Заселение Америки
Заселение Америки началось, когда палеолитические охотники-собиратели ( палеоиндейцы ) проникли в Северную Америку из североазиатской Мамонтовой степи через Берингийский сухопутный мост , образовавшийся между северо-восточной Сибирью и западной Аляской в связи с понижением уровня моря во время последней Ледниковый максимум (26 000–19 000 лет назад). [2] Эти популяции распространились к югу от Лаврентийского ледникового щита и быстро распространились на юг, заселив как Северную, так и Южную Америку , 12–14 000 лет назад. [3] [4] [5] [6] [7] Самые ранние популяции в Америке, возникшие примерно 10 000 лет назад, известны как палеоиндейцы . Коренные народы Америки были связаны с сибирским населением лингвистическими факторами , распределением групп крови и генетическим составом , что отражено молекулярными данными, такими как ДНК . [8] [9]
Хотя существует общее мнение, что Америка была впервые заселена из Азии, характер миграции и место(а) происхождения в Евразии народов, мигрировавших в Америку, остаются неясными. [4] Традиционная теория состоит в том, что древние берингийцы переселились, когда уровень моря значительно снизился из-за четвертичного оледенения . [10] [11] преследуя стада ныне вымершей плейстоценовой мегафауны по свободным ото льда коридорам , простирающимся между ледниковыми щитами Лаврентиды и Кордильеров . [12] Другой предложенный маршрут заключается в том, что пешком или на лодках они мигрировали вдоль побережья Тихого океана в Южную Америку до Чили . [13] Любые археологические свидетельства оккупации побережья во время последнего ледникового периода теперь были бы скрыты повышением уровня моря , которое с тех пор достигло ста метров. [14]
Точная дата заселения Америки является давним открытым вопросом, и хотя достижения в археологии , плейстоцена геологии , физической антропологии и анализе ДНК постепенно проливают больше света на эту тему, важные вопросы остаются нерешенными. [15] [16] «Первая теория Хлодвига» относится к гипотезе о том, что культура Хлодвига представляет собой самое раннее присутствие человека на Америке около 13 000 лет назад. [17] Свидетельства существования культур, существовавших до Хлодвига, накопились и отодвинули возможную дату первого заселения Америки. [18] [19] [20] [21] Академики обычно полагают, что люди достигли Северной Америки к югу от ледникового щита Лаврентида где-то между 15 000 и 20 000 лет назад. [15] [18] [22] [23] [24] [25] Некоторые новые противоречивые археологические данные предполагают, что прибытие людей в Америку могло произойти до последнего ледникового максимума, более 20 000 лет назад. [18] [26] [27] [28] [29] [30]
Окружающая среда во время позднего оледенения
[ редактировать ]Возникновение и затопление Берингии.
[ редактировать ]
Во время оледенения Висконсина вода океана Земли в разной степени с течением времени хранилась в ледниковом льду. По мере накопления воды в ледниках объем воды в океанах соответственно уменьшался, что приводило к понижению глобального уровня моря . Изменение уровня моря во времени было реконструировано с помощью анализа изотопов кислорода глубоководных кернов, датирования морских террас и отбора проб изотопов кислорода с высоким разрешением из океанских бассейнов и современных ледяных шапок. Падение эвстатического уровня моря примерно на 60–120 метров (от 200 до 390 футов) по сравнению с современным уровнем, начавшееся примерно за 30 000 лет до настоящего времени (BP), создало Берингию , прочный и обширный географический объект, соединяющий Сибирь с Аляской. [31] С подъемом уровня моря после Последнего ледникового максимума (LGM) Берингийский сухопутный мост снова оказался под водой. Оценки окончательного повторного затопления Берингийского сухопутного моста, основанные исключительно на современной батиметрии Берингова пролива и эвстатической кривой уровня моря, относят это событие примерно к 11 000 лет назад (рис. 1). Продолжающиеся исследования по реконструкции палеогеографии Берингии во время дегляциации могут изменить эту оценку, а возможное более раннее затопление может еще больше ограничить модели миграции людей в Северную Америку. [31]
Ледники
[ редактировать ]
Наступление последнего ледникового максимума спустя 30 000 лет назад привело к расширению альпийских ледников и континентальных ледниковых щитов, которые заблокировали пути миграции из Берингии. К 21 000 лет назад, а возможно, и на тысячи лет раньше, ледяные щиты Кордильеров и Лаврентидов слились к востоку от Скалистых гор , закрыв потенциальный путь миграции в центр Северной Америки. [32] [33] [34] Альпийские ледники в прибрежных хребтах и на Аляске изолировали внутреннюю часть Берингии от побережья Тихого океана. Прибрежные альпийские ледники и доли кордильерского льда объединились в предгорные ледники , которые покрыли большие участки береговой линии вплоть до острова Ванкувер и образовали ледяную долю через пролив Хуан-де-Фука к 18 000 лет назад. [35] [36] Прибрежные альпийские ледники начали отступать около 19 000 лет назад. [37] в то время как лед Кордильеров продолжал наступать в низменности Пьюджет до 16 800 лет назад. [36] Даже во время максимальной протяженности прибрежных льдов незамерзающие рефугиумы , служащие средой обитания наземных и морских млекопитающих. на современных островах сохранялись [34] По мере таяния ледников рефугиум расширялся, пока побережье не освободилось ото льда к 15 000 лет назад. [34] Отступление ледников на Аляске примерно на 17 000 лет назад обеспечило доступ из Берингии к побережью Тихого океана. [38] Ледяной барьер между внутренней Аляской и побережьем Тихого океана разрушился примерно 16 200 лет назад. [35] Свободный ото льда коридор во внутренние районы Северной Америки открылся между 13 000 и 12 000 лет назад. [32] [33] [34] Оледенение Восточной Сибири во время МГМ ограничивалось альпийскими и долинными ледниками горных хребтов и не блокировало доступ между Сибирью и Берингией. [31]
Климат и биологическая среда
[ редактировать ]
Палеоклиматы и растительность Восточной Сибири и Аляски во время Висконсинского оледенения были определены на основе данных изотопов кислорода высокого разрешения и пыльцы стратиграфии . [31] [39] [40] До последнего ледникового максимума климат в Восточной Сибири колебался между условиями, приближающимися к современным условиям, и более холодными периодами. Теплые циклы, предшествовавшие LGM, в арктической Сибири были отмечены расцветом мегафауны. [31] Записи изотопов кислорода в ледяной шапке Гренландии позволяют предположить, что эти циклы после примерно 45 000 лет назад длились от сотен до одной-двух тысяч лет, причем большая продолжительность холодных периодов начиналась примерно с 32 000 лет назад. [31] Данные о пыльце из озера Эликчан, расположенного к северу от Охотского моря , показывают заметный сдвиг от пыльцы деревьев и кустарников к пыльце трав до 30 000 лет назад, поскольку травяная тундра заменила бореальные леса и кустарниковую степь, перейдя в LGM. [31] Аналогичная запись о замене пыльцы деревьев/кустарников пыльцой трав по мере приближения LGM была обнаружена вблизи реки Колыма в арктической Сибири. [40] Заброшенность северных регионов Сибири из-за быстрого похолодания или отступления охотничьих видов с наступлением ЛГМ было предложено объяснить отсутствие в этом регионе археологических памятников, относящихся к ЛГМ. [40] [41] Запись пыльцы со стороны Аляски показывает сдвиги между травянисто-кустарниковой и кустарниковой тундрой до LGM, что позволяет предположить менее резкие эпизоды потепления, чем те, которые позволили колонизировать леса на сибирской стороне. разнообразная, хотя и не обязательно многочисленная, мегафауна В этих средах присутствовала . В период LGM преобладала травяная тундра из-за холодных и засушливых условий. [39]
Прибрежная среда во время последнего ледникового максимума была сложной. Пониженный уровень моря и изостатическая выпуклость, уравновешенная депрессией под Кордильерским ледниковым щитом, обнажили континентальный шельф, образовав прибрежную равнину. [42] В то время как большая часть прибрежной равнины была покрыта предгорными ледниками, неледниковые рефугиумы, поддерживающие наземных млекопитающих, были обнаружены на Хайда-Гвайи , острове Принца Уэльского и внешних островах архипелага Александра . [39] Ныне затопленная прибрежная равнина имеет потенциал для создания новых убежищ. [39] Данные по пыльце указывают на то, что тундровая растительность в основном разнотравяная/кустарниковая на незамерзающих территориях, с некоторыми бореальными лесами ближе к южному концу хребта Кордильерских льдов. [39] Прибрежная морская среда оставалась продуктивной, о чем свидетельствуют окаменелости ластоногих . [42] Высокопродуктивные леса водорослей на каменистых морских мелководьях могли быть приманкой для прибрежной миграции. [43] [44] Реконструкция южного побережья Берингии также предполагает наличие потенциала высокопродуктивной прибрежной морской среды. [44]
Изменения окружающей среды во время дегляциации
[ редактировать ]
Данные по пыльце указывают на теплый период, достигающий кульминации между 17 000 и 13 000 лет назад, за которым последовало похолодание между 13 000 и 11 500 лет назад. [42] Прибрежные районы быстро исчезли, поскольку прибрежные альпийские ледники, а затем куски кордильерского льда, отступили. Отступление ускорилось, поскольку уровень моря поднялся и всплыли концы ледников. Было подсчитано, что береговой хребет был полностью свободен ото льда между 16 000 и 15 000 лет назад. [42] [34] Прибрежные морские организмы колонизировали береговую линию, когда океанская вода заменила талую ледниковую воду. Замена травянисто-кустарниковой тундры хвойными лесами началась к 15 000 лет назад к северу от Хайда-Гвайи. Эвстатическое повышение уровня моря вызвало наводнение, которое ускорилось по мере увеличения скорости его темпов. [42]
Внутренние ледниковые щиты Кордильеров и Лаврентид отступали медленнее, чем прибрежные ледники. Открытие свободного ото льда коридора произошло только через 13–12 000 лет назад. [32] [33] [34] На начальном этапе существования свободного ото льда коридора преобладали ледниковые смывы и талая вода, с озерами, покрытыми льдом, и периодическими наводнениями в результате выброса талой воды, запруженной льдом. [32] Биологическая продуктивность дегляциального ландшафта возрастала медленно. [34] Самая ранняя возможная жизнеспособность незамерзающего коридора как маршрута миграции людей оценивается в 11 500 лет назад. [34]
Березовые леса распространились по бывшей разнотравной тундре в Берингии примерно на 17 000 л.н. в ответ на улучшение климата, что указывает на повышение продуктивности ландшафта. [40]
Анализ биомаркеров и микрофоссилий, сохранившихся в отложениях озер Е5 и Могильного озера на севере Аляски, позволяет предположить, что ранние люди сожгли берингийские ландшафты еще 34 000 лет назад. [45] [46] Авторы этих исследований предполагают, что огонь использовался как средство охоты на мегафауну.
Хронология, причины и источники миграции
[ редактировать ]Коренные народы Америки обнаружили археологическое присутствие на территории Америки примерно 15 000 лет назад. [47] [48] Однако более поздние исследования предполагают присутствие человека между 18 000 и 26 000 лет назад, во время последнего ледникового максимума. [49] [50] [7] Остается неопределенность относительно точной датировки отдельных участков и выводов, сделанных на основе исследований популяционной генетики современных коренных американцев.
Хронология
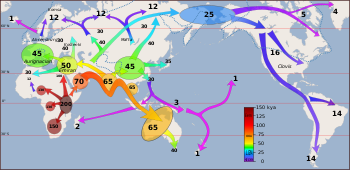
[ редактировать ]![«Карты, изображающие каждую фазу трехэтапной ранней миграции человека для заселения Америки. (A) Постепенное расширение населения предков индейцев из их генофонда в Центральной и Восточной Азии (синяя стрелка). (B) Заселение протоамериканцами Америки. Берингия с незначительным приростом населения или его отсутствием в течение ≈20 000 лет (C) Быстрая колонизация Нового Света группой основателей, мигрирующей на юг через свободный ото льда внутренний коридор между восточным Лаврентидским и западным Кордильерскими ледниковыми щитами (зеленая стрелка) и// или вдоль побережья Тихого океана (красная стрелка). На (B) обнаженное морское дно показано в наибольшей степени во время последнего ледникового максимума ≈20–18 000 лет назад [25], на (A) и (C). обнаженное морское дно изображено ≈40 000 лет назад и ≈16 000 лет назад, когда доисторический уровень моря был сопоставим. Уменьшенная версия современной Берингии (снижение A – C на 60%) представлена в левом нижнем углу. На карте меньшего размера показан Берингов пролив, который географически отделял Новый Свет от Азии примерно 11–10 000 лет назад».](http://upload.wikimedia.org/wikipedia/commons/thumb/d/d7/Journal.pone.0001596.g004.png/280px-Journal.pone.0001596.g004.png)
В начале XXI века модели хронологии миграции разделяются на два общих подхода. [52] [53]
Первая — это теория краткой хронологии , согласно которой первая миграция произошла после LGM, которая пришла в упадок примерно 19 000 лет назад. [37] а затем последовали последовательные волны иммигрантов. [54]
Вторая теория — это теория длинной хронологии, которая предполагает, что первая группа людей проникла в Берингию, включая свободные ото льда части Аляски, гораздо раньше, возможно, 40 000 лет назад. [55] [56] [57] за которым последовала гораздо более поздняя вторая волна иммигрантов. [53] [58]
Теория Хлодвига Первого , которая доминировала в антропологии Нового Света на протяжении большей части 20-го века, была оспорена в 2000-х годах в результате надежного датирования археологических памятников в Америке возрастом менее 13 000 лет назад. [32] [33] [34] [59] [48]
Все археологические памятники в Америке, самые древние даты которых получили широкое признание, соответствуют возрасту около 15 000 лет. Сюда входит комплекс Баттермилк-Крик в Техасе, [47] участок Медоукрофт Рокшелтер в Пенсильвании и участок Монте-Верде на юге Чили. [48] Археологические данные о существовании людей до Хлодвига указывают на то, что Топпер-Участку в Южной Каролине было 16 000 лет, в то время, когда ледниковый максимум теоретически допускал бы более низкие береговые линии.
Часто предполагалось, что свободный ото льда коридор на территории нынешней Западной Канады мог позволить миграцию до начала голоцена . Однако исследование 2016 года выступило против этого, предполагая, что заселение Северной Америки через такой коридор вряд ли произошло значительно раньше самых ранних стоянок Кловиса. В исследовании делается вывод, что свободный ото льда коридор на территории нынешней Альберты и Британской Колумбии «постепенно был захвачен бореальным лесом, в котором преобладают ели и сосновые деревья» и что «люди Кловиса, вероятно, пришли с юга, а не с севера, возможно, следуя за дикими животными, такими как бизоны ». [60] [61] Альтернативной гипотезой заселения Америки является прибрежная миграция , которая могла быть осуществима вдоль обезледеневшей (но теперь затопленной) береговой линии северо-запада Тихого океана примерно 16 000 лет назад.
Доказательства присутствия человека до LGM
[ редактировать ]
Миграция до LGM через Берингию была предложена для объяснения предполагаемого возраста археологических памятников в Америке, таких как пещеры Bluefish, до LGM. [56] и квартиры Олд Кроу [57] на территории Юкона и Meadowcroft Rock Shelter в Пенсильвании. [53] [58] Самые старые археологические памятники на аляскинской стороне Берингии датируются примерно 14 000 лет назад. [40] [62] Возможно, что до этого времени в Берингию проникла небольшая популяция основателей. Однако археологические памятники, датируемые ближе к LGM, ни на сибирской, ни на аляскинской стороне Берингии отсутствуют. Анализ биомаркеров и микрофоссилий отложений озер Е5 и озера Могила на севере Аляски позволяет предположить присутствие человека в восточной Берингии еще 34 000 лет назад. [45] Было высказано предположение, что эти осадочные анализы являются единственными возможными извлекаемыми остатками людей, живших на Аляске во время последнего ледникового периода. [46]
В Олд-Кроу-Флэтс были обнаружены кости мамонта, сломанные особым образом, что указывает на человеческую бойню. Радиоуглеродные даты на них варьируются от 25 000 до 40 000 лет назад. Также на участке обнаружены каменные микроотщепы, свидетельствующие о орудийном производстве. [63] Однако интерпретация следов мясника и геологическая ассоциация костей на стоянках Bluefish Cave и Old Crow Flats, а также на связанном с ними участке Bonnet Plume были поставлены под сомнение. [29] Никаких следов человеческих останков на этих участках обнаружено не было. Помимо спорных археологических памятников, подтверждение присутствия человека до LGM было обнаружено в записях озерных отложений на севере Аляски. Биомаркерный анализ и анализ микрофоссилий отложений из озер Е5 и Могильного озера позволяют предположить присутствие человека в восточной Берингии еще 34 000 лет назад. [45] [46] Эти анализы действительно убедительны, поскольку они подтверждают выводы, сделанные на основании местонахождений в пещере Блюфиш и равнинах Олд-Кроу.
В 2020 году появились доказательства существования нового участка до LGM в северо-центральной Мексике . Пещера Чикиуите , археологический памятник в штате Сакатекас , датируется 26 000 лет назад на основании многочисленных каменных артефактов, обнаруженных там. [64] Однако ученые спорят о том, следует ли считать эти артефакты свидетельством человеческой деятельности или они образовались естественным путем. [65] [28] Никаких доказательств присутствия человеческой ДНК или очага обнаружено не было. [66]
Присутствие человека в Южной Америке до LGM частично основано на хронологии скандального каменного убежища Педра Фурада в Пиауи , Бразилия . Совсем недавно исследования на археологических раскопках Санта-Элина (27-10 тыс. л.н.) [67] на Среднем Западе и Ринкан I (20 000–12 000 лет назад) [68] на юго-востоке Бразилии также обнаруживаются ассоциации свидетельств присутствия человека с отложениями, датируемыми до LGM. Исследование 2003 года датировало доказательства контролируемого использования огня более 40 000 лет назад. [69] Дополнительные доказательства были получены на основе морфологии окаменелости женщины Лузии , которая была описана как австрало-меланезийская . Эта интерпретация была оспорена в обзоре 2003 года, в котором был сделан вывод, что рассматриваемые особенности также могли возникнуть в результате генетического дрейфа. [70] В ноябре 2018 года ученые Университета Сан-Паулу и Гарвардского университета опубликовали исследование, которое противоречит предполагаемому австрало-меланезийскому происхождению Лузии. С помощью секвенирования ДНК результаты показали, что предки Лузии были полностью индейскими. [71] [72]
Камни, описанные как вероятные инструменты, молотки и наковальни , были найдены в южной Калифорнии , на стоянке Мастодонта Черутти , которые связаны со скелетом мастодонта , который, по-видимому, был обработан людьми. Скелет мастодонта был датирован радиометрическим анализом тория-230/урана с использованием моделей датирования диффузии-адсорбции-распада примерно 130 тысячами лет назад. [73] Никаких человеческих костей обнаружено не было, и реакция экспертов была неоднозначной; назвал «неправдоподобными» Заявления об инструментах и обработке костей профессор Том Диллехей . [74]
Местонахождение на реке Яна Рога носорога (RHS) датирует заселение человеком восточной части Арктической Сибири 31 300 лет назад. [75] Некоторые интерпретировали эту дату как свидетельство неизбежности миграции в Берингию, что подтверждает факт оккупации Берингии во время LGM. [76] [77] Однако дата Yana RHS относится к началу периода охлаждения, который привел к LGM. [31] Подборка дат археологических раскопок по всей Восточной Сибири позволяет предположить, что период похолодания вызвал отступление людей на юг. [40] [41] Каменные свидетельства до LGM в Сибири указывают на оседлый образ жизни, основанный на местных ресурсах, тогда как каменные свидетельства после LGM указывают на более мигрирующий образ жизни. [41]
Обнаружение в 2021 году человеческих следов в отложениях реликтового озера возле национального парка Уайт-Сэндс в Нью-Мексико позволяет предположить, что присутствие человека датировалось LGM между 18 000 и 26 000 лет назад. [49] [50] Более поздние исследования, о которых сообщалось в октябре 2023 года, подтвердили, что возраст человеческих следов составляет «до 23 000 лет». [78] [79]
Сторонники Хлодвига прежде всего не признали правдивость этих выводов. В 2022 году они заявили: «Самые старые свидетельства археологических раскопок в Новом Свете с большим количеством артефактов, встречающихся в дискретных и минимально нарушенных стратиграфических контекстах, встречаются в восточной Берингии между 13 000 и 14 200 лет назад. К югу от ледниковых щитов самые старые такие памятники Если людям удалось прорваться через континентальные ледниковые щиты значительно раньше 13 000 лет назад, то должны быть четкие доказательства этого в виде, по крайней мере, некоторых стратиграфически дискретных археологических компонентов с относительно высоким количеством артефактов. таких доказательств не существует». [80]
Оценка геномного возраста
[ редактировать ]
В генетических исследованиях использовались аналитические методы высокого разрешения, применяемые к образцам ДНК современных коренных американцев и азиатских популяций, считающихся их исходными популяциями, для реконструкции развития гаплогрупп ДНК Y-хромосомы человека ( гаплогруппы yDNA ) и гаплогрупп митохондриальной ДНК человека (гаплогруппы мтДНК), характерных для Индейское население. [55] [76] [77] Модели скорости молекулярной эволюции использовались для оценки возраста, в котором линии ДНК коренных американцев отделились от своих родительских линий в Азии, а также для определения возраста демографических событий. Одна модель (Tammetal 2007), основанная на гаплотипах мтДНК коренных американцев (рис. 2), предполагает, что миграция в Берингию произошла между 30 000 и 25 000 лет назад, а миграция в Америку произошла примерно через 10 000–15 000 лет после изоляции небольшой популяции-основателя . [76] Другая модель (Китчен и др., 2008) предполагает, что миграция в Берингию произошла примерно 36 000 лет назад, после чего последовала 20 000 лет изоляции в Берингии. [77] Третья модель (Nomatto et al. 2009) предполагает, что миграция в Берингию произошла между 40 000 и 30 000 лет назад, с миграцией до LGM в Америку, за которой последовала изоляция северной популяции после закрытия незамерзающего коридора. [55] Доказательства примеси австрало-меланезийцев в популяциях Амазонии были обнаружены Скоглундом и Райхом (2016). [81]
Исследование диверсификации гаплогрупп C и D мтДНК из южной Сибири и Восточной Азии соответственно предполагает, что родительская линия (субгаплогруппа D4h) субгаплогруппы D4h3, линии, обнаруженной среди коренных американцев и ханьских китайцев, [82] [83] возникло около 20 000 лет назад, ограничивая появление D4h3 пост-LGM. [84] По оценкам возраста, основанным на разнообразии микросателлитов Y-хромосомы, происхождение американской гаплогруппы Q1a3a (Y-ДНК) происходит примерно от 15 000 до 10 000 лет назад. [85] Большей согласованности моделей скорости молекулярной эволюции ДНК друг с другом и с археологическими данными можно добиться за счет использования датированной ископаемой ДНК для калибровки скорости молекулярной эволюции. [82]
Мегафаунальные миграции
[ редактировать ]Хотя нет никаких археологических свидетельств, которые можно было бы использовать для прямого подтверждения маршрута прибрежной миграции во время последнего ледникового максимума , генетический анализ был использован для подтверждения этого тезиса. В дополнение к генетическому происхождению человека, линия ДНК мегафауны может быть использована для отслеживания перемещений мегафауны – крупных млекопитающих – а также первых групп людей, которые охотились на них.
Зубры , тип мегафауны, были идентифицированы как идеальный кандидат для отслеживания миграций людей из Европы из-за их обилия в Северной Америке, а также из-за того, что они являются одной из первых мегафауны, для которой древняя ДНК использовалась для отслеживания закономерностей. движение населения. В отличие от других видов фауны, перемещавшихся между Америкой и Евразией ( мамонтов , лошадей и львов ), бизоны пережили вымирание в Северной Америке , произошедшее в конце плейстоцена. Однако в их геноме есть свидетельства узкого места, которое можно использовать для проверки гипотезы о миграции между двумя континентами. [90] Ранние человеческие группы были в основном кочевниками , полагаясь на следующие источники пищи для выживания. Мобильность была частью того, что сделало людей успешными. Будучи кочевыми группами, древние люди, вероятно, следовали за пищей из Евразии в Америку – это одна из причин, почему отслеживание ДНК мегафауны так полезно для понимания этих миграционных моделей. [91]
Серый волк зародился в Америке и мигрировал в Евразию до последнего ледникового максимума , во время которого считалось, что оставшиеся популяции серого волка, проживающие в Северной Америке, оказались под угрозой исчезновения и были изолированы от остальной популяции. Однако это может быть не так. Радиоуглеродное датирование останков древних серых волков, найденных в отложениях вечной мерзлоты на Аляске, показывает непрерывный обмен популяцией с 12 500 радиоуглеродных лет назад до пределов возможностей радиоуглеродного датирования. Это указывает на то, что существовала возможность обмена популяциями серых волков между двумя континентами. [92]
Способность этих фаун обмениваться популяциями в период последнего ледникового максимума, а также генетические данные, обнаруженные в останках ранних людей в Америке, служат доказательством поддержки миграций в Америку до Кловиса.
Исходные популяции
[ редактировать ]Исходная индейская популяция сформировалась в Сибири путем смешения двух различных популяций: древних жителей Северной Евразии и древней популяции Восточной Азии (ESEA). [93] [94] По словам Дженнифер Рафф, древнее население Северной Евразии смешалось с дочерним населением древних жителей Восточной Азии, с которым они столкнулись около 25 000 лет назад, что привело к появлению коренных американцев предкового населения . Однако точное место, где произошло смешение, неизвестно, а миграционные движения, объединившие две популяции, являются предметом споров. [95]
Одна теория предполагает, что древние жители Северной Евразии мигрировали на юг, в Восточную Азию или Южную Сибирь , где они встретились и смешались с древними жителями Восточной Азии. Генетические данные из озера Байкал в России подтверждают, что именно этот район имел место примеси. [96]
Однако третья теория, «гипотеза застоя Берингии», предполагает, что выходцы из Восточной Азии вместо этого мигрировали на север, в Северо-Восточную Сибирь, где они смешались с АНЕ, а затем разошлись в Берингии, где сформировались отдельные индейские линии. Эта теория подтверждается данными материнской и ядерной ДНК . [97] По словам Гребенюка, после 20 000 лет назад ветвь древних восточноазиатов мигрировала в Северо-Восточную Сибирь и смешалась с потомками АНЕ, что привело к появлению древних палеосибирских и индейских популяций в Крайнем Северо-Восточной Азии. [98]
Однако гипотеза застоя в Берингии не подтверждается данными отцовской ДНК, которые могут отражать различную популяционную историю отцовских и материнских линий у коренных американцев, что не является редкостью и наблюдалось в других популяциях. [99]
Исследование 2019 года показало, что коренные американцы являются ближайшими ныне живущими родственниками окаменелостей возрастом 10 000 лет, найденных у реки Колыма на северо-востоке Сибири. [100]
Исследование, опубликованное в июле 2022 года, показало, что люди на юге Китая, возможно, внесли свой вклад в генофонд коренных американцев, основываясь на открытии и анализе ДНК окаменелостей человека возрастом 14 000 лет. [101] [102]
Контраст между генетическими профилями скелетов Дзёмон с Хоккайдо и современных айнов иллюстрирует еще одну неопределенность в моделях источников, полученных на основе современных образцов ДНК. [103]
Митохондриальные (мтДНК) линии
[ редактировать ]Развитие геномного анализа высокого разрешения предоставило возможности для дальнейшего определения субкладов коренных американцев и сужения круга азиатских субкладов, которые могут быть родительскими или сестринскими субкладами.
Давно признано широкое распространение гаплогрупп A, B, C и D мтДНК среди населения Восточной Азии и коренных американцев, а также наличие X. гаплогруппы [104] В целом наибольшая частота четырех индейских гаплогрупп встречается в Алтае - Байкальском регионе Южной Сибири. [105] Некоторые субклады C и D, более близкие к индейским субкладам, встречаются среди монгольских, амурских, японских, корейских и айнских популяций. [104] [106]
С дальнейшим определением субкладов, связанных с индейским населением, требования к выборке азиатских популяций для поиска наиболее тесно связанных субкладов становятся более конкретными. Субгаплогруппы D1 и D4h3 считались специфическими для коренных американцев из-за их отсутствия среди большой выборки популяций, считающихся потенциальными потомками исходных популяций на обширной территории Азии. [76] Среди 3764 выборок Сахалино - Нижнеамурская область была представлена 61 ороком . [76] выявлена подгаплогруппа D1a (4 из 87 отобранных, или 4,6%), а также подгаплогруппа C1a (1 из 87, или 1,1%). В другом исследовании среди ульчей нижнего Приамурья [106] Субгаплогруппа C1a считается близкой сестринской кладой индейской подгаплогруппы C1b. [106]
Субгаплогруппа D1a также была обнаружена среди древних скелетов Дзёмон с Хоккайдо. [103] Современные айны считаются потомками Дзёмон. [103] Появление субгаплогрупп D1a и C1a в нижнем Приамурье предполагает наличие исходной популяции из этого региона, отличной от исходных популяций Алтая и Байкала, где выборка не выявила эти два конкретных субклада. [106] Сделаны выводы относительно подгаплогруппы D1, указывающие на потенциальные исходные популяции в нижнем Амуре. [106] и Хоккайдо [103] области контрастируют с моделью миграции с одним источником. [55] [76] [77]
выявлена подгаплогруппа D4h3 У ханьских китайцев . [82] [83] Субгаплогруппа D4h3 из Китая не имеет такого же географического значения, как субгаплотип D1a из Амур-Хоккайдо, поэтому его значение для исходных моделей более умозрительно. Считается, что его родительская линия, субгаплотип D4h, возникла в Восточной Азии, а не в Сибири, около 20 000 лет назад. [84] Субгаплогруппа D4h2, сестринская клада D4h3, также была обнаружена среди скелетов Дзёмон с Хоккайдо. [107] D4h3 имеет прибрежный след в Америке. [83]
X — одна из пяти гаплогрупп мтДНК, обнаруженных у коренных американцев. Коренные американцы в основном относятся к кладе X2a, которая никогда не встречалась в Старом Свете . [108] По мнению Дженнифер Рафф , X2a, вероятно, произошел от той же сибирской популяции, что и другие четыре материнские линии-основатели, и нет убедительных оснований полагать, что он связан с линиями X, обнаруженными в Европе или Западной Евразии. Было обнаружено, что ископаемое человека из Кенневика несет самую глубокую ветвь гаплогруппы X2a, и у него не было никакого европейского происхождения, которое можно было бы ожидать от европейского происхождения этой линии. [109]
Геномика HTLV-1
[ редактировать ]Лимфотрофный вирус 1 Т-клеток человека ( HTLV-1 ) — это вирус, передающийся при обмене жидкостями организма и от матери к ребенку через грудное молоко. Передача от матери ребенку имитирует наследственный признак, хотя такая передача от матери-носителя составляет менее 100%. [110] Геном вируса HTLV был картирован, что позволило идентифицировать четыре основных штамма и проанализировать их древность посредством мутаций. Самые высокие географические концентрации штамма HLTV-1 наблюдаются в странах Африки к югу от Сахары и Японии. [111] В Японии наибольшая концентрация встречается на острове Кюсю . [111] Он также присутствует среди потомков африканцев и коренного населения Карибского региона и Южной Америки. [111] В Центральной Америке и Северной Америке это редкость. [111] Считается, что его распространение в Америке произошло в результате завоза в рамках работорговли. [112]
У айнов выработались антитела к HTLV-1, что указывает на его эндемичность для айнов и его древность в Японии. [113] Подтип «А» был определен и идентифицирован среди японцев (включая айнов ), а также среди изолятов из Карибского бассейна и Южной Америки. [114] Подтип «В» выявлен в Японии и Индии. [114] В 1995 году у коренных американцев прибрежной Британской Колумбии были обнаружены оба подтипа А и В. [115] Сообщалось, что образцы костного мозга андской мумии возрастом около 1500 лет показали наличие подтипа А. [116] Это открытие вызвало споры: утверждалось, что образец ДНК был недостаточно полным для такого вывода и что результат отражает современное загрязнение. [117] Однако повторный анализ показал, что последовательности ДНК соответствовали, но не определенно принадлежали «космополитической кладе» (подтип А). [117] Присутствие подтипов A и B в Америке наводит на мысль о том, что исходная популяция коренных американцев связана с предками айнов, Дзёмон .
Физическая антропология
[ редактировать ]Палеоиндийские скелеты в Америке, такие как череп человека из Кенневика (штат Вашингтон), скелет негра Хойя (Юкатан), женщины Лузии и другие черепа из стоянки Лагоа-Санта (Бразилия), женщины из Буля (Айдахо), женщины из Пеньона III , [118] два черепа из стоянки Тлапакойя (Мехико), [118] и 33 черепа из Нижней Калифорнии. [119] продемонстрировали определенные черепно-лицевые черты, отличные от большинства современных коренных американцев, что побудило физических антропологов предположить более раннюю «палеоамериканскую» популяционную волну. [120] Самым основным измеренным отличительным признаком является долихоцефалия черепа. Некоторые современные изолированные популяции, такие как перикуэсы Нижней Калифорнии и огнеземельцы Огненной Земли, обладают той же морфологической чертой. [119]
Другие антропологи выступают за альтернативную гипотезу о том, что эволюция исходного берингийского фенотипа привела к возникновению особой морфологии, которая была сходной во всех известных палеоамериканских черепах, с последующей конвергенцией к современному фенотипу коренных американцев. [121] [122]
Археогенетические исследования не подтверждают двухволновую модель или палеоамериканскую гипотезу австрало-меланезийского происхождения и твердо относят всех палеоиндейцев и современных коренных американцев к одной древней популяции, которая проникла в Америку в результате единой миграции из Берингии. Только у одного древнего экземпляра (Лагоа Санта) и нескольких современных популяций в регионе Амазонки небольшой компонент австралийского происхождения ок. Обнаружено 3%, что остается необъясненным текущим состоянием исследований (по состоянию на 2021 г. [update]), но может быть объяснено наличием более базовой родословной, связанной с Тяньюанем , глубокой восточноазиатской линией, которая не внесла прямого вклада в современных аборигенов Восточной Азии, но, возможно, внесла свой вклад в предков коренных американцев в Сибири, поскольку такое происхождение также найден среди предыдущих палеолитических сибиряков ( древних жителей Северной Евразии ). [81] [123] [93]
В отчете, опубликованном в Американском журнале физической антропологии в январе 2015 года, были рассмотрены черепно-лицевые вариации с упором на различия между ранними и поздними коренными американцами и их объяснения, основанные либо на морфологии черепа, либо на молекулярной генетике. Аргументы, основанные на молекулярной генетике, в основном, по мнению авторов, допускают единственную миграцию из Азии с вероятной паузой в Берингии, а также более поздний двунаправленный поток генов. В некоторых исследованиях, посвященных черепно-лицевой морфологии, ранее утверждалось, что палеоамериканские останки были описаны как более близкие к австрало-меланезийцам и полинезийцам, чем к современной группе коренных американцев, что позволяет предположить два проникновения в Америку, раннее из которых произошло до того, как возникла характерная восточноазиатская морфология. (именуемая в статье «Модель двух компонентов»). Другая «третья модель», модель «Повторяющегося потока генов» (RGF), пытается примирить две, утверждая, что циркуарктический поток генов после первоначальной миграции может объяснять морфологические изменения. В нем специально переоценивается первоначальный отчет о скелете негра хойя, который поддерживал модель RGF. Авторы не согласились с первоначальным выводом, в котором предполагалось, что форма черепа не соответствует форме черепа современных коренных американцев, утверждая, что «череп попадает в субрегион морфопространства, занимаемого как палеоамериканцами, так и некоторыми современными коренными американцами». [124]
Стволовые точки
[ редактировать ]Острие на ножке — это каменная технология, отличная от берингийского и кловисского типов. Их распространение варьируется от прибрежных районов Восточной Азии до тихоокеанского побережья Южной Америки. [43] Появление наконечников на ножках было прослежено в Корее в эпоху верхнего палеолита. [125] Происхождение и распространение стволовых точек были интерпретированы как культурный маркер, связанный с исходной популяцией из прибрежной Восточной Азии. [43]
Миграционные маршруты
[ редактировать ]Внутренний маршрут
[ редактировать ]
Теория Хлодвига-Первого
[ редактировать ]Исторически теории о миграции в Америку вращались вокруг миграции из Берингии через внутренние районы Северной Америки. Обнаружение артефактов, связанных с останками плейстоценовой фауны недалеко от Кловиса, штат Нью-Мексико, в начале 1930-х годов потребовало продления временных рамок заселения Северной Америки до периода, в течение которого ледники все еще были обширными. Это привело к появлению гипотезы о миграционном маршруте между ледниковыми щитами Лаврентиды и Кордильеров, объясняющей раннее заселение. На стоянке Кловис использовалась каменная технология, характеризующаяся наконечниками копий с углублением или канавкой, где острие прикреплялось к древку. Каменный комплекс, характеризующийся технологией Кловис-Пойнт, впоследствии был обнаружен на большей части территории Северной Америки и в Южной Америке. Связь сложной технологии Кловиса с остатками фауны позднего плейстоцена привела к появлению теории о том, что она ознаменовала прибытие охотников на крупную дичь, которые мигрировали из Берингии, а затем рассеялись по всей Америке, иначе известной как теория Кловиса Первого.
Недавнее радиоуглеродное датирование стоянок Кловиса позволило установить возраст от 13 000 до 12 600 лет назад, что несколько позже, чем даты, полученные с помощью более старых методов. [126] Переоценка более ранних радиоуглеродных дат привела к выводу, что не менее 11 из 22 стоянок Кловиса с радиоуглеродными датами являются «проблемными» и их следует игнорировать, включая типовое место в Кловисе, штат Нью-Мексико. Численное датирование памятников Хлодвига позволило сравнить даты Хлодвига с датами других археологических памятников по всей Америке, а также с датами открытия свободного ото льда коридора. Оба приводят к серьезным проблемам с теорией Хлодвига Первого. Местонахождение Монте-Верде на юге Чили датируется 14 800 лет назад. [48] Участок в пещере Пейсли в восточном Орегоне дал 14 500 лет назад на копролите с человеческой ДНК и радиоуглеродными датами 13 200 и 12 900 лет назад на горизонтах, содержащих западные стволовые точки. [127] Горизонты артефактов с каменными комплексами, не принадлежащими Кловису, и дохловисским возрастом встречаются в восточной части Северной Америки, хотя максимальный возраст, как правило, плохо ограничен. [59] [128]
Недавние исследования показали, что свободный ото льда коридор открылся позже (около 13 800 ± 500 лет назад), чем самые ранние широко признанные археологические памятники в Америке, что позволяет предположить, что он не мог использоваться в качестве миграционного маршрута для миграции первых народов. юг. [129]
Каменные свидетельства миграций до Хлодвига
[ редактировать ]
Геологические находки о времени появления свободного ото льда коридора также бросают вызов представлению о том, что заселение Северной и Южной Америки людьми Хлодвига и до него было результатом миграции по этому маршруту после последнего ледникового максимума. Закрытие коридора до LGM может приблизиться к 30 000 лет назад, а оценки отступления льда из коридора находятся в диапазоне от 13 000 до 12 000 лет назад. [32] [33] [34] Жизнеспособность коридора как пути миграции людей оценивается в 11 500 лет назад, что позже, чем возраст Кловиса и дохлодвигских стоянок. [34] Датированные археологические памятники Кловиса предполагают распространение культуры Кловис с юга на север. [32]
Миграция до-LGM во внутренние районы была предложена для объяснения дохловисского возраста археологических памятников в Америке. [53] [58] хотя такие места, существовавшие до Кловиса, как Meadowcroft Rock Shelter, [59] [128] Монте Верде, [48] и Пещера Пейсли не дали подтвержденного возраста до LGM.
, есть много стоянок до Хлодвига На юго-западе Америки, особенно в пустыне Мохаве . В карьерах озера Мохаве, относящихся к плейстоцену, хранятся каменные остатки снарядов Силвер-Лейк и снарядов озера Мохаве. Это указывает на внутреннее движение в регион уже в 13 800 лет назад, если не раньше. [130]
Предложение дене-енисейской языковой семьи
[ редактировать ]Связь между языками на-дене Северной Америки (такими как навахо и апачи) и енисейскими языками Сибири была впервые предложена еще в 1923 году и получила дальнейшее развитие другими. Подробное исследование было проведено Эдвардом Вайдой и опубликовано в 2010 году. [131] Эта теория получила поддержку многих лингвистов, а археологические и генетические исследования предоставили ей дополнительную поддержку. [ нужна ссылка ]
Традиция изготовления арктических малых инструментов на Аляске и в канадской Арктике, возможно, зародилась в Восточной Сибири около 5000 лет назад. Это связано с древними палеоэскимосскими народами Арктики.
Источником традиции арктического стрелкового инструмента могла быть культурная последовательность Сялах -Белькачи- Ымяхтах Восточной Сибири, датируемая 6500–2800 л.н. [132]
Внутренний маршрут соответствует распространению языковой группы на-дене. [131] и субгаплогруппа X2a в Америку после самой ранней палеоамериканской миграции. [83]
Тем не менее, некоторые ученые предполагают, что предки жителей западной Северной Америки, говорящие на языках на-дене, совершили прибрежную миграцию на лодке. [133]
Тихоокеанский прибрежный маршрут
[ редактировать ]Теория тихоокеанской прибрежной миграции предполагает, что люди впервые достигли Америки водным путем, следуя по береговым линиям из северо-восточной Азии в Америку, первоначально предложенную в 1979 году Кнутом Флэдмарком в качестве альтернативы гипотетической миграции через незамерзающий внутренний коридор. [134] Эта модель поможет объяснить быстрое распространение вируса на прибрежные территории, чрезвычайно удаленные от региона Берингова пролива, включая такие места, как Монте-Верде на юге Чили и Тайма-Тайма на западе Венесуэлы .
Очень похожая гипотеза морской миграции представляет собой вариант прибрежной миграции; по сути, его единственное отличие состоит в том, что он постулирует, что лодки были основным средством передвижения. Предлагаемое использование лодок добавляет определенную гибкость хронологии прибрежной миграции, поскольку тогда не потребуется непрерывное свободное ото льда побережье (16–15 000 калиброванных лет назад): Мигранты в лодках могли легко обойти ледяные преграды и поселиться в разрозненных прибрежных рефугиумах до завершения дегляциации прибрежного сухопутного маршрута. Популяция, компетентная в области морских перевозок, в прибрежных районах Восточной Азии является важной частью гипотезы морской миграции. [43] [44]
В статье 2007 года в « Журнале островной и прибрежной археологии» была предложена «гипотеза шоссе из водорослей», вариант прибрежной миграции, основанный на эксплуатации лесов из водорослей вдоль большей части Тихоокеанского региона от Японии до Берингии, северо-запада Тихого океана и Калифорнии, а также до Андского побережья Южной Америки. После того как береговые линии Аляски и Британской Колумбии исчезли около 16 000 лет назад, эти леса водорослей (наряду с устьевыми, мангровыми зарослями и коралловыми рифами) стали экологически однородным миграционным коридором, полностью находящимся на уровне моря и практически беспрепятственным.Анализ ДНК растений и животных, проведенный в 2016 году, показал, что прибрежный маршрут возможен. [135] [136]
Митохондриальная субгаплогруппа D4h3a, редкий субклад D4h3 , встречающийся вдоль западного побережья Америки, была идентифицирована как клада, связанная с прибрежной миграцией. [83] Эта гаплогруппа была обнаружена в скелете, известном как Анзик-1 , найденном в Монтане в тесной связи с несколькими артефактами Кловиса, датированными 12 500 лет назад. [137]
Проблемы с оценкой моделей прибрежной миграции
[ редактировать ]Модели прибрежной миграции предлагают другой взгляд на миграцию в Новый Свет, но они не лишены своих проблем: одна из таких проблем заключается в том, что глобальный уровень моря поднялся более чем на 120 метров (390 футов). [138] с момента окончания последнего ледникового периода, и это привело к затоплению древних береговых линий, по которым морские люди могли бы следовать в Америку. Найти места, связанные с ранними прибрежными миграциями, чрезвычайно сложно, а систематические раскопки любых мест, обнаруженных в более глубоких водах, являются сложной и дорогостоящей задачей. Стратегии поиска мест ранней миграции включают выявление потенциальных участков на затопленных палеобереговых линиях , поиск участков в районах, поднятых либо в результате тектоники, либо изостатического отскока, а также поиск речных участков в районах, которые могли привлечь прибрежных мигрантов. [43] [139] С другой стороны, есть свидетельства морских технологий, обнаруженных на холмах Нормандских островов в Калифорнии около 12 000 лет назад. [140] Если была ранняя прибрежная миграция до Хлодвига, всегда существует вероятность «неудавшейся колонизации».
См. также
[ редактировать ]- Ранние миграции человека
- Генетическая история коренных народов Америки
- Список первых населенных пунктов
- История населения коренных народов Америки
- Теории доколумбовых трансокеанских контактов
Ссылки
[ редактировать ]- ^ Буренхульт, Горан (2000). Первые люди . Издательство «Вельтбилд». ISBN 978-3-8289-0741-6 .
- ^ Прингл, Хизер (8 марта 2017 г.). «Что происходит, когда археолог бросает вызов общепринятому научному мышлению?» . Смитсоновский институт .
- ^ Фэган, Брайан М. и Дуррани, Надя (2016). Предыстория мира: краткое введение . Рутледж. п. 124. ИСБН 978-1-317-34244-1 .
- ^ Jump up to: а б Гебель, Тед; Уотерс, Майкл Р.; О'Рурк, Деннис Х. (2008). «Расселение современных людей в Америке в позднем плейстоцене» (PDF) . Наука . 319 (5869): 1497–1502. Бибкод : 2008Sci...319.1497G . CiteSeerX 10.1.1.398.9315 . дои : 10.1126/science.1153569 . ПМИД 18339930 . S2CID 36149744 . Архивировано из оригинала (PDF) 2 января 2014 г. Проверено 5 февраля 2010 г.
- ^ Циммер, Карл (3 января 2018 г.). «В костях похороненного ребенка — признаки массовой миграции людей в Америку» . Нью-Йорк Таймс . Проверено 3 января 2018 г.
- ^ Морено-Майар, СП; Поттер, бакалавр; Виннер, Л; и др. (2018). «Геном Аляски терминального плейстоцена показывает первую популяцию коренных американцев» (PDF) . Природа . 553 (7687): 203–207. Бибкод : 2018Natur.553..203M . дои : 10.1038/nature25173 . ПМИД 29323294 . S2CID 4454580 .
- ^ Jump up to: а б Нуньес Кастильо, Мелида Инес (20 декабря 2021 г.). Древний генетический ландшафт археологических человеческих останков из Панамы, Южной Америки и Океании, описанный с помощью частот генотипов STR и последовательностей митохондриальной ДНК . Диссертация (докторская диссертация). doi : 10.53846/goediss-9012 . S2CID 247052631 .
- ^ Эш, Патрисия Дж. и Робинсон, Дэвид Дж. (2011). Появление людей: исследование эволюционной временной шкалы . Джон Уайли и сыновья . п. 289. ИСБН 978-1-119-96424-7 .
- ^ Робертс, Алиса (2010). Невероятное человеческое путешествие . А&С Черный. стр. 101–103. ISBN 978-1-4088-1091-0 .
- ^ Фитцхью, доктора. Уильям; Годдард, Айвз; Оусли, Стив; Оусли, Дуг; Стэнфорд, Деннис. «Палеоамериканский» . Отделение антропологии Смитсоновского института. Архивировано из оригинала 5 января 2009 г. Проверено 15 января 2009 г.
- ^ «Атлас человеческого пути-Генографический проект» . Национальное географическое общество. 1996–2008 гг. Архивировано из оригинала 1 мая 2011 г. Проверено 27 января 2017 г.
- ^ «Население Америки: генетическое происхождение влияет на здоровье» . Научный американец . Проверено 8 мая 2019 г.
- ^ Фладмарк, КР (1979). «Альтернативные миграционные коридоры для древних людей в Северной Америке». Американская древность . 44 (1): 55–69. дои : 10.2307/279189 . JSTOR 279189 . S2CID 162243347 .
- ^ «68 ответов на «Море поднимется до уровня последнего ледникового периода»» . Центр исследования климатических систем Колумбийского университета . 26 января 2009 года . Проверено 17 ноября 2009 г.
- ^ Jump up to: а б Ноль (27 июня 2022 г.). «Население Америки» . Труды Национальной академии наук Соединенных Штатов Америки . Проверено 19 декабря 2022 г.
- ^ Вагеспак, Николь (2012). «Ранние палеоиндейцы: от колонизации до Фолсома» . В Тимоти Р. Паукетат (ред.). Оксфордский справочник по североамериканской археологии . Издательство Оксфордского университета. стр. 86–95. ISBN 978-0-19-538011-8 .
- ^ Суровелл, штат Калифорния; Аллаун, ЮАР; Джинджерич, ДЖЕМ; Граф, К.Э.; Холмс, компакт-диск (2022). «Поздняя дата прибытия человека в Северную Америку» . ПЛОС ОДИН . 17 (4): e0264092. дои : 10.1371/journal.pone.0264092 . ПМК 9020715 . ПМИД 35442993 .
- ^ Jump up to: а б с Ясински, Эмма (2 мая 2022 г.). «Новые данные усложняют историю заселения Америки» . Журнал Ученый . Проверено 19 декабря 2022 г.
- ^ Арделин, Чиприан Ф.; Бесерра-Вальдивия, Лорена; Педерсен, Миккель Винтер; Швеннингер, Жан-Люк; Овиатт, Чарльз Г.; Масиас-Кинтеро, Хуан И.; Арройо-Кабралес, Хоакин; Сикора, Мартин; Окампо-Диас, Ям Зул Э.; Рубио-Сиснерос, Игорь И.; Уотлинг, Дженнифер Г.; Де Медейрос, Ванда Б.; Де Оливейра, Пауло Э.; Барба-Пингарон, Луис; Ортис-Бутрон, Агустин; Бланкас-Васкес, Хорхе; Ривера-Гонсалес, Иран; Солис-Росалес, Корина; Родригес-Сеха, Мария; Ганди, Девлин А.; Наварро-Гутьеррес, Замара; де ла Роса-Диас, Хесус Х.; Уэрта-Арельяно, Владимир; Маррокин-Фернандес, Марко Б.; Мартинес-Риохас, Л. Мартин; Лопес-Хименес, Алехандро; Хайэм, Томас; Виллерслев, Эске (2020). «Свидетельства человеческой оккупации Мексики во время последнего ледникового максимума» . Природа . 584 (7819): 87–92. Бибкод : 2020Natur.584...87A . дои : 10.1038/s41586-020-2509-0 . ПМИД 32699412 . S2CID 220697089 .
- ^ Бесерра-Вальдивия, Лорена; Хайэм, Томас (2020). «Время и последствия первых посещений людей в Северной Америке». Природа . 584 (7819): 93–97. Бибкод : 2020Natur.584...93B . дои : 10.1038/s41586-020-2491-6 . ПМИД 32699413 . S2CID 220715918 .
- ^ Грюн, Рут (22 июля 2020 г.). «Появляется все больше свидетельств того, что заселение Америки началось более 20 000 лет назад». Природа . 584 (7819): 47–48. Бибкод : 2020Natur.584...47G . дои : 10.1038/d41586-020-02137-3 . ПМИД 32699366 . S2CID 220717778 .
- ^ Спенсер Уэллс (2006). Глубокое происхождение: внутри генографического проекта . Национальные географические книги. стр. 222–. ISBN 978-0-7922-6215-2 . OCLC 1031966951 .
- ^ Джон Х. Релетфорд (17 января 2017 г.). 50 великих мифов об эволюции человека: понимание заблуждений о нашем происхождении . Джон Уайли и сыновья. стр. 192–. ISBN 978-0-470-67391-1 . OCLC 1238190784 .
- ^ Х. Джеймс Биркс, изд. (10 июня 2010 г.). Антропология XXI века: Справочник . Публикации SAGE. ISBN 978-1-4522-6630-5 . OCLC 1102541304 .
- ^ Джон Э. Кица; Ребекка Хорн (3 ноября 2016 г.). Устойчивые культуры: коренные народы Америки противостоят европейской колонизации 1500–1800 гг. (2-е изд.). Рутледж. ISBN 978-1-315-50987-7 .
- ^ Байсас, Лаура (16 ноября 2022 г.). «Ученые все еще пытаются выяснить, как состарить древние следы в национальном парке Уайт-Сэндс» . Популярная наука . Проверено 18 сентября 2023 г.
- ^ Сомервилл, Эндрю Д.; Касар, Изабель; Арройо-Кабралес, Хоакин (2021). «Новый радиоуглеродный возраст AMS на докерамических уровнях пещеры Кошкатлан, Пуэбла, Мексика: плейстоценовое заселение долины Теуакан?» . Латиноамериканская древность . 32 (3): 612–626. дои : 10.1017/laq.2021.26 .
- ^ Jump up to: а б Чаттерс, Джеймс К.; Поттер, Бен А.; Прентисс, Анна Мари; Фидель, Стюарт Дж.; Хейнс, Гэри; Келли, Роберт Л.; Килби, Дж. Дэвид; Ланоэ, Франсуа; Холланд-Лулевич, Джейкоб; Миллер, Д. Шейн; Морроу, Джульетта Э.; Перри, Анджела Р.; Радемейкер, Курт М.; Ройтер, Джошуа Д.; Ритчисон, Брэндон Т.; Санчес, Гваделупе; Санчес-Моралес, Исмаэль; Спайви-Фолкнер, С. Маргарет; Мелодия, Джесси В.; Хейнс, К. Вэнс (23 октября 2021 г.). «Оценка утверждений о заселении ранними людьми пещеры Чикиуите, Мексика». ПалеоАмерика . 8 (1). Информа UK Limited: 1–16. дои : 10.1080/20555563.2021.1940441 . ISSN 2055-5563 . S2CID 239853925 .
- ^ Jump up to: а б Брайант, Вон М. младший (1998). «До Кловиса» . У Гая Гиббона; и др. (ред.). Археология доисторической Америки: энциклопедия . Справочная библиотека «Гирлянда» по гуманитарным наукам. Том. 1537. стр. 682–683. ISBN 978-0-8153-0725-9 .
- ^ Хант, Кэти (05 октября 2023 г.). «Ученые говорят, что они подтвердили доказательства того, что люди прибыли в Америку гораздо раньше, чем считалось ранее» . CNN . Проверено 22 июля 2024 г.
- ^ Jump up to: а б с д и ж г час Бригам-Гретт, Джули; Ложкин Анатолий Владимирович; Андерсон, Патрисия М. и Глушкова, Ольга Ю. (2004). «Палеоэкологические условия Западной Берингии перед последним ледниковым максимумом» . В Д.Б. Мэдсене (ред.). Вхождение в Америку: Северо-Восточная Азия и Берингия перед последним ледниковым максимумом . Университет Юты Пресс. ISBN 978-0-87480-786-8 .
- ^ Jump up to: а б с д и ж г Джексон, Лайонел Э. младший и Уилсон, Майкл К. (февраль 2004 г.). «Возвращение к незамерзающему коридору» . Геотаймс . Американский геологический институт .
- ^ Jump up to: а б с д и Джексон, Л.Е. младший; Филлипс, FM; Шимамура, К. и Литтл, ЕС (1997). «Космогенный 36 Cl датировка поезда Foothills Erratics, Альберта, Канада». Geology . 25 (3): 195–198. Bibcode : 1997Geo....25..195J . doi : 10.1130/0091-7613(1997)025<0195: ccdotf>2.3.co;2 .
- ^ Jump up to: а б с д и ж г час я дж к Мандрик, Кэрол А.С.; Йозенханс, Хайнер; Федже, Дэрил В. и Мэтьюз, Рольф В. (январь 2001 г.). «Позднечетвертичная палеосреда северо-запада Северной Америки: последствия для внутренних и прибрежных миграционных маршрутов» . Четвертичные научные обзоры . 20 (1): 301–314. Бибкод : 2001QSRv...20..301M . дои : 10.1016/s0277-3791(00)00115-3 .
- ^ Jump up to: а б Дайк, А.С.; Мур А. и Робертсон Л. (2003). Дегляциация Северной Америки (Отчет). Открыть файл 1574. Геологическая служба Канады . дои : 10.4095/214399 .
- ^ Jump up to: а б Бут, Дерек Б.; Трост, Кэти Гетц; Клэг, Джон Дж. и Уэйт, Ричард Б. (2003). «Кордильерский ледниковый щит». Четвертичный период в США . Развитие четвертичных наук. Том. 1. стр. 17–43. дои : 10.1016/S1571-0866(03)01002-9 . ISBN 978-0-4445-1470-7 .
- ^ Jump up to: а б Блез, Б.; Клэг, Джей-Джей и Мэтьюз, RW (1990). «Время максимального оледенения позднего Висконсина, западное побережье Канады». Четвертичные исследования . 34 (3): 282–295. Бибкод : 1990QuRes..34..282B . дои : 10.1016/0033-5894(90)90041-i . S2CID 129658495 .
- ^ Мисарти, Николь; Финни, Брюс П.; Джордан, Джеймс В.; и др. (10 августа 2012 г.). «Раннее отступление ледникового комплекса полуострова Аляска и последствия для прибрежных миграций первых американцев». Четвертичные научные обзоры . 48 : 1–6. Бибкод : 2012QSRv...48....1M . doi : 10.1016/j.quascirev.2012.05.014 .
- ^ Jump up to: а б с д и Клэг, Джон Дж.; Мэтьюз, Рольф В. и Агер, Томас А. (2004). «Окружающая среда северо-запада Северной Америки до последнего ледникового максимума» . В Д.Б. Мэдсене (ред.). Вхождение в Америку: Северо-Восточная Азия и Берингия перед последним ледниковым максимумом . Университет Юты Пресс. ISBN 978-0-87480-786-8 .
- ^ Jump up to: а б с д и ж Васильев Сергей А.; Кузьмин Ярослав Васильевич; Орлова, Любовь А. и Дементьев, Вячеслав Н. (2002). «Радиоуглеродная хронология палеолита Сибири и ее значение для заселения Нового Света» . Радиоуглерод . 44 (2): 503–530. Бибкод : 2002Radcb..44..503V . дои : 10.1017/s0033822200031878 .
- ^ Jump up to: а б с Граф, Келли Э. (2009). «Современная человеческая колонизация Сибирской мамонтовой степи: взгляд из юга Центральной Сибири» (PDF) . В лагерях Марта; Парт Чаухан (ред.). Справочник по палеолитическим переходам . Спрингер. стр. 479–501. дои : 10.1007/978-0-387-76487-0_32 . ISBN 978-0-387-76478-8 .
- ^ Jump up to: а б с д и Федже, Дэрил В.; Маки, Квентин; Диксон, Э. Джеймс и Хитон, Тимоти Х. (2004). «Окружающая среда позднего Висконсина и археологическая видимость вдоль северного северо-западного побережья» . В Д.Б. Мэдсене (ред.). Вхождение в Америку: Северо-Восточная Азия и Берингия перед последним ледниковым максимумом . Университет Юты Пресс. ISBN 978-0-87480-786-8 .
- ^ Jump up to: а б с д и Эрландсон, Джон М. и Брайе, Тодд Дж. (2011). «Из Азии в Америку на лодке? Палеогеография, палеоэкология и стволовые точки северо-западной части Тихого океана». Четвертичный интернационал . 239 (1–2): 28–37. Бибкод : 2011QuInt.239...28E . дои : 10.1016/j.quaint.2011.02.030 .
- ^ Jump up to: а б с Эрландсон, Джон М.; Грэм, Майкл Х.; Бурк, Брюс Дж.; и др. (2007). «Гипотеза шоссе Келп: морская экология, теория прибрежной миграции и заселение Америки». Журнал островной и прибрежной археологии . 2 (2): 161–174. дои : 10.1080/15564890701628612 . S2CID 140188874 .
- ^ Jump up to: а б с Вачула, РС; Хуанг, Ю.; Рассел, Дж. М.; и др. (20 мая 2020 г.). «Осадочные биомаркеры подтверждают воздействие человека на экосистемы северной Берингии в последний ледниковый период» . Борей . 49 (3): 514–525. Бибкод : 2020Борея..49..514В . дои : 10.1111/bor.12449 .
- ^ Jump up to: а б с Вачула, РС; Хуанг, Ю.; Лонго, ВМ; и др. (13 декабря 2018 г.). «Свидетельства существования людей ледникового периода в восточной Берингии позволяют предположить раннюю миграцию в Северную Америку» . Четвертичные научные обзоры . 205 : 35–44. doi : 10.1016/j.quascirev.2018.12.003 . S2CID 134519782 .
- ^ Jump up to: а б Каплан, Сара (24 октября 2018 г.). «Старейшие наконечники копий континента дают новые сведения о первых американцах» . Вашингтон Пост .
- ^ Jump up to: а б с д и Диллехей, Томас (2000). Заселение Америки: новая предыстория . Нью-Йорк: Основные книги. ISBN 978-0-465-07669-7 .
- ^ Jump up to: а б Циммер, Карл (23 сентября 2021 г.). «Древние следы отодвигают дату прибытия человека в Америку» . Нью-Йорк Таймс . Проверено 23 сентября 2021 г.
- ^ Jump up to: а б Мэтью Беннетт; и др. (23 сентября 2021 г.). «Свидетельства существования людей в Северной Америке во время последнего ледникового максимума» . Наука . 373 (6562): 1528–1531. Бибкод : 2021Sci...373.1528B . дои : 10.1126/science.abg7586 . ПМИД 34554787 . S2CID 237616125 . Проверено 24 сентября 2021 г.
- ^ Рисунок 4 Андрей, Кухня (2008). «Трехэтапная модель колонизации для заселения Америки» . ПЛОС ОДИН . 3 (2): e1596. Бибкод : 2008PLoSO...3.1596K . дои : 10.1371/journal.pone.0001596 . ПМК 2223069 . ПМИД 18270583 .
- ^ Уайт, Филипп М. (2006). Хронология американских индейцев: хронологии американской мозаики . Гринвуд. п. 1. ISBN 978-0-313-33820-5 .
- ^ Jump up to: а б с д Уэллс, Спенсер и Рид, Марк (2002). Путешествие человека – генетическая одиссея . Случайный дом. стр. 138–140. ISBN 978-0-8129-7146-0 .
- ^ Ловгрен, Стефан (13 марта 2008 г.). «Америка заселена 15 000 лет назад, говорится в исследовании» . Нэшнл Географик . Архивировано из оригинала 14 марта 2008 года.
- ^ Jump up to: а б с д Бонатто, Сандро Л. и Сальзано, Франциско М. (1997). «Единичная и ранняя миграция населения Америки, подтвержденная данными о последовательностях митохондриальной ДНК» . Труды Национальной академии наук . 94 (5): 1866–1871. Бибкод : 1997PNAS...94.1866B . дои : 10.1073/pnas.94.5.1866 . ПМК 20009 . ПМИД 9050871 .
- ^ Jump up to: а б Синк-Марс, Дж. (1979). «Пещера луфиша 1: отложения восточно-берингийской пещеры позднего плейстоцена в Северном Юконе». Канадский журнал археологии (3): 1–32. JSTOR 41102194 .
- ^ Jump up to: а б Боннихсен, Робсон (1978). «Критические аргументы в пользу плейстоценовых артефактов из бассейна Олд-Кроу, Юкон: предварительное заявление» . У Алана Л. Брайана (ред.). Ранний человек в Америке с точки зрения Тихоокеанского региона . Периодические статьи № 1. Эдмонтон: Международный отдел антропологии археологических исследований Университета Альберты . стр. 102–118. ISBN 978-0-88864-999-7 .
- ^ Jump up to: а б с Оппенгеймер, Стивен. «Путешествие человечества» . Фонд Брэдшоу .
- ^ Jump up to: а б с Гудиер, Альберт К. (2005). «Свидетельства о стоянках до Хлодвига на востоке Соединенных Штатов» . В Робсоне Боннихсене; и др. (ред.). Палеоамериканское происхождение: за пределами Хлодвига . Заселение Америки. Центр изучения первых американцев Техасского университета A&M. стр. 103–112. ISBN 978-1-60344-812-3 .
- ^ Педерсен, Миккель В.; Рутер, Энтони; Швегер, Чарльз; и др. (10 августа 2016 г.). «Постледниковая жизнеспособность и колонизация в свободном ото льда коридоре Северной Америки» . Природа . 537 (7618): 45–49. Бибкод : 2016Natur.537...45P . дои : 10.1038/nature19085 . ПМИД 27509852 . S2CID 4450936 .
- ^ Чунг, Эмили (10 августа 2016 г.). «Популярная теория о том, как люди заселили Северную Америку, не может быть верной, как показывает исследование: свободный ото льда коридор через Альберту, Британская Колумбия, не может использоваться людьми до тех пор, пока не прибудут люди Кловис» . Новости ЦБК . Проверено 10 августа 2016 г.
- ^ Гебель, Тед и Бувит, Ян (2011). От Енисея до Юкона: интерпретация изменчивости каменных комплексов в позднеплейстоценовой/раннеголоценовой Берингии . Центр изучения первых американцев, издательство Техасского университета A&M. п. 5. ISBN 978-1-60344-384-5 .
- ^ Морлан, Ричард Э. (4 марта 2015 г.). «Тассейн Старой Вороны» . Канадская энциклопедия . Историка Канады .
- ^ Хандверк, Брайан (22 июля 2020 г.). «Открытие в мексиканской пещере может радикально изменить известные сроки прибытия людей в Америку» . Смитсоновский журнал .
- ^ Костопулос, Андре (10 ноября 2021 г.). «30-тысячный археологический материал в пещере Чикиуите, раунд 2: все еще не имеет значения, насколько некоторые предметы похожи на каменные орудия» . АрхеоМысли . Проверено 16 сентября 2023 г.
- ^ https://www.science.org/content/article/were-humans-living-mexican-cave-during-last-ice-age
- ^ Виалоу, Д.; Перья; Фонтюнь; Виалоу (2017). «Заселение центра Южной Америки: место позднего плейстоцена Санта-Элина». Античность . 91 (358): 865–884. дои : 10.15184/aqy.2017.101 .
- ^ Родригес, Челиз; Джаннини (2003). «Ранняя антропогенная оккупация и геоморфологические изменения в Южной Америке: взаимодействие человека и окружающей среды и данные OSL с участка Ринкан I, юго-восток Бразилии». Журнал четвертичной науки . 38 (5): 685–701. дои : 10.1002/jqs.3505 . S2CID 258572647 .
- ^ Сантос, генеральный менеджер; Бёрд, Мичиган; Паренти, Ф.; и др. (2003). «Пересмотренная хронология самого нижнего населенного слоя каменного убежища Педра Фурада, Пиауи, Бразилия: плейстоценовое заселение Америки». Четвертичные научные обзоры . 22 (21–22): 2303–2310. Бибкод : 2003QSRv...22.2303S . дои : 10.1016/S0277-3791(03)00205-1 .
- ^ ван Варк, GN; Куизенга, Д. и Уильямс, Флорида (июнь 2003 г.). «Кенневик и Лузия: уроки европейского верхнего палеолита». Американский журнал физической антропологии . 121 (2): 181–184, обсуждение 185–188. дои : 10.1002/ajpa.10176 . ПМИД 12740961 .
• Фидель, Стюарт Дж. (2004). «Кенневикские безумства: «новые» теории о заселении Америки». Журнал антропологических исследований . 60 (1): 75–110. дои : 10.1086/jar.60.1.3631009 . JSTOR 3631009 . S2CID 163722475 .
• Гонсалес-Хосе, Р.; Бортолини, MC; Сантос, Франция и Бонатто, СЛ (октябрь 2008 г.). «Население Америки: изменение формы черепа в континентальном масштабе и его интерпретация с междисциплинарной точки зрения». Американский журнал физической антропологии . 137 (2): 175–187. дои : 10.1002/ajpa.20854 . hdl : 11336/101290 . ПМИД 18481303 . S2CID 32748672 . - ^ Морено-Майяр, Дж. Виктор; Победитель, Лассе; де Баррос Дамгаард, Питер; Фонтана, Констанция; и др. (7 декабря 2018 г.). «Раннее расселение людей по Америке» . Наука . 362 (6419): eaav2621. Бибкод : 2018Sci...362.2621M . дои : 10.1126/science.aav2621 . ПМИД 30409807 .
- ^ Пост, Козимо; Накацука, Натан; Лазаридис, Иосиф; Скоглунд, Понт; и др. (15 ноября 2018 г.). «Реконструкция глубокой истории населения Центральной и Южной Америки» . Клетка . 175 (5): 1185–1197.e22. дои : 10.1016/j.cell.2018.10.027 . ISSN 0092-8674 . ПМК 6327247 . ПМИД 30415837 .
- ^ Холен, Стивен Р.; Демере, Томас А.; Фишер, Дэниел С.; и др. (2017). «Археологический памятник возрастом 130 000 лет в южной Калифорнии, США». Природа . 544 (7651): 479–483. Бибкод : 2017Natur.544..479H . дои : 10.1038/nature22065 . ПМИД 28447646 . S2CID 205255425 .
- ^ Ринкон, Пол (26 апреля 2017 г.). «Первые заявления американцев вызывают споры» . Новости Би-би-си . Проверено 30 апреля 2017 г. .
Майкл Р. Уотерс прокомментировал: «Чтобы продемонстрировать такое раннее заселение Америки, необходимо наличие однозначных каменных артефактов. Не существует однозначных каменных орудий, связанных с костями... это место, вероятно, просто интересное палеонтологическое местонахождение». Крис Стрингер сказал, что «экстраординарные заявления требуют экстраординарных доказательств – каждый аспект требует самого тщательного изучения», добавив, что «должны были потребоваться высокие и концентрированные силы, чтобы разбить самые толстые кости мастодонта, а среда отложения низкой энергии, по-видимому, не обеспечивает очевидной альтернативы людям». используя тяжелые булыжники, найденные вместе с костями.
- ^ Питулько В.В.; Никольский, Пенсильвания; Гиря, Е. Ю; и др. (2 января 2004 г.). «Участок Яны RHS: Люди в Арктике до последнего ледникового максимума». Наука . 303 (5654): 52–56. Бибкод : 2004Sci...303...52P . дои : 10.1126/science.1085219 . ISSN 0036-8075 . ПМИД 14704419 . S2CID 206507352 .
- ^ Jump up to: а б с д и ж Тамм, Эрика; Кивисилд, Тоомас; Рейдла, Маере; и др. (2007). «Берингийское затишье и распространение коренных американцев-основателей» . ПЛОС ОДИН . 2 (9): е829. Бибкод : 2007PLoSO...2..829T . дои : 10.1371/journal.pone.0000829 . ПМК 1952074 . ПМИД 17786201 .
- ^ Jump up to: а б с д Кухня, Эндрю; Миямото, Михал М. и Маллиган, Конни Дж. (2008). «Трехэтапная модель колонизации для заселения Америки» . ПЛОС ОДИН . 3 (2): e1596. Бибкод : 2008PLoSO...3.1596K . дои : 10.1371/journal.pone.0001596 . ПМК 2223069 . ПМИД 18270583 .
- ^ Вей-Хаас, Майя (5 октября 2023 г.). «Новые доказательства того, что древние следы отодвигают прибытие человека в Северную Америку. После исследования следов, найденных в Нью-Мексико в 2021 году, исследователи использовали дополнительные методы, чтобы подтвердить утверждение о том, что этим следам около 23 000 лет» . Нью-Йорк Таймс . Архивировано из оригинала 5 октября 2023 года . Проверено 6 октября 2023 г.
- ^ Пигати, Джеффри С.; и др. (5 октября 2023 г.). «Независимые оценки возраста разрешают споры о древних человеческих следах в Уайт-Сэндс» . Наука . 382 (6666): 73–75. Бибкод : 2023Sci...382...73P . дои : 10.1126/science.adh5007 . ПМИД 37797035 . S2CID 263672291 . Архивировано из оригинала 7 октября 2023 года . Проверено 6 октября 2023 г.
- ^ Суровелл 2022 .
- ^ Jump up to: а б Скоглунд, Понтус и Райх, Дэвид (декабрь 2016 г.). «Геномный взгляд на заселение Америки» (PDF) . Текущее мнение в области генетики и развития . 41 : 27–35. дои : 10.1016/j.gde.2016.06.016 . ПМК 5161672 . ПМИД 27507099 .
Недавно мы провели строгую проверку нулевой гипотезы о единой основной популяции аборигенов Центральной и Южной Америки, используя полногеномные данные разных коренных американцев. Мы обнаружили статистически четкий сигнал, связывающий коренных американцев в амазонском регионе Бразилии с современными австрало-меланезийцами и жителями Андаманских островов («австралазийцами»). В частности, мы обнаружили, что австралийцы имеют значительно больше генетических вариантов с некоторыми популяциями Амазонии, в том числе говорящими на языках тупи, чем с другими коренными американцами. Мы назвали эту предполагаемую древнюю индейскую линию «Популяция Y» в честь Ypykuéra, что означает «предок» в языковой семье тупи.
- ^ Jump up to: а б с Кемп, Брайан М.; Малхи, Рипан С.; Макдонаф, Джон; и др. (2007). «Генетический анализ останков скелетов раннего голоцена с Аляски и его значение для заселения Америки» (PDF) . Американский журнал физической антропологии . 132 (4): 605–621. CiteSeerX 10.1.1.576.7832 . дои : 10.1002/ajpa.20543 . ПМИД 17243155 .
- ^ Jump up to: а б с д и Перего, Уго А.; Ахилли, Алессандро; Ангерхофер, Норман; и др. (2009). «Особенные палеоиндийские миграционные маршруты из Берингии, отмеченные двумя редкими гаплогруппами мтДНК» . Современная биология . 19 (1): 1–8. Бибкод : 2009CBio...19....1P . дои : 10.1016/j.cub.2008.11.058 . ПМИД 19135370 . S2CID 9729731 .
- ^ Jump up to: а б Деренко, Мирослава; Малярчук Борис; Гжибовский, Томаш; и др. (21 декабря 2010 г.). «Происхождение и постледниковое распространение гаплогрупп C и D митохондриальной ДНК в Северной Азии» . ПЛОС ОДИН . 5 (12): e15214. Бибкод : 2010PLoSO...515214D . дои : 10.1371/journal.pone.0015214 . ПМК 3006427 . ПМИД 21203537 .
- ^ Бортолини, Мария-Катира; Сальзано, Франсиско М.; Томас, Марк Г.; и др. (2003). «Свидетельства Y-хромосомы о различных древних демографических историях в Америке» (PDF) . Американский журнал генетики человека . 73 (3): 524–539. дои : 10.1086/377588 . ПМК 1180678 . ПМИД 12900798 .
- ^ Морено-Майяр, Дж. Виктор; Поттер, Бен А.; Виннер, Лассе; Штайнрюкен, Матиас; Расмуссен, Саймон; Терхорст, Джонатан; Камм, Джон А.; Альбрехцен, Андерс; Маласпинас, Анна-Сапфо; Сикора, Мартин; Ройтер, Джошуа Д.; Ирландец, Джоэл Д.; Малхи, Рипан С.; Орландо, Людовик; Сун, Юн С.; Нильсен, Расмус; Мельцер, Дэвид Дж.; Виллерслев, Эске (2018), «Геном терминального плейстоцена Аляски обнаруживает первую популяцию коренных американцев» , Nature , 553 (7687), Macmillan Publishers Limited: 203–207, Бибкод : 2018Natur.553..203M , doi : 10.1038/nature25173 , PMID 29323294 , S2CID 4454580 , получено 3 января 2018 г.
- ^ Доверительные интервалы приведены в Moreno-Mayar et al. (2018):
- 26,1-23-9 тыс. лет назад для отделения восточноазиатской линии ANA от современных восточноазиатских популяций;
- 25-20 тыс. лет назад для события примеси ANE и ранних восточноазиатских линий, предков ANA;
- 22,0-18,1 тыс. лет назад для отделения древнеберингийских от других палеоиндийских линий;
- 17,5-14,6 тыс. лет назад для разделения палеоиндейцев на коренных американцев Северной Америки (ННА) и коренных американцев Южной Америки (СНА).
- ^ Гилл, Хэчан; Ли, Джухён; Чон, Чунгвон (2 апреля 2024 г.). «Реконструкция генетических взаимоотношений между древними и современными популяциями Сибири» . Геномная биология и эволюция . 16 (4): evae063. дои : 10.1093/gbe/evae063 . ISSN 1759-6653 . ПМЦ 10999361 . ПМИД 38526010 .
- ^ Пост, Козимо; Накацука, Натан; Лазаридис, Иосиф; Скоглунд, Понт; Маллик, Свапан; Ламнидис, Тисей К.; Роланд, Надин; Нэгеле, Катрин; Адамски, Николь; Бертолини, Эмили; Брумандхошбахт, Насрин; Купер, Алан; Каллетон, Брендан Дж.; Феррас, Тьяго; Ферри, Мэтью (15 ноября 2018 г.). «Реконструкция глубокой истории населения Центральной и Южной Америки» . Клетка . 175 (5): 1185–1197.e22. дои : 10.1016/j.cell.2018.10.027 . ISSN 0092-8674 . ПМК 6327247 . ПМИД 30415837 .
- ^ Хайнцман, Питер Д.; Фрёзе, Дуэйн; Айвз, Джон В.; Соарес, Андре ЭР; Зазула, Грант Д.; Леттс, Брэндон; Эндрюс, Томас Д.; Драйвер, Джонатан С.; Холл, Элизабет; Заяц, П. Грегори; Джасс, Кристофер Н. (19 июля 2016 г.). «Филогеография бизонов ограничивает распространение и жизнеспособность Свободного ото льда коридора в западной Канаде». Труды Национальной академии наук . 113 (29): 8057–8063. дои : 10.1073/pnas.1601077113 . ISSN 0027-8424 . ПМИД 27274051.
- ^ Общество, National Geographic (19 августа 2019 г.). «Культура охотников-собирателей». Национальное географическое общество . Проверено 18 февраля 2022 г.
- ^ Леонард, Дженнифер А.; Вила, Карлес; Фокс-Доббс, Кена; Кох, Пол Л.; Уэйн, Роберт К.; Ван Валкенбург, Блэр (3 июля 2007 г.). «Вымирание мегафауны и исчезновение специализированного волчьего экоморфа». Современная биология . 17 (13): 1146–1150. дои : 10.1016/j.cub.2007.05.072 . ISSN 0960-9822 .
- ^ Jump up to: а б Ян, Мелинда А. (6 января 2022 г.). «Генетическая история миграции, диверсификации и примеси в Азии» . Популяционная генетика и геномика человека . 2 (1): 1–32. дои : 10.47248/hpgg2202010001 . ISSN 2770-5005 . Рагхаван и др. [51] использовали f3-статистику аутгруппы, чтобы показать, что Мальта1 была наиболее тесно связана с современными коренными американцами. Эта закономерность привела к модели, согласно которой расселение людей по Америке произошло из популяции Сибирь произошла от смеси двух различных предков, одна из которых связана с сегодняшними выходцами из Восточной Азии, а другая — с Мальтой1. Недавние исследования ДНК древних сибиряков [69,73] предоставили прямые доказательства смешанных популяций в Сибири, которые тесно связаны с ними. Родословная коренных американцев».
- ^ Чжан, Сяомин Цзи; Ли, Чуньмэй; Хуан, Цзяхуэй; У, Юнь; 07-25). Геном человека позднего плейстоцена из Юго-Западного Китая» 32 . « ( ): 3095–3109.e5 Бибкод : 2022CBio...32E3095Z . doi : 10.1016/j.cub.2022.06.016 14 ISSN 0960-9822 PMID 35839766 S2CID 250502011 .
- ^ Рафф, Дженнифер (8 февраля 2022 г.). Происхождение: Генетическая история Америки . Издательство Гранд Сентрал. п. 188. ИСБН 978-1-5387-4970-8 . «Текущие оценки показывают, что примерно 63% предков первых народов происходят из восточноазиатской группы, а остальные — из древних северных сибиряков. Мы не уверены, где произошло это взаимодействие. Некоторые археологи полагают, что оно произошло в Восточной Азии. предполагая, что именно сюда сибиряки переселились во время LGM" [...] "На основании генетических данных также можно предположить, что это взаимодействие имело место в районе озера Байкал в Сибири" [...] " Но другие археологи и генетики утверждают, что встреча двух прародительских популяций коренных американцев произошла потому, что люди двинулись на север, а не на юг, в ответ на LGM».
- ^ Рафф 2022 , с. 188: «Население Мальты — древние северные сибиряки, по-видимому, столкнулось с дочерней восточноазиатской популяцией, описанной в начале этой главы, около 25 000 лет назад и скрестилось с ними».… «Мы не уверены, где это взаимодействие имело место. Некоторые археологи полагают, что оно произошло в Восточной Азии, предполагая, что именно сюда сибиряки переселились во время LGM»... «Есть также основания полагать, что это взаимодействие имело место в районе озера Байкал. Сибирь также основана на генетических данных. Древние палеосибиряки, как упоминалось ранее в этой главе, отделились от восточноазиатских предков коренных американцев примерно 25 000 лет назад. Они известны нам по геномам человека верхнего палеолита. Регион озера Байкал, известный нам по геномам человека верхнего палеолита из региона озера Байкал, известного как UKY, и человека из Северо-Восточной Сибири, датируемого примерно 9800 лет назад, известного как Колыма1».
- ^ Рафф 2022 , стр. 188–189: «Но другие археологи и генетики утверждают, что встреча двух прародительских популяций коренных американцев произошла потому, что люди двинулись на север, а не на юг, в ответ на LGM. В этом сценарии палео-сибирский потомки, такие как UKY, могли быть результатом повторного заселения Сибири на юг из Берингии. Причина этого в том, что как митохондриальный, так и ядерный геномы коренных американцев показывают, что они были изолированы от всех других популяций в течение длительного периода времени. Это открытие, первоначально основанное на классических генетических маркерах и митохондриальных данных, стало известно как гипотеза Берингийской инкубации, Берингийской паузы или Берингийской гипотезы застоя».
- ^ Гребенюк Павел С.; Федорченко Александр Юрьевич; Дьяконов Виктор М.; Лебединцев Александр Иванович; Малярчук, Борис А. (2022). «Древние культуры и миграции в Северо-Восточной Сибири» . Люди в сибирских пейзажах . Спрингер География. Международное издательство Спрингер. п. 96. дои : 10.1007/978-3-030-90061-8_4 . ISBN 978-3-030-90060-1 .
По новейшим палеогенетическим данным, восточноазиатские популяции мигрировали в Северо-Восточную Сибирь ок. 20 000–18 000 кал BP. Миграция сопровождалась их смешением с потомками «древних северных сибиряков», представленными геномом янских и мальтийских особей. Эти процессы отразились в широком распространении берингийской традиции в регионе и привели к появлению на Крайнем Северо-Востоке Азии нескольких родовых линий (рис. 1): древнего палеосибирского населения, представленного геномом особи из Дуванного Яра, и предкового Коренные американцы. Последний тип впоследствии разделился на древних берингийцев и всех остальных коренных американцев (Морено-Майар и др., 2018; Сикора и др., 2019).
- ^ Хоффекер, Джон Ф. (4 марта 2016 г.). «Берингия и глобальное расселение современного человека: Берингия и глобальное расселение современного человека» . Эволюционная антропология: проблемы, новости и обзоры . 25 (2): 64–78. дои : 10.1002/evan.21478 . ПМИД 27061035 . S2CID 3519553 . «Паттерн, наблюдаемый в данных мтДНК Таммом и его коллегами, еще не проявился с одинаковой ясностью ни в Y-ДНК, ни в аутосомной ДНК. Отсутствие аналогичного паттерна в данных Y-ДНК, по-видимому, отражает различную историю отцовской и материнской линии. — закономерность, обнаруженная в других частях мира (например, в Южной Азии).33 Некоторое подтверждение гипотезы застоя в аутосомной ДНК можно найти в распределении аллеля в микросателлитном локусе на хромосоме 9 (D9S1120), которое демонстрирует закономерность аналогично тому, что наблюдается в данных мтДНК.34»
- ^ Сикора, Мартин; Питулко Владимир Владимирович; Соуза, Витор К.; и др. (2019). «История населения Северо-Восточной Сибири начиная с плейстоцена» . Природа . 570 (7760): 182–188. Бибкод : 2019Natur.570..182S . дои : 10.1038/s41586-019-1279-z . hdl : 1887/3198847 . ПМИД 31168093 . S2CID 174809069 .
- ^ Чжан, Сяомин Цзи; Ли, Чуньмэй; Хуан, Цзяхуэй; У, Юнь; Пан, Хуан, Яньи; Су, Бин; . «Геном человека позднего плейстоцена из Юго-Западного Китая» Текущая биология . 3095–3109.e5 . Bibcode : 2022CBio...32E3095Z doi : 10.1016 / . J.CUB.2022.06.016 . 9822. PMID 35839766. . S2CID 250502011 .
- ^ «ДНК древнего населения Южного Китая предполагает восточноазиатские корни коренных американцев» .
- ^ Jump up to: а б с д Адачи, Нобору; Шинода, Кен-ичи; Уметсу, Кадзуо и Мацумура, Хирофуми (март 2009 г.). «Анализ митохондриальной ДНК скелетов Дзёмон из стоянки Фунадомари, Хоккайдо, и его значение для происхождения коренных американцев». Американский журнал физической антропологии . 138 (3): 255–265. дои : 10.1002/ajpa.20923 . ПМИД 18951391 .
- ^ Jump up to: а б Шурр, Теодор Г. (май 2000 г.). «Митохондриальная ДНК и заселение Нового Света» (PDF) . Американский учёный . 88 (3): 246. Бибкод : 2000AmSci..88..246S . дои : 10.1511/2000.3.246 . S2CID 7527715 .
- ^ Захаров И.А.; Деренко, М.В.; Малярчук, Б.А.; и др. (12 января 2006 г.). «Вариации митохондриальной ДНК аборигенных популяций Алтае-Байкалья: значение для генетической истории Северной Азии и Америки». Анналы Нью-Йоркской академии наук . 1011 (1): 21–35. Бибкод : 2004NYASA1011...21Z . дои : 10.1196/анналы.1293.003 . ПМИД 15126280 . S2CID 37139929 .
- ^ Jump up to: а б с д и Стариковская Елена Борисовна; Сукерник, Рем И.; Дербенева Ольга Александровна; и др. (январь 2005 г.). «Разнообразие митохондриальной ДНК у коренных народов южной части Сибири и происхождение гаплогрупп коренных американцев» . Анналы генетики человека . 69 (Часть 1): 67–89. дои : 10.1046/j.1529-8817.2003.00127.x . ПМЦ 3905771 . ПМИД 15638829 .
- ^ Адачи, Нобору; Шинода, Кен-ичи; Уметсу, Кадзуо; и др. (ноябрь 2011 г.). «Анализ митохондриальной ДНК скелетов Дзёмон Хоккайдо: остатки архаичных материнских линий на юго-западной окраине бывшей Берингии». Американский журнал физической антропологии . 146 (3): 346–360. дои : 10.1002/ajpa.21561 . ПМИД 21953438 .
- ^ Дай волю, Роберто; Баттонс, Патриция; Джардина, Бруно (8 марта 2012 г.). Достижения митохондриальной медицины . Springer Science & Business Media. п. 446. ИСБН 978-94-007-2869-1 .
- ^ Рафф, Дженнифер А.; Больник, Дебора А. (октябрь 2015 г.). «Указывает ли митохондриальная гаплогруппа X на древнюю трансатлантическую миграцию в Америку? Критическая переоценка» . ПалеоАмерика . 1 (4): 297–304. дои : 10.1179/2055556315Z.00000000040 . ISSN 2055-5563 . S2CID 85626735 . «Все эти исследования пришли к одному и тому же выводу и предполагают, что гаплогруппа X2a, вероятно, возникла в той же популяции (популяциях), что и другие американские гаплогруппы-основатели, в силу сопоставимых дат слияния и демографической истории» ... «X2a имеет сопоставимые даты слияния и демографическую историю». не был обнаружен нигде в Евразии, и филогеография не дает нам убедительных оснований полагать, что он, скорее, пришел из Европы, чем из Сибири. Более того, анализ полного генома кенневикского человека, который принадлежит к самой базовой линии X2a, когда-либо идентифицированной. , не дает никаких указаний на недавнее европейское происхождение и перемещает расположение самой глубокой ветви X2a на западное побережье, что соответствует принадлежности X2a к той же наследственной популяции, что и другие митохондриальные гаплогруппы-основатели».
- ^ Ли, Хун-Чуань; Биггар, Роберт Дж.; Майли, Венделл Дж.; и др. (2004). «Провирусная нагрузка в грудном молоке и риск передачи Т-лимфотропного вируса человека типа I от матери ребенку» . Журнал инфекционных болезней . 190 (7): 1275–1278. дои : 10.1086/423941 . ПМИД 15346338 .
- ^ Jump up to: а б с д Вердонк, К.; Гонсалес Э.; Ван Доорен, С.; и др. (апрель 2007 г.). «Человеческий Т-лимфотропный вирус 1: последние знания о древней инфекции». Ланцет инфекционных заболеваний . 7 (4): 266–281. дои : 10.1016/S1473-3099(07)70081-6 . ПМИД 17376384 .
- ^ Гессен, А.; Галло, Р.К. и Франкини, Г. (апрель 1992 г.). «Низкая степень генетического дрейфа вируса Т-клеточного лейкоза/лимфомы человека типа I in vivo как средство мониторинга передачи вируса и перемещения древних человеческих популяций» . Журнал вирусологии . 66 (4): 2288–2295. doi : 10.1128/JVI.66.4.2288-2295.1992 . ПМК 289023 . ПМИД 1548762 .
- ^ Исида, Такафуми; Ямамото, Котаро; Омото, Кэйичи; и др. (сентябрь 1985 г.). «Распространенность человеческого ретровируса у коренных японцев: свидетельства возможного древнего происхождения». Журнал инфекции . 11 (2): 153–157. дои : 10.1016/s0163-4453(85)92099-7 . ПМИД 2997332 .
- ^ Jump up to: а б Миура, Т.; Фукунага, Т.; Игараси, Т.; и др. (февраль 1994 г.). «Филогенетические подтипы Т-лимфотропного вируса человека типа I и их связь с антропологическим фоном» . Труды Национальной академии наук Соединенных Штатов Америки . 91 (3): 1124–1127. Бибкод : 1994PNAS...91.1124M . дои : 10.1073/pnas.91.3.1124 . ПМК 521466 . ПМИД 8302841 .
- ^ Пикард, Ф.Дж.; Култхарт, МБ; Огер, Дж.; и др. (ноябрь 1995 г.). «Человеческий Т-лимфотропный вирус типа 1 у прибрежных жителей Британской Колумбии: филогенетическое сходство и возможное происхождение» . Журнал вирусологии . 69 (11): 7248–56. doi : 10.1128/JVI.69.11.7248-7256.1995 . ЧВК 189647 . ПМИД 7474147 .
- ^ Ли, Хун-Чуань; Фудзёси, Тосинобу; Лу, Хонг; и др. (декабрь 1999 г.). «Наличие ДНК провируса древнего человеческого Т-клеточного лимфотропного вируса типа I в андской мумии». Природная медицина . 5 (12): 1428–1432. дои : 10.1038/71006 . ПМИД 10581088 . S2CID 12893136 .
- ^ Jump up to: а б Култхарт, Майкл Б.; Посада, Дэвид; Крэндалл, Кейт А. и Декабанд, Грегори А. (март 2006 г.). «О филогенетическом размещении последовательностей вируса Т-клеточного лейкоза человека типа 1, связанных с андской мумией» . Инфекция, генетика и эволюция . 6 (2): 91–96. дои : 10.1016/j.meegid.2005.02.001 . ЧВК 1983367 . ПМИД 16503510 .
- ^ Jump up to: а б Гонсалеса, Сильвия; Хаддарт, Дэвид; Исраде-Алькантара, Изабель; и др. (30 марта 2015 г.). «Палеоиндийские памятники из Мексиканского бассейна: данные стратиграфии, тефрохронологии и датировки» (PDF) . Четвертичный интернационал . 363 : 4–19. Бибкод : 2015QuInt.363....4G . дои : 10.1016/j.quaint.2014.03.015 .
- ^ Jump up to: а б Гонсалес-Хосе, Роландо; Гонсалес-Мартин, Антонио; Эрнандес, Микель; и др. (4 сентября 2003 г.). «Краниометрические доказательства выживания палеоамериканцев в Нижней Калифорнии». Природа . 425 (6953): 62–65. Бибкод : 2003Natur.425...62G . дои : 10.1038/nature01816 . ПМИД 12955139 . S2CID 4423359 .
- ^ Диллехей, Томас Д. (4 сентября 2003 г.). «По следам первых американцев». Природа . 425 (6953): 23–24. дои : 10.1038/425023а . ПМИД 12955120 . S2CID 4421265 .
- ^ Фидель, Стюарт Дж. (весна 2004 г.). «Безумие Кенневика: «новые» теории о заселении Америки». Журнал антропологических исследований . 60 (1): 75–110. дои : 10.1086/jar.60.1.3631009 . JSTOR 3631009 . S2CID 163722475 .
- ^ Чаттерс, Джеймс К.; Кеннетт, Дуглас Дж.; Асмером, Йеман; и др. (16 мая 2014 г.). «Скелет человека позднего плейстоцена и мтДНК связывают палеоамериканцев и современных коренных американцев» (PDF) . Наука . 344 (6185): 750–754. Бибкод : 2014Sci...344..750C . дои : 10.1126/science.1252619 . ПМИД 24833392 . S2CID 206556297 . Архивировано из оригинала (PDF) 13 июля 2015 г.
- ^ Виллерслев, Эске; Мельцер, Дэвид Дж. (17 июня 2021 г.). «Заселение Америки по данным древней геномики». Природа . 594 (7863): 356–364. Бибкод : 2021Natur.594..356W . дои : 10.1038/s41586-021-03499-y . ПМИД 34135521 . S2CID 235460793 .
- ^ де Азведо, Соледад; Бортолини, Мария К.; Бонатто, Сандро Л.; и др. (январь 2015 г.). «Древние останки и первое заселение Америки: переоценка черепа негра Хойо». Американский журнал физической антропологии . 148 (3): 514–521. дои : 10.1002/ajpa.22801 . hdl : 11336/42446 . ПМИД 26174009 .
• Азеведу, Соледад де; Квинто-Санчес, Мирша; Пашетта, Каролина и Гонсалес-Хосе, Роландо (28 февраля 2017 г.). « Первое человеческое поселение Нового Света. Более пристальный взгляд на черепно-лицевые вариации и эволюцию групп коренных американцев раннего и позднего голоцена» . Четвертичный интернационал . 431 (часть Б): 152–167. Бибкод : 2017QuInt.431..152D . дои : 10.1016/j.quaint.2015.11.012 . hdl : 11336/44745 . - ^ Сон, Чунтэк (декабрь 2008 г.). «Острия, микролезвия и охота позднего палеолита в Корее». Античность . 82 (318): 871–883. дои : 10.1017/s0003598x00097647 . S2CID 127994558 .
- ^ Уотерс, Майкл Р. и Стаффорд, Томас В. (23 февраля 2007 г.). «Переопределение возраста Хлодвига: последствия для заселения Америки». Наука . 315 (5815): 1122–1126. Бибкод : 2007Sci...315.1122W . дои : 10.1126/science.1137166 . ПМИД 17322060 . S2CID 23205379 .
- ^ Дженкинс, Деннис Л.; Дэвис, Лорен Г.; Стаффорд, Томас В. младший; и др. (13 июля 2012 г.). «Западные наконечники снарядов эпохи Хлодвига и человеческие копролиты в пещерах Пейсли» . Наука . 337 (6091): 223–228. Бибкод : 2012Sci...337..223J . дои : 10.1126/science.1218443 . ПМИД 22798611 . S2CID 40706795 .
- ^ Jump up to: а б Адовазио, Дж. М.; Донахью Дж. и Стукенрат Р. (1990). «Радиоуглеродная хронология Медоукрофт Рокшелтер 1975–1990». Американская древность . 55 (2): 348–354. дои : 10.2307/281652 . JSTOR 281652 . S2CID 163541173 .
- ^ Кларк, Джори; Карлсон, Андерс Э.; Рейес, Альберто В.; Карлсон, Элизабет CB; Гийом, Луиза; Милн, Гленн А.; Тарасов Лев; Кофе, Марк; Вилкен, Клаус; Руд, Дилан Х. (5 апреля 2022 г.). «Эпоха открытия незамерзающего коридора и последствия для заселения Америки» . Труды Национальной академии наук . 119 (14). дои : 10.1073/pnas.2118558119 . ISSN 0027-8424 . ПМЦ 9168949 . ПМИД 35312340 .
- ^ Альтшул, Джеффри; Джонсон, Уильям К.; Стернер, Мэтью А.; Инженерный корпус армии, округ Лос-Анджелес; Инженерный корпус армии, округ Лос-Анджелес; Статистические исследования, Inc.; НИИ Пресс (1989). Участок Дип-Крик (CA-SBr-176): поздний доисторический базовый лагерь в районе Мохаве-Ривер-Форкс, округ Сан-Бернардино, Калифорния . Техническая серия статистических исследований. Пол Д. Буи, Томас М. Оригер. Тусон, Аризона: SRI Press.
- ^ Jump up to: а б Вайда, Эдвард Дж. (18 апреля 2017 г.). «Дене-Енисейский» . Оксфордские библиографии в Интернете . дои : 10.1093/OBO/9780199772810-0064 .
- ^ Флегонтов Павел; Алтынышик, Н. Эзги; Чангмай, Пия; и др. (13 октября 2017 г.). «Палеоэскимосское генетическое наследие Северной Америки» . bioRxiv : 203018. doi : 10.1101/203018 . hdl : 21.11116/0000-0004-5D08-C . S2CID 90288469 .
• Флегонтов Павел; Алтынышик, Н. Эзги; Чангмай, Пия; и др. (5 июня 2019 г.). «Палео-эскимосское генетическое происхождение и заселение Чукотки и Северной Америки» (PDF) . Природа . 570 (7760): 236–240. Бибкод : 2019Natur.570..236F . дои : 10.1038/s41586-019-1251-y . ISSN 0028-0836 . ПМЦ 6942545 . ПМИД 31168094 . - ^ Хандверк, Брайан (12 февраля 2010 г.). «Лицо древнего человека, нарисованное из ДНК волос; геном рисует картину человека вымершей гренландской культуры» . Национальные географические новости . Архивировано из оригинала 1 октября 2019 года.
- ^ Фладмарк, Кнут Р. (январь 1979 г.). «Маршруты: альтернативные миграционные коридоры для раннего человека в Северной Америке». Американская древность . 44 (1): 55–69. дои : 10.2307/279189 . JSTOR 279189 . S2CID 162243347 .
- ^ Каллауэй, Юэн (11 августа 2016 г.). «ДНК растений и животных позволяет предположить, что первыми американцы выбрали прибрежный маршрут» . Природа . 536 (7615): 138. Бибкод : 2016Natur.536..138C . дои : 10.1038/536138а . ПМИД 27510205 .
- ^ Лето, Томас (10 августа 2016 г.). «Люди, возможно, выбрали другой путь в Америку, чем думали, что арктический проход не обеспечил бы достаточно еды для первых путешествий американцев» . Новости науки .
- ^ Расмуссен, Мортен; Анзик, Сара Л.; Уотерс, Майкл Р.; Скоглунд, Понт; ДеДжорджио, Майкл; Стаффорд, Томас В. младший; и др. (февраль 2014 г.). «Геном человека позднего плейстоцена из захоронения Кловиса в западной Монтане» . Природа . 506 (7487): 225–229. Бибкод : 2014Natur.506..225R . дои : 10.1038/nature13025 . ПМЦ 4878442 . ПМИД 24522598 .
- ^ Горниц, Вивиан (январь 2007 г.). «Подъем уровня моря после таяния льда и сегодня» . Годдардский институт космических исследований . НАСА . Архивировано из оригинала 2 февраля 2007 г. Проверено 23 апреля 2015 г.
- ^ Хетерингтон, Рене; Барри, Дж. Вон; Маклауд, Роджер и Уилсон, Майкл (февраль 2004 г.). «В поисках затерянной земли» . Геотаймс .
- ^ «Калифорнийские острова отказываются от свидетельств раннего мореплавания: многочисленные артефакты, найденные на стоянках позднего плейстоцена на Нормандских островах» . Наука Дейли . Университет Орегона. 3 марта 2011 г.
Дальнейшее чтение
[ редактировать ]- Бесерра-Вальдивия, Лорена и Томас Хайэм. (2020) «Время и последствия самого раннего прибытия человека в Северную Америку». Природа 584.7819 (2020): 93-97. онлайн
- Брэдли, Брюс и Стэнфорд, Деннис Дж. (2004). «Коридор кромки льда в Северной Атлантике: возможный палеолитический путь в Новый Свет». Мировая археология . 36 (4): 459–478. CiteSeerX 10.1.1.694.6801 . дои : 10.1080/0043824042000303656 . S2CID 161534521 .
- Брэдли, Брюс и Стэнфорд, Деннис Дж. (2006). «Связь Солютрея и Хлодвига: ответ Штраусу, Мельцеру и Гебелю». Мировая археология . 38 (4): 704–714. дои : 10.1080/00438240601022001 . JSTOR 40024066 . S2CID 162205534 .
- Стэнфорд, Деннис Дж.; Брэдли, Брюс (2012). Первые американцы до Хлодвига: происхождение американской культуры Кловиса . Издательство Калифорнийского университета. ISBN 978-0-520-22783-5 .
- Стэнфорд, Деннис Дж. и Брэдли, Брюс А. (2013). Через Атлантический лед: Происхождение американской культуры Кловис . Издательство Калифорнийского университета. ISBN 978-0-520-27578-2 .
- Диксон, Э. Джеймс (1993). Поиски происхождения первых американцев . Университет Нью-Мексико. ISBN 978-0-8263-1406-2 .
- Диксон, Э. Джеймс (1999). Кости, лодки и бизоны: археология и первая колонизация западной части Северной Америки . Издательство Университета Нью-Мексико. ISBN 978-0-8263-2138-1 .
- Эрландсон, Джон М. (2013). Ранние охотники-собиратели побережья Калифорнии . Springer Science & Business Media. ISBN 978-1-4757-5042-3 .
- Эрландсон, Джон М. (2001). «Археология водных адаптаций: парадигмы нового тысячелетия». Журнал археологических исследований . 9 (4): 287–350. дои : 10.1023/а:1013062712695 . S2CID 11120840 .
- Эрландсон, Джон М. (2002). «Анатомически современные люди, морские путешествия и плейстоценовая колонизация Америки» . В Нине Г. Яблонски (ред.). Первые американцы: плейстоценовая колонизация Нового Света . Калифорнийская академия наук. стр. 59–92. ISBN 978-0-940228-50-4 .
- Эрландсон, Джон. М.; Грэм, Миннесота; Бурк, Брюс Дж.; и др. (30 октября 2007 г.). «Гипотеза о шоссе из водорослей: морская экология, теория прибрежной миграции и заселение Америки». Журнал островной и прибрежной археологии . 2 (2): 161–174. дои : 10.1080/15564890701628612 . S2CID 140188874 .
- Эшлеман, Джейсон А.; Малхи, Рипан С. и Гленн Смит, Дэвид (2003). «Исследования митохондриальной ДНК коренных американцев: концепции и заблуждения о предыстории населения Америки». Эволюционная антропология . 12 (1): 7–18. дои : 10.1002/evan.10048 . S2CID 17049337 .
- Федже, Дэрил В. и Кристенсен, Тина (октябрь 1999 г.). «Моделирование палеобереговых линий и обнаружение прибрежных участков раннего голоцена в Хайда-Гвайи». Американская древность . 64 (4): 635–652. дои : 10.2307/2694209 . JSTOR 2694209 . S2CID 163478479 .
- Гринман, Э.Ф. (февраль 1963 г.). «Верхний палеолит и Новый Свет». Современная антропология . 4 (1): 41–66. дои : 10.1086/200337 . JSTOR 2739818 . S2CID 144250630 .
- Привет, Джоди (25 мая 2005 г.). «О количестве основателей нового мира: популяционно-генетический портрет населения Америки» . ПЛОС Биология . 3 (6): e193. doi : 10.1371/journal.pbio.0030193 . ПМЦ 1131883 . ПМИД 15898833 .
- Яблонски, Нина Г. (2002). Первые американцы: плейстоценовая колонизация Нового Света . Калифорнийская академия наук . ISBN 978-0-940228-50-4 .
- Джонс, Питер Н. (2005). Уважение к предкам: культурная принадлежность американских индейцев на американском Западе . Институт Бауу. ISBN 978-0-9721349-2-7 .
- Коротаев Андрей; Березкин Юрий Евгеньевич; Боринская Светлана А.; Давлетшин Альберт И.; Халтурина, Дарья А. (2017). «Какие гены и мифы принесли в Новый Свет разные волны заселения Америки?» . У Леонида Евгеньевича Гринина; Андрей В. Коротаев; Юрий Евгеньевич Березкин (ред.). История и математика: экономика, демография, культура и космические цивилизации . ООО «Издательство «Учитель». С. 9–77. ISBN. 978-5-7057-5247-8 .
- Лаубер, Патрисия (2003). Кто пришел первым: новые сведения о доисторических американцах . Детские книги National Geographic Soc. ISBN 978-0-7922-8228-0 .
- Мэтсон, Р.Г. и Коупленд, Гэри (2016). Предыстория Северо-Западного побережья . Тейлор и Фрэнсис. ISBN 978-1-315-41739-4 .
- Мельцер, Дэвид Дж. (2009). Первые народы в новом мире: колонизация Америки ледникового периода . Издательство Калифорнийского университета. ISBN 978-0-520-94315-5 .
- Сноу, Дин Р. (1996). «Первые американцы и дифференциация культур охотников-собирателей» . В Брюсе Г. Триггере; Уилкомб Э. Уошберн (ред.). Кембриджская история коренных народов Америки: Северная Америка . Том. 1: Часть 1. Издательство Кембриджского университета. стр. 125–199. ISBN 978-0-521-57392-4 .
- Уэллс, Спенсер (2002). Путешествие человека: генетическая одиссея . Издательство Принстонского университета. ISBN 0-691-11532-Х .
Внешние ссылки
[ редактировать ]- База данных палеоиндейцев – Университет Теннесси, факультет антропологии .
- «Когда люди пришли в Америку?» – Журнал Smithsonian Magazine , февраль 2013 г. Архивировано 9 декабря 2013 г. в Wayback Machine.
- Палеоиндийский период - Министерство внутренних дел США, Служба национальных парков.
- Шепард Креч III, Палеоиндейцы и великое вымирание плейстоцена – Американская академия искусств и наук, Национальный гуманитарный центр , 2008.
- «Путешествие человека: Генетическая одиссея» (фильм) на YouTube – Спенсер Уэллс – PBS и National Geographic Channel , 2003 – 120 минут, UPC/EAN: 841887001267