Светлая кожа


Светлая кожа — это цвет кожи человека с низким уровнем пигментации эумеланина , что является адаптацией к среде с низким УФ-излучением . [1] [2] Из-за миграций людей в последние столетия светлокожие популяции сегодня встречаются по всему миру. [2] [3] Светлая кожа чаще всего встречается у коренного населения Европы , Восточной Азии , [4] [5] [6] Западная Азия , Центральная Азия и Сибирь , измеренные по отражательной способности кожи . [7] Людей со светлой пигментацией кожи часто называют « белыми ». [8] [9] хотя эти обычаи могут быть неоднозначными в некоторых странах, где они используются конкретно для обозначения определенных этнических групп или групп населения. [10]
У людей со светлой пигментацией кожи кожа с низким содержанием эумеланина и меньше меланосом , чем у людей с темной пигментацией кожи. Светлая кожа обеспечивает лучшее поглощение ультрафиолетового излучения, что помогает организму синтезировать большее количество витамина D для таких процессов в организме, как выработка кальция. [2] [11] С другой стороны, светлокожие люди, живущие вблизи экватора , где много солнечного света , подвергаются повышенному риску истощения запасов фолиевой кислоты . В результате истощения запасов фолиевой кислоты они подвергаются более высокому риску повреждения ДНК , врожденных дефектов и многих видов рака , особенно рака кожи . [12] Люди с более темной кожей, живущие дальше от тропиков , могут иметь более низкий уровень витамина D, что также может привести к осложнениям со здоровьем, как физическим, так и психическим , включая больший риск развития шизофрении . [13] Эти два наблюдения формируют «гипотезу витамина D-фолата», которая пытается объяснить, почему популяции, мигрировавшие из тропиков в районы с низким УФ-излучением. [14] эволюционировали, чтобы иметь светлую пигментацию кожи. [2] [15] [16]
Распределение светлокожих популяций тесно коррелирует с низким уровнем ультрафиолетового излучения регионов их проживания. Исторически сложилось так, что светлокожие популяции почти исключительно жили вдали от экватора, в высоких широтах с низкой интенсивностью солнечного света. [17]
Эволюция
[ редактировать ]
Принято считать, что темная кожа возникла как защита от воздействия УФ-излучения ; Эумеланин защищает как от истощения запасов фолиевой кислоты , так и от прямого повреждения ДНК . [2] [20] [21] [22] Этим объясняется темная пигментация кожи Homo sapiens во время их развития в Африке; основные миграции из Африки с целью колонизации остального мира также были темнокожими. [23] Широко распространено мнение, что легкая пигментация кожи возникла из-за важности поддержания выработки витамина D3 в коже. [24] сильное избирательное давление для развития светлой кожи в областях с низким УФ-излучением. Ожидается [15]
После того, как предки западных евразийцев и восточноевразийцев разошлись более 40 000 лет назад, более светлые тона кожи развились независимо в подмножестве каждой из двух популяций. У жителей Западной Евразии А111Т аллель rs1426654 полиморфизма гена пигментации SLC24A5 обладает наибольшим эффектом осветления кожи и широко распространен в Европе, Южной Азии, Центральной Азии, на Ближнем Востоке и в Северной Африке. [25]
В исследовании 2013 года Canfield et al. установили, что SLC24A5 находится в блоке гаплотипов , один из которых (C11) является общим практически для всех хромосом, несущих вариант A111T . Эта «эквивалентность» между C11 и A111T указывает на то, что все люди, несущие этот аллель осветления кожи, происходят от общего происхождения: от одного носителя, который, скорее всего, жил «между Ближним Востоком и Индийским субконтинентом». Кэнфилд и др. попытался датировать мутацию A111T , но ограничил возрастной диапазон только до эпохи неолита. [25] Однако второе исследование того же года (Басу Маллик и др.) оценило возраст слияния (дату разделения) для этого аллеля между ~ 28 000 и ~ 22 000 лет назад. [26]
Вторым по важности фактором осветления кожи у жителей Западной Евразии является депигментирующий аллель F374 полиморфизма rs16891982, расположенный в меланина гене синтеза SLC45A2 . Из-за низкого разнообразия гаплотипов Yuasa et al. (2006) также пришли к выводу, что эта мутация ( L374F ) «произошла только один раз в предках европеоидов». [27]
Подводя итог этим исследованиям, Ханель и Карлберг (2020) пришли к выводу, что аллели двух генов SLC24A5 и SLC45A2 , которые больше всего связаны с более светлым цветом кожи у современных европейцев, возникли в Западной Азии примерно 22 000–28 000 лет назад, и каждая из этих двух мутаций возникла в один перевозчик. [23] Это согласуется с данными Джонса и др. (2015), которые реконструировали взаимоотношения между ближневосточными неолитическими земледельцами и кавказскими охотниками-собирателями : двумя популяциями, которые несли светлокожий вариант SLC24A5 . Анализируя недавно секвенированные древние геномы, Jones et al. оценил дату разделения примерно в 24 000 лет назад и локализовал разделение где-то к югу от Кавказа. [28] Однако совместный анализ этого аллеля, проведенный Crawford et al. (2017) дали более узкую и более раннюю дату разделения - ~ 29 000 лет назад (с окном достоверности 95% от 28 000 до 31 000 лет назад). [29]
Варианты светлой кожи SLC24A5 и SLC45A2 присутствовали в Анатолии 9000 лет назад, где они стали ассоциироваться с неолитической революцией . Отсюда их носители распространили неолитическое земледелие по Европе. [30] Более светлая кожа и светлые волосы также появились у древнего населения Северной Евразии . [31]
Дальнейшая волна светлокожих популяций по всей Европе (и в других местах) связана с ямной культурой и индоевропейскими миграциями, имеющими древнее североевразийское происхождение и аллель KITLG для светлых волос. Более того, ген SLC24A5, связанный со светлой пигментацией у европейцев, был завезен в Восточную Африку из Европы более пяти тысяч лет назад. Эти аллели теперь можно найти у жителей Сан , Эфиопии и Танзании с афро-азиатским происхождением. [25] [32] [33] SLC24A5 , говорящими на в Эфиопии сохраняет значительную частоту среди на семитском и кушитском языках говорящих омотическом , нилотском или нигерско-конголезском по сравнению с группами языках. Предполагается, что он мог попасть в регион в результате миграции из Леванта, что также подтверждается лингвистическими данными. [34] У народа сан он был приобретен в результате взаимодействия со скотоводами Восточной Африки. [35] Между тем, в Восточной Азии и Америке MFSD12 . за более светлый цвет кожи отвечает вариант гена [31] Таким образом, современная связь между тоном кожи и широтой возникла относительно недавно. [23]
По данным Кроуфорда и др. (2017), большинство генетических вариантов, связанных со светлой и темной пигментацией, по-видимому, возникли более 300 000 лет назад. [36] Африканские, южноазиатские и австрало-меланезийские популяции также несут производные аллели темной пигментации кожи, которые не встречаются у европейцев или жителей Восточной Азии. [32] Хуанг и др. (2021) обнаружили существование «селективного давления на светлую пигментацию у предкового населения европейцев и жителей Восточной Азии» до их расхождения друг от друга. Также было обнаружено, что на пигментацию кожи влияет направленный отбор в сторону более темной кожи у африканцев, а также более светлой кожи у евразийцев. [37] Кроуфорд и др. (2017) аналогичным образом обнаружили доказательства отбора в сторону светлой пигментации до расхождения жителей Западной Евразии и Восточной Азии. [32]
Мутация A111T в гене SLC24A5 преобладает в популяциях западно-евразийского происхождения. Географическое распространение показывает, что оно почти зафиксировано во всей Европе и на большей части Ближнего Востока, простираясь на восток до некоторых групп населения в современном Пакистане и Северной Индии. Он демонстрирует широтное снижение к экватору с высокими частотами в Северной Африке (80%) и средними (40–60%) в Эфиопии и Сомали. [25]
Древние популяции
[ редактировать ]Некоторые авторы выразили осторожность в отношении прогнозов SNP по пигментации кожи в группах раннего палеолита. По данным Джу и др. (2021): «Относительно темная пигментация кожи в Европе раннего верхнего палеолита могла бы соответствовать тому, что эти популяции были относительно плохо адаптированы к условиям высоких широт в результате недавней миграции из более низких широт. С другой стороны, хотя мы показали, что Эти популяции несли несколько аллелей светлой пигментации, которые выделяются в современной Европе. Они могли нести разные аллели, которые мы сейчас не можем обнаружить. Крайний пример: неандертальцы и алтайские денисовцы демонстрируют генетические показатели, находящиеся в одинаковом диапазоне. к людям раннего верхнего палеолита, но весьма вероятно, что эти популяции, жившие в высоких широтах в течение сотен тысяч лет, независимо адаптировались к низким уровням УФ-излучения. По этой причине мы не можем с уверенностью делать заявления о пигментации кожи у людей. древнее население». [38]
В 2015 году было обнаружено, что образцы кавказских охотников-собирателей (CHG) из Грузии возрастом 13 000 лет несут мутацию и производные аллели пигментации очень светлой кожи, аналогичные ранним фермерам (EF). Говорят, что эта черта имеет относительно долгую историю в Евразии и стала часто встречаться во время неолитической экспансии , причем ее происхождение, вероятно, предшествовало последнему ледниковому максимуму (LGM). [39]
В том же году исследование показало, что гены, способствующие светлой коже, были почти зафиксированы у анатолийских фермеров эпохи неолита: «Вторым по силе сигналом в нашем анализе является производный аллель rs16891982 в SLC45A2, который способствует светлой пигментации кожи и почти зафиксировано у современных европейцев, но встречается с гораздо меньшей частотой в древних популяциях. Напротив, производный аллель SLC24A5, который является другой основной детерминантой светлой пигментации кожи в современной Европе, по-видимому, зафиксирован в анатолийском неолите, что позволяет предположить, что его быстрое увеличение. частота примерно до 0,9 (90%) в Европе раннего неолита была в основном связана с миграцией». [40]
В 2018 году было опубликовано исследование, показывающее, что многие скандинавы позднего мезолита, жившие 9500 лет назад в Северной Европе, имели светлые волосы и светлую кожу, что контрастировало с некоторыми из их современников, более темными западными охотниками-собирателями (WHG). [41] Тем не менее, в статье 2024 года было обнаружено, что фенотипически большинство изученных ими особей WHG имели темную кожу и голубые глаза, характерные для WHG, но некоторые другие WHG во Франции, которые они секвенировали, также имели пигментацию кожи от бледной до средней. [42] Другая запись в 2018 году показала, что восточные охотники-собиратели (EHG), скандинавские охотники-собиратели (SHG) и балтийские собиратели имели производные аллели светлой пигментации кожи. [43]
Исследование популяций энеолитического Леванта (6000-7000 лет назад) показало, что аллель rs1426654 в гене SLC24A5 , который является одной из наиболее важных детерминант светлой пигментации у жителей Западной Евразии, был зафиксирован для производных вариантов во всем Леванте. Образцы энеолита, позволяющие предположить, что светлокожий фенотип мог быть обычным явлением в сообществе. У этих людей также была высокая частота геномных маркеров, связанных с цветом голубых глаз. [44] [45]
В статье, проведенной Фрегелем, Розой и др. (2018) показали, что в Северной Африке марокканцы позднего неолита имели европейскую/кавказскую мутацию SLC24A5 , а также другие аллели и гены, которые предрасполагают людей к более светлому цвету кожи и глаз. [46]
Географическое распространение; ультрафиолет и витамин D
[ редактировать ]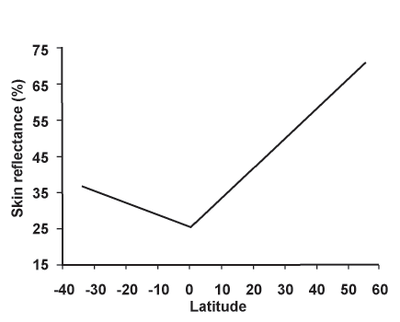

В 1960-х годах биохимик У. Фарнсворт Лумис предположил, что цвет кожи связан с потребностью организма в витамине D. Основным положительным эффектом УФ-излучения у наземных позвоночных является способность синтезировать витамин D3 из него . Определенное количество витамина D помогает организму усваивать больше кальция , который необходим для построения и поддержания костей, особенно для развития эмбрионов . Производство витамина D зависит от воздействия солнечного света. Люди, живущие в широтах вдали от экватора, развили светлую кожу, чтобы помочь усваивать больше витамина D. Люди со светлой кожей ( тип II ) могут вырабатывать превитамин D3 в своей коже в 5–10 раз быстрее, чем темнокожие ( тип V ). люди. [47] [48] [49] [50] [51]
В 1998 году антрополог Нина Яблонски и ее муж Джордж Чаплин собрали данные спектрометра для измерения уровней УФ-излучения по всему миру и сравнили их с опубликованной информацией о цвете кожи коренного населения более чем 50 стран. Результаты показали очень высокую корреляцию между УФ-излучением и цветом кожи; чем слабее солнечный свет в географическом регионе, тем светлее была кожа коренного населения. Яблонски отмечает, что люди, живущие выше 50 градусов широты, имеют самый высокий риск развития дефицита витамина D. Она предполагает, что у людей, живущих далеко от экватора, развилась светлая кожа, позволяющая производить достаточное количество витамина D зимой при низком уровне ультрафиолетового излучения. Генетические исследования показывают, что светлокожие люди отбирались несколько раз. [52] [53] [54]

Полярные регионы, витамин D и диета
[ редактировать ]
Полярные регионы Северного полушария большую часть года получают мало УФ-излучения и еще меньше УФВ-излучения, производящего витамин D. Эти регионы были необитаемы людьми примерно 12 000 лет назад. (По крайней мере, в северную Фенноскандию человеческое население прибыло вскоре после таяния ледников.) [55] В таких регионах, как Скандинавия и Сибирь , очень низкая концентрация ультрафиолетового излучения, а все коренное население светлокожее. [2] [48]
Однако диетические факторы могут обеспечивать достаточность витамина D даже у темнокожих людей. [56] [57] Многие коренные народы Северной Европы и Северной Азии выживают за счет потребления оленей , за которыми они следуют и выпасают . Мясо, органы и жир северного оленя содержат большое количество витамина D, который олени получают, поедая значительное количество лишайника . [58] Некоторые народы полярных регионов , например инуиты ( эскимосы ), сохранили тёмную кожу; , богатые витамином D они ели морепродукты , такие как рыба и жир морских млекопитающих . [59]
Более того, эти люди живут на Крайнем Севере менее 7000 лет. Поскольку в их популяциях-основателях отсутствовали аллели светлого цвета кожи, у них, возможно, не было достаточно времени для того, чтобы значительно более низкое производство меланина было выбрано природой после того, как они были введены случайными мутациями. [60] «Это был один из последних барьеров в истории человеческих поселений», — заявляет Яблонски. «Только после того, как люди научились ловить рыбу и, следовательно, получили доступ к пище, богатой витамином D, они смогли заселить регионы высоких широт ». Кроме того, весной инуиты получают высокий уровень ультрафиолетового излучения в результате отражения от снега, а их относительно темная кожа защищает их от солнечного света. [2] [15] [11]
Более ранние гипотезы
[ редактировать ]Для объяснения развития светлой пигментации кожи были выдвинуты две другие основные гипотезы: устойчивость к холодовому повреждению и генетический дрейф; сейчас считается, что оба они не являются основным механизмом эволюции светлой кожи. [2]
Гипотеза устойчивости к холодовым травмам утверждала, что темная кожа предпочтительнее в холодном климате вдали от экватора и на больших высотах, поскольку темная кожа больше подвержена обморожению . [61] Было обнаружено, что реакция кожи на экстремально холодный климат на самом деле больше связана с другими аспектами, такими как распределение соединительной ткани и распределение жира. [62] [63] и с реакцией периферических капилляров на разницу температур, а не с пигментацией. [2]
Предположение о том, что темная кожа развилась в отсутствие селективного давления, было выдвинуто гипотезой вероятного эффекта мутации . [64] Основным фактором, инициирующим развитие светлой кожи, считалось следствие генетической мутации без эволюционного отбора . Считалось, что последующее распространение светлой кожи было вызвано ассортативным спариванием. [63] а половой отбор способствовал еще более светлой пигментации у самок. [65] [66] Эта гипотеза была подвергнута сомнению, поскольку можно было бы ожидать более случайных моделей окраски кожи в отличие от наблюдаемой структурной светлой пигментации кожи в областях с низким УФ-излучением. [54] Клинальное . (постепенное) распределение пигментации кожи, наблюдаемое в восточном полушарии и в меньшей степени в западном полушарии, является одной из наиболее значимых характеристик пигментации кожи человека Популяции со все более светлой кожей распределяются по территориям со все более низким уровнем УФ-излучения. [67] [68]
Генетические ассоциации
[ редактировать ]Вариации гена KITL положительно связаны примерно с 20% различий в концентрации меланина между африканскими и неафриканскими популяциями. Один из аллелей гена встречается в 80% популяциях Евразии. [69] [70] Ген ASIP имеет уровень вариаций 75–80% среди евразийских популяций по сравнению с 20–25% в африканских популяциях. [71] Вариации гена SLC24A5 составляют 20–25% различий между темнокожими и светлокожими популяциями Африки. [72] и, по-видимому, возникли совсем недавно, в течение последних 10 000 лет. [73] Полиморфизм Ala111Thr или rs1426654 в кодирующей области гена SLC24A5 достигает фиксации в Европе , но встречается по всему земному шару, особенно среди популяций Северной Африки , Африканского Рога , Западной Азии , Центральной Азии и Южной Азии . [74] [75] [76]
Биохимия
[ редактировать ]Меланин является производным аминокислоты тирозина . Эумеланин является доминирующей формой меланина, обнаруженной в коже человека . Эумеланин защищает ткани и ДНК от радиационного повреждения ультрафиолетом . Меланин производится в специализированных клетках, называемых меланоцитами , которые находятся на самом нижнем уровне эпидермиса . [77] Меланин вырабатывается внутри небольших мембранных пакетов, называемых меланосомами . Люди с естественной светлой кожей имеют различное количество более мелкого и редко распределенного эумеланина и его более светлого родственника феомеланина . [52] [78] Концентрация феомеланин сильно варьируется в пределах популяции от индивидуума к индивидууму, но он чаще встречается среди слабопигментированных европейцев, жителей Восточной Азии и коренных американцев. [24] [79]
В одной и той же части тела люди, независимо от цвета кожи, имеют одинаковое количество меланоцитов (однако различия между разными частями тела значительны), но органеллы, содержащие пигменты, называемые меланосомами, у светлокожих людей меньше и менее многочисленны. [80]
У людей с очень светлой кожей кожа приобретает большую часть своего цвета за счет голубовато-белой соединительной ткани дермы и связанных гемоглобином с клеток крови , циркулирующих в капиллярах дермы. Цвет, связанный с циркулирующим гемоглобином, становится более очевидным, особенно на лице, когда артериолы расширяются и наполняются кровью в результате длительной физической нагрузки или стимуляции симпатической нервной системы (обычно смущение или гнев ). [81] До 50% UVA могут проникать глубоко в дерму у людей со светлой пигментацией кожи и небольшим количеством защитного пигмента меланина. [58]
Сочетание светлой кожи, рыжих волос и веснушек связано с высоким содержанием феомеланина и небольшим количеством эумеланина. Этот фенотип вызван мутацией потери функции в гене рецептора меланокортина 1 (MC1R). [82] [83] Однако вариации в последовательности гена MC1R оказывают значительное влияние на пигментацию только в популяциях, где преобладают рыжие волосы и очень светлая кожа. [54] Основной эффект вариации гена заключается в стимулировании синтеза эумеланина за счет синтеза феомеланина, хотя это способствует очень небольшим различиям в отражательной способности кожи между разными этническими группами. [84] Меланоциты из светлых клеток кожи, совместно культивированные с кератиноцитами, приводят к типу распределения, характерному для светлой кожи. [85]
Веснушки обычно возникают только у людей с очень слабопигментированной кожей. Они варьируются от очень темного до коричневого цвета и образуют случайный рисунок на коже человека. [86] Солнечные лентиго , другой вид веснушек, встречаются у пожилых людей независимо от цвета кожи. [2] Люди с очень светлой кожей ( типы I и II ) производят очень мало меланина в своих меланоцитах и имеют очень малую способность или вообще не производят меланин под воздействием УФ-излучения. [87] Это может привести к частым солнечным ожогам и более опасным, но невидимым повреждениям соединительной ткани и ДНК, лежащей под кожей. Это может способствовать преждевременному старению и раку кожи . [88] [89] Сильно красный цвет слегка пигментированной кожи в ответ на высокие уровни УФ-излучения вызван увеличением диаметра, количества и кровотока капилляров. [24]
Люди с умеренно пигментированной кожей ( типы III-IV ) способны вырабатывать меланин в коже в ответ на УФ-излучение. Нормальный загар обычно задерживается, поскольку меланину требуется время, чтобы подняться в эпидермис . Сильный загар не достигает фотозащитного эффекта от повреждения ДНК, вызванного УФР, по сравнению с естественной темной кожей . [90] [91] однако он обеспечивает отличную защиту от сезонных колебаний УФ-излучения. Постепенно развивающийся загар весной предотвращает солнечные ожоги летом. Этот механизм почти наверняка является эволюционной причиной развития загара. [2]
Последствия для здоровья
[ редактировать ]Пигментация кожи — это эволюционная адаптация к различным уровням УФ-излучения по всему миру. Светлокожие люди, живущие в условиях высокого УФ-излучения, имеют последствия для здоровья. Различные культурные традиции усугубляют проблемы, связанные со здоровьем светлокожих, например, загорание среди светлокожих. [2]
Преимущества при слабом солнечном свете
[ редактировать ]У людей со светлой пигментацией кожи, живущих в условиях слабого солнечного света, синтез витамина D увеличивается по сравнению с людьми с темной пигментацией кожи из-за способности поглощать больше солнечного света. Почти каждая часть человеческого тела, включая скелет, иммунную систему и мозг, нуждается в витамине D. Производство витамина D в коже начинается, когда УФ-излучение проникает в кожу и взаимодействует с холестериноподобной молекулой, образуя превитамин D3. Эта реакция происходит только в присутствии УФР средней длины, УФВ. Большинство лучей UVB и UVC разрушаются или отражаются озоном, кислородом и пылью в атмосфере. UVB достигает поверхности Земли в наибольшем количестве, когда его путь прямой и проходит через небольшой слой атмосферы.
Чем дальше место находится от экватора, тем меньше ультрафиолетового излучения поступает, и снижается потенциал выработки витамина D. Некоторые регионы, расположенные далеко от экватора, вообще не получают UVB-излучения в период с осени по весну. [58] Дефицит витамина D не убивает своих жертв быстро и, как правило, не убивает вообще. Скорее, он ослабляет иммунную систему, кости и ставит под угрозу способность организма бороться с неконтролируемым делением клеток, что приводит к раку. Форма витамина D является мощным ингибитором роста клеток; таким образом, хронический дефицит витамина D, по-видимому, связан с более высоким риском развития некоторых видов рака. Это активная тема исследований рака, и она до сих пор обсуждается. [58] Дефицит витамина D, связанный с темной кожей, приводит к более высокому уровню шизофрении в таких группах населения, проживающих в северных широтах. [92]
С увеличением синтеза витамина D снижается частота заболеваний, связанных с распространенными состояниями дефицита витамина D у людей с темной пигментацией кожи, живущих в условиях низкого УФ-излучения: рахит , остеопороз , многочисленные виды рака (включая рак толстой кишки и молочной железы). рак ) и сбои в работе иммунной системы. Витамин D способствует выработке кателицидина , который помогает защитить организм человека от грибковых, бактериальных и вирусных инфекций , включая грипп . [2] [3] При воздействии ультрафиолета B вся открытая часть кожи тела человека с относительно светлой кожей способна вырабатывать от 10 до 20 000 МЕ витамина D. [58]
Недостатки при ярком солнечном свете
[ редактировать ]
Светлокожие люди, живущие в условиях яркого солнечного света, более восприимчивы к вредным ультрафиолетовым лучам солнечного света из-за недостатка меланина , вырабатываемого кожей. Наиболее распространенным риском, связанным с чрезмерным воздействием солнечного света, является повышенный риск солнечных ожогов . Этот повышенный риск связан с культурной практикой принятия солнечных ванн, которая популярна среди светлокожего населения. Эта культурная практика получения загорелой кожи, если ее не регулировать должным образом, может привести к солнечным ожогам, особенно среди людей с очень светлой кожей. Чрезмерное воздействие солнечного света также может привести к базальноклеточной карциноме , которая является распространенной формой рака кожи .
Еще одним последствием для здоровья является истощение запасов фолиевой кислоты в организме, при этом чрезмерное воздействие ультрафиолета может привести к мегалобластной анемии . Дефицит фолиевой кислоты у беременных женщин может нанести вред здоровью их новорожденных детей в виде дефектов нервной трубки , выкидышей и расщелины позвоночника — врожденного дефекта, при котором позвоночник и позвоночный канал не закрываются до рождения. [93] Пик возникновения дефектов нервной трубки является самым высоким в период май-июнь в Северном полушарии . [2] Фолат необходим для репликации ДНК в делящихся клетках, и его дефицит может привести к нарушениям нормального эмбриогенеза и сперматогенеза . [2] [3] [48]
У людей со слабопигментированной кожей, неоднократно подвергающихся сильному УФ-излучению, наблюдается более быстрое старение кожи, что проявляется в увеличении морщин и аномалиях пигментации. Окислительное повреждение вызывает деградацию защитной ткани дермы , которая придает коже прочность. [24] Было высказано предположение, что у белых женщин морщины могут появиться быстрее, чем у чернокожих, после менопаузы потому что белые женщины более восприимчивы к солнечному излучению на протяжении всей жизни. Доктор Хью С. Тейлор из Йельской школы медицины пришел к выводу, что исследование не может доказать полученные результаты, но они подозревают основную причину. Предполагается, что светлая кожа является одним из факторов, способствующих образованию морщин. [94] [95]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ «Светлокожий» . thefreedictionary.com . Проверено 24 января 2017 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Яблонски, Нина Г. (29 июля 2010 г.), Мюленбейн, Майкл П. (редактор), «Окраска кожи» , «Эволюционная биология человека » (1-е изд.), Cambridge University Press, стр. 192–213, doi : 10.1017/cbo9780511781193 .016 , ISBN 978-0-521-70510-3 , получено 1 июня 2024 г.
- ^ Jump up to: а б с О'Нил, Деннис. «Адаптация цвета кожи» . Биологическая адаптивность человека: цвет кожи как адаптация . Паломар. Архивировано из оригинала 18 декабря 2012 года . Проверено 10 декабря 2012 г.
- ^ Хоу, Сен (март 2024 г.). «Цвет кожи китаянок в разных регионах Китая: анализ, основанный как на индивидуальном типологическом угле, так и на угле оттенка» . Журнал дерматологических наук и косметических технологий . 1 (1). дои : 10.1016/j.jdsct.2024.100003 .
- ^ Чо, Чанхуэй (январь 2015 г.). «Сравнение цвета кожи двух азиатских популяций: в зависимости от широты и воздействия ультрафиолета» . Журнал косметической дерматологии . 14 (1): 22–26. дои : 10.1111/jocd.12130 . ПМИД 25573440 .
- ^ Ву, Юэ (июль 2020 г.). «Объективное измерение и сравнение цвета кожи лица человека у женщин Восточной Азии» . Исследования кожи и технологии . 26 (4): 584–590. дои : 10.1111/srt.12838 . ПМИД 31943387 .
- ^ Релетфорд, Джон (1997). Основы биологической антропологии . Издательская компания Мэйфилд. п. 270. ИСБН 978-1559346672 .
- ^ Оксфордские словари. Апрель 2010 г. Издательство Оксфордского университета. «принадлежащий к группе людей со светлой кожей или обозначающий ее», «белый» (по состоянию на 6 августа 2012 г.).
- ^ Dictionary.com: белый 3.a «отмечен легкой пигментацией кожи»
- ^ «Всемирная перепись» . Американская антропологическая ассоциация. Архивировано из оригинала 14 сентября 2018 года . Проверено 10 декабря 2012 г.
- ^ Jump up to: а б Кирхвегер, Джина. «Биология цвета кожи: черное и белое» . Библиотека эволюции . ПБС . Проверено 22 сентября 2018 г.
- ^ Вольф, С. Тони; Кенни, В. Ларри (1 сентября 2019 г.). «Гипотеза витамина D-фолата о здоровье сосудов человека» . Американский журнал физиологии. Регуляторная, интегративная и сравнительная физиология . 317 (3). Американское физиологическое общество: R491–R501. дои : 10.1152/ajpregu.00136.2019 . ISSN 0363-6119 . ПМК 6766707 . ПМИД 31314544 .
- ^ Цуй, Сяоин; Дж. МакГрат, Джон; Х. Дж. Бёрн, Томас (26 января 2021 г.). «Витамин D и шизофрения: 20 лет спустя» . Природа . 26 (7): 2708–2720. дои : 10.1038/s41380-021-01025-0 . ПМЦ 8505257 . ПМИД 33500553 .
- ^ Аппенцеллер, Тим (2012). «Человеческие миграции: Восточная одиссея» . Природа . 485 (7396): 24–26. Бибкод : 2012Natur.485...24A . дои : 10.1038/485024а . ПМИД 22552074 .
- ^ Jump up to: а б с Релетфорд, Дж. Х. (2000). «Разнообразие цвета кожи человека является самым высоким среди населения Африки к югу от Сахары». Биология человека; Международный отчет об исследованиях . 72 (5): 773–80. ПМИД 11126724 .
- ^ Джонс, П.; Лукок, М.; Вейси, М.; Беккет, Э. (2018). «Гипотеза витамина D⁻фолата как эволюционная модель пигментации кожи: обновление и интеграция современных идей» . Питательные вещества . 10 (5): 554. дои : 10.3390/nu10050554 . ПМЦ 5986434 . ПМИД 29710859 .
- ^ «Современная человеческая вариация: обзор» . Архивировано из оригинала 5 ноября 2012 года.
- ^ Гюнтер, Торстен; Мальмстрем, Хелена; Свенссон, Эмма М.; Омрак, Айча; Санчес-Кинто, Федерико; Кылынч, Гюльшах М.; Кшевиньская, Майя; Эрикссон, Гунилла; Фрейзер, Магдалена; Эдлунд, Ханна; Мунтерс, Ариэль Р. (9 января 2018 г.). «Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам» . ПЛОС Биология . 16 (1). Из подтверждающего информационного документа S8, стр. 5/28. дои : 10.1371/journal.pbio.2003703 . ISSN 1545-7885 . ПМК 5760011 . ПМИД 29315301 .
Геномные данные позволили нам дополнительно изучить внешний вид ГСП; например, они демонстрируют сочетание цвета глаз от голубого до светло-коричневого и светлой пигментации кожи. Это разительно отличается от WHG, которым предлагалось иметь специфическое сочетание голубых глаз и темной кожи, и от EHG, которым предлагалось быть кареглазыми и светлокожими.
- ^ Джу, Дэн; Мэтисон, Ян (2021). «Эволюция вариаций, связанных с пигментацией кожи, в Западной Евразии» . ПНАС . 118 (1): e2009227118. Бибкод : 2021PNAS..11809227J . дои : 10.1073/pnas.2009227118 . ПМЦ 7817156 . ПМИД 33443182 .
Относительно темная пигментация кожи в Европе раннего верхнего палеолита могла бы соответствовать тому, что эти популяции были относительно плохо адаптированы к условиям высоких широт в результате недавней миграции из более низких широт. С другой стороны, хотя мы показали, что эти популяции несли несколько аллелей светлой пигментации, которые выделяются в современной Европе, они могли нести и другие аллели, которые мы сейчас не можем обнаружить.
- ^ Вит, Рейнхольд (2003). «Влияние витамина D на кости и естественный отбор цвета кожи: о каком количестве витамина D мы говорим?». В Агарвале Сабрина К.; Стаут, Сэмюэл Д. (ред.). Потеря костной массы и остеопороз: антропологический взгляд . Нью-Йорк: Kluwer Academic / Plenum Press. стр. 139–154. дои : 10.1007/978-1-4419-8891-1_9 . ISBN 978-0-306-47767-6 .
- ^ Хэчкок, JN; Шао, А.; Вит, Р.; Хини, Р.; и др. (2007). «Оценка риска витамина D» . Американский журнал клинического питания . 72 (1): 451–462. дои : 10.1093/ajcn/85.1.6 . ПМИД 17209171 .
- ^ Кимбалл, Саманта; Фулейхан, Гада Эль-Хадж; Вит, Рейнхольд (2008). «Витамин D: перспектива роста». Критические обзоры клинических лабораторных наук . 45 (4): 339–414. дои : 10.1080/10408360802165295 . ISSN 1040-8363 . ПМИД 18568854 . S2CID 57808076 .
- ^ Jump up to: а б с Ханель, Андреа; Карлберг, Карстен (2020). «Цвет кожи и витамин D: обновленная информация» . Экспериментальная дерматология . 29 (9): 864–875. дои : 10.1111/exd.14142 . ПМИД 32621306 . S2CID 220335539 .
- ^ Jump up to: а б с д Яблонски, Нина Г. (октябрь 2004 г.). «Эволюция кожи человека и цвета кожи». Ежегодный обзор антропологии . 33 (1): 585–623. дои : 10.1146/annurev.anthro.33.070203.143955 . ISSN 0084-6570 .
- ^ Jump up to: а б с д Кэнфилд, Виктор А.; Берг, Артур; Пекинс, Стивен; Вентцель, Стивен М.; Анг, Кай Чунг; Оппенгеймер, Стивен; Ченг, Кейт К. (1 ноября 2013 г.). «Молекулярная филогеография аутосомного локуса цвета кожи человека в условиях естественного отбора» . G3: Гены, геномы, генетика . 3 (11): 2059–2067. дои : 10.1534/g3.113.007484 . ISSN 2160-1836 . ПМК 3815065 . ПМИД 24048645 .
- ^ Басу Маллик, Чандана; Илиеску, Флорин Мирча; Мёлс, Мярт; Хилл, Сара; Таманг, Ракеш; Чаубей, Гьянешвер; Гото, Рие; Хо, Саймон Ю.В.; Гальего Ромеро, Ирен; Кривелларо, Федерика; Худжашов, Георгий; Рай, Нирадж; Мецпалу, Мейт; Мэйси-Тейлор, К.Г. Николас; Питчаппан, Рамасами; Сингх, Лалджи; Миразон-Лар, Марта; Тангарадж, Кумарасами; Виллемс, Ричард; Кивисилд, Тоомас (7 ноября 2013 г.). «Аллель светлой кожи SLC24A5 у жителей Южной Азии и европейцев разделяет идентичность по происхождению» . ПЛОС Генетика . 9 (11): e1003912. дои : 10.1371/journal.pgen.1003912 . ISSN 1553-7404 . ПМЦ 3820762 . ПМИД 24244186 .
- ^ Юаса, И.; Уметсу, К.; Харихара, С.; Кидо, А.; Миёси, А.; Сайто, Н.; Дашням, Б.; Джин, Ф.; Люкотт, Г.; Чаттопадхьяй, ПК; Хенке, Л.; Хенке, Дж. (ноябрь 2006 г.). «Распространение аллели F374 гена SLC45A2 (MATP) и анализ гаплотипов-основателей». Анналы генетики человека . 70 (6): 802–811. дои : 10.1111/j.1469-1809.2006.00261.x . ISSN 0003-4800 . ПМИД 17044855 .
- ^ Джонс, Эппи Р.; Гонсалес-Фортес, Глория; Коннелл, Сара; Сиска, Вероника; Эрикссон, Андерс; Мартиниано, Руи; Маклафлин, Рассел Л.; Гальего Льоренте, Маркос; Кэссиди, Лара М.; Гамба, Кристина; Мешвелиани, Тенгиз; Бар-Йосеф, Офер; Мюллер, Вернер; Белфер-Коэн, Анна; Мацкевич, Зиновий; Джакели, Нино; Хайэм, Томас Ф.Г.; Куррат, Матиас; Лордкипанидзе, Давид; Хофрейтер, Михаэль; Маника, Андреа; Пинхаси, Рон; Брэдли, Дэниел Г. (16 ноября 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 (1): 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ISSN 2041-1723 . ПМК 4660371 . ПМИД 26567969 .
- ^ Кроуфорд, Николас Г.; Келли, Дерек Э.; Хансен, Мэтью Э.Б.; Бельтрам, Марсия Х.; Фань, Шаохуа; Боуман, Шанна Л.; Джуэтт, Итан; Ранчиаро, Алессия; Томпсон, Саймон; Ло, Янси; Пфайфер, Сюзанна П.; Дженсен, Джеффри Д.; Кэмпбелл, Майкл С.; Беггс, Уильям; Хормоздиари, Фархад; Мполока, Сунунгуко Вата; Моконе, Гаоньядиве Джордж; Ньямбо, Томас; Мескель, Давит Вольде; Белай, Гурджа; Хаут, Джейк; Программа сравнительного секвенирования NISC; Ротшильд, Харриет; Зон, Леонард; Чжоу, И; Ковач, Майкл А.; Сюй, Май; Чжан, Тунву; Бишоп, Кевин; Синклер, Джейсон; Ривас, Сесилия; Эллиот, Юджин; Чхве, Джиён; Ли, Шэнчао А.; Хикс, Белинда; Берджесс, Шон; Абнет, Кристиан; Уоткинс-Чоу, Дон Э.; Океана, Елена; Сун, Юн С.; Эскин, Элеазар; Браун, Кевин М.; Маркс, Майкл С.; Лофтус, Стейси К.; Паван, Уильям Дж.; Йегер, Мередит; Чанок, Стивен; Тишкофф, Сара А. (17 ноября 2017 г.). «Локусы, связанные с пигментацией кожи, выявленные у африканских популяций» . Наука . 358 (6365). doi : 10.1126/science.aan8433 . ISSN 0036-8075 . ПМЦ 5759959 . ПМИД 29025994 .
На основе коалесцентного анализа с данными последовательностей из Проекта геномного разнообразия Саймонса (SGDP) время появления самого недавнего общего предка (TMRCA) большинства евразийских линий, содержащих аллель rs1426654 (A), составляет 29 тысяч лет назад (ka) [95 % критического интервала (CI), от 28 до 31 тыс. лет назад], что соответствует предыдущим исследованиям.
- ^ Пашу, Перистера; Дриней, Петрос; Яннаки, Евангелия; Разу, Анна; Канаки, Катерина; Цецос, Фотис; Падхманабуни, Шанмукха; Михалодимитракис, Манолис; Ренда, Мария; Паволович, Соня; Анагностопулос, Ахиллес; Стаматояннопулос, Джон; Кидд, Кеннет; Стаматояннопулос, Джордж (24 июня 2014 г.). «Морской путь колонизации Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 111 (25): 9211–9216. Бибкод : 2014PNAS..111.9211P . дои : 10.1073/pnas.1320811111 . ПМК 4078858 . ПМИД 24927591 .
- ^ Jump up to: а б Даунс, Наташа (21 января 2019 г.). «Генетические исследования дают новое представление об эволюции цвета кожи» . Новости UCL (пресс-релиз). Университетский колледж Лондона . Проверено 4 декабря 2021 г.
- ^ Jump up to: а б с Кроуфорд, Николас Г.; Келли, Дерек Э.; Хансен, Мэтью Э.Б.; Бельтрам, Марсия Х.; Фань, Шаохуа; Боуман, Шанна Л.; Джуэтт, Итан; Ранчиаро, Алессия; Томпсон, Саймон; Ло, Янси; Пфайфер, Сюзанна П.; Дженсен, Джеффри Д.; Кэмпбелл, Майкл С.; Беггс, Уильям; Хормоздиари, Фархад (17 ноября 2017 г.). «Локусы, связанные с пигментацией кожи, выявленные у африканских популяций» . Наука . 358 (6365): eaan8433. дои : 10.1126/science.aan8433 . ISSN 1095-9203 . ПМЦ 5759959 . ПМИД 29025994 .
- ^ Фэн, Юаньцин; Маккуиллан, Майкл А.; Тишкофф, Сара А. (26 апреля 2021 г.). «Эволюционная генетика пигментации кожи у африканских популяций» . Молекулярная генетика человека . 30 (R1): R88–R97. дои : 10.1093/hmg/ddab007 . ISSN 1460-2083 . ПМЦ 8117430 . ПМИД 33438000 .
- ^ Пагани, Лука; Кивисилд, Томас; Тарекегн, Айеле; Эконг, Розмари; Пластырь, Крис; Гальего Розмари, Ирен; Иов, Касим; Мехди, С. Касим; Томас, Марк Г.; Луиза, Донна; Бекеле, Эндашоу (13 июля 2012 г.). «Генетическое разнообразие Эфиопии свидетельствует о языковой стратификации и сложном влиянии на генофонд Эфиопии» . Американский журнал генетики человека . 91 (1): 83–96. дои : 10.1016/j.ajhg.2012.05.015 . ISSN 1537-6605 . ПМЦ 3397267 . ПМИД 22726845 .
- ^ Линь, Мэн; Сифорд, Ребекка Л.; Мартин, Алисия Р.; Накагоме, Сигэки; Мёллер, Марло; Хоал, Эйлин Г.; Бустаманте, Карлос Д.; Жиньу, Кристофер Р.; Хенн, Бренна М. (26 декабря 2018 г.). «Быстрая эволюция аллеля осветления кожи в южноафриканском Кхоесане» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (52): 13324–13329. Бибкод : 2018PNAS..11513324L . дои : 10.1073/pnas.1801948115 . ISSN 1091-6490 . ПМК 6310813 . ПМИД 30530665 .
- ^ Бэйли, Кэтрин Унгер (12 октября 2017 г.). «Идентифицированы гены, ответственные за разнообразие цветов кожи человека» . Penn Today (пресс-релиз). Пенсильванский университет. Переиздано ScienceDaily .
- ^ Хуан, Синь (2021). «Анализ динамики и различий селективного давления в эволюции пигментации человека» . Биология Открытая . 10 (2). дои : 10.1242/bio.056523 . ПМЦ 7888712 . ПМИД 33495209 .
- ^ Джу, Дэн; Мэтисон, Ян (2021). «Эволюция вариаций, связанных с пигментацией кожи, в Западной Евразии» . ПНАС . 118 (1): e2009227118. Бибкод : 2021PNAS..11809227J . дои : 10.1073/pnas.2009227118 . ПМЦ 7817156 . ПМИД 33443182 .
- ^ Джонс, Эппи Р.; Гонсалес-Фортес, Глория; Коннелл, Сара; Сиска, Вероника; Эрикссон, Андерс; Мартиниано, Руи; Маклафлин, Рассел Л.; Гальего Льоренте, Маркос; Кэссиди, Лара М.; Гамба, Кристина; Мешвелиани, Тенгиз; Бар-Йосеф, Офер; Мюллер, Вернер; Белфер-Коэн, Анна; Мацкевич, Зиновий (16 ноября 2015 г.). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 (1): 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . hdl : 2262/76623 . ISSN 2041-1723 .
- ^ Мэтисон, Иэн; Лазаридис, Иосиф; Роланд, Надин; Маллик, Свапан; Паттерсон, Ник; Руденберг, Сонгюль Алпаслан; Харни, Эдаоин; Стюардсон, Кристин; Фернандес, Даниэль; Новак, Марио; Сирак, Кендра; Гамба, Кристина; Джонс, Эппи Р.; Ламас, Бастьен; Дрёмов, Станислав (декабрь 2015 г.). «Общегеномные закономерности отбора у 230 древних евразийцев» . Природа . 528 (7583): 499–503. Бибкод : 2015Natur.528..499M . дои : 10.1038/nature16152 . ISSN 1476-4687 . ПМЦ 4918750 . ПМИД 26595274 .
- ^ Гюнтер, Торстен; Мальмстрем, Хелена; Свенссон, Эмма М.; Омрак, Айча; Санчес-Кинто, Федерико; Кылынч, Гюльшах М.; Кшевиньская, Майя; Эрикссон, Гунилла; Фрейзер, Магдалена; Эдлунд, Ханна; Мунтерс, Ариэль Р.; Коутиньо, Александра; Симоэнс, Лусиана Г.; Висенте, Марио; Шёландер, Андерс (9 января 2018 г.). «Популяционная геномика мезолитической Скандинавии: исследование маршрутов ранней послеледниковой миграции и адаптации к высоким широтам» . PLoS Биология . 16 (1): e2003703. дои : 10.1371/journal.pbio.2003703 . ISSN 1545-7885 . ПМК 5760011 . ПМИД 29315301 .
- ^ Симоэнс, Лусиана Г.; Пейротео-Стьерна, Рита; Маршан, Грегор; Бернхардссон, Каролина; Виалет, Амели; Четти, Даршан; Алачамлы, Эркин; Эдлунд, Ханна; Букен, Дени; Дина, Кристиан; Гармонд, Николас; Гюнтер, Торстен; Якобссон, Маттиас (5 марта 2024 г.). «Геномное происхождение и социальная динамика последних охотников-собирателей Атлантической Франции» . Труды Национальной академии наук . 121 (10): e2310545121. Бибкод : 2024PNAS..12110545S . дои : 10.1073/pnas.2310545121 . ISSN 0027-8424 . ПМЦ 10927518 . ПМИД 38408241 .
- ^ Миттник, Алисса; Ван, Чуан-Чао; Пфрангле, Саския; Даубарас, Мантас; Зариня, Гунита; Халлгрен, Фредрик; Аллмяэ, Райли; Хартанович, Валерий; Моисеев Вячеслав; Тырв, Мари; Фуртвенглер, Аня; Андрадес Валтуэнья, Аида; Фельдман, Михал; Эконому, Христос; Ойнонен, Маркку (30 января 2018 г.). «Генетическая предыстория региона Балтийского моря» . Природные коммуникации . 9 (1): 442. Бибкод : 2018NatCo...9..442M . дои : 10.1038/s41467-018-02825-9 . ISSN 2041-1723 . ПМЦ 5789860 . ПМИД 29382937 .
- ^ Харни, Эдаоин; Мэй, Хила; Шалем, Дина; Роланд, Надин; Маллик, Свапан; Лазаридис, Иосиф; Сариг, Рэйчел; Стюардсон, Кристин; Норденфельт, Сюзанна; Паттерсон, Ник; Гершковитц, Израиль; Райх, Дэвид (20 августа 2018 г.). «Древняя ДНК из халколита Израиля раскрывает роль смешения населения в культурной трансформации» . Природные коммуникации . 9 (1): 3336. Бибкод : 2018NatCo...9.3336H . дои : 10.1038/s41467-018-05649-9 . ISSN 2041-1723 . ПМК 6102297 . ПМИД 30127404 .
- ^ «Анализ ДНК останков человека возрастом 6500 лет с мутацией голубого глаза» . ScienceDaily . Проверено 27 мая 2024 г.
- ^ Фрегель, Роза; Мендес, Фернандо Л.; Бокбот, Юсеф; Мартин-Сокас, Димас; Камалич-Массие, Мария Д.; Сантана, Джонатан; Моралес, Джейкоб; Авила-Аркос, Мария К.; Андерхилл, Питер А.; Шапиро, Бет; Войчик, Женевьева; Расмуссен, Мортен; Соарес, Андре ЭР; Капп, Джошуа; Сокелл, Александра (26 июня 2018 г.). «Древние геномы из Северной Африки свидетельствуют о доисторических миграциях в Магриб как из Леванта, так и из Европы» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (26): 6774–6779. Бибкод : 2018PNAS..115.6774F . дои : 10.1073/pnas.1800851115 . ISSN 1091-6490 . ПМК 6042094 . ПМИД 29895688 .
- ^ Клементс, ТЛ; Адамс, Дж. С.; Хендерсон, СЛ; Холик, МФ; и др. (1982). «Повышение пигментации кожи снижает способность кожи синтезировать витамин D» (PDF) . Ланцет . 1 (8263): 74–76. дои : 10.1016/S0140-6736(82)90214-8 . ПМИД 6119494 . S2CID 41818974 .
- ^ Jump up to: а б с Яблонски, Н.Г.; Чаплин, Г. (2000). «Эволюция окраски кожи человека». Журнал эволюции человека . 39 (1): 57–106. Бибкод : 2000JHumE..39...57J . дои : 10.1006/jhev.2000.0403 . ПМИД 10896812 .
- ^ Уэбб, Арканзас (2006). «Кто, что, где и когда: влияет на кожный синтез витамина D». Прогресс биофизики и молекулярной биологии . 92 (1): 17–25. doi : 10.1016/j.pbiomolbio.2006.02.004 . ПМИД 16766240 .
- ^ Армас, Луизиана; Доуэлл, С.; Ахтер, М.; Дутулуру, С.; Хуэртер, К.; Холлис, BW; Лунд, Р.; Хини, Р.П.; и др. (2007). «Ультрафиолетовое излучение B повышает уровень 25-гидроксивитамина D в сыворотке: влияние дозы UVB и цвета кожи». Журнал Американской академии дерматологии . 57 (4): 588–593. дои : 10.1016/j.jaad.2007.03.004 . ПМИД 17637484 .
- ^ Чен, TC; и др. (2007). «Факторы, влияющие на кожный синтез и пищевые источники витамина D» . Архив биохимии и биофизики . 460 (2): 213–217. дои : 10.1016/j.abb.2006.12.017 . ПМЦ 2698590 . ПМИД 17254541 .
- ^ Jump up to: а б Ламасон, РЛ; Мохидин, Массачусетс; Мест, младший; Вонг, AC; Нортон, Х.Л.; Арос, MC; Юринец, MJ; Мао, X.; Хамфривилл, Вирджиния; Гумберт, JE; Синха, С.; Мур, Дж.Л.; Джагадисваран, П.; Чжао, В.; Нин, Г.; Макаловская, И.; Маккейг, премьер-министр; О'Доннелл, Д.; Киттлс, Р.; Парра, Э.Дж.; Манджини, Нью-Джерси; Грюнвальд, диджей; Шрайвер, доктор медицины; Кэнфилд, Вирджиния; Ченг, КЦ; и др. (2005). «SLC24A5, предполагаемый катионообменник, влияет на пигментацию у рыбок данио и людей». Наука . 310 (5755): 1782–1786. Бибкод : 2005Sci...310.1782L . дои : 10.1126/science.1116238 . ПМИД 16357253 . S2CID 2245002 .
- ^ Лалуэса-Фокс; Рёмплер, Х.; Карамелли, Д.; Штойберт, Дж.; Каталано, Г.; Хьюз, Д; Роланд, Н.; Пилли, Э.; Лонг, Л.; Кондеми, С.; Расилла, М.; Фортеа, Дж.; Росас, А.; Стоункинг, М.; Шенеберг, Т.; Бертранпети, Ж.; Хофрейтер, М.; и др. (2007). «Аллель рецептора меланокортина-1 предполагает различную пигментацию у неандертальцев». Наука . 318 (5855): 1453–1455. Бибкод : 2007Sci...318.1453L . дои : 10.1126/science.1147417 . ПМИД 17962522 . S2CID 10087710 .
- ^ Jump up to: а б с Нортон, Х.Л.; Киттлс, РА; Парра, Э.; Маккейг, П.; Мао, X.; Ченг, К.; Кэнфилд, Вирджиния; Брэдли, генеральный директор; МакЭвой, Б.; Шрайвер, доктор медицины; и др. (2007). «Генетические доказательства конвергентной эволюции светлой кожи у европейцев и жителей Восточной Азии» . Молекулярная биология и эволюция . 24 (3): 710–722. дои : 10.1093/molbev/msl203 . ПМИД 17182896 .
- ^ Бергман, Ингела; Олофссон, Андерс; Хёрнберг, Грегер; Закриссен, Олле; Хеллберг, Эрик (июнь 2004 г.). «Дегляциация и колонизация: пионерские поселения в северной Фенноскандии». Журнал мировой предыстории . 18 (2): 155–177. дои : 10.1007/s10963-004-2880-z . S2CID 129136655 .
- ^ Бьёрн, Ло; Ван, Т; и др. (2000). «Витамин D в экологическом контексте». Международный журнал циркумполярного здоровья . 59 (1): 26–32. ПМИД 10850004 .
- ^ Ван Дир Меер; Буке, Эй Джей; Липс, П.; Гроотьянс-Геертс, И.; Вуистер, доктор медицинских наук; Девиль, WL; Вилдерс, JP; Баутер, LM; Мидделькооп, Б.Дж.; и др. (2007). «Жирная рыба и пищевые добавки вносят наибольший вклад в концентрацию 25-гидроксивитамина D в сыворотке крови у многоэтнического населения» . Клиническая эндокринология . 68 (3): 466–472. дои : 10.1111/j.1365-2265.2007.03066.x . hdl : 1871/22170 . ПМИД 17941903 . S2CID 15728496 .
- ^ Jump up to: а б с д и Яблонски, Нина (2012). Живой цвет . Беркли, Лос-Анджелес, Лондон: Издательство Калифорнийского университета. ISBN 978-0-520-25153-3 .
- ^ Почему цвет кожи различается. Кафедра физики: факультет математики и естественных наук. Авторы: Йохан Моан, Аста Юзениене.
- ^ «Биологическая адаптивность человека: цвет кожи как адаптация» . www2.palomar.edu .
- ^ Почта; Дэниелс-младший, форвард; Бинфорд-младший, RT; и др. (1975). «Холодовая травма и эволюция «белой» кожи». Биология человека . 47 (1): 65–80. ПМИД 1126703 .
- ^ Стигман, А. Т. младший (1967). «Обморожение человеческого лица как избирательная сила». Биология человека . 39 (2): 131–144. ПМИД 6056270 .
- ^ Jump up to: а б Киттлс, Р. (1995). «Природа, происхождение и вариации пигментации человека». Журнал исследований чернокожих . 26 : 36–61. дои : 10.1177/002193479502600104 . S2CID 145636646 .
- ^ Брейс, CL (1963). «Структурная редукция эволюции». Американский натуралист . 97 (892): 39–49. дои : 10.1086/282252 . S2CID 85732039 .
- ^ Фрост, П. (1988). «Цвет кожи человека: возможная связь между его половым диморфизмом и социальным восприятием». Перспективы биологии и медицины . 32 (1): 38–59. дои : 10.1353/pbm.1988.0010 . ПМИД 3059317 . S2CID 36144428 .
- ^ Аоки, К. (2002). «Половой отбор как причина изменения цвета кожи человека: новый взгляд на гипотезу Дарвина». Анналы биологии человека . 29 (6): 589–608. дои : 10.1080/0301446021000019144 . ПМИД 12573076 . S2CID 22703861 .
- ^ Релетфорд, Дж. Х. (1997). «Разница полушарий в цвете кожи человека» . Американский журнал физической антропологии . 104 (4): 449–457. doi : 10.1002/(SICI)1096-8644(199712)104:4<449::AID-AJPA2>3.0.CO;2-N . ПМИД 9453695 .
- ^ Чаплин, Г.; Яблонски, Н. (1998). «Различия полушарий в цвете кожи человека». Американский журнал физической антропологии . 107 (2): 221–224. doi : 10.1002/(SICI)1096-8644(199810)107:2<221::AID-AJPA8>3.0.CO;2-X . ПМИД 9786336 .
- ^ Миллер, Крейг Т.; Белеза, Сандра; Пыльца, Алекс А.; Шлютер, Дольф; Киттлс, Рик А.; Шрайвер, Марк Д.; Кингсли, Дэвид М. (2007). «Цис-регуляторные изменения в экспрессии Kit-лиганда и параллельная эволюция пигментации у колюшек и людей» . Клетка . 131 (6): 1179–89. дои : 10.1016/j.cell.2007.10.055 . ПМК 2900316 . ПМИД 18083106 .
- ^ HapMap: отчет SNP для rs642742 . Hapmap.ncbi.nlm.nih.gov (19 октября 2009 г.). Проверено 27 февраля 2011 г.
- ^ «Отчет SNP для rs2424984» . Международный проект HapMap . Национальный центр биотехнологической информации США . Проверено 11 декабря 2012 г.
- ^ Ламасон, РЛ; Мохидин, Массачусетс; Мест, младший; Вонг, AC; Нортон, Х.Л.; Арос, MC; Юринец, MJ; Мао, X.; и др. (2005). «SLC24A5, предполагаемый катионообменник, влияет на пигментацию у рыбок данио и людей». Наука . 310 (5755): 1782–17886. Бибкод : 2005Sci...310.1782L . дои : 10.1126/science.1116238 . ПМИД 16357253 . S2CID 2245002 .
- ^ Гиббонс, А. (2007). «ВСТРЕЧА АМЕРИКАНСКОЙ АССОЦИАЦИИ ФИЗИЧЕСКИХ АНТРОПОЛОГОВ: Кожа европейцев побледнела только недавно, - предполагает Джин». Наука . 316 (5823): 364а. дои : 10.1126/science.316.5823.364a . ПМИД 17446367 . S2CID 43290419 .
- ^ «Графическое отображение частот аллелей Ala111Thr» . База данных частот аллелей . Проверено 10 октября 2012 г.
- ^ «АЛЬФРЕД — Информация о полиморфизме — Ala111Thr» . База данных частот аллелей . Проверено 22 сентября 2018 г.
- ^ Пагани, Лука; Томас Кивисилд; Айеле Тарекегн; Розмари Эконг; Крис Пластер; Ирен Гальего Ромеро; Касим Аюб; С. Касим Мехди; Марк Г. Томас; Доната Луиселли; Эндашоу Бекеле; Нил Брэдман; Дэвид Дж. Болдинг; Крис Тайлер-Смит (21 июня 2012 г.). «Генетическое разнообразие в Эфиопии свидетельствует о языковой стратификации и сложном влиянии на генофонд» . Американский журнал генетики человека . 91 (1): Том 91, выпуск 1, 83–96, 21 июня 2012 г. doi : 10.1016/j.ajhg.2012.05.015 . ПМЦ 3397267 . ПМИД 22726845 .
- ^ Хаас и др., 2005.
- ^ Стринги, HY; и др. (2003). «Закономерности распределения меланосом в кератиноцитах кожи человека как один из факторов, определяющих цвет кожи». Британский журнал дерматологии . 149 (3): 498–505. дои : 10.1046/j.1365-2133.2003.05473.x . ПМИД 14510981 . S2CID 43355316 .
- ^ Вондрак, Георг (2016), Пути реакции кожи на стресс: факторы окружающей среды и молекулярные возможности , Springer International Publishing, стр. 159, ISBN 9783319431574 , получено 6 апреля 2020 г.
{{citation}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Сабо, Г.; и др. (1969). «Расовые различия в судьбе меланосом в эпидермисе человека». Природа . 222 (5198): 1081–1082. Бибкод : 1969Natur.222.1081S . дои : 10.1038/2221081a0 . ПМИД 5787098 . S2CID 4223552 .
- ^ Яблонски, Н.Г. (2006). Кожа: естественная история . Беркли, Калифорния: Издательство Калифорнийского университета.
- ^ Штурм, РА; и др. (2003). «Генетическая ассоциация и клеточная функция аллелей варианта MC1R при пигментации человека». Анналы Нью-Йоркской академии наук . 994 (1): 348–358. Бибкод : 2003NYASA.994..348S . дои : 10.1111/j.1749-6632.2003.tb03199.x . ПМИД 12851335 . S2CID 6156245 .
- ^ Рис, Дж.Л. (2003). «Генетика цвета волос и кожи». Ежегодный обзор генетики . 37 : 67–90. дои : 10.1146/annurev.genet.37.110801.143233 . ПМИД 14616056 .
- ^ Алалуф, С.; и др. (2002). «Этнические различия в содержании и составе меланина в фотоэкспонированных и фотозащищенных человеческих сджинах». Исследование пигментных клеток . 15 (2): 112–118. дои : 10.1034/j.1600-0749.2002.1o071.x . ПМИД 11936268 .
- ^ Минвала, С.; и др. (2001). «Кератиноциты играют роль в регулировании закономерностей распределения меланосом-реципиентов in vitro» . Журнал исследовательской дерматологии . 117 (2): 341–347. дои : 10.1046/j.0022-202x.2001.01411.x . ПМИД 11511313 .
- ^ Родос, Арканзас; и др. (1991). «Солнечные веснушки у детей и молодых людей: корреляция клинических и гистопатологических особенностей» . Рак . 67 (7): 1990–2001. doi : 10.1002/1097-0142(19910401)67:7<1990::aid-cncr2820670728>3.0.co;2-p . ПМИД 2004316 .
- ^ Фитцпатрик, ТБ; Ортонн, JP (2003). «Нормальный цвет кожи и общие соображения о пигментных нарушениях». В книге Фитцпатрика «Дерматология в общей медицине» . 6 : 819–825.
- ^ Кливер, Дж. Э.; Кроули, Э. (2002). «УФ-повреждение, восстановление ДНК и канцерогенез кожи» . Границы бионауки . 7 (1–3): 1024–1043. doi : 10.2741/тесак . ПМИД 11897551 .
- ^ Мацумура, Ясухиро; Анантавами, Хоннавара Н. (2004). «Токсическое действие ультрафиолетового излучения на кожу». Токсикология и прикладная фармакология . 195 (3): 298–308. дои : 10.1016/j.taap.2003.08.019 . ПМИД 15020192 .
- ^ Тадокоро, Т.; и др. (2005). «Механизмы загара кожи у разных расовых/этнических групп в ответ на ультрафиолетовое излучение» . Журнал исследовательской дерматологии . 124 (6): 1326–1332. дои : 10.1111/j.0022-202X.2005.23760.x . ПМИД 15955111 .
- ^ Нильсен, КП; и др. (2006а). «Важность глубины распределения меланина в коже для защиты ДНК и других фотобиологических процессов». Журнал фотохимии и фотобиологии B: Биология . 82 (3): 194–198. Бибкод : 2006JPPB...82..194N . doi : 10.1016/j.jphotobiol.2005.11.008 . ПМИД 16388960 .
- ^ Цуй, Сяоин; МакГрат, Джон Дж.; Бёрн, Томас Х.Дж.; Эйлс, Дэррил В. (26 января 2021 г.). «Витамин D и шизофрения: 20 лет спустя» . Природа . 26 (7): 2708–2720. дои : 10.1038/s41380-021-01025-0 . ПМЦ 8505257 . ПМИД 33500553 .
Отдельное наблюдение о том, что потомки мигрантов с темной кожей, которые мигрируют в холодный климат, имеют повышенный риск развития шизофрении, также может быть связано с низким уровнем витамина D во время беременности и в раннем возрасте, поскольку темная кожа требует большего воздействия солнечного света для достижения адекватного уровня витамина D. прегормон.
- ^ Джукич, А. (2007). «Фолитозависимые неврологические заболевания». Детская неврология . 37 (6): 387–397. doi : 10.1016/j.pediatrneurol.2007.09.001 . ПМИД 18021918 .
- ^ Нортон, Эми (10 ноября 2010 г.). «На коже белых женщин быстрее появляются морщины» . Рейтер . Проверено 22 сентября 2018 г.
- ^ Коул, Гэри. «Морщины» . MedicineNet.com . Проверено 22 сентября 2018 г.