Y-хромосома
| Y-хромосома человека | |
|---|---|
Y-хромосома человека (после G-бэндинга ) | |
 Y-хромосома в мужской кариограмме человека | |
| Функции | |
| Длина ( б.н. ) | 62 460 029 п.н. (CHM13) |
| Количество генов | 63 ( ССДС ) [ 1 ] |
| Тип | Аллосома |
| Положение центромеры | Акроцентрический [ 2 ] (10,4 Мбит/с) [ 3 ] ) |
| Полные списки генов | |
| CCDS | Список генов |
| HGNC | Список генов |
| ЮниПрот | Список генов |
| NCBI | Список генов |
| Внешние программы просмотра карт | |
| Вместе | Хромосома Y |
| Входить | Хромосома Y |
| NCBI | Хромосома Y |
| УКСК | Хромосома Y |
| Полные последовательности ДНК | |
| RefSeq | NC_000024 ( ИСПРАВЛЕНО ) |
| ГенБанк | CM000686 ( ИСПРАВЛЕНО ) |
— Y-хромосома одна из двух половых хромосом у териевых млекопитающих и других организмов . Наряду с X-хромосомой , она является частью системы определения пола XY , в которой Y является определяющей пол хромосомой, поскольку наличие Y-хромосомы приводит к тому, что потомство, полученное в результате полового размножения, будет мужского пола . У млекопитающих Y-хромосома содержит ген SRY , который запускает развитие мужских гонад . Y-хромосома передается только от родителей мужского пола потомству мужского пола.
Обзор
[ редактировать ]Открытие
[ редактировать ]Y-хромосома была идентифицирована как определяющая пол хромосома Нетти Стивенс из колледжа Брин-Мор в 1905 году во время исследования мучного червя Tenebrio molitor . Эдмунд Бичер Уилсон независимо открыл те же механизмы в том же году, работая с полужесткокрылыми . Стивенс предположил, что хромосомы всегда существовали парами и что меньшая хромосома (теперь обозначенная буквой «Y») была парой Х-хромосомы, открытой в 1890 году Германом Хенкингом . Она поняла, что предыдущая идея Кларенса Эрвина МакКланга о том, что Х-хромосома определяет пол, была ошибочной и что определение пола на самом деле происходит благодаря наличию или отсутствию Y-хромосомы. В начале 1920-х годов Теофил Пейнтер установил, что X- и Y-хромосомы определяют пол человека (и других млекопитающих). [ 4 ]
Хромосоме было присвоено имя «Y» просто для того, чтобы следовать по алфавиту от «X» Хенкинга. [ 5 ] [ 6 ] Идея о том, что Y-хромосома была названа из-за внешнего сходства с буквой «Y», ошибочна. Все хромосомы обычно выглядят под микроскопом как аморфная капля и принимают четко очерченную форму только во время митоза . Эта форма отдаленно напоминает Х-образную для всех хромосом. Совершенно случайно, что Y-хромосома во время митоза имеет две очень короткие ветви, которые под микроскопом могут выглядеть слившимися и выглядеть как потомок Y-образной ветви. [ 5 ] : 65–66
Вариации
[ редактировать ]У большинства териевых млекопитающих в каждой клетке имеется только одна пара половых хромосом. У мужчин есть одна Y-хромосома и одна Х-хромосома , а у женщин — две Х-хромосомы. У млекопитающих Y-хромосома содержит ген SRY , который запускает эмбриональное развитие самца. Y-хромосомы человека и других млекопитающих также содержат другие гены, необходимые для нормального производства спермы. [ нужна ссылка ]
Однако есть исключения. Среди людей у некоторых мужчин рождаются два X и Y («XXY», см. Синдром Клайнфельтера ), один X и два Y (см. Синдром XYY ). У некоторых женщин есть три Х ( трисомия Х ), а у некоторых — один Х вместо двух («Х0», см. синдром Тернера ). Существуют и другие варианты, при которых во время развития ген WNT4 эмбрионального [ 7 ] активируется и/или ген SRY повреждается, что приводит к рождению самки XY (синдром Свайера [ 7 ] ). AY-хромосома также может присутствовать, но не приводит к развитию мужского фенотипа у людей с синдромом нечувствительности к андрогенам , а вместо этого приводит к женскому или неоднозначному фенотипу. В других случаях ген SRY копируется в X, что приводит к рождению самца XX . [ 8 ]
Происхождение и эволюция
[ редактировать ]Перед Y-хромосомой
[ редактировать ]Многие эктотермные позвоночные не имеют половых хромосом. [ 9 ] Если эти виды имеют разный пол, пол определяется окружающей средой, а не генетически. У некоторых видов, особенно рептилий , пол зависит от температуры инкубации. [ 10 ] Некоторые позвоночные являются гермафродитами , хотя гермафродитные виды чаще всего являются последовательными , то есть организм меняет пол, производя мужские или женские гаметы в разные моменты своей жизни, но никогда не производя их одновременно. Это противоположность одновременному гермафродитизму, когда один и тот же организм производит мужские и женские гаметы одновременно. Большинство видов одновременных гермафродитов являются беспозвоночными, а среди позвоночных одновременный гермафродитизм обнаружен лишь у нескольких отрядов рыб. [ 11 ]
Источник
[ редактировать ]Считается, что Х- и Y-хромосомы произошли от пары идентичных хромосом. [ 12 ] [ 13 ] называемые аутосомами , когда у предкового животного развилась аллельная вариация (так называемый «половой локус») и простое обладание этой аллелью сделало организм мужским. [ 14 ] Хромосома с этим аллелем стала Y-хромосомой, а другой член пары — Х-хромосомой. Со временем гены, которые были полезны для мужчин и вредны для женщин (или не влияли на них), либо развивались на Y-хромосоме, либо были приобретены Y-хромосомой в процессе транслокации . [ 15 ]
До недавнего времени считалось, что X- и Y-хромосомы млекопитающих разошлись около 300 миллионов лет назад. [ 16 ] Однако исследование, опубликованное в 2008 году и анализирующее утконоса , геном [ 17 ] предположили, что система определения пола XY не существовала более 166 миллионов лет назад, когда однопроходные отделились от других млекопитающих. [ 18 ] Эта переоценка возраста терианской системы XY основана на обнаружении того, что последовательности, находящиеся в Х-хромосомах сумчатых и плацентарных млекопитающих, не присутствуют в аутосомах утконоса и птиц. [ 18 ] Более старая оценка была основана на ошибочных сообщениях о том, что Х-хромосомы утконоса содержат эти последовательности. [ 19 ] [ 20 ]
Ингибирование рекомбинации
[ редактировать ]Большинство хромосом рекомбинируют во время мейоза. Однако у мужчин пары X и Y находятся в общей области, известной как псевдоаутосомная область (PAR). [ 21 ] PAR подвергается частой рекомбинации между X- и Y-хромосомами. [ 21 ] но рекомбинация подавляется в других участках Y-хромосомы. [ 14 ] Эти регионы содержат гены, определяющие пол, и другие гены, специфичные для мужчин. [ 22 ] Без этого подавления эти гены могут быть потеряны из Y-хромосомы в результате рекомбинации и вызвать такие проблемы, как бесплодие. [ 23 ]
Отсутствие рекомбинации в большей части Y-хромосомы делает ее полезным инструментом в изучении эволюции человека , поскольку рекомбинация усложняет математические модели, используемые для отслеживания предков. [ 24 ]
Вырождение
[ редактировать ]По одной из оценок, Y-хромосома человека за время своего существования потеряла 1393 из 1438 исходных генов, и линейная экстраполяция этой потери 1393 генов за 300 миллионов лет дает скорость генетической потери 4,6 генов на миллион лет. [ 25 ] Продолжающаяся потеря генов со скоростью 4,6 генов на миллион лет приведет к тому, что Y-хромосома останется без функциональных генов - то есть Y-хромосома потеряет полную функцию - в течение следующих 10 миллионов лет или половины этого времени по текущей оценке возраста. 160 миллионов лет. [ 14 ] [ 26 ] Сравнительный геномный анализ показывает, что многие виды млекопитающих испытывают аналогичную потерю функции гетерозиготной половой хромосомы. Дегенерация может стать судьбой всех нерекомбинирующих половых хромосом из-за трех общих эволюционных сил: высокой частоты мутаций , неэффективного отбора и генетического дрейфа . [ 14 ]
Y-хромосома, при 30% разнице между людьми и шимпанзе, является одной из наиболее быстро развивающихся частей человеческого генома . [ 27 ] Однако эти изменения ограничились некодирующими последовательностями, а сравнения Y-хромосом человека и шимпанзе (впервые опубликованные в 2005 году) показывают, что Y-хромосома человека не потеряла ни одного гена со времени расхождения людей и шимпанзе между 6–7 миллионами человек. много лет назад. [ 28 ] Кроме того, в научном отчете 2012 года говорилось, что только один ген был потерян с тех пор, как люди отделились от макаки-резус 25 миллионов лет назад. [ 29 ] Эти факты являются прямым доказательством того, что модель линейной экстраполяции ошибочна, и позволяют предположить, что нынешняя человеческая Y-хромосома либо больше не сокращается, либо сокращается гораздо медленнее, чем 4,6 генов на миллион лет, оцененные моделью линейной экстраполяции. [ нужна ссылка ]
Высокая скорость мутаций
[ редактировать ]Y-хромосома человека особенно подвержена высокой частоте мутаций из-за окружающей среды, в которой она находится. Y-хромосома передается исключительно через сперматозоиды , которые подвергаются множественным клеточным делениям во время гаметогенеза . Каждое клеточное деление предоставляет дополнительную возможность накапливать мутации пар оснований. Кроме того, сперматозоиды хранятся в высокоокислительной среде яичка , что способствует дальнейшей мутации. Сочетание этих двух условий подвергает Y-хромосому большей вероятности мутации, чем остальная часть генома. [ 14 ] Увеличение возможности мутации Y-хромосомы оценивается Грейвсом как фактор 4,8. [ 14 ] Однако в ее исходной ссылке это число получено для относительной частоты мутаций в мужских и женских зародышевых линиях линии, ведущей к человеку. [ 30 ]
Наблюдение того, что Y-хромосома подвергается незначительной мейотической рекомбинации и имеет повышенную скорость мутаций и деградационных изменений по сравнению с остальной частью генома, предполагает эволюционное объяснение адаптивной функции мейоза по отношению к основной части генетической информации. Брандейс [ 31 ] предположил, что основной функцией мейоза (особенно мейотической рекомбинации) является сохранение целостности генома, это предположение согласуется с идеей о том, что мейоз является адаптацией для восстановления повреждений ДНК . [ 32 ]
Неэффективный отбор
[ редактировать ]Без способности рекомбинировать во время мейоза Y-хромосома не может подвергать отдельные аллели естественному отбору. Вредным аллелям разрешено «путешествовать автостопом» с полезными соседями, тем самым распространяя неадаптированные аллели в следующее поколение. И наоборот, выгодные аллели могут быть отобраны, если они окружены вредными аллелями (фоновый отбор). Из-за этой неспособности сортировать содержимое своих генов Y-хромосома особенно склонна к накоплению «мусорной» ДНК . По всей территории Y разбросаны массивные скопления ретротранспозируемых элементов. [ 14 ] Случайная вставка сегментов ДНК часто нарушает закодированные последовательности генов и делает их нефункциональными. Однако Y-хромосома не способна отсеять эти «прыгающие гены». Без способности изолировать аллели отбор не может эффективно воздействовать на них. [ нужна ссылка ]
Ясным количественным показателем этой неэффективности является уровень энтропии Y-хромосомы. В то время как все остальные хромосомы в геноме человека имеют уровень энтропии 1,5–1,9 бит на нуклеотид (по сравнению с теоретическим максимумом, равным ровно 2 при отсутствии избыточности), уровень энтропии Y-хромосомы составляет всего 0,84. [ 33 ] Это означает, что Y-хромосома имеет гораздо меньшую информативность по сравнению с ее общей длиной; это более избыточно. [ нужна ссылка ]
Генетический дрейф
[ редактировать ]Даже если хорошо адаптированной Y-хромосоме удастся сохранить генетическую активность, избегая накопления мутаций, нет никакой гарантии, что она перейдет к следующему поколению. Размер популяции Y-хромосомы по своей природе ограничен 1/4 размера аутосом: диплоидные организмы содержат две копии аутосомных хромосом, в то время как только половина популяции содержит 1 Y-хромосому. Таким образом, генетический дрейф представляет собой исключительно сильную силу, воздействующую на Y-хромосому. Из-за абсолютно случайного распределения взрослый мужчина никогда не сможет передать свою Y-хромосому, если у него будет потомство только женского пола. Таким образом, хотя у самца может быть хорошо адаптированная Y-хромосома без чрезмерных мутаций, она может никогда не попасть в следующий генофонд. [ 14 ] Повторяющаяся случайная потеря хорошо адаптированных Y-хромосом в сочетании с тенденцией Y-хромосомы к эволюции с появлением более вредных мутаций, а не меньших по причинам, описанным выше, способствует общевидовой дегенерации Y-хромосом через механизм Мюллера . [ 34 ]
Конверсия генов
[ редактировать ]Как уже упоминалось, Y-хромосома не способна рекомбинировать во время мейоза, как другие хромосомы человека; однако в 2003 году исследователи из Массачусетского технологического института обнаружили процесс, который может замедлить процесс деградации. Они обнаружили, что человеческая Y-хромосома способна «рекомбинировать» сама с собой, используя палиндромные последовательности пар оснований . [ 35 ] Такая «рекомбинация» называется конверсией генов .
В случае Y-хромосом палиндромы не являются некодирующей ДНК ; эти цепочки оснований содержат функционирующие гены, важные для мужской фертильности. Большинство пар последовательностей идентичны более чем на 99,97%. Широкое использование генной конверсии может сыграть роль в способности Y-хромосомы исправлять генетические ошибки и поддерживать целостность относительно небольшого количества генов, которые она несет. Другими словами, поскольку Y-хромосома одна, она имеет дубликаты своих генов вместо второй, гомологичной хромосомы. При возникновении ошибок он может использовать другие части себя в качестве шаблона для их исправления. [ 35 ]
Результаты были подтверждены путем сравнения аналогичных участков Y-хромосомы человека с Y-хромосомами шимпанзе , бонобо и горилл . Сравнение показало, что тот же феномен конверсии генов, по-видимому, действовал более 5 миллионов лет назад, когда люди и приматы, не относящиеся к человеку, разошлись друг с другом. [ 35 ]
генной конверсии, Тракты образующиеся во время мейоза , длинные, около 2068 пар оснований, и значительно смещены в сторону фиксации нуклеотидов G или C (смещение GC). [ 36 ] Было обнаружено, что промежуточные продукты рекомбинации, предшествующие конверсии генов, редко выбирают альтернативный путь кроссоверной рекомбинации. [ 36 ] Коэффициент конверсии гена YY у человека составляет около 1,52 х 10. -5 конверсии/база/год. [ 37 ] Эти события генной конверсии могут отражать основную функцию мейоза — сохранение целостности генома.
Будущая эволюция
[ редактировать ]По некоторым теориям, на терминальных стадиях дегенерации Y-хромосомы другие хромосомы могут все больше перенимать ранее связанные с ней гены и функции и, наконец, в рамках этой теории Y-хромосома полностью исчезает и возникает новый пол. - возникает определяющая система. [ 14 ] [ нейтралитет оспаривается ] [ неправильный синтез? ]
Несколько видов грызунов сестринских семейств Muridae и Cricetidae достигли стадии, когда система XY была модифицирована. [ 38 ] [ 39 ] следующими способами:
- Закавказская слепушонка и Ellobius lutescens , зайсанская слепушонка Ellobius tancrei японские остистые деревенские крысы Tokudaia osimensis и Tokudaia tokunoshimensis утратили Y-хромосому и SRY . полностью [ 14 ] [ 40 ] [ 41 ] Токудая виды. переместили некоторые другие гены, изначально присутствовавшие на Y-хромосоме, на Х-хромосому. [ 41 ] Оба пола Tokudaia spp. и Ellobius lutescens имеют генотип ХО ( синдром Тернера ), [ 41 ] тогда как все Ellobius tancrei обладают генотипом XX. [ 14 ] Новая система(ы) определения пола этих грызунов остаётся неясной.
- Лесной лемминг Myopus schisticolor , арктический лемминг , Dicrostonyx torquatus и несколько видов травяных мышей из рода Akodon развили плодовитых самок, которые обладают генотипом, обычно кодирующим самцов, XY, в дополнение к предковой самке XX, посредством различных модификаций. к Х- и Y-хромосомам. [ 38 ] [ 42 ] [ 43 ]
- У ползучей полевки Microtus oregoni самки, имеющие только одну Х-хромосому, производят только Х-гаметы, а самцы XY производят Y-гаметы или гаметы, лишенные какой-либо половой хромосомы, посредством нерасхождения . [ 44 ]
За исключением грызунов, черный мунтжак , Muntiacus crinifrons , развил новые X- и Y-хромосомы путем слияния предковых половых хромосом и аутосом . [ 45 ]
Современные данные ставят под сомнение гипотезу об исчезновении Y-хромосомы. [ 16 ] К такому выводу пришли ученые, изучавшие Y-хромосомы макак-резус. При геномном сравнении Y-хромосомы макак-резус и человека ученые обнаружили очень мало различий, учитывая, что люди и макаки-резус разошлись 30 миллионов лет назад. [ 46 ] [ нужны разъяснения ]
За исключением млекопитающих, некоторые организмы потеряли Y-хромосому, например, большинство видов нематод. Однако для того, чтобы произошла полная элиминация Y, необходимо было разработать альтернативный способ определения пола (например, путем определения пола по соотношению Х-хромосомы и аутосом), а любые гены, необходимые для мужской функции, были переместиться в другие хромосомы. [ 16 ] Между тем, современные данные демонстрируют сложные механизмы эволюции Y-хромосомы и тот факт, что исчезновение Y-хромосомы не гарантировано.
Соотношение полов 1:1
[ редактировать ]Принцип Фишера объясняет, почему почти все виды, использующие половое размножение, имеют соотношение полов 1:1. У. Д. Гамильтон дал следующее основное объяснение в своей статье 1967 года «Чрезвычайное соотношение полов»: [ 47 ] при условии, что производство самцов и самок обходится одинаково:
- Предположим, что рождаемость мужского пола встречается реже, чем женского.
- Тогда новорожденный самец имеет лучшие шансы на спаривание, чем новорожденная самка, и поэтому может рассчитывать на большее потомство.
- Таким образом, у родителей, генетически предрасположенных к появлению на свет мужского пола, обычно рождается больше внуков, чем в среднем.
- Таким образом, гены склонности к воспроизведению мужского пола распространяются, и рождение мальчиков становится более распространенным явлением.
- По мере достижения соотношения полов 1:1 преимущество, связанное с производством самцов, исчезает.
- Те же рассуждения справедливы, если повсюду заменять мужчин женщинами. Следовательно, 1:1 является равновесным соотношением.
Нетерианская Y-хромосома
[ редактировать ]Многие группы организмов, помимо териевых млекопитающих, имеют Y-хромосомы, но эти Y-хромосомы не имеют общего происхождения с териальными Y-хромосомами. К таким группам относятся однопроходные, дрозофилы , некоторые другие насекомые, некоторые рыбы, некоторые рептилии и некоторые растения. У Drosophila melanogaster Y-хромосома не запускает развитие мужского пола. Вместо этого пол определяется количеством Х-хромосом. Y- хромосома D. melanogaster действительно содержит гены, необходимые для мужской фертильности. Таким образом, XXY D. melanogaster — самки, а D. melanogaster с единственным X (X0) — самцы, но бесплодны. Существуют виды дрозофил, у которых самцы Х0 одновременно жизнеспособны и плодовиты. [ нужна ссылка ]
ZW-хромосомы
[ редактировать ]У других организмов половые хромосомы имеют зеркальное отображение: гомогенный пол — мужской с двумя Z-хромосомами, а женский — гетерогенный пол с Z-хромосомой и W-хромосомой. [ 48 ] Например, система определения пола ZW встречается у птиц , змей и бабочек ; у женщин есть половые хромосомы ZW, а у мужчин - половые хромосомы ZZ. [ 48 ] [ 49 ] [ 50 ]
Неинвертированная Y-хромосома
[ редактировать ]Есть некоторые виды, такие как японская рисовая рыба , у которых система XY все еще развивается, и скрещивание между X и Y все еще возможно. Поскольку специфичная для самцов область очень мала и не содержит существенных генов, можно даже искусственно индуцировать XX самцов и YY самок без каких-либо вредных последствий. [ 51 ]
Несколько пар XY
[ редактировать ]Однопроходные, такие как утконосы, обладают четырьмя или пятью парами половых хромосом XY, каждая пара состоит из половых хромосом с гомологичными участками. Хромосомы соседних пар частично гомологичны, так что во время митоза образуется цепочка . [ 19 ] Первая Х-хромосома в цепи также частично гомологична последней Y-хромосоме, что указывает на то, что в истории происходили глубокие перестройки, в том числе с добавлением новых фрагментов из аутосом. [ 52 ] [ 53 ] : инжир. 5
Половые хромосомы утконоса имеют сильное сходство последовательностей с Z-хромосомой птиц (что указывает на близкую гомологию ). [ 17 ] а ген SRY, столь важный для определения пола у большинства других млекопитающих, по-видимому, не участвует в определении пола у утконоса. [ 18 ]
Y-хромосома человека
[ редактировать ]Этот раздел может потребовать очистки Википедии , чтобы соответствовать стандартам качества . Конкретная проблема: слишком много подразделов. Статья могла бы выиграть от перемещения подразделов h3 в разделы h2, если мы сможем каким-то образом устранить разрыв между всеми терианцами и людьми. Раздел «Происхождение и эволюция» ориентирован на человека, но в обсуждение включены все терианцы. ( Октябрь 2021 г. ) |
Y-хромосома человека состоит примерно из 62 миллионов пар оснований ДНК , что делает ее похожей по размеру на хромосому 19 и составляет почти 2% от общего количества ДНК в мужской клетке . [ 54 ] [ 55 ] Y-хромосома человека несет 693 гена , 107 из которых кодируют белки . [ 56 ] Однако некоторые гены повторяются, в результате чего число эксклюзивных генов , кодирующих белок, составляет всего 42. [ 56 ] Проект консенсусной кодирующей последовательности (CCDS) классифицирует только 63 из 107 генов, хотя оценки CCDS часто считаются нижними границами из-за их консервативной стратегии классификации. [ 57 ] Все однокопийные Y-сцепленные гены гемизиготны (присутствуют только на одной хромосоме), за исключением случаев анеуплоидии, таких как синдром XYY или синдром XXYY . Признаки, которые наследуются через Y-хромосому, называются Y-сцепленными признаками, или голандрическими признаками (от древнегреческого ὅλος hólos , «целый» + ἀνδρός andrós , «мужской»). [ 58 ]
Последовательность Y-хромосомы человека
[ редактировать ]По окончании проекта «Геном человека» (и после многих обновлений) почти половина Y-хромосомы осталась несеквенированной даже в 2021 году; Y-хромосома, отличная от генома HG002 (GM24385), была полностью секвенирована в январе 2022 года и включена в новую «полную геномную» последовательность эталонного генома человека , CHM13. [ 56 ] Было показано, что полное секвенирование Y-хромосомы человека содержит 62 460 029 пар оснований и 41 дополнительный ген . [ 56 ] Это добавило 30 миллионов пар оснований, [ 56 ] но было обнаружено, что Y-хромосома может сильно различаться по размеру у разных людей: от 45,2 миллиона до 84,9 миллиона пар оснований. [ 59 ]
Поскольку почти половина человеческой Y-последовательности была неизвестна до 2022 года, ее нельзя было выявить как загрязнение в проектах микробного секвенирования. В результате база данных бактериального генома NCBI RefSeq ошибочно включает некоторые данные Y-хромосомы. [ 56 ]
Структура
[ редактировать ]В этой статье отсутствует информация о структуре NRY/MSY — о том, как в q имеется огромный кусок гетерохроматина , номенклатура палиндромов и ампликонов , транскрипты TTTY и т. д. Лучше всего, если мы добавим фигуру, объединяющую верхушки Colaco 2018, рис. 1 и PMID 12815422 рис. 3. ( октябрь 2021 г. ) |
Цитогенетическая полоса
[ редактировать ]
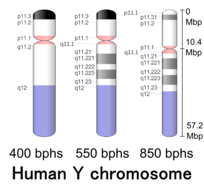
| Хр. | Рука [ 64 ] | Группа [ 65 ] | ISCN начинать [ 66 ] |
ISCN останавливаться [ 66 ] |
Базовая пара начинать |
Базовая пара останавливаться |
Пятно [ 67 ] | Плотность |
|---|---|---|---|---|---|---|---|---|
| И | п | 11.32 | 0 | 149 | 1 | 300,000 | хороший | |
| И | п | 11.31 | 149 | 298 | 300,001 | 600,000 | GPS | 50 |
| И | п | 11.2 | 298 | 1043 | 600,001 | 10,300,000 | хороший | |
| И | п | 11.1 | 1043 | 1117 | 10,300,001 | 10,400,000 | акцент | |
| И | д | 11.1 | 1117 | 1266 | 10,400,001 | 10,600,000 | акцент | |
| И | д | 11.21 | 1266 | 1397 | 10,600,001 | 12,400,000 | хороший | |
| И | д | 11.221 | 1397 | 1713 | 12,400,001 | 17,100,000 | GPS | 50 |
| И | д | 11.222 | 1713 | 1881 | 17,100,001 | 19,600,000 | хороший | |
| И | д | 11.223 | 1881 | 2160 | 19,600,001 | 23,800,000 | GPS | 50 |
| И | д | 11.23 | 2160 | 2346 | 23,800,001 | 26,600,000 | хороший | |
| И | д | 12 | 2346 | 3650 | 26,600,001 | 57,227,415 | гвар |
Необъединяющая область Y (NRY)
[ редактировать ]Y-хромосома человека обычно не способна рекомбинировать с X-хромосомой, за исключением небольших участков псевдоаутосомных областей (PAR) на теломерах (которые составляют около 5% длины хромосомы). Эти области являются остатками древней гомологии между X и Y-хромосомами. Основная часть Y-хромосомы, которая не рекомбинирует, называется «NRY» или нерекомбинирующей областью Y-хромосомы. [ 68 ] Однонуклеотидные полиморфизмы (SNP) в этом регионе используются для отслеживания прямых отцовских наследственных линий.
Точнее, PAR1 составляет 0,1–2,7 МБ. PAR2 составляет 56,9–57,2 МБ. Нерекомбинирующая область (NRY) или мужская область (MSY) находится между ними. Их размеры теперь прекрасно известны из CHM13: 2,77 Мб и 329,5 Кб. До CHM13 данные в PAR1 и PAR2 просто копировались из X-хромосомы. [ 59 ]
Классы последовательностей
[ редактировать ]Гены
[ редактировать ]Количество генов
[ редактировать ]Ниже приведены некоторые оценки количества генов Y-хромосомы человека. Поскольку исследователи используют разные подходы к аннотированию генома, их прогнозы количества генов в каждой хромосоме различаются (технические подробности см. в разделе «Прогнозирование генов» ). Среди различных проектов CCDS придерживается крайне консервативной стратегии. Таким образом, прогноз числа генов CCDS представляет собой нижнюю границу общего количества генов, кодирующих человеческие белки. [ 69 ]
| По оценкам | Гены, кодирующие белки | Некодирующие гены РНК | Псевдогены | Источник | Дата выпуска |
|---|---|---|---|---|---|
| CCDS | 63 | — | — | [ 1 ] | 2016-09-08 |
| HGNC | 45 | 55 | 381 | [ 70 ] | 2017-05-12 |
| Вместе | 63 | 109 | 392 | [ 71 ] | 2017-03-29 |
| ЮниПрот | 47 | — | — | [ 72 ] | 2018-02-28 |
| NCBI | 73 | 122 | 400 | [ 73 ] [ 74 ] [ 75 ] | 2017-05-19 |
Список генов
[ редактировать ]В целом Y-хромосома человека чрезвычайно бедна генами — это одна из крупнейших генных пустынь в геноме человека. Если не учитывать псевдоаутосомные гены, гены, кодируемые Y-хромосомой человека, включают:
| Имя | X- паралог | Примечание |
|---|---|---|
| СРЮ | SOX3 | Пол-определяющая область. Это пара [Yp]. |
| ЗФЮ | ZFX | Цинковый палец . |
| РПС4Y1 | РПС4Х | Рибосомальный белок S4. |
| КОТОРЫЙ | АМЕЛКС | Амелогенин . |
| ТБЛ1Y | ТБЛ1Х | |
| PCDH11Y | PDCH11X | X-транспонированная область (XTR) из Xq21, одного из двух генов. Когда-то получивший название «PAR3». [ 77 ] но позже опровергнуто. [ 78 ] |
| TGIF2LY | TGIF2LX | Другой X-транспонированный ген. |
| ЦПY1 , ЦПY2 | ТСПХ | Белок, специфичный для семенников. |
| АЗФа | (никто) | Не ген. Первая часть региона AZF (фактор азооспермии) на руке q. Содержит четыре следующих гена. X-двойники избегают инактивации. |
| USP9Y | USP9X | Убиквитиновая протеаза. |
| DDX3Y | DDX3X | Хеликаза. |
| УТЙ | ЮТХ | Гистондеметилаза. |
| ТБ4Y | ТБ4X | |
| АЗФб | (никто) | Второй регион AZF на рукаве q. Склонен к NAHR (неаллельной гомологичной рекомбинации) с AZFc. Пересекается с AZFc. Содержит три однокопийных участка гена и повторы. |
| CYorf15 | CXorf15 | |
| РПС4Y2 | РПС4Х | Еще одна копия рибосомного белка S4. |
| EIF1AY | EIF4AX | |
| КДМ5Д | КДМ5С | |
| XKRY | ХК (белок) | Найден в «желтом» ампликоне . |
| ХСФГ1 , ХСФГ2 | ХСFX1 , ХСFX2 | Найден в «голубом» ампликоне. |
| PRY , PRY2 | Найден в «голубом» ампликоне. Идентифицируется по сходству с PTPN13 (гл. 4). | |
| РБМИ1А1 | РБМИ | Большое количество копий. Часть семейства генов RBM белков мотива распознавания РНК (RRM). |
| АЗФК | (никто) | Конечная (дистальная) часть АЗФ. Множественные палиндромы. |
| ДАЗ1 , ДАЗ2 , ДАЗ3 , ДАЗ4 | Гены RRM в двух палиндромных кластерах. BOLL и DAZLA являются аутосомными гомологами. | |
| CDY1 , CDY2 | CDY1 на самом деле представляет собой две идентичные копии. CDY2 — это две близкородственные копии в палиндроме P5. Вероятно, происходит из аутосомного CDYL . | |
| ВЦИ1 , ВЦИ2 | VCX1–3 | Три копии VCX2 (BPY2). Входит в семейство VCX/VCY . Вместо этого две копии BPY1 находятся в Yq11.221/AZFa. |
Заболевания, сцепленные с Y-хромосомой
[ редактировать ]Заболевания, связанные с Y-хромосомой, обычно включают анеуплоидию — атипичное количество хромосом.
Потеря Y-хромосомы
[ редактировать ]Мужчины могут потерять Y-хромосому в подмножестве клеток, это называется мозаичной потерей. Утрата мозаики тесно связана с возрастом, [ 79 ] и курение является еще одним важным фактором риска мозаичной потери. [ 80 ]
Потеря мозаики может быть связана с последствиями для здоровья, указывая на то, что Y-хромосома играет важную роль помимо определения пола. [ 80 ] [ 81 ] Мужчины с более высоким процентом гемопоэтических стволовых клеток, лишенных Y-хромосомы, имеют более высокий риск развития некоторых видов рака и имеют меньшую продолжительность жизни. [ 81 ] Во многих случаях причинно-следственная связь между Y-хромосомой и последствиями для здоровья не определена, а некоторые предполагают, что потеря Y-хромосомы может быть «нейтральным кариотипом, связанным с нормальным старением ». [ 82 ] Однако исследование 2022 года показало, что мозаичная потеря Y-хромосомы причинно способствует фиброзу , сердечно-сосудистым рискам и смертности. [ 83 ]
Необходимы дальнейшие исследования, чтобы понять, как мозаичная потеря Y-хромосомы может способствовать другим половым различиям в состоянии здоровья, например, почему у курильщиков-мужчин в 1,5–2 раза выше риск развития нереспираторного рака, чем у курящих женщин. [ 84 ] [ 85 ] Потенциальные контрмеры, выявленные на данный момент, включают отказ от курения или прекращение курения , и по крайней мере один потенциальный препарат, который «может помочь противодействовать вредным последствиям потери хромосом», находится в стадии расследования. [ 86 ] [ 87 ] [ нужен лучший источник ]
Микроделеция Y-хромосомы
[ редактировать ]Микроделеция Y-хромосомы (YCM) — это семейство генетических нарушений, вызванных отсутствием генов в Y-хромосоме. Многие больные мужчины не проявляют никаких симптомов и ведут нормальную жизнь. Однако известно, что YCM присутствует у значительного числа мужчин со сниженной фертильностью или уменьшенным количеством сперматозоидов. [ нужна ссылка ]
Дефектная Y-хромосома
[ редактировать ]Это приводит к тому, что человек имеет женский фенотип (т. е. рождается с женскими гениталиями), даже если этот человек обладает кариотипом XY . Отсутствие второго Х приводит к бесплодию. Другими словами, если смотреть с противоположной стороны, человек проходит дефеминизацию , но не может завершить маскулинизацию . [ нужна ссылка ]
Причиной может быть неполная Y-хромосома: обычный кариотип в этих случаях - 45X плюс фрагмент Y. Обычно это приводит к дефектному развитию яичек, так что у младенца могут быть или не быть полностью сформированные мужские гениталии внутри или снаружи. . Может возникнуть весь спектр неоднозначности структуры, особенно если мозаицизм присутствует . Когда Y-фрагмент минимален и нефункционален, ребенок обычно является девочкой с признаками синдрома Тернера или смешанной дисгенезии гонад . [ нужна ссылка ]
ХХ
[ редактировать ]Синдром Клайнфельтера (47, XXY) – это не анеуплоидия Y -хромосомы, а состояние наличия дополнительной Х-хромосомы, что обычно приводит к нарушению постнатальной функции яичек. Механизм не до конца понятен; по-видимому, это не связано с прямым вмешательством дополнительного X в экспрессию генов Y. [ нужна ссылка ]
XYY
[ редактировать ]47, синдром XYY (просто известный как синдром XYY) вызван наличием одной дополнительной копии Y-хромосомы в каждой мужской клетке. 47, самцы XYY имеют одну Х-хромосому и две Y-хромосомы, всего 47 хромосом на клетку. Исследователи обнаружили, что дополнительная копия Y-хромосомы связана с увеличением роста и увеличением частоты проблем с обучением у некоторых мальчиков и мужчин, но эффекты варьируются, часто минимальны, и подавляющее большинство не знает своего кариотипа. [ 88 ]
В 1965 и 1966 годах Патрисия Джейкобс и ее коллеги опубликовали исследование хромосом 315 пациентов мужского пола в клинике. Шотландии Единственная в больница специального режима для людей с отклонениями в развитии , обнаружение большего, чем ожидалось, количества пациентов с дополнительной Y-хромосомой. [ 89 ] Авторы этого исследования задались вопросом, «предрасполагает ли лишняя Y-хромосома своих носителей к необычно агрессивному поведению», и эта гипотеза «положила начало следующим пятнадцати годам исследований Y-хромосомы человека». [ 90 ]
Исследования, проведенные в течение следующего десятилетия, показали, что эта гипотеза неверна: повышенный уровень преступности среди мужчин XYY обусловлен более низким средним интеллектом, а не повышенной агрессией. [ 91 ] а увеличенный рост был единственной характеристикой, которую можно было достоверно связать с самцами XYY. [ 92 ] Таким образом, концепция «криминального кариотипа» неверна. [ 88 ]
Существуют также синдром XXXY и синдром XXXXY .
Редкий
[ редактировать ]Следующие заболевания, сцепленные с Y-хромосомой, редки, но примечательны тем, что они проливают свет на природу Y-хромосомы.
Более двух Y-хромосом
[ редактировать ]Полисомия Y-хромосомы большей степени (наличие более одной дополнительной копии Y-хромосомы в каждой клетке, например XYYY) встречается значительно реже. Дополнительный генетический материал в этих случаях может привести к аномалиям скелета, зубным аномалиям, снижению IQ, задержке развития и респираторным проблемам, но степень тяжести этих состояний варьируется. [ 93 ]
XX мужской синдром
[ редактировать ]Мужской синдром ХХ возникает из-за генетической рекомбинации при формировании мужских гамет , в результате чего часть SRY Y-хромосомы перемещается в Х-хромосому. [ 8 ] Когда такая Х-хромосома присутствует в зиготе, мужские гонады развиваются благодаря гену SRY. [ 8 ]
Генетическая генеалогия
[ редактировать ]В генетической генеалогии человека (применении генетики к традиционной генеалогии ) использование информации, содержащейся в Y-хромосоме, представляет особый интерес, поскольку, в отличие от других хромосом, Y-хромосома передается исключительно от отца к сыну, по отцовской линии. Митохондриальная ДНК , наследуемая по материнской линии как сыновьям, так и дочерям, используется аналогичным образом для отслеживания материнской линии. [ нужна ссылка ]
Функция мозга
[ редактировать ]В настоящее время исследования выясняют, является ли развитие нейронов по мужскому типу прямым следствием экспрессии генов, связанных с Y-хромосомой, или косвенным результатом выработки андрогенных гормонов , связанных с Y-хромосомой . [ 94 ]
Микрохимеризм
[ редактировать ]В 1974 году мужские хромосомы были обнаружены в клетках плода в кровообращении женщин. [ 95 ]
В 1996 году было обнаружено, что клетки-предшественники плода мужского пола могут сохраняться после родов в кровотоке матери до 27 лет. [ 96 ]
Исследование, проведенное в 2004 году в Онкологическом исследовательском центре Фреда Хатчинсона в Сиэтле, изучало происхождение мужских хромосом, обнаруженных в периферической крови женщин, у которых не было потомства мужского пола. Всего было обследовано 120 субъектов (женщин, у которых никогда не было сыновей), и было обнаружено, что у 21% из них в периферической крови была мужская ДНК. Субъекты были разделены на четыре группы на основе их историй болезни: [ 97 ]
- Группа А (8%) имела потомство только женского пола.
- У пациенток группы Б (22%) в анамнезе был один или несколько выкидышей.
- Пациентки группы С (57%) прервали беременность медикаментозными методами.
- Группа D (10%) никогда раньше не была беременна.
Исследование отметило, что 10% женщин никогда раньше не были беременны, что поднимает вопрос о том, откуда могли взяться Y-хромосомы в их крови. Исследование предполагает, что возможными причинами возникновения микрохимеризма мужских хромосом могут быть одна из следующих: [ 97 ]
- выкидыши,
- беременность,
- исчезнувший близнец мужского пола,
- возможно, от полового акта.
Исследование, проведенное в 2012 году в том же институте, обнаружило клетки с Y-хромосомой во многих областях мозга умерших женщин. [ 98 ]
См. также
[ редактировать ]- Генеалогический ДНК-тест
- Генетическая генеалогия
- Гаплодиплоидная система определения пола
- Гаплогруппы ДНК Y-хромосомы человека
- Список маркеров Y-STR
- трещотка Мюллера
- Однонуклеотидный полиморфизм
- Короткие тандемные повторы Y-хромосомы (STR)
- Y-образная связь
- Y-хромосома Аарон
- Y-хромосомный Адам
- Гаплогруппы Y-хромосомы в популяциях мира
Ссылки
[ редактировать ]- ^ Jump up to: а б «Гены Y-хромосомы человека разумного» . CCDS Release 20 для Homo Sapiens . 08.09.2016 . Проверено 28 мая 2017 г.
- ^ Страчан Т., Читать A (2 апреля 2010 г.). Молекулярная генетика человека . Гирляндная наука. п. 45. ИСБН 978-1-136-84407-2 .
- ^ Jump up to: а б с «Данные идеограммы Homo sapience (850 ударов в час, сборка GRCh38.p3)» . Страница оформления генома . Национальный центр биотехнологической информации США (NCBI). 03.06.2014 . Проверено 26 апреля 2017 г.
- ^ Стекло Б (1990). «Художник Теофил Шикель: 22 августа 1889 г. - 5 октября 1969 г.» (PDF) . Биографические мемуары Национальной академии наук . 59 : 309–37. ПМИД 11616163 .
- ^ Jump up to: а б Бейнбридж Д. (2003). X в сексе: как контролируется X-хромосома . Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-01621-7 .
- ^ Шварц Дж. (2009). В поисках гена: от Дарвина к ДНК . Кембридж, Массачусетс: Издательство Гарвардского университета. стр. 170–2. ISBN 978-0-674-03491-4 .
- ^ Jump up to: а б «Синдром Свайера — симптомы, причины, лечение | NORD» . Rarediseases.org . Проверено 12 сентября 2023 г.
- ^ Jump up to: а б с «ХХ Мужской синдром — обзор» . Наука Директ . 12 сентября 2023 г.
Домингес А.А., Рейхо Пера Р.А. (1 января 2013 г.). «Бесплодие» . В Малой С., Хьюз К. (ред.). Энциклопедия генетики Бреннера (2-е изд.). Академическая пресса. стр. 71–74. дои : 10.1016/B978-0-12-374984-0.00793-2 . ISBN 978-0-12-374984-0 . Проверено 12 сентября 2023 г. - ^ Девлин Р.Х., Нагахама Ю (21 июня 2002 г.). «Определение пола и дифференциация пола у рыб: обзор генетических, физиологических и экологических влияний» . Аквакультура . Определение пола и половая дифференциация у рыб. 208 (3): 191–364. Бибкод : 2002Aquac.208..191D . дои : 10.1016/S0044-8486(02)00057-1 . ISSN 0044-8486 .
- ^ Барреси М.Дж., Гилберт С.Ф. (2023). «6. Определение пола и гаметогенез §6.4 Определение пола у рептилий в окружающей среде» . Биология развития (13-е изд.). Издательство Оксфордского университета. ISBN 978-0-19-757459-1 .
Гилберт С.Ф. (2000). «Определение пола в условиях окружающей среды» . Биология развития (6-е изд.). Синауэр Ассошиэйтс. п. 60. ИСБН 0-87893-243-7 . НБК9989. - ^ Кувамура Т., Сунобе Т., Сакаи Ю., Кадота Т., Савада К. (01.07.2020). «Гермафродитизм у рыб: аннотированный список видов, филогения и система спаривания» . Ихтиологические исследования . 67 (3): 341–360. Бибкод : 2020IchtR..67..341K . дои : 10.1007/s10228-020-00754-6 . ISSN 1616-3915 . S2CID 254168012 .
- ^ Мюллер HJ (1914). «Ген четвертой хромосомы дрозофилы» . Журнал экспериментальной зоологии . 17 (3): 325–336. Бибкод : 1914JEZ....17..325M . дои : 10.1002/jez.1400170303 .
- ^ Лан Б.Т., Пейдж, округ Колумбия (октябрь 1999 г.). «Четыре эволюционных слоя Х-хромосомы человека». Наука . 286 (5441): 964–7. дои : 10.1126/science.286.5441.964 . ПМИД 10542153 .
- ^ Jump up to: а б с д и ж г час я дж к Грейвс Дж. А. (март 2006 г.). «Специализация и дегенерация половых хромосом у млекопитающих» . Клетка . 124 (5): 901–914. дои : 10.1016/j.cell.2006.02.024 . ПМИД 16530039 . S2CID 8379688 .
- ^ Грейвс Дж. А., Койна Э., Санкович Н. (июнь 2006 г.). «Как развивался генный состав половых хромосом человека». Текущее мнение в области генетики и развития . 16 (3): 219–224. дои : 10.1016/j.где.2006.04.007 . ПМИД 16650758 .
- ^ Jump up to: а б с Бахтрог Д. (февраль 2013 г.). «Эволюция Y-хромосомы: новое понимание процессов дегенерации Y-хромосомы» . Обзоры природы. Генетика . 14 (2): 113–124. дои : 10.1038/nrg3366 . ПМК 4120474 . ПМИД 23329112 .
- ^ Jump up to: а б Уоррен В.К., Хиллиер Л.В., Маршалл Грейвс Дж.А., Бирни Э., Понтинг К.П., Грюцнер Ф. и др. (май 2008 г.). «Анализ генома утконоса выявил уникальные признаки эволюции» . Природа . 453 (7192): 175–183. Бибкод : 2008Natur.453..175W . дои : 10.1038/nature06936 . ПМК 2803040 . ПМИД 18464734 .
- ^ Jump up to: а б с Вейрунес Ф., Уотерс П.Д., Митке П., Ренс В., Макмиллан Д., Олсоп А.Е. и др. (июнь 2008 г.). «Птичьи половые хромосомы утконоса предполагают недавнее происхождение половых хромосом млекопитающих» . Геномные исследования . 18 (6): 965–973. дои : 10.1101/гр.7101908 . ПМК 2413164 . ПМИД 18463302 .
- ^ Jump up to: а б Грюцнер Ф., Ренс В., Ценд-Аюш Э., Эль-Могарбель Н., О'Брайен ПК, Джонс Р.К. и др. (декабрь 2004 г.). «У утконоса мейотическая цепь из десяти половых хромосом разделяет гены с Z-хромосомами птиц и X-хромосомами млекопитающих». Природа . 432 (7019): 913–7. Бибкод : 2004Natur.432..913G . дои : 10.1038/nature03021 . ПМИД 15502814 . S2CID 4379897 .
- ^ Уотсон Дж. М., Риггс А., Грейвс Дж. А. (октябрь 1992 г.). «Исследования по картированию генов подтверждают гомологию между хромосомами X утконоса и X1 ехидны и идентифицируют консервативную предковую однопроходную X-хромосому». Хромосома . 101 (10): 596–601. дои : 10.1007/BF00360536 . ПМИД 1424984 . S2CID 26978106 .
- ^ Jump up to: а б Лемье Дж. (29 мая 2020 г.). «О PAR: как X- и Y-хромосомы рекомбинируют во время мейоза» . GEN — Новости генной инженерии и биотехнологии . Проверено 13 ноября 2023 г.
- ^ Пенедер П., Валлнер Б., Фогль С. (октябрь 2017 г.). «Обмен генетической информацией между терианскими гаметологами X и Y-хромосом в старых эволюционных слоях» . Экология и эволюция . 7 (20): 8478–8487. Бибкод : 2017EcoEv...7.8478P . дои : 10.1002/ece3.3278 . ПМЦ 5648654 . ПМИД 29075464 .
- ^ Дханоа Дж.К., Мукхопадьяй К.С., Арора Дж.С. (июль 2016 г.). «Гены Y-хромосомы, влияющие на мужскую фертильность: обзор» . Ветеринарный мир . 9 (7): 783–91. дои : 10.14202/vetworld.2016.783-791 . ПМЦ 4983133 . ПМИД 27536043 .
- ^ Брукфилд Дж. Ф. (октябрь 1995 г.). «Эволюция человека. Y-хромосома является ключом к происхождению человека» . Современная биология . 5 (10): 1114–1115. Бибкод : 1995CBio....5.1114B . дои : 10.1016/S0960-9822(95)00224-7 . ПМИД 8548280 . S2CID 16081591 .
- ^ Грейвс Дж. А. (2004). «Выродившаяся Y-хромосома – может ли конверсия спасти ее?». Воспроизводство, рождаемость и развитие . 16 (5): 527–534. дои : 10.1071/RD03096 . ПМИД 15367368 .
- ^ Гото Х., Пэн Л., Макова К.Д. (февраль 2009 г.). «Эволюция генов X-вырожденной Y-хромосомы у высших приматов: сохранение содержания генов у человека и гориллы, но не у шимпанзе». Журнал молекулярной эволюции . 68 (2): 134–144. Бибкод : 2009JMolE..68..134G . дои : 10.1007/s00239-008-9189-y . ПМИД 19142680 . S2CID 24010421 .
- ^ Уэйд Н. (13 января 2010 г.). «Мужская хромосома может эволюционировать быстрее» . Нью-Йорк Таймс .
- ^ Хьюз Дж. Ф., Скалецкий Х., Пинтикова Т., Минкс П. Дж., Грейвс Т., Розен С. и др. (сентябрь 2005 г.). «Консервация Y-связанных генов в ходе эволюции человека выявлена путем сравнительного секвенирования у шимпанзе». Природа . 437 (7055): 100–103. Бибкод : 2005Natur.437..100H . дои : 10.1038/nature04101 . ПМИД 16136134 . S2CID 4418662 .
- ^ Сюй К. «Биологи опровергают теорию «гниения» Y-хромосомы: мужчины все еще будут существовать» . Медицинская газета. Архивировано из оригинала 25 февраля 2012 г. Проверено 23 февраля 2012 г.
- ^ Линдблад-То К., Уэйд С.М., Миккельсен Т.С., Карлссон Е.К., Яффе Д.Б., Камаль М. и др. (декабрь 2005 г.). «Последовательность генома, сравнительный анализ и структура гаплотипов домашней собаки» . Природа . 438 (7069): 803–819. Бибкод : 2005Natur.438..803L . дои : 10.1038/nature04338 . ПМИД 16341006 .
- ^ Брандейс М. (май 2018 г.). «Новые идеи о древнем сексе: отделение мейоза от спаривания может решить загадку вековой давности». Биологические обзоры Кембриджского философского общества . 93 (2): 801–810. дои : 10.1111/brv.12367 . ПМИД 28913952 . S2CID 4764175 .
- ^ Бернштейн Х., Хопф Ф.А., Мишод Р.Э. (1987). «Молекулярные основы эволюции пола». Молекулярная генетика развития . Достижения генетики. Том. 24. С. 323–70. дои : 10.1016/S0065-2660(08)60012-7 . ISBN 9780120176243 . ПМИД 3324702 .
- ^ Лю З, Венкатеш СС, Малей СС (октябрь 2008 г.). «Охват пространства последовательностей, энтропия геномов и возможность обнаружения нечеловеческой ДНК в образцах человека» . БМК Геномика . 9 (1): 509. дои : 10.1186/1471-2164-9-509 . ПМЦ 2628393 . ПМИД 18973670 . Рис. 6 с использованием Лемпеля-Зива . оценок скорости энтропии
- ^ Чарльзуорт Б., Чарльзуорт Д. (ноябрь 2000 г.). «Дегенерация Y-хромосомы» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 355 (1403): 1563–1572. дои : 10.1098/rstb.2000.0717 . ПМК 1692900 . ПМИД 11127901 .
- ^ Jump up to: а б с Розен С., Скалецкий Х., Маршалек Дж.Д., Минкс П.Дж., Кордум Х.С., Уотерстон Р.Х. и др. (июнь 2003 г.). «Обильная конверсия генов между плечами палиндромов в Y-хромосомах человека и обезьяны». Природа . 423 (6942): 873–876. Бибкод : 2003Natur.423..873R . дои : 10.1038/nature01723 . ПМИД 12815433 . S2CID 4323263 .
- ^ Jump up to: а б Халласт П., Балареск П., Боуден Г.Р., Баллеро С., Джоблинг М.А. (2013). «Динамика рекомбинации палиндрома Y-хромосомы человека: быстрая конверсия генов, основанная на GC, многокилобазные конверсионные пути и редкие инверсии» . ПЛОС Генет . 9 (7): e1003666. дои : 10.1371/journal.pgen.1003666 . ПМЦ 3723533 . ПМИД 23935520 .
- ^ Бонито М, Д'Атанасио Э, Равасини Ф, Кариати С, Финоккио А, Новеллетто А, Тромбетта Б, Кручиани Ф (ноябрь 2021 г.). «Новое понимание эволюции палиндромов Y-хромосомы человека посредством мутаций и конверсии генов» . Хум Мол Жене 30 (23): 2272–2285. дои : 10.1093/hmg/ddab189 . ПМК 8600007 . ПМИД 34244762 .
- ^ Jump up to: а б Маршал Х.А., Акоста М.Дж., Бульехос М., Диас де ла Гуардиа Р., Санчес А. (2003). «Половые хромосомы, определение пола и сцепленные с полом последовательности у Microtidae». Цитогенетические и геномные исследования . 101 (3–4): 266–273. дои : 10.1159/000074347 . ПМИД 14684993 . S2CID 10526522 .
- ^ Уилсон М.А., Макова К.Д. (2009). «Геномный анализ эволюции половых хромосом». Ежегодный обзор геномики и генетики человека . 10 (1): 333–354. doi : 10.1146/annurev-genom-082908-150105 . ПМИД 19630566 .
- ^ Джаст В., Баумстарк А., Зюсс А., Графодатский А., Ренс В., Шефер Н. и др. (2007). «Ellobius lutescens: определение пола и половая хромосома». Сексуальное развитие . 1 (4): 211–221. дои : 10.1159/000104771 . ПМИД 18391532 . S2CID 25939138 .
- ^ Jump up to: а б с Аракава Ю., Нисида-Умехара С., Мацуда Ю., Суто С., Судзуки Х. (2002). «Х-хромосомная локализация Y-связанных генов млекопитающих у двух видов XO колючей крысы Рюкю». Цитогенетические и геномные исследования . 99 (1–4): 303–9. дои : 10.1159/000071608 . ПМИД 12900579 . S2CID 39633026 .
- ^ Хоекстра Х.Э., Эдвардс С.В. (сентябрь 2000 г.). «Множественное происхождение самок мышей XY (род Akodon): филогенетические и хромосомные данные» . Слушания. Биологические науки . 267 (1455): 1825–31. дои : 10.1098/rspb.2000.1217 . ПМК 1690748 . ПМИД 11052532 .
- ^ Ортис М.И., Пинна-Сенн Э., Далмассо Дж., Лисанти Дж.А. (2009). «Хромосомные аспекты и наследование женского состояния XY у Akodon azarae (Rodentia, Sigmodontinae)». Биология млекопитающих . 74 (2): 125–9. Бибкод : 2009MamBi..74..125O . дои : 10.1016/j.mambio.2008.03.001 .
- ^ Чарльзуорт Б., Демпси, Северная Дакота (апрель 2001 г.). «Модель эволюции необычной системы половых хромосом Microtus oregoni» . Наследственность . 86 (Часть 4): 387–394. дои : 10.1046/j.1365-2540.2001.00803.x . ПМИД 11520338 . S2CID 34489270 .
- ^ Чжоу Ц, Ван Дж, Хуан Л, Не В, Ван Дж, Лю Ю и др. (2008). «Неополовые хромосомы черного мунтжака повторяют зарождающуюся эволюцию половых хромосом млекопитающих» . Геномная биология . 9 (6): 98 рандов. дои : 10.1186/gb-2008-9-6-r98 . ПМЦ 2481430 . ПМИД 18554412 .
- ^ Хьюз Дж. Ф., Скалецкий Х., Пейдж DC (декабрь 2012 г.). «Секвенирование Y-хромосомы макака-резуса проясняет происхождение и эволюцию генов DAZ (удаленных при АЗооспермии)» . Биоэссе . 34 (12): 1035–44. doi : 10.1002/bies.201200066 . ПМЦ 3581811 . ПМИД 23055411 .
- ^ Гамильтон У.Д. (апрель 1967 г.). «Необычайное соотношение полов. Теория соотношения полов для связи полов и инбридинга имеет новые последствия в цитогенетике и энтомологии». Наука . 156 (3774): 477–488. Бибкод : 1967Sci...156..477H . дои : 10.1126/science.156.3774.477 . ПМИД 6021675 .
- ^ Jump up to: а б Смит Дж. Дж., Восс С. Р. (сентябрь 2007 г.). «Ортологи половых хромосом птиц и млекопитающих соответствуют одной и той же аутосомной области у саламандры (амбистомы)» . Генетика . 177 (1): 607–613. doi : 10.1534/genetics.107.072033 . ПМК 2013703 . ПМИД 17660573 .
- ^ Виана П.Ф., Эзаз Т., де Белло Чоффи М., Лиер Т., Аль-Рикаби А., Голл Л.Г. и др. (июль 2020 г.). «Ландшафт эволюции половых хромосом змей, охватывающий 85 миллионов лет назад, обнаруживает происхождение последовательностей, несмотря на различные эволюционные траектории» . Научные отчеты . 10 (1): 12499. Бибкод : 2020НатСР..1012499В . дои : 10.1038/s41598-020-69349-5 . ПМЦ 7385105 . ПМИД 32719365 .
- ^ Сахара К., Ёсидо А., Траут В. (январь 2012 г.). «Эволюция половых хромосом у мотыльков и бабочек» . Хромосомные исследования . 20 (1): 83–94. дои : 10.1007/s10577-011-9262-z . hdl : 2115/49121 . ПМИД 22187366 . S2CID 15130561 .
- ^ Шартл М. (июль 2004 г.). «Сравнительный взгляд на определение пола в медаке». Механизмы развития . 121 (7–8): 639–645. дои : 10.1016/j.mod.2004.03.001 . ПМИД 15210173 . S2CID 17401686 .
- ^ Кортес Д., Марин Р., Толедо-Флорес Д., Фруадево Л., Лихти А., Уотерс П.Д. и др. (апрель 2014 г.). «Происхождение и функциональная эволюция Y-хромосом у млекопитающих». Природа . 508 (7497): 488–493. Бибкод : 2014Natur.508..488C . дои : 10.1038/nature13151 . ПМИД 24759410 . S2CID 4462870 .
- ^ Дикин Дж. Э., Грейвс Дж. А., Ренс В. (2012). «Эволюция сумчатых и однопроходных хромосом» . Цитогенетические и геномные исследования . 137 (2–4): 113–129. дои : 10.1159/000339433 . hdl : 1885/64794 . ПМИД 22777195 .
- ^ «Ensembl Human MapView, выпуск 43» . Февраль 2014 года . Проверено 14 апреля 2007 г.
- ^ «Домашний генетический справочник Национальной медицинской библиотеки» . Архивировано из оригинала 29 марта 2012 г. Проверено 9 ноября 2009 г.
- ^ Jump up to: а б с д и ж Ри А., Нурк С., Чехова М., Хойт С.Дж., Тейлор Дж., Альтемос Н. и др. (сентябрь 2023 г.). «Полная последовательность Y-хромосомы человека» . Природа . 621 (7978): 344–354. Бибкод : 2023Natur.621..344R . дои : 10.1038/s41586-023-06457-y . ПМЦ 10752217 . ПМИД 37612512 . S2CID 254181409 .
- ^ Пертеа М., Зальцберг С.Л. (5 мая 2010 г.). «Между курицей и виноградом: оценка количества генов человека» . Геномная биология . 11 (5): 206. doi : 10.1186/gb-2010-11-5-206 . ПМК 2898077 . ПМИД 20441615 .
- ^ «Определение голандрического слова» . Словарь.com . Проверено 21 января 2020 г.
- ^ Jump up to: а б Халласт П., Эберт П., Лофтус М., Йилмаз Ф., Аудано П.А., Логсдон Г.А. и др. (сентябрь 2023 г.). «Сборка 43 Y-хромосом человека демонстрирует огромную сложность и разнообразие» . Природа . 621 (7978): 355–364. Бибкод : 2023Natur.621..355H . дои : 10.1038/s41586-023-06425-6 . ПМЦ 10726138 . ПМИД 37612510 . S2CID 261098546 .
- ^ «Данные идеограммы Homo sapience (400 ударов в час, сборка GRCh38.p3» . Страница украшения генома . Национальный центр биотехнологической информации США (NCBI). 04 июня 2014 г. Проверено 26 апреля 2017 г.
- ^ «Данные идеограммы Homo sapience (550 ударов в час, сборка GRCh38.p3)» . Страница оформления генома . Национальный центр биотехнологической информации США (NCBI). 11 августа 2015 г. Проверено 26 апреля 2017 г.
- ^ Международный постоянный комитет по цитогенетической номенклатуре человека (2013 г.). ISCN 2013: Международная система цитогенетической номенклатуры человека (2013) . Каргерское медицинское и научное издательство. ISBN 978-3-318-02253-7 .
- ^ Сетхакулвичай В., Манитпорнсут С., Вибунрат М., Лилакиацакун В., Ассавамакин А., Тонгсима С. (2012). «Оценка разрешения на уровне полос изображений хромосом человека». 2012 Девятая Международная конференция по информатике и программной инженерии (JCSSE) . стр. 276–282. дои : 10.1109/JCSSE.2012.6261965 . ISBN 978-1-4673-1921-8 . S2CID 16666470 .
- ^ " p ": Короткая рука; « q »: Длинная рука.
- ^ Номенклатуру цитогенетических полос см. в статье locus .
- ^ Jump up to: а б Эти значения (начало/конец ISCN) основаны на длине полос/идеограмм из книги ISCN «Международная система цитогенетической номенклатуры человека» (2013). Произвольная единица .
- ^ gpos : область, положительно окрашенная G-полосами , обычно богатая АТ и бедная генами; gneg : область, отрицательно окрашенная G-полосами, обычно богатая CG и богатая генами; acen Центромера . var : переменная область; стебель : Стебель.
- ^ «Ученые изменили форму дерева гаплогруппы Y-хромосомы, получив новое представление о происхождении человека» . Наука Дейли . 3 апреля 2008 г.
- ^ Пертеа М., Зальцберг С.Л. (2010). «Между курицей и виноградом: оценка количества генов человека» . Геномная биология . 11 (5): 206. doi : 10.1186/gb-2010-11-5-206 . ПМК 2898077 . ПМИД 20441615 .
- ^ «Статистика и загрузки для хромосомы Y» . Комитет по генной номенклатуре Хьюго . 12 мая 2017 г. Архивировано из оригинала 29 июня 2017 г. Проверено 19 мая 2017 г.
- ^ «Хромосома Y: Краткое описание хромосом — Homo sapiens» . Ансамбль Выпуск 88 . 2017-03-29 . Проверено 19 мая 2017 г.
- ^ «Человеческая хромосома Y: записи, названия генов и перекрестные ссылки на MIM» . ЮниПрот . 28 февраля 2018 г. Проверено 16 марта 2018 г.
- ^ «Гены, кодирующие Y-хромосому Homo sapiens» . База данных генов Национального центра биотехнологической информации . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ «Некодирующие гены Y-хромосомы человека разумного» . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ «Некодирующие псевдогены Y-хромосомы человека разумного» . 19 мая 2017 г. Проверено 20 мая 2017 г.
- ^ Колако С., Моди Д. (февраль 2018 г.). «Генетика Y-хромосомы человека и ее связь с мужским бесплодием» . Репродуктивная биология и эндокринология . 16 (1): 14. дои : 10.1186/s12958-018-0330-5 . ПМЦ 5816366 . ПМИД 29454353 .
- ^ Вираппа А.М., Падаканная П., Рамачандра Н.Б. (август 2013 г.). «Полиморфизм, основанный на вариациях числа копий, в новой псевдоаутосомной области 3 (PAR3) транспонированной X-хромосомой области человека (XTR) в Y-хромосоме». Функциональная и интегративная геномика . 13 (3): 285–293. дои : 10.1007/s10142-013-0323-6 . ПМИД 23708688 . S2CID 13443194 .
- ^ Раудсепп Т., Чоудхари Б.П. (6 января 2016 г.). «Эвтерова псевдоаутосомная область» . Цитогенетические и геномные исследования . 147 (2–3): 81–94. дои : 10.1159/000443157 . hdl : 10576/22940 . ПМИД 26730606 .
- ^ Зейхер А., Браун Т. (июль 2022 г.). «Мозаичная потеря Y-хромосомы при старении». Наука . 377 (6603): 266–7. Бибкод : 2022Sci...377..266Z . дои : 10.1126/science.add0839 . ПМИД 35857599 . S2CID 250579530 .
- ^ Jump up to: а б Форсберг Л.А. (май 2017 г.). «Потеря хромосомы Y (LOY) в клетках крови связана с повышенным риском заболеваний и смертности у стареющих мужчин» . Генетика человека . 136 (5): 657–663. дои : 10.1007/s00439-017-1799-2 . ПМК 5418310 . ПМИД 28424864 .
- ^ Jump up to: а б Форсберг Л.А., Раси С., Малмквист Н., Дэвис Х., Пасупулати С., Пакалапати Г. и др. (июнь 2014 г.). «Мозаичная потеря хромосомы Y в периферической крови связана с более короткой выживаемостью и более высоким риском рака» . Природная генетика . 46 (6): 624–8. дои : 10.1038/ng.2966 . ПМЦ 5536222 . ПМИД 24777449 .
- ^ Го X, Дай X, Чжоу Т, Ван Х, Ни Дж, Сюэ Дж, Ван X (апрель 2020 г.). «Мозаичная потеря Y-хромосомы человека: что, как и почему». Генетика человека . 139 (4): 421–446. дои : 10.1007/s00439-020-02114-w . ПМИД 32020362 . S2CID 211036885 .
- ^ Сано С., Хоритани К., Огава Х., Халвардсон Дж., Чавкин Н.В., Ван Ю. и др. (июль 2022 г.). «Гематопоэтическая потеря Y-хромосомы приводит к сердечному фиброзу и смертности от сердечной недостаточности» . Наука . 377 (6603): 292–7. Бибкод : 2022Sci...377..292S . дои : 10.1126/science.abn3100 . ПМЦ 9437978 . ПМИД 35857592 .
- Новостная статья: Колата Г (14 июля 2022 г.). «Поскольку Y-хромосомы исчезают с возрастом, риск сердечно-сосудистых заболеваний может возрастать» . Нью-Йорк Таймс . Проверено 21 августа 2022 г.
- ^ Коглан А. (13 декабря 2014 г.). «У мужчин больше шансов заболеть раком, чем у женщин» . Новый учёный : 17.
- ^ Думански Дж. П., Раси С., Лённ М., Дэвис Х., Ингельссон М., Гидрайтис В. и др. (январь 2015 г.). «Мутагенез. Курение связано с мозаичной потерей хромосомы Y» . Наука . 347 (6217): 81–83. Бибкод : 2015Sci...347...81D . дои : 10.1126/science.1262092 . ПМЦ 4356728 . ПМИД 25477213 .
- ^ «Потеря мужской половой хромосомы приводит к более ранней смерти мужчин» . Университет Вирджинии . Проверено 31 августа 2022 г.
- ^ «Потеря мужской половой хромосомы может привести к более ранней смерти мужчин – исследование» . Независимый . 14 июля 2022 г. Проверено 31 августа 2022 г.
- ^ Jump up to: а б Нуссбаум, Роберт Л. (2007). Генетика Томпсона и Томпсона в медицине . Макиннес, Родерик Р., Уиллард, Хантингтон Ф., Хамош, Ада, Томпсон, Маргарет В. (Маргарет Уилсон), 1920- (7-е изд.). Филадельфия: Сондерс/Эльзевир. ISBN 978-1416030805 . OCLC 72774424 .
- ^ Джейкобс П.А., Брантон М., Мелвилл М.М., Бриттен Р.П., Макклемонт В.Ф. (декабрь 1965 г.). «Агрессивное поведение, психическое отклонение и мужчина XYY». Природа . 208 (5017): 1351–2. Бибкод : 1965Natur.208.1351J . дои : 10.1038/2081351a0 . ПМИД 5870205 . S2CID 4145850 .
- ^ Ричардсон СС (2013). Сам секс: поиск мужского и женского пола в геноме человека . Чикаго: Университет Чикаго Press. п. 84. ИСБН 978-0-226-08468-8 .
- ^ Уиткин Х.А., Медник С.А., Шульсингер Ф., Баккестром Е., Кристиансен К.О., Гуденаф Д.Р. и др. (август 1976 г.). «Преступность у мужчин XYY и XXY» Наука 193 (4253): 547–555. Бибкод : 1976Sci...193..547W . дои : 10.1126/science.959813 . ПМИД 959813 .
- ^ Уиткин Х.А., Гуденаф Д.Р., Хиршхорн К. (октябрь 1977 г.). «Мужчины XYY: они преступно агрессивны?». Науки . 17 (6): 10–13. дои : 10.1002/j.2326-1951.1977.tb01570.x . ПМИД 11662398 .
- ^ Абеди М., Салманинежад А., Сахиния Э. (январь 2018 г.). «Редкий синдром 48, XYYY: описание случая и обзор литературы» . Отчеты о клинических случаях . 6 (1): 179–184. дои : 10.1002/ccr3.1311 . ПМК 5771943 . ПМИД 29375860 .
- ^ Копсида Э., Стергиакули Э., Линн П.М., Уилкинсон Л.С., Дэвис В. (2009). «Роль Y-хромосомы в работе мозга» . Открытый нейроэндокринологический журнал . 2 : 20–30. дои : 10.2174/1876528900902010020 . ПМЦ 2854822 . ПМИД 20396406 .
- ^ Шредер Дж., Тииликайнен А., Де ла Шапель А. (апрель 1974 г.). «Лейкоциты плода в кровотоке матери после родов. I. Цитологические аспекты» . Трансплантация . 17 (4): 346–354. дои : 10.1097/00007890-197404000-00003 . ПМИД 4823382 . S2CID 35983351 .
- ^ Бьянки Д.В., Зиквольф Г.К., Вейл Г.Дж., Сильвестр С., ДеМария М.А. (январь 1996 г.). «Клетки-предшественники плода мужского пола сохраняются в материнской крови в течение 27 лет после родов» . Труды Национальной академии наук Соединенных Штатов Америки . 93 (2): 705–8. Бибкод : 1996PNAS...93..705B . дои : 10.1073/pnas.93.2.705 . ПМК 40117 . ПМИД 8570620 .
- ^ Jump up to: а б Ян З., Ламберт Н.К., Гатри К.А., Портер А.Дж., Лубьер Л.С., Мадлен М.М. и др. (август 2005 г.). «Мужской микрохимеризм у женщин без сыновей: количественная оценка и корреляция с анамнезом беременности». Американский медицинский журнал . 118 (8): 899–906. doi : 10.1016/j.amjmed.2005.03.037 . ПМИД 16084184 .
- ^ Чан В.Ф., Гурно С., Монтин Т.Дж., Соннен Дж.А., Гатри К.А., Нельсон Дж.Л. (26 сентября 2012 г.). «Мужской микрохимеризм в женском мозге человека» . ПЛОС ОДИН . 7 (9): e45592. Бибкод : 2012PLoSO...745592C . дои : 10.1371/journal.pone.0045592 . ПМЦ 3458919 . ПМИД 23049819 .
Внешние ссылки
[ редактировать ]- Y-хромосома CHM13v2.0
- Геномный браузер Ensembl
- Информация о проекте «Геном человека» — стартовая панель «Хромосома Y человека»
- По теме: Y-хромосома — данные Института биомедицинских исследований Уайтхеда.
- Природа — сосредоточьтесь на Y-хромосоме
- Национальный институт исследования генома человека (NHGRI) — использование нового механизма сохраняет гены Y-хромосомы
- Ysearch.org - Публичная база данных Y-ДНК. Архивировано 4 января 2011 г. на Wayback Machine.
- Консорциум Y-хромосомы (YCC). Архивировано 16 января 2017 г. в Wayback Machine.
- Мужчина-человек NPR: работа все еще продолжается
- Генетическая генеалогия: об использовании анализа мтДНК и Y-хромосомы при проверке происхождения