, возможно, возникла не в Восточной Европе, а среди богатых CHG популяций Западной Азии к югу от Кавказа, которые позже поглотили богатые EHG группы к северу от Кавказа. Было отмечено, что гаплогруппы могут не коррелировать с компонентами аутосомного происхождения и историческим распространением языков. [30]
Внешний вид
[ редактировать ]
Предполагается, что у EHG в основном были карие глаза и светлая кожа. [24] [32] с «промежуточными частотами голубоглазых вариантов» и «высокими частотами светлокожих вариантов». [33] определил, что ЭХГ из Карелии Гюнтер (2018) имеет высокую вероятность быть кареглазыми и темноволосыми с прогнозируемым промежуточным тоном кожи. [34] Другой EHG из Самары, по прогнозам, был светлокожим, и было установлено, что он с высокой вероятностью будет голубоглазым и светлым оттенком волос, с расчетной вероятностью 75% - блондином. [35] [33]
rs12821256 Аллель гена KITLG , контролирующего развитие меланоцитов и синтез меланина , [36] который связан со светлыми волосами и впервые обнаружен у человека из Сибири, датируемый примерно 17 000 лет назад, обнаружен у трех восточных охотников-собирателей из Самары, Моталы и Украины c. 10 000 лет назад , что позволяет предположить, что этот аллель возник у древнего населения Северной Евразии, а затем распространился на Западную Евразию. [37]
Многие останки восточных охотников-собирателей, датируемые примерно 8100 годом до нашей эры (6100 лет до нашей эры), также были раскопаны на Южный Олений острове в Онежском озере . [38] Древние североевразийские (ANE) предки, безусловно, являются основным компонентом группы Южных Олений и являются одними из самых высоких среди остальных восточных охотников-собирателей (EHG). [4]
![Реконструкция могильника № 132 Оленеостровского могильника (остров Южный Олений, Онежское озеро). Экспонат Национального музея Республики Карелия.[39]](//upload.wikimedia.org/wikipedia/commons/thumb/3/39/Burial_ground._Karelia.jpg/200px-Burial_ground._Karelia.jpg) Реконструкция могильника № 132 Оленеостровского могильника (остров Южный Олений, Онежское озеро). Экспонат Национального музея Республики Карелия. [39]
Реконструкция могильника № 132 Оленеостровского могильника (остров Южный Олений, Онежское озеро). Экспонат Национального музея Республики Карелия. [39]![Артефакты и реконструкция Герасимова восточных охотников-собирателей с острова Южный Олений.[39]](//upload.wikimedia.org/wikipedia/commons/thumb/d/de/National_museum_of_Karelia_10.JPG/200px-National_museum_of_Karelia_10.JPG) Артефакты и реконструкция Южный Олений . Герасимова восточных охотников-собирателей с острова [39]
Артефакты и реконструкция Южный Олений . Герасимова восточных охотников-собирателей с острова [39] Карельский петроглиф с изображением пяти лыжников и оленя. Эти петроглифы датируются 7000–6000 лет назад.
Карельский петроглиф с изображением пяти лыжников и оленя. Эти петроглифы датируются 7000–6000 лет назад.
![Реконструкция могильника № 132 Оленеостровского могильника (остров Южный Олений, Онежское озеро). Экспонат Национального музея Республики Карелия.[39]](http://upload.wikimedia.org/wikipedia/commons/thumb/3/39/Burial_ground._Karelia.jpg/200px-Burial_ground._Karelia.jpg)
![Артефакты и реконструкция Герасимова восточных охотников-собирателей с острова Южный Олений.[39]](http://upload.wikimedia.org/wikipedia/commons/thumb/d/de/National_museum_of_Karelia_10.JPG/200px-National_museum_of_Karelia_10.JPG)

Материальная культура
[ редактировать ]
Будучи охотниками-собирателями, EHG изначально полагались на каменные орудия труда и артефакты, полученные из слоновой кости, рогов или рогов. Примерно с 5900 г. до н. э. гончарное дело начали перенимать в районе северного Каспия или, возможно, из-за Урала. Всего за три-четыре столетия керамика распространилась на расстояние около 3000 километров, достигнув Балтийского моря . Это технологическое распространение было намного быстрее, чем распространение самого сельского хозяйства, и в основном происходило за счет передачи технологий между группами охотников-собирателей, а не за счет демического распространения земледельцев. [41]
См. также
[ редактировать ]- Днепровско-Донецкая культура
- Гребень Керамическая культура
- Sredny Stog culture
- Дереевка
- Самарская культура
- Kvalynsk culture
Примечания
[ редактировать ]- ^ Лазаридис и др. (2016) обнаружили, что ГСП представляют собой смесь EHG и WHG: «Восточные охотники-собиратели (EHG) на 3/4 происходят от ANE... Скандинавские охотники-собиратели (SHG) представляют собой смесь EHG и WHG; и WHG представляют собой смесь EHG и верхнепалеолитического бишона из Швейцарии». [13]
Ссылки
[ редактировать ]- ^ Экспонат Национального музея Карелии.
- ^ Перейти обратно: а б с Пост, Козимо; Ю, Хэ; Галичи, Айшин (март 2023 г.). «Палеогеномика от верхнего палеолита до неолита европейских охотников-собирателей» . Природа . 615 (7950): 117–126. Бибкод : 2023Natur.615..117P . дои : 10.1038/s41586-023-05726-0 . ISSN 1476-4687 . ПМЦ 9977688 . ПМИД 36859578 .
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; Харни, Эдаоин; Стюардсон, Кристин; Фу, Цяомэй (1 июня 2015 г.). «Массовая миграция из степи стала источником индоевропейских языков в Европе». Природа. 522 (7555): 207–211. дои: 10.1038/nature14317. ISSN 1476-4687. ПМЦ 5048219. ПМИД 25731166.
- ^ Перейти обратно: а б Козинцев А.Г. (4 января 2022 г.). «Закономерности истории населения Северной Евразии от мезолита до раннего бронзового века по данным краниометрии и генетики» . Археология, этнология и антропология Евразии . 49 (4): 141. дои : 10.17746/1563-0110.2021.49.4.140-151 .
АНЭ составляет основную долю аутосомного компонента ЭХГ (Восточный охотник-собиратель), содержание которого особенно велико в геномах мезолита и раннего неолита жителей Северо-Востока Европы, захороненных в Южном Оленом Острове, Попово, Сиделькино, Лебяжинке IV и др. (Хаак и др., 2015; Дамгаард и др., 2018). Они передали ЭХГ ямникам, от которых ее унаследовали несколько сыновних популяций, в том числе афанасьевцы. Еще в эпоху мезолита ЭХГ был завезен из северной России в Скандинавию, о чем свидетельствуют геномы народа мотала на юге Швеции. Их предки мигрировали туда с востока вдоль побережья Норвегии, потому что доля ЭГГ у более южных популяций, таких как более ранний народ кунда восточной Балтики, ниже (Haak et al., 2015; Mittnik et al., 2018).
- ^ Перейти обратно: а б Фельдман, Михал; Некки-Русконе, Гвидо А.; Ламнидис, Тисей К.; Пост, Козимо (2021). «Там, где Азия встречается с Европой – последние открытия в области геномики древнего человека». Анналы биологии человека . 48 (3): 191–202. дои : 10.1080/03014460.2021.1949039 . ПМИД 34459345 . S2CID 237348859 .
- ^ Нэгеле, Катрин; Риволлат, Майте; Ю, Хэ; Ван, Кэ (2022). «Древние геномные исследования - от широких мазков до тонких реконструкций прошлого». Журнал антропологических наук . 100 (100): 193–230. дои : 10.4436/jass.10017 . ПМИД 36576953 .
- ^ Энтони 2019b , с. 27.
- ^ Кашуба 2019 : «Более ранние исследования адДНК предполагают наличие трех генетических групп в ранней послеледниковой Европе: западных охотников-собирателей (WHG), восточных охотников-собирателей (EHG) и скандинавских охотников-собирателей (SHG)4. SHG были смоделировано как смесь WHG и EHG».
- ^ Энтони 2019b , с. 28.
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; Харни, Эдаоин; Стюардсон, Кристин; Фу, Цяомэй (1 июня 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ISSN 1476-4687 . ПМК 5048219 . ПМИД 25731166 .
- ^ из Лоосдрехта, Марике; Бузуггар, Абдельджалил; Хамфри, Луиза; Пост, Козимо; Бартон, Ник; Аксим-Петри, Айинуэр; Никель, Биргит; Нагель, Сара; Тальби, Эль-Хасан; Эль-Хаджрауи, Мохаммед Абдельджалил; Амзази, Саид; Юблин, Жан-Жак; Паабо, Сванте; Шиффельс, Стивен; Мейер, Матиас (4 мая 2018 г.). «Плейстоценовые геномы Северной Африки, связанные с человеческими популяциями Ближнего Востока и Африки к югу от Сахары» . Наука 360 (6388): 548–552. Бибкод : 2018Науч... 360..548В дои : 10.1126/science.year8380 . ISSN 0036-8075 . ПМИД 29545507 .
- ^ Лазаридис, Иосиф; Алпаслан-Роденберг, Сонгюль; Ачар, Айше; Ачикколь, Айшен; Агеларакис, Анагностис; Агикян, Левон; Акюз, Угур; Андреева, Десислава; Андрияшевич, Гойко; Антонович, Драгана; Армит, Ян; Атмака, Альпер; Аветисян Павел; Айтек, Ахмет Ихсан; Бачваров, Крум (26 августа 2022 г.). «Генетическая история Южной дуги: мост между Западной Азией и Европой» . Наука . 377 (6609): eabm4247. дои : 10.1126/science.abm4247 . ISSN 0036-8075 . ПМЦ 10064553 . ПМИД 36007055 . S2CID 251843620 .
- ^ Лазаридис 2016 .
- ^ Перейти обратно: а б Вязание крючком 2015г .
- ^ Ирвинг-Пиз, Эван К.; Рефойо-Мартинес, Альба; Барри, Уильям; Ингасон, Андрес; Пирсон, Элис; Фишер, Андерс; Шегрен, Карл-Йоран; Халгрен, Альма С.; Маклеод, Руайрид; Деметра, Фабрис; Хенриксен, Расмус А.; Вимала, Тарсика; Макколл, Хью; Вон, Эндрю Х.; Шпайдель, Лео (24 января 2024 г.). «Селекционный ландшафт и генетическое наследие древних евразийцев» . Природа . 625 (7994): 312–320. Бибкод : 2024Natur.625..312I . дои : 10.1038/s41586-023-06705-1 . ISSN 1476-4687 . ПМЦ 10781624 . ПМИД 38200293 .
- ^ Лазаридис, Иосиф (1 декабря 2018 г.). «Эволюционная история человеческих популяций в Европе» . Текущее мнение в области генетики и развития . Генетика происхождения человека. 53 : 21–27. arXiv : 1805.01579 . дои : 10.1016/j.gde.2018.06.007 . ISSN 0959-437X . ПМИД 29960127 . S2CID 19158377 .
- ^ Хаак, Вольфганг; Лазаридис, Иосиф; Паттерсон, Ник; Роланд, Надин; Маллик, Свапан; Ламас, Бастьен; Брандт, Гвидо; Норденфельт, Сюзанна; Харни, Эдаоин; Стюардсон, Кристин; Фу, Цяомэй; Миттник, Алисса; Банфи, Эстер; Эконому, Христос; Франкен, Майкл (июнь 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ISSN 1476-4687 . ПМК 5048219 . ПМИД 25731166 .
Хаак и др. (2015): 38–40% ANE (MA-1), 60–62% WHG (рис. S8.6). (Альтернативные топологии, в которых EHG и ANE представляют собой несмешанные сестринские линии с примесью WHG, не отвергаются)
- ^ Перейти обратно: а б Аллентофт, Мортен Э.; Сикора, Мартин; Рефойо-Мартинес, Альба; Ирвинг-Пиз, Эван К.; Фишер, Андерс; Барри, Уильям; Ингасон, Андрес; Стендерап, Йеспер; Шегрен, Карл-Йоран; Пирсон, Элис; Соуза да Мота, Барбара; Шульц Паулссон, Беттина; Халгрен, Альма; Маклеод, Руайрид; Йорков, Мария Луиза Шеллеруп (январь 2024 г.). «Популяционная геномика послеледниковой Западной Евразии» . Природа . 625 (7994): 301–311. Бибкод : 2024Natur.625..301A . дои : 10.1038/s41586-023-06865-0 . ISSN 1476-4687 . ПМЦ 10781627 . ПМИД 38200295 .
- ^ Вильяльба-Моуко, Ванесса; ван де Лоосдрехт, Марике С.; Рорлах, Адам Б.; Фьюласс, Хелен; Таламо, Сахра; Ю, Хэ; Арон, Франциска; Лалуэса-Фокс, Карлес; Кабельо, Лидия; Канталехо Дуарте, Педро; Рамос-Муньос, Хосе; Пост, Козимо; Краузе, Йоханнес; Венигер, Герд-Кристиан; Хаак, Вольфганг (апрель 2023 г.). «23 000-летний житель Южного Иберийского полуострова связывает группы людей, которые жили в Западной Европе до и после последнего ледникового максимума» . Экология и эволюция природы . 7 (4): 597–609. Бибкод : 2023NatEE...7..597В . дои : 10.1038/s41559-023-01987-0 . ISSN 2397-334X . ПМК 10089921 . ПМИД 36859553 .
В настоящее время самая сильная близость к Тяньюаню среди европейских HG голоцена зарегистрирована для восточноевропейских HG (EHG). Это связано с тем, что предки, обнаруженные на Мальте и Афонтовой Горе (древние североевразийские предки), произошли от популяций UP Восточной Азии / Юго-Восточной Азии54, которые затем внесли существенный вклад в EHG55.
- ^ Чилдебаева, Айнаш; Фрике, Фабиан; Рорлах, Адам Бенджамин; Хуан, Лей; Шиффельс, Стефан; Весакоски, Оути; Маннермаа, Кристина; Семерау, Лена; Арон, Франциска; Солодовников Константин; Рыкун, Марина; Моисеев Вячеслав; Хартанович, Валерий; Ковтун Игорь; Краузе, Йоханнес (11 июня 2024 г.). «Генетика Северной Евразии бронзового века в контексте развития металлургии и сибирской родословной» . Коммуникационная биология . 7 (1): 1–12. дои : 10.1038/s42003-024-06343-x . ISSN 2399-3642 . ПМЦ 11166947 .
Затем мы смоделировали поток генов из линии, ведущей к CHB, в EEHG на уровне 9,4% (95% ДИ 4,4–14,7%).
- ^ Перейти обратно: а б с д и Мэтисон и др. 2018 .
- ^ Джонс 2017 .
- ^ Видел 2017 год .
- ^ Перейти обратно: а б Гюнтер 2018 .
- ^ Чжан, Фань; Нин, Чао; Скотт, Эшли (ноябрь 2021 г.). «Геномное происхождение мумий Таримской котловины бронзового века» . Природа . 599 (7884): 256–261. Бибкод : 2021Natur.599..256Z . дои : 10.1038/s41586-021-04052-7 . ISSN 1476-4687 . ПМЦ 8580821 . ПМИД 34707286 .
- ^ Миттник 2018 .
- ^ Нарасимхан 2019 .
- ^ Энтони 2019a , с. 14.
- ^ Энтони 2019a , стр. 7, 14.
- ^ Лазаридис, Иосиф; Алпаслан-Роденберг, Сонгюль; Ачар, Айше; Ачикколь, Айшен; Агеларакис, Анагностис; Агикян, Левон; Акюз, Угур; Андреева, Десислава; Андрияшевич, Гойко; Антонович, Драгана; Армит, Ян; Атмака, Альпер; Аветисян Павел; Айтек, Ахмет Ихсан; Бачваров, Крум (26 августа 2022 г.). «Генетическая история Южной дуги: мост между Западной Азией и Европой» . Наука . 377 (6609): eabm4247. дои : 10.1126/science.abm4247 . ISSN 0036-8075 . ПМЦ 10064553 . ПМИД 36007055 .
- ^ Ханель, Андреа; Карлберг, Карстен (3 июля 2020 г.). «Цвет кожи и витамин D: обновленная информация» . Экспериментальная дерматология . 29 (9): 864–875. дои : 10.1111/exd.14142 . ПМИД 32621306 . S2CID 220335539 .
- ^ Ханель, Андреа; Карлберг, Карстен (2020). «Цвет кожи и витамин D: обновленная информация» . Экспериментальная дерматология . 29 (9): 864–875. дои : 10.1111/exd.14142 . ПМИД 32621306 . S2CID 220335539 . «Интересно, что восточные и скандинавские охотники-собиратели имели светлую кожу,[48] в отличие от балтийских охотников-собирателей, которые сохраняли свою темную кожу только до 3800 лет назад, когда земледелие в этом регионе было введено в результате экспансии жителей русских степей в бронзовом веке. происхождения.[56, 57]»
- ^ Перейти обратно: а б Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам S8 Text. Функциональная вариация древних образцов. , doi : 10.1371/journal.pbio.2003703.s013
- ^ Гюнтер 2018 , с. 28 апреля: Из дополнительного документа S8: «У карельского человека высока вероятность быть кареглазым (0,99) и тёмным цветом волос (0,96). Не размышляя о генетической архитектуре пигментации кожи, мы предполагаем промежуточный тип кожи — фенотип пигментации для жителя Карелии, поскольку он нес предковый аллель rs16891982 и производный аллель rs1426654 (таблица S1). Наличие аллеля светлой кожи rs1426654 в дополнение к пяти дополнительным аллелям, связанным с C11, в гаплотипе, определяющем SNP (). Таблица S1) предполагает, что карельский человек нес гаплотип светлой кожи C11».
- ^ Гюнтер 2018 , с. 28 апреля: Из дополнительного документа S8: «У самарца высока вероятность быть голубоглазым (0,88), светлым оттенком волос (0,99); скорее всего, блондином (0,75)».
- ^ Сулем, Патрик; Гудбьартссон, Дэниел Ф.; Стейси, Саймон Н.; Хельгасон, Агнар; Рафнар, Торунн; Магнуссон, Кристинн П.; Манолеску, Андрей; Карасон, Ари; Палссон, Арнар; Торлейфссон, Гудмар; и др. (декабрь 2007 г.). «Генетические детерминанты пигментации волос, глаз и кожи у европейцев» . Природная генетика . 39 (12): 1443–1452. дои : 10.1038/ng.2007.13 . ISSN 1546-1718 . ПМИД 17952075 . S2CID 19313549 .
- ^ Мэтисон и др. 2018 «Дополнительная информация, стр. 52: «Производный аллель KITLG SNP rs12821256, который связан и, вероятно, является причиной светлых волос у европейцев, присутствует у одного охотника-собирателя из Самары, Моталы и Украины (I0124, I0014 и I1763). ), а также несколько более поздних особей степного происхождения. Поскольку аллель обнаруживается в популяциях с происхождением EHG, но не WHG, это позволяет предположить, что его происхождение происходит от популяции Древней Северной Евразии (ANE). В соответствии с этим мы наблюдаем, что самой ранней известной особью с производным аллелем (подтвержденной двумя прочтениями) является особь ANE Афонтова Гора 3, которая напрямую датируется 16130-15749 кал. до н.э. (14710±60 BP, MAMS-27186: a ранее неопубликованная дата, о которой мы недавно сообщаем здесь). Мы не можем определить статус rs12821256 в Афонтова Гора 2 и МА-1 из-за отсутствия покрытия последовательности в этом SNP».
- ^ Миттник, Алисса; Ван, Чуан-Чао; Пфренгл, Саския (30 января 2018 г.). «Генетическая предыстория региона Балтийского моря» . Природные коммуникации . 9 (1): Рис. 1. Бибкод : 2018NatCo...9..442M . дои : 10.1038/s41467-018-02825-9 . ISSN 2041-1723 . ПМЦ 5789860 . ПМИД 29382937 .
- ^ Перейти обратно: а б Козинцев Александр (1 января 2021 г.). «Закономерности истории населения Северной Евразии от мезолита до ранней бронзы» . Археология, этнология и антропология Евразии . 49 (4): 140–151. дои : 10.17746/1563-0110.2021.49.4.140-151 .
АНЭ составляет основную долю аутосомного компонента ЭХГ (Восточный охотник-собиратель), содержание которого особенно велико в геномах мезолита и раннего неолита жителей Северо-Востока Европы, захороненных в Южном Оленом Острове, Попово, Сиделькино, Лебяжинке IV и др. (Хаак и др., 2015; Дамгаард и др., 2018).», «Мезолит, Север Русской равнины, Южный Олений остров (Алексеев, Гохман, 1984)
- ^ Долбунова Екатерина; Люкен, Александр (февраль 2023 г.). «Передача гончарной технологии среди доисторических европейских охотников-собирателей» . Природа человеческого поведения . 7 (2): 171–183. дои : 10.1038/s41562-022-01491-8 . ISSN 2397-3374 . ПМЦ 9957732 . ПМИД 36550220 .
- ^ Долбунова Екатерина; Люкен, Александр (февраль 2023 г.). «Передача гончарной технологии среди доисторических европейских охотников-собирателей» . Природа человеческого поведения . 7 (2): 171–183. дои : 10.1038/s41562-022-01491-8 . ISSN 2397-3374 . ПМЦ 9957732 . ПМИД 36550220 .
Хотя демическое распространение может сыграть свою роль, на основании его скорости мы утверждаем, что производство керамики быстро распространилось посредством передачи знаний через устоявшиеся сети между рассредоточенными сообществами охотников-собирателей.
Библиография
[ редактировать ]- Энтони, Дэвид (весна – лето 2019a). «Археология, генетика и язык в степях: комментарий к Бомхарду» . Журнал индоевропейских исследований . 47 (1–2) . Проверено 9 января 2020 г.
- Энтони, Дэвид В. (2019b). «Древняя ДНК, сети спаривания и анатолийский раскол». В Серанджели Матильда; Оландер, Томас (ред.). Рассредоточение и диверсификация: лингвистические и археологические перспективы ранних этапов индоевропейского происхождения . БРИЛЛ . стр. 21–54. ISBN 978-9004416192 .
- Гюнтер, Торстен (1 января 2018 г.). «Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам» . ПЛОС Биология . 16 (1). ПЛОС : e2003703. дои : 10.1371/journal.pbio.2003703 . ПМК 5760011 . ПМИД 29315301 .
- Хаак, Вольфганг (11 июня 2015 г.). «Массовая миграция из степи явилась источником индоевропейских языков в Европе» . Природа . 522 (7555): 207–211. arXiv : 1502.02783 . Бибкод : 2015Natur.522..207H . дои : 10.1038/nature14317 . ПМК 5048219 . ПМИД 25731166 .
- Джонс, Эппи Р. (20 февраля 2017 г.). «Переход к неолиту в Прибалтике не был вызван смешением с ранними европейскими земледельцами» . Современная биология . 27 (4). Cell Press : 576–582. Бибкод : 2017CBio...27..576J . дои : 10.1016/j.cub.2016.12.060 . ПМК 5321670 . ПМИД 28162894 .
- Кашуба, Наталья (15 мая 2019 г.). «Древняя ДНК из мастики укрепляет связь между материальной культурой и генетикой мезолитических охотников-собирателей в Скандинавии» . Коммуникационная биология . 2 (105). Исследования природы : 185. doi : 10.1038/s42003-019-0399-1 . ПМК 6520363 . ПМИД 31123709 .
- Лазаридис, Иосиф (25 июля 2016 г.). «Геномный взгляд на происхождение земледелия на древнем Ближнем Востоке» . Природа . 536 (7617): 419–424. Бибкод : 2016Natur.536..419L . дои : 10.1038/nature19310 . ПМЦ 5003663 . ПМИД 27459054 .
- Мэтисон, Иэн (23 ноября 2015 г.). «Общегеномные закономерности отбора у 230 древних евразийцев» . Природа . 528 (7583): 499–503. Бибкод : 2015Natur.528..499M . дои : 10.1038/nature16152 . ПМЦ 4918750 . ПМИД 26595274 .
- Мэтисон, Иэн; Алпаслан-Роденберг, Сонгюль; Пост, Козимо; Сечени-Надь, Анна; и др. (март 2018 г.). «Геномная история Юго-Восточной Европы» . Природа . 555 (7695): 197–203. Бибкод : 2018Natur.555..197M . дои : 10.1038/nature25778 . ПМК 6091220 . ПМИД 29466330 .
- Миттник, Алиса (30 января 2018 г.). «Генетическая предыстория региона Балтийского моря» . Природные коммуникации . 16 (1): 442. Бибкод : 2018NatCo...9..442M . дои : 10.1038/s41467-018-02825-9 . ПМЦ 5789860 . ПМИД 29382937 .
- Нарасимхан, Вагиш М. (6 сентября 2019 г.). «Формирование человеческих популяций в Южной и Центральной Азии» . Наука . 365 (6457). Американская ассоциация содействия развитию науки : eaat7487. bioRxiv 10.1101/292581 . дои : 10.1126/science.aat7487 . ПМЦ 6822619 . ПМИД 31488661 .
- Сааг, Лехти (24 июля 2017 г.). «Экстенсивное земледелие в Эстонии началось благодаря миграции из степи по признаку пола» . Современная биология . 27 (14). Cell Press : 2185–2193. Бибкод : 2017CBio...27E2185S . дои : 10.1016/j.cub.2017.06.022 . ПМИД 28712569 .
- Ван, Чуан-Чао (4 февраля 2019 г.). «Древние данные по всему геному человека за 3000-летний интервал на Кавказе соответствуют эколого-географическим регионам Евразии» . Природные коммуникации . 10 (1): 590. Бибкод : 2019NatCo..10..590W . дои : 10.1038/s41467-018-08220-8 . ПМК 6360191 . ПМИД 30713341 .
Дальнейшее чтение
[ редактировать ]- Энтони, Дэвид (весна – лето 2019 г.). «Археология, генетика и язык в степях: комментарий к Бомхарду» . Журнал индоевропейских исследований . 47 (1–2) . Проверено 9 января 2020 г.
- Энтони, Дэвид В. (2019b). «Древняя ДНК, сети спаривания и анатолийский раскол». В Серанджели Матильда; Оландер, Томас (ред.). Рассредоточение и диверсификация: лингвистические и археологические перспективы ранних этапов индоевропейского происхождения . БРИЛЛ . стр. 21–54. ISBN 978-9004416192 .
- Аллентофт, Мортен Э.; Сикора, Мартин; Шегрен, Карл-Горан; Расмуссен, Саймон; Расмуссен, Мортен; Стендерап, Йеспер; Дамгаард, Питер Б.; Шредер, Ханнес; Альстрем, Торбьёрн; Победитель, Лассе; Маласпинас, Анна-Сапфо (2015). «Популяционная геномика Евразии бронзового века» . Природа . 522 (7555): 167–172. Бибкод : 2015 Природа.522..167А . дои : 10.1038/nature14507 . ISSN 1476-4687 . ПМИД 26062507 . S2CID 4399103 .
- Лазаридис, Иосиф (декабрь 2018 г.). «Эволюционная история человеческих популяций в Европе» . Текущее мнение в области генетики и развития . 53 . Эльзевир : 21–27. arXiv : 1805.01579 . дои : 10.1016/j.gde.2018.06.007 . ПМИД 29960127 . S2CID 19158377 . Проверено 15 июля 2020 г.
| Подтемы | |
|---|---|
| Генетическая история по регионам | |
| Популяционная генетика по группе |
|