Гаплогруппа ДНК Y-хромосомы человека

| Часть серии о |
| Генетическая генеалогия |
|---|
| Концепции |
| Связанные темы |
В генетике человека гаплогруппа ДНК Y-хромосомы человека представляет собой гаплогруппу , определяемую мутациями в нерекомбинирующих участках ДНК мужской Y -хромосомы (называемой Y-ДНК). Многие люди в гаплогруппе имеют одинаковое количество коротких тандемных повторов (STR) и типы мутаций, называемые однонуклеотидными полиморфизмами (SNP). [ 2 ]
Y-хромосома человека накапливает примерно две мутации за поколение. [ 3 ] Гаплогруппы Y-ДНК представляют собой основные ветви филогенетического дерева Y-хромосомы , которые имеют сотни или даже тысячи мутаций, уникальных для каждой гаплогруппы.
Самый последний общий предок Y-хромосомы (Y-MRCA, неофициально известный как Y-хромосомный Адам ) — это самый последний общий предок (MRCA), от которого все ныне живущие люди произошли по отцовской линии . По оценкам, Адам с Y-хромосомой жил примерно 236 000 лет назад в Африке. [ нужна ссылка ] . Изучая другие узкие места, большинство евразийских мужчин (мужчин из популяций за пределами Африки) происходят от человека, жившего в Африке 69 000 лет назад ( гаплогруппа CT ). Юго-Восточная Азия считается источником всех Y-хромосом неафриканского человека. [ 4 ] однако это было названо маловероятным. [ 5 ] Другие крупные узкие места произошли около 50 000 и 5 000 лет назад, и впоследствии происхождение большинства евразийских мужчин можно проследить до четырех предков, живших 50 000 лет назад и являвшихся потомками африканцев ( E-M168 ). [ 6 ] [ 7 ] [ 8 ]
Соглашение об именах
[ редактировать ]В этом разделе есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
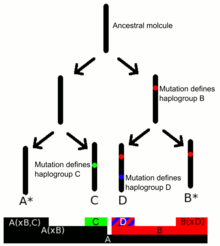
Гаплогруппы Y-ДНК определяются наличием ряда маркеров SNP Y-ДНК . Субклады определяются терминальным SNP , самым дальним SNP в филогенетическом дереве Y-хромосомы. [ 9 ] [ 10 ] Консорциум Y-хромосомы (YCC) разработал систему наименования основных гаплогрупп Y-ДНК с заглавными буквами от A до T, с дальнейшими субкладами, названными с использованием цифр и строчных букв (длинная номенклатура YCC ). В сокращенной номенклатуре YCC гаплогруппы Y-ДНК и их субклады называются с помощью первой буквы основной гаплогруппы Y-ДНК, за которой следует тире и имя определяющего терминального SNP. [ 11 ]
Номенклатура гаплогрупп Y-ДНК со временем меняется, чтобы учесть растущее число открываемых и тестируемых SNP и, как следствие, расширение филогенетического дерева Y-хромосомы. Это изменение номенклатуры привело к тому, что в разных источниках используется непоследовательная номенклатура. [ 2 ] Эта непоследовательность и все более громоздкая стенографическая номенклатура побудили к использованию более простой сокращенной номенклатуры.
Филогенетическая структура
[ редактировать ]
- Филогенетическое древо гаплогрупп Y-ДНК [ 12 ]
Основные гаплогруппы Y-ДНК
[ редактировать ]Гаплогруппы А и В
[ редактировать ]Гаплогруппа A — это макрогаплогруппа NRY ( нерекомбинирующая Y ), от которой происходят все современные отцовские гаплогруппы. Он редко распространен в Африке, концентрируясь среди койсанских популяций на юго-западе и нилотских популяций на северо-востоке в долине Нила. BT представляет собой субклад гаплогруппы A, точнее клады A1b (A2-T в Cruciani et al., 2011), а именно:
- Гаплогруппа А
- Гаплогруппа А00
- Гаплогруппа A0 (ранее также A1b)
- Гаплогруппа A1 (также A1a-T)
- Гаплогруппа A1a (M31)
- Гаплогруппа A1b (также A2-T; P108, V221)
- Гаплогруппа A1b1a1 (также A2; M14)
- Гаплогруппа A1b1b (также A3; M32)
- Гаплогруппа БТ (М91, М42, М94, М139, М299)
- Гаплогруппа B (M60)
- Гаплогруппа КТ
Гаплогруппа CT (P143)
[ редактировать ]Определяющими мутациями, разделяющими CT (все гаплогруппы, кроме A и B), являются M168 и M294. Место происхождения, вероятно, находится в Африке. Его возраст оценивается примерно в 88 000 лет. [ 13 ] [ 14 ] а в последнее время около 100 000 [ 15 ] или 101 000 лет. [ 16 ]
Гаплогруппа С (M130)
[ редактировать ]- Гаплогруппа C (M130, M216) встречается в Азии , Океании и Северной Америке.
- Гаплогруппа C1 (F3393/Z1426)
- Гаплогруппа C1a (CTS11043)
- Гаплогруппа C1a1 (M8, M105, M131) встречается с низкой частотой в Японии.
- Гаплогруппа C1a2 (V20) Встречается с низкой частотой в Европе , армянах , Алжире и Непале.
- Гаплогруппа C1b (F1370, Z16480)
- Гаплогруппа C1b1 (AM00694/K281)
- Гаплогруппа C1b1a (B66/Z16458)
- Гаплогруппа C1b1a1 (M356) Встречается с низкой частотой в Южной Азии , Юго-Западной Азии и северном Китае.
- Гаплогруппа C1b1a2 (B65)
- Гаплогруппа C1b1a2a (B67) обнаружена среди народа Леббо на Борнео, Индонезия.
- Гаплогруппа C1b1a2b (F725) обнаружена среди ханьцев (Гуандун, Хунань и Шэньси), народа дай (Юньнань), народа мурут (Бруней), малайцев (Сингапур) и народа аэта (Филиппины).
- Гаплогруппа C1b1a3 (Z16582) встречается с низкой частотой в Саудовской Аравии и Ираке.
- Гаплогруппа C1b1b (B68) обнаружена у народа Дусун (Бруней).
- Гаплогруппа C1b1a (B66/Z16458)
- Гаплогруппа C1b2 (C-Z16582)
- Гаплогруппа C1b3 (B477/Z31885)
- Гаплогруппа C1b3a (M38) обнаружена в Индонезии , Новой Гвинее , Меланезии , Микронезии и Полинезии.
- Гаплогруппа C1b3b (M347, P309) Встречается у коренных народов Австралии .
- Гаплогруппа C1b1 (AM00694/K281)
- Гаплогруппа C1a (CTS11043)
- Гаплогруппа C2 (M217, P44) Встречается по всей Евразии и Северной Америке , но особенно среди монголов , казахов , тунгусских народов , палеосибирцев и на-дене. народов, говорящих
- Гаплогруппа C1 (F3393/Z1426)
Гаплогруппа D (CTS3946)
[ редактировать ]- Гаплогруппа D (CTS3946)
- Гаплогруппа D1 (M174) Встречается в Японии , Китае (особенно в Тибете ), на Андаманских островах.
- Гаплогруппа D1a (CTS11577)
- Гаплогруппа D1a1 (Z27276, Z27283, Z29263)
- Гаплогруппа D1a1a (M15). Встречается в основном у тибетцев , народов Цянги , И и Хмонг-Миен. народов
- Гаплогруппа D1a1b (P99) Встречается в основном у тибетцев , цянских народов , наси и тюркских народов.
- Гаплогруппа D1a2 (M55, M57, M64.1, M179, P12, P37.1, P41.1 (M359.1), 12f2.2) Встречается преимущественно в Японии.
- Гаплогруппа D1a3 (Y34637) Встречается у андаманских народов ( Онге , Джарава )
- Гаплогруппа D1a1 (Z27276, Z27283, Z29263)
- Гаплогруппа D1b (L1366, L1378, M226.2) найдена на острове Мактан , Филиппины.
- Гаплогруппа D1a (CTS11577)
- Гаплогруппа D2 (A5580.2) обнаружена в Нигерии , Саудовской Аравии и Сирии.
- Гаплогруппа D1 (M174) Встречается в Японии , Китае (особенно в Тибете ), на Андаманских островах.
Гаплогруппа Е (M96)
[ редактировать ]- Гаплогруппа E (M40, M96) встречается в Африке, некоторых частях Ближнего Востока и Европы.
- Гаплогруппа E1 (P147)
- Гаплогруппа E1a (M33, M132), ранее E1
- Гаплогруппа E1b (P177)
- Гаплогруппа E1b1 (P2, DYS391p); ранее E3
- Гаплогруппа E1b1a (V38)
- Гаплогруппа E1b1a1 (M2). Встречается в Африке, особенно среди нигерско-конгоязычного населения. ; ранее E3a
- Гаплогруппа E1b1a2 (M329) Встречается в Африке, особенно в Эфиопии среди омотикоязычного населения. ; ранее E3*
- Гаплогруппа E1b1b (M215)
- Гаплогруппа E1b1b1 (M35) встречается на Африканском Роге , в Северной Африке , на Ближнем Востоке и в Европе (особенно в районах вблизи Средиземноморья и Балкан ) ; ранее E3b
- Гаплогруппа E1b1a (V38)
- Гаплогруппа E1b1 (P2, DYS391p); ранее E3
- Гаплогруппа E2 (M75)
- Гаплогруппа E1 (P147)
Гаплогруппа F (M89)
[ редактировать ]Группы, происходящие от гаплогруппы F, встречаются примерно у 90% населения мира, но почти исключительно за пределами Африки к югу от Сахары.
F xG,H,I,J,K редко встречается у современных популяций и пиков Южной Азии , особенно Шри-Ланки . [ 12 ] Похоже, что он также уже давно присутствует в Юго-Восточной Азии ; Сообщается, что уровень заболеваемости составляет 4–5% в Сулавеси и Лембате . Одно исследование, в котором не проводился всесторонний скрининг других субкладов F-M89 (включая некоторые субклады GHIJK), показало, что индонезийские мужчины с SNP P14/PF2704 (который эквивалентен M89) составляют 1,8% мужчин в Западном Тиморе . 1,5% Флореса, 5,4% Лембаты, 2,3% Сулавеси и 0,2% Суматры . [ 17 ] [ 18 ] F* (F xF1,F2,F3) зарегистрирован у 10% мужчин в Шри-Ланке и Южной Индии , у 5% в Пакистане, а также более низкие уровни среди народа таманг (Непал) и в Иране . F1 (P91), F2 (M427) и F3 (M481; ранее F5) очень редки и практически эксклюзивны для регионов/этнических меньшинств в Шри-Ланке, Индии, Непале, Южном Китае , Таиланде , Бирме и Вьетнаме . Однако в таких случаях вероятность ошибочной идентификации считается относительно высокой, и некоторые из них могут принадлежать к ошибочно идентифицированным субкладам гаплогруппы GHIJK . [ 19 ]
Гаплогруппа G (M201)
[ редактировать ]Гаплогруппа G (M201) возникла около 48 000 лет назад, а ее последний общий предок, вероятно, жил 26 000 лет назад на Ближнем Востоке. Он распространился в Европе с неолитической революцией .
Он встречается у многих этнических групп Евразии; наиболее распространен на Кавказе , в Иране , Анатолии и Леванте . Встречается почти во всех европейских странах, но наиболее распространен в Гагаузии , юго-восточной Румынии , Греции , Италии , Испании , Португалии , Тироле и Богемии , с самой высокой концентрацией на некоторых средиземноморских островах; редко встречается в Северной Европе . [ 20 ] [ 21 ]
G-M201 также встречается в небольших количествах на северо-западе Китая и в Индии , Бангладеш , Пакистане , Шри-Ланке , Малайзии и Северной Африке .
Гаплогруппа H (M69)
[ редактировать ]Гаплогруппа H (M69), вероятно, возникла в южной части Центральной Азии , Южной Азии или Западной Азии около 48 000 лет назад и остается там широко распространенной в формах H1 (M69) и H3 (Z5857). Его субклады также с меньшей частотой встречаются в Иране, Центральной Азии, на Ближнем Востоке и на Аравийском полуострове.
Однако H2 (P96) присутствует в Европе со времен неолита, а H1a1 (M82) распространился на запад в эпоху Средневековья с миграцией цыган .
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( сентябрь 2016 г. ) |
Гаплогруппа I (M170)
[ редактировать ]Гаплогруппа I (М170, М258) встречается преимущественно в Европе и на Кавказе .
- Гаплогруппа I1 нордиды / нордические европеоиды (M253). Встречаются в основном в Северной Европе.
- Гаплогруппа I2 Динариды/Динарские европиды (P215) Встречается в основном на Балканах, в юго-восточной Европе и Сардинии, за исключением I2B1 (m223), который с умеренной частотой встречается в Западной, Центральной и Северной Европе.
Гаплогруппа J (M304)
[ редактировать ]Гаплогруппа J (M304, S6, S34, S35) встречается преимущественно на Ближнем Востоке , Кавказе и в Юго-Восточной Европе .
- Гаплогруппа J* (J-M304*) редко встречается за пределами острова Сокотра .
- Гаплогруппа J1 семитид/бедуиниды арабиды (M267) связаны с Северо-Восточного Кавказа народами в Дагестане и людьми, говорящими на семитских языках на Ближнем Востоке, в Эфиопии и Северной Африке , а также встречаются в Средиземноморской Европе с меньшими частотами, так же, как и гаплогруппа T.
- Гаплогруппа J2 Сириды/Нахраиниды Арабиды (M172) встречается в основном у семитоязычных народов , Анатолии , Греции , Балкан , Италии , Ирана , Кавказа , Южной Азии и Средней Азии .
Гаплогруппа К (М9)
[ редактировать ]Гаплогруппа К (М9) распространена по всей Евразии , Океании и среди коренных американцев .
K(xLT,K2a,K2b) – то есть K*, K2c, K2d или K2e – встречается главным образом в Меланезии , у австралийских аборигенов , Индии , Полинезии и на островах Юго-Восточной Азии .
Гаплогруппы L и Т (К1)
[ редактировать ]Гаплогруппа L (M20) встречается в Южной Азии, Центральной Азии, Юго-Западной Азии и Средиземноморье.
Гаплогруппа T (M184, M70, M193, M272) встречается на высоком уровне на Африканском Роге (в основном у народов, говорящих на кушитском языке ), в некоторых частях Южной Азии , на Ближнем Востоке и в Средиземноморье . Т-М184 также встречается у значительных меньшинств шакченси , штильфсеров , египтян , оманцев , евреев-сефардов , [ 22 ] Ибицы (Eivissencs) и Тубу . Он также встречается на низких частотах в других частях Средиземноморья и Южной Азии .
Гаплогруппа К2 (К-М526)
[ редактировать ]Единственные живые самцы, которые, как сообщается, являются носителями базальной парагруппы K2*, являются коренными австралийцами . Крупные исследования, опубликованные в 2014 и 2015 годах, показывают, что до 27% мужчин австралийских аборигенов являются носителями K2 *, в то время как другие несут субклад K2.
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( сентябрь 2016 г. ) |
Гаплогруппы K2a, K2a1, NO и NO1
[ редактировать ]Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( сентябрь 2016 г. ) |
Гаплогруппа N
[ редактировать ]Гаплогруппа N (М231) встречается в Северной Евразии, особенно среди носителей уральских языков .
Гаплогруппа N, возможно, возникла в Восточной Азии и распространилась как на север, так и на запад, в Сибирь , являясь наиболее распространенной группой, встречающейся у некоторых уральскоязычных народов .
Гаплогруппа О
[ редактировать ]Гаплогруппа O (M175) с наибольшей частотой встречается в Восточной и Юго-Восточной Азии , с более низкой частотой — в южной части Тихого океана , Центральной Азии , Южной Азии и на островах Индийского океана ( например , на Мадагаскаре, Коморских островах).
- Гаплогруппа O1 (F265/M1354, CTS2866, F75/M1297, F429/M1415, F465/M1422)
- Гаплогруппа O1a (M119, CTS31, F589/Page20, L246, L466). Встречается в восточном, центральном и южном материковом Китае , на Тайване и в Юго-Восточной Азии , особенно среди австронезийских и тай-кадайских народов .
- Гаплогруппа O1b (P31, M268)
- Гаплогруппа O1b1 (M95). Встречается в Японии , Китае, Тайване, Юго-Восточной Азии и на Индийском субконтиненте , особенно среди австроазиатских и тай-кадайских народов, малайцев и индонезийцев.
- Гаплогруппа O1b2 (SRY465, M176) обнаружена в Японии, Корее , Маньчжурии и Юго-Восточной Азии.
- Гаплогруппа O2 (M122) встречается по всей Восточной Азии, Юго-Восточной Азии и Австронезии , включая Полинезию.
Гаплогруппы K2b1, M и S
[ редактировать ]Примеров базальной парагруппы K2b1* не обнаружено. Мужчины, несущие субклады K2b1, встречаются преимущественно среди папуасских народов , микронезийских народов , коренных австралийцев и полинезийцев .
Его основные субклады представляют собой две основные гаплогруппы:
- Гаплогруппа S (B254), также известная как K2b1a: найдена в высокогорьях Папуа-Новой Гвинеи и;
- Гаплогруппа M (P256), также известная как K2b1b: обнаружена в Новой Гвинее и Меланезии.
Этот раздел нуждается в расширении . Вы можете помочь, добавив к нему . ( сентябрь 2016 г. ) |
Гаплогруппа P (K2b2)
[ редактировать ]Гаплогруппа P (P295) имеет две основные ветви: P1 (P-M45) и чрезвычайно редкую P2 (P-B253). [ 23 ]
P*, P1* и P2 встречаются вместе только на острове Лусон на Филиппинах . [ 23 ] В частности, P* и P1* в значительной степени встречаются среди представителей народа аэта (или агта) на Лусоне. [ 24 ] Хотя P1* в настоящее время более распространен среди живых людей в Восточной Сибири и Центральной Азии , он также встречается в небольших количествах в материковой части Юго-Восточной Азии и Южной Азии . В совокупности эти распределения позволяют предположить, что P* возник из K2b в Юго-Восточной Азии. [ 24 ] [ 25 ]
P1 также является родительским узлом двух основных клад:
- Гаплогруппа Q (Q-M242) и;
- Гаплогруппа R (R-M207). Они имеют общий маркер M45 в дополнение как минимум к 18 другим SNP.
Гаплогруппа Q (MEH2, M242, P36) обнаружена в Сибири и Америке. Гаплогруппа R (M207, M306): встречается в Европе , Западной Азии , Центральной Азии и Южной Азии.
Гаплогруппа Q M242
[ редактировать ]Q определяется SNP M242. Считается, что он возник в Центральной Азии примерно 32 000 лет назад. [ 26 ] [ 27 ] Субклады гаплогруппы Q с их определяющими мутациями согласно ISOGG 2008 года. дереву [ 28 ] приведены ниже. ss4 bp, rs41352448, не представлен в дереве ISOGG 2008, поскольку это значение для STR. Это низкочастотное значение было обнаружено как новая линия Q (Q5) в индийском населении. [ 29 ]
Дерево ISOGG 2008 г.
- Вопрос (M242)
- Вопрос*
- К1 (P36.2)
- Q1*
- Q1a (MEH2)
- Вопрос 1а*
- Q1a1 (M120, M265/N14) Встречается с низкой частотой среди бутанцев , [ 30 ] дунгане , ханьцы , японцы , корейцы , монголы , [ 31 ] Наси и тибетцы [ 32 ] [ 33 ]
- Q1a2 (M25, M143) Встречается с низкой и умеренной частотой среди некоторых популяций Юго-Западной Азии , Центральной Азии и Сибири.
- Q1a3 (M346)
- Q1a3* Встречается с низкой частотой в Пакистане , Индии и Тибете.
- Q1a3a (M3) Типично для коренных народов Америки
- Q1a3a*
- Q1a3a1 (M19) Встречается у некоторых коренных народов Южной Америки , таких как тикуна и вайуу. [ 34 ]
- Q1a3a2 (M194)
- Q1a3a3 (M199, P106, P292)
- Q1a4 (P48)
- Q1a5 (P89)
- Q1a6 (M323) Встречается у значительного меньшинства йеменских евреев.
- Q1b (M378) Встречается с низкой частотой среди образцов хазарейцев и синдхов.
Гаплогруппа R (M207)
[ редактировать ]
Гаплогруппа R определяется SNP M207. Основная часть гаплогруппы R представлена в субкладе-потомке R1 (M173), зародившемся в Сибири . У R1 есть два субклада-потомка: R1a и R1b .
R1a связан с протоиндоиранскими и балтославянскими народами и сейчас встречается преимущественно в Центральной Азии , Южной Азии и Восточной Европе .
Гаплогруппа R1b является доминирующей гаплогруппой Западной Европы, а также встречается редко распространенной среди различных народов Азии и Африки . Его субклад R1b1a2 (M269) представляет собой гаплогруппу, которая чаще всего встречается среди современного населения Западной Европы и связана с итало-кельтскими и германскими народами.
- Гаплогруппа R1 (M173) встречается по всей Западной Евразии.
- Гаплогруппа R1a (M420) встречается в Центральной Азии, Южной Азии, Центральной, Северной и Восточной Европе, на Балканах.
- Гаплогруппа R1b (M343) встречается в Западной Европе, Западной Азии , Центральной Азии , Северной Африке и северном Камеруне.
- Гаплогруппа R2 (M124) встречается в Южной Азии, на Кавказе, в Центральной Азии и Восточной Европе.
Хронологическое развитие гаплогрупп
[ редактировать ]| Гаплогруппа | Возможное время происхождения | Возможное место происхождения | Возможный TMRCA [ 35 ] [ 14 ] |
|---|---|---|---|
| А00 | 235,900 [ 7 ] или 275 000 лет назад [ 36 ] | Африка [ 37 ] | 235 900 лет назад |
| БТ | 130 700 лет назад [ 7 ] | Африка | 88000 лет назад |
| Коннектикут | 88,000 [ 7 ] или 101-100 000 лет назад [ 15 ] [ 16 ] | Африка | 68 500 лет назад |
| И | 65,200, [ 7 ] 69,000, [ 38 ] или 73000 лет назад [ 39 ] | Восточная Африка [ 40 ] или Азия [ 17 ] | 53 100 лет назад |
| Ф | 65 900 лет назад [ 7 ] | Евразия | 48 800 лет назад |
| Г | 48 500 лет назад [ 7 ] | Средний Восток | 26 200 лет назад |
| Эй Джей | 47 200 лет назад [ 7 ] | Средний Восток | 42900 лет назад |
| К | 47 200 лет назад [ 7 ] | Азия | 45 400 лет назад |
| П | 45 400 лет назад [ 7 ] | Азия | 31900 лет назад |
| Дж | 42900 лет назад [ 7 ] [ 41 ] | Средний Восток | 31600 лет назад |
| я | 42900 лет назад [ 7 ] | Европа | 27 500 лет назад |
| Э-М215 (E1b1b) | 42 300 лет назад [ 7 ] [ 42 ] | Восточная Африка | 34800 лет назад |
| Э-В38 (E1b1a) | 42 300 лет назад [ 7 ] [ 42 ] | Восточная Африка | 40 100 лет назад |
| Н | 36 800 лет назад [ 7 ] [ 43 ] | Азия | 22 100 лет назад |
| E1b1b-M35 | 34800 лет назад [ 7 ] [ 42 ] | Восточная Африка | 24 100 лет назад |
| Р | 31900 лет назад [ 7 ] | Азия | 28 200 лет назад |
| J-M267 (J1) | 31600 лет назад [ 7 ] [ 41 ] | Средний Восток | 18 500 лет назад |
| J-M172 (J2) | 31600 лет назад [ 7 ] [ 41 ] | Средний Восток | 27 800 лет назад [ 7 ] [ 44 ] |
| Р-М173 (Р1) | 28 200 лет назад [ 7 ] | Азия | 22800 лет назад |
| И-М253 (И1) | 27 500 лет назад [ 7 ] [ 45 ] [ 46 ] | Европа | 4600 лет назад |
| И-М438 (И2) | 27 500 лет назад [ 7 ] [ 46 ] | Европа | 21800 лет назад |
| Р-М420 (Р1а) | 22800 лет назад [ 7 ] [ 47 ] | Евразия | 18 300 лет назад |
| Р-М343 (Р1б) | 22800 лет назад [ 7 ] [ 48 ] | Евразия [ 49 ] | 20 400 лет назад |
| И2-Л460 (И2а) | 21800 лет назад [ 7 ] [ 50 ] | Европа | 21 100 лет назад |
| И2а-П37 | 21 100 лет назад [ 7 ] [ 45 ] [ 51 ] | Европа | 18 500 лет назад |
| E1b1b-M78 | 19 800 лет назад [ 7 ] [ 42 ] [ 52 ] | Северо-Восточная Африка [ 52 ] | 13 400 лет назад [ 7 ] [ 52 ] |
| И2а-М423 | 18 500 лет назад [ 7 ] [ 51 ] | Европа | 13 500 лет назад |
| И2а-М223 | 17 400 лет назад [ 7 ] | Европа | 12 100 лет назад |
| N1c-M178 | 14 200 лет назад [ 7 ] [ 43 ] | Азия | 11900 лет назад |
| Р1а-М17 | 14 100 лет назад [ 7 ] [ 47 ] [ 53 ] | Восточная Европа | 8500 лет назад |
| Р1б-М269 | 13 300 лет назад [ 7 ] | Восточная Европа | 6400 лет назад [ 54 ] |
| E1b1b-V12 | 11800 лет назад [ 7 ] [ 52 ] | Северная Африка | 9900 лет назад |
| E-U175 (E1b1a8) | 9200 лет назад [ 7 ] [ 42 ] | Восточная Африка | 8500 лет назад |
| E1b1b-V13 | 8100 лет назад [ 7 ] [ 52 ] | Южная Европа | 4800 лет назад |
| Э-М191 (E1b1a7) | 7400 лет назад [ 7 ] [ 42 ] | Восточная Африка | 6400 лет назад |
| Е-U174 (E1b1a-U174) | 6400 лет назад [ 7 ] [ 42 ] | Восточная Африка | 5300 лет назад |
| R1b-L151 | 5800 лет назад [ 7 ] | Восточная Европа | 4800 лет назад |
| R1a-Z280 | 5000 лет назад [ 7 ] | Восточная Европа | 4600 лет назад [ 55 ] |
| Р1а-М458 | 4700 лет назад [ 7 ] | Восточная Европа | 4700 лет назад [ 55 ] |
См. также
[ редактировать ]- Список гаплогрупп Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК в населении Европы
- Генетическая история Европы
- Список однонуклеотидных полиморфизмов Y-ДНК
- Список маркеров Y-STR
- Гаплогруппа митохондриальной ДНК человека
- * (гаплогруппа)
- Молекулярная филогения
- Генетическая генеалогия
- Генеалогический ДНК-тест
- Таблица перевода гаплогрупп Y-хромосомы
Ссылки
[ редактировать ]- ^ Халласт П., Агдхоян А., Балановский О., Сюэ Ю., Тайлер-Смит С. (2020). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/ s00439-020-02204-9 ПМЦ 7864842 . ПМИД 32666166 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Перейти обратно: а б «Понимание гаплогрупп: как называются гаплогруппы?» . ДНК генеалогического древа. Архивировано из оригинала 21 июня 2012 года . Проверено 31 марта 2013 г.
- ^ Долгин, Эли (2009). «Выявлена частота мутаций у человека» . Природа . дои : 10.1038/news.2009.864 . Проверено 18 сентября 2017 г. «одна мутация на каждые 30 миллионов пар оснований»
- ^ Халласт, Таблетки; Грин, Анастасия; Балановский Олег; Сюэ, Он был; Тайлер-Смит, Крис (2021). «Происхождение современных неафриканских Y-хромосом человека из Юго-Восточной Азии» . Генетика человека . 140 (2): 299–307. дои : 10.1007/ s00439-020-02204-9 ISSN 0340-6717 . ПМЦ 7864842 . ПМИД 32666166 .
- ^ Сент-Клер, Майкл (2021). Предыстория языка: взгляд на триангулированную Y-хромосому . Штутгарт, Германия: Генетико-лингвистический интерфейс. п. 29. ISBN 978-3-00-071045-2 .
- ^ Кармин; и др. (2015). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 . «Мы датируем самого последнего общего предка Y-хромосомы (MRCA) в Африке 254 (95% ДИ 192–307) тысячелетий назад и обнаруживаем кластер основных неафриканских гаплогрупп-основателей в узком временном интервале в 47–52 тысячелетий, что соответствует с моделью быстрой первоначальной колонизации Евразии и Океании после узкого места за пределами Африки. В отличие от демографических реконструкций, основанных на мтДНК, мы предполагаем второе сильное узкое место в линиях Y-хромосомы, датируемое последними 10 тысячелетиями. «Узкое место» вызвано культурными изменениями, влияющими на различия в репродуктивном успехе среди мужчин».
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а к ап «YFull YTree» . YПолный . Проверено 15 сентября 2017 г.
- ^ «Что-то странное случилось с людьми 7000 лет назад, и мы наконец-то знаем, почему» . 31 мая 2018 г.
Около 7000 лет назад, еще в эпоху неолита, с генетическим разнообразием человека произошло нечто действительно необычное. В течение следующих 2000 лет, что наблюдалось в Африке, Европе и Азии, генетическое разнообразие Y-хромосомы рухнуло, и на каждые 17 женщин приходился только один мужчина.
- ^ «Понимание результатов: однонуклеотидный полиморфизм Y-ДНК (SNP): что такое гаплогруппа ДНК Y-хромосомы (Y-ДНК)?» . ДНК генеалогического древа . Проверено 31 марта 2013 г.
Гаплогруппы ДНК Y-хромосомы (Y-ДНК) являются основными ветвями отцовского генеалогического древа человека. Каждая гаплогруппа имеет множество ответвлений. Это субклады.
- ^ «Руководство пользователя myFTDNA 2.0: Y-ДНК: Что такое страница Y-ДНК — совпадения?» . ДНК генеалогического древа . Проверено 31 марта 2013 г.
Терминальный SNP определяет конечную (конечную) подветвь дерева Y-ДНК, к которой принадлежит человек.
- ^ «Понимание результатов: однонуклеотидный полиморфизм Y-ДНК (SNP): как называются гаплогруппы и их субклады?» . ДНК генеалогического древа . Проверено 31 марта 2013 г.
- ^ Перейти обратно: а б Авторские права: ISOGG, 2015 г. «Ствол дерева гаплогруппы Y-ДНК ISOGG 2015» . isogg.org .
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Андерхилл и Кивисилд; Кивисилд, Т (2007). «Использование структуры популяции Y-хромосомы и митохондриальной ДНК для отслеживания миграций человека». Анну. Преподобный Жене . 41 (1): 539–64. дои : 10.1146/annurev.genet.41.110306.130407 . ПМИД 18076332 .
- ^ Перейти обратно: а б Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, МФ (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–38. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Перейти обратно: а б Камин М., Сааг Л., Винсенте М. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–66. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Перейти обратно: а б Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арсьеро Э., Хуанмин Ю., Томас М.Г., Сюэ Ю., Тайлер-Смит С. (июнь 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–28. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- ^ Перейти обратно: а б Кьярони, Жак; Андерхилл, Питер А.; Кавалли-Сфорца, Лука Л. (1 декабря 2009 г.). «Разнообразие Y-хромосомы, человеческая экспансия, дрейф и культурная эволюция» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (48): 20174–79. Бибкод : 2009PNAS..10620174C . дои : 10.1073/pnas.0910803106 . ПМЦ 2787129 . ПМИД 19920170 .
- ^ Тумонггор, Мерианн К. (2014). «Изоляция, контакты и социальное поведение сформировали генетическое разнообразие в Западном Тиморе» . Журнал генетики человека . 59 (9): 494–503. дои : 10.1038/jhg.2014.62 . ПМЦ 4521296 . ПМИД 25078354 .
- ^ Так было, например, с исходным субкладом F3 (M96), который с тех пор был переименован в гаплогруппу H2 .
- ^ Пассарино Г., Каваллери Г.Л., Лин А.А., Кавалли-Сфорца Л.Л., Борресен-Дейл А.Л., Андерхилл П.А. (2002). «Различные генетические компоненты норвежской популяции, выявленные с помощью анализа полиморфизмов мтДНК и Y-хромосомы» . Европейский журнал генетики человека . 10 (9): 521–29. дои : 10.1038/sj.ejhg.5200834 . ПМИД 12173029 .
- ^ Карлссон, Андреас О; Валлерстрем, Томас; Гётерстрем, Андерс; Холмлунд, Гунилла (2006). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива» . Европейский журнал генетики человека . 14 (8): 963–70. дои : 10.1038/sj.ejhg.5201651 . ПМИД 16724001 .
- ^ Ногейро, Инес (2009). «Филогеографический анализ отцовских линий в еврейских общинах северо-восточной Португалии» . Американский журнал физической антропологии . 141 (3): 373–81. дои : 10.1002/ajpa.21154 . ПМИД 19918998 .
- ^ Перейти обратно: а б ISOGG, 2016, Гаплогруппа P Y-ДНК и ее субклады – 2016 (20 июня 2016 г.).
- ^ Перейти обратно: а б Тумонггор, Мерианн К; Карафет Татьяна М; Дауни, Шон; Лансинг, Дж. Стивен; Норквест, Питер; Судойо, Херавати; Хаммер, Майкл Ф; Кокс, Мюррей П. (31 июля 2014 г.). «Изоляция, контакты и социальное поведение сформировали генетическое разнообразие в Западном Тиморе» . Журнал генетики человека . 59 (9): 494–503. дои : 10.1038/jhg.2014.62 . ПМЦ 4521296 . ПМИД 25078354 .
- ^ Татьяна М Карафет; и др. (2015). «Улучшенное филогенетическое разрешение и быстрая диверсификация гаплогруппы Y-хромосомы K-M526 в Юго-Восточной Азии» . Европейский журнал генетики человека . 23 (3): 369–73. дои : 10.1038/ejhg.2014.106 . ПМЦ 4326703 . ПМИД 24896152 .
- ^ Фагундес, Нельсон-младший; Каниц, Рикардо; Эккерт, Роберта; Валлс, Ана К.С.; Бого, Маурисио Р.; Сальзано, Франсиско М.; Смит, Дэвид Гленн; Сильва, Уилсон А.; Заго, Марко А.; Рибейру-дус-Сантос, Андреа К.; Сантос, Сидни Э.Б.; Петцль-Эрлер, Мария Луиза ; Бонатто, Сандро Л. (2008). «Геномика митохондриальной популяции подтверждает единое происхождение до Кловиса с прибрежным маршрутом заселения Америки» (PDF) . Американский журнал генетики человека . 82 (3): 583–92. дои : 10.1016/j.ajhg.2007.11.013 . ПМЦ 2427228 . ПМИД 18313026 . Архивировано из оригинала (PDF) 25 марта 2009 г. Проверено 22 мая 2013 г.
Со времени первых исследований было обнаружено, что современные индейские популяции имеют почти исключительно пять «гаплогрупп мтДНК» (A–D и X)6, классифицированных в автохтонные гаплогруппы A2, B2, C1, D1 и X2a.7 Гаплогруппы A– D встречаются по всему Новому Свету и часто встречаются в Азии, что подтверждает северо-восточное азиатское происхождение этих линий.
- ^ Зегура, СЛ; Карафет, ТМ; Животовский, Л.А.; Хаммер, МФ (2003). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Молекулярная биология и эволюция . 21 (1): 164–75. дои : 10.1093/molbev/msh009 . ПМИД 14595095 .
- ^ «Дерево гаплогрупп Y-ДНК 2010» . Международное общество генетической генеалогии . Проверено 1 июля 2010 г.
- ^ Шарма, Сваркар; Рай, Экта; Бхат, Аудеш К; Бханвер, Амарджит С; Соответствующий Бамезай, Рамешвар Н.К. (2007). «Новая подгруппа Q5 гаплогруппы Q Y-хромосомы человека в Индии» . БМК Эвол Биол . 7 (1): 232. Бибкод : 2007BMCEE...7..232S . дои : 10.1186/1471-2148-7-232 . ПМК 2258157 . ПМИД 18021436 .
- ^ Халласт, П.; Батини, К.; Задик, Д.; Местонахождение Дельсер, П.; Веттон, Дж. Х.; Браун-Стрим, Э.; Каваллери, ГЛ; Де Книфф, П.; Дестро Бисол, Г.; Дюпюи, БМ; Эриксен, ХА; Джорде, LB; Кинг, TE; Лармюсо, Миннесота; Лопес Де Мунаин, А.; Лопес-Парра, AM; Лутрадис, А.; Милас, Дж.; Новеллетто, А.; Памджав, Х.; Саджантила, А.; Шемпп, В.; Сирс, М.; Толун, А.; Тайлер-Смит, К.; Ван Гейстелен, А.; Уоткинс, С.; Винни, Б.; Джоблинг, Массачусетс (2015). «Дерево Y-хромосомы превращается в лист: 13 000 SNP с высокой степенью достоверности, охватывающие большинство известных клад» . Молекулярная биология и эволюция . 32 (3): 661–73. дои : 10.1093/molbev/msu327 . ПМЦ 4327154 . ПМИД 25468874 .
- ^ Ди Кристофаро, Джули; Пеннарун, Эрван; Мазьер, Стефан; Майрес, Натали М.; Лин, Алиса А.; Темори, Шах Ага; Мецпалу, Мейт; Мецпалу, Эне; Витцель, Майкл; Кинг, Рой Дж.; Андерхилл, Питер А.; Виллемс, Ричард; Кьярони, Жак (2013). «Афганский Гиндукуш: место, где сходятся потоки генов Евразийского субконтинента» . ПЛОС ОДИН . 8 (10): е76748. Бибкод : 2013PLoSO...876748D . дои : 10.1371/journal.pone.0076748 . ПМЦ 3799995 . ПМИД 24204668 .
- ^ Вэнь Б., Ли Х., Лу Д. и др. (сентябрь 2004 г.). «Генетические данные подтверждают демическое распространение ханьской культуры». Природа . 431 (7006): 302–05. Бибкод : 2004Natur.431..302W . дои : 10.1038/nature02878 . ПМИД 15372031 . S2CID 4301581 .
- ^ Уэллс Р.С., Юлдашева Н., Рузибакиев Р. и др. (август 2001 г.). «Центр Евразии: континентальный взгляд на разнообразие Y-хромосомы» . Учеб. Натл. акад. наук. США . 98 (18): 10244–49. Бибкод : 2001PNAS...9810244W . дои : 10.1073/pnas.171305098 . ПМК 56946 . ПМИД 11526236 .
- ^ Бортолини М.К., Салцано Ф.М., Томас М.Г. и др. (сентябрь 2003 г.). «Y-хромосома свидетельствует о различиях в древней демографической истории Америки» . Являюсь. Дж. Хум. Жене . 73 (3): 524–39. дои : 10.1086/377588 . ПМК 1180678 . ПМИД 12900798 .
- ^ Батини, Кьяра; Халласт, Пилле; Задик, Дэниел; Дельсер, Пьерпаоло Майзано; Бенаццо, Андреа; Гиротто, Сильвия; Арройо-Пардо, Эдуардо; Каваллери, Джанпьеро Л.; Де Книфф, Питер; Дюпюи, Берит Мире; Эриксен, Хайди А.; Кинг, Тури Э.; Де Мунаин, Адольфо Лопес; Лопес-Парра, Ана М.; Лутрадис, Афродита; Миласин, Елена; Новеллетто, Андреа; Памджав, Хоролма; Саджантила, Антти; Толун, Аслихан; Винни, Брюс; Джоблинг, Марк А. (2015). «Крупномасштабное недавнее расширение европейских отцов, продемонстрированное изменением последовательности населения» . Природные коммуникации . 6 : 7152. Бибкод : 2015NatCo...6.7152B . дои : 10.1038/ncomms8152 . ПМЦ 4441248 . ПМИД 25988751 .
- ^ Мендес, Л.; и др. (2016). «Расхождение Y-хромосом неандертальца и современного человека» . Американский журнал генетики человека . 98 (4): 728–34. дои : 10.1016/j.ajhg.2016.02.023 . ПМЦ 4833433 . ПМИД 27058445 .
- ^ «Отцу всех людей 340 000 лет» .
- ^ Камин М., Сааг Л., Винсенте М. и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Хабер М., Джонс А.Л., Коннел Б.А., Асан, Арсьеро Э., Хуанмин Ю., Томас М.Г., Сюэ Ю., Тайлер-Смит С. (июнь 2019 г.). «Редкая глубоко укоренившаяся африканская гаплогруппа Y-хромосомы D0 и ее значение для расселения современного человека за пределы Африки» . Генетика . 212 (4): 1421–1428. дои : 10.1534/genetics.119.302368 . ПМК 6707464 . ПМИД 31196864 .
- ^ Семино, Орнелла; Магри, Кьяра; Бенуцци, Джорджия; Лин, Алиса А.; Аль-Захери, Надя; Батталья, Винченца; Макчиони, Лилиана; Триантафиллидис, Костас; и др. (2004). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Американский журнал генетики человека . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Перейти обратно: а б с Семино, О; Магри, К; Бенуцци, Дж; и др. (май 2004 г.). «Происхождение, распространение и дифференциация гаплогрупп E и J Y-хромосомы: выводы о неолитизации Европы и более поздних миграционных событиях в Средиземноморье» . Являюсь. Дж. Хум. Жене . 74 (5): 1023–34. дои : 10.1086/386295 . ПМК 1181965 . ПМИД 15069642 .
- ^ Перейти обратно: а б с д и ж г Тромбетта, Бениамино; д'Атанасио, Евгения; Домохозяйка Андреа; Ипполити, Марко; Коппа, Альфредо; Кандилио, Франческа; Койя, Валентина; Руссо, Джанлука; Дюгужон, Жан-Мишель; Мораль, Педро; Акар, Неджат; Селитто, Даниэле; Валезини, Гвидо; Новеллетто, Андреа; Скоццари, Розария; Кручиани, Фульвио (2015). «Филогеографическое уточнение и крупномасштабное генотипирование гаплогрупп человека и хромосом и новое понимание расселения ранних скотоводов на африканском континенте» . Геномная биология и эволюция . 7 (7): 1940–50. дои : 10.1093/gbe/evv118 . ПМЦ 4524485 . ПМИД 26108492 .
- ^ Перейти обратно: а б Ши, Хун; Ци, Сюэбин; Чжун, Хуа; Пэн, Йи; Чжан, Сяомин; Ма, Ранлин З.; Су, Бинг (2013). «Генетические доказательства восточноазиатского происхождения и палеолитической миграции на север гаплогруппы N Y-хромосомы» . ПЛОС ОДИН . 8 (6): е66102. Бибкод : 2013PLoSO...866102S . дои : 10.1371/journal.pone.0066102 . ПМЦ 3688714 . ПМИД 23840409 .
- ^ Батини, Кьяра; Халласт, Пилле; Задик, Дэниел; Дельсер, Пьерпаоло Майзано; Бенаццо, Андреа; Гиротто, Сильвия; Арройо-Пардо, Эдуардо; Каваллери, Джанпьеро Л.; Де Книфф, Питер; Дюпюи, Берит Мире; Эриксен, Хайди А.; Кинг, Тури Э.; Де Мунаин, Адольфо Лопес; Лопес-Парра, Ана М.; Лутрадис, Афродита; Миласин, Елена; Новеллетто, Андреа; Памджав, Хоролма; Саджантила, Антти; Толун, Аслихан; Винни, Брюс; Джоблинг, Марк А. (2015). «Крупномасштабное недавнее расширение европейских отцов, продемонстрированное изменением последовательности населения» . Природные коммуникации . 6 : 7152. Бибкод : 2015NatCo...6.7152B . дои : 10.1038/ncomms8152 . ПМЦ 4441248 . ПМИД 25988751 .
- ^ Перейти обратно: а б Роотси, Сиири; и др. (2004). «Филогеография гаплогруппы I Y-хромосомы раскрывает различные области доисторического потока генов в Европе» (PDF) . Американский журнал генетики человека . 75 (1): 128–37. дои : 10.1086/422196 . ПМК 1181996 . ПМИД 15162323 . Архивировано из оригинала (PDF) 19 июня 2009 г. Проверено 4 мая 2016 г.
- ^ Перейти обратно: а б П. Андерхилл, Н. М. Майрес, С. Рутси, Ч. Т. Чоу, А. А. Лин, Р. П. Отиллар, Р. Кинг, Л. А. Животовский, О. Балановский, А. Пшеничнов, К. Х. Ричи, Л. Л. Кавалли-Сфорца, Т. Кивисилд, Р. Виллемс, С.Р. Вудворд, Новые филогенетические отношения для гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории, в П. Мелларсе, К. Бойле, О. Бар-Йосефе и К. Стрингере (ред.), Переосмысление эволюции человека (2007), стр. 33–42.
- ^ Перейти обратно: а б Шарма, Сваркар; Рай, Экта; Шарма, Притхвирадж; Йена, Мамата; Сингх, Света; Дарвиши, Катаюн; Бхат, Аудеш К.; Бханвер, Эй Джей С.; Тивари, Прамод Кумар; Бамезай, Рамешвар Н.К. (2009). «Индийское происхождение отцовской гаплогруппы R1a1* обосновывает автохтонное происхождение брахманов и кастовой системы» . Журнал генетики человека . 54 (1): 47–55. дои : 10.1038/jhg.2008.2 . ПМИД 19158816 .
- ^ фтДНК
- ^ Майрес2010
- ^ Джонс, Эппи Р.; Гонсалес-Фортес, Глория; Коннелл, Сара; Сиска, Вероника; Эрикссон, Андерс; Мартиниано, Руи; Маклафлин, Рассел Л.; Гальего Льоренте, Маркос; Кэссиди, Лара М.; Гамба, Кристина; Мешвелиани, Тенгиз; Бар-Йосеф, Офер; Мюллер, Вернер; Белфер-Коэн, Анна; Мацкевич, Зиновий; Джакели, Нино; Хайэм, Томас Ф.Г.; Куррат, Матиас; Лордкипанидзе, Давид; Хофрейтер, Михаэль; Маника, Андреа; Пинхаси, Рон; Брэдли, Дэниел Г. (2015). «Геномы верхнего палеолита раскрывают глубокие корни современных евразийцев» . Природные коммуникации . 6 : 8912. Бибкод : 2015NatCo...6.8912J . дои : 10.1038/ncomms9912 . ПМК 4660371 . ПМИД 26567969 .
- ^ Перейти обратно: а б «Мезолитическая ДНК Западной Евразии» . Путешествия предков . Архивировано из оригинала 7 апреля 2016 г. Проверено 4 мая 2016 г.
- ^ Перейти обратно: а б с д и Кручиани (2007), «Отслеживание прошлых перемещений мужчин среди людей в Северной / Восточной Африке и Западной Евразии: новые подсказки по гаплогруппам Y-хромосомы E-M78 и J-M12», Molecular Biology and Evolution , 24 (6): 1300–11, doi : 10.1093/molbev/msm049 , PMID 17351267. Также см. дополнительные данные , заархивированные 5 декабря 2012 г. на archive.today.
- ^ Эватт, Дэнни (1 ноября 2013 г.). «Клан Эватт: Всемирный исторический обзор фамилии семьи Эватт» .
- ^ Сьёдин, Пер; Франсуа, Оливье (2011). «Модели волны прогресса распространения гаплогруппы R1b1b2 y-хромосомы в Европе» . ПЛОС ОДИН . 6 (6): e21592. Бибкод : 2011PLoSO...621592S . дои : 10.1371/journal.pone.0021592 . ПМЦ 3123369 . ПМИД 21720564 .
- ^ Перейти обратно: а б Андерхилл, Питер А.; Майрес, Натали М.; Рутси, Сирия; Мецпалу, Мейт; Животовский Лев А.; Кинг, Рой Дж.; Лин, Алиса А.; Чоу, Шерил-Эмилиан Т.; Семино, Орнелла; Баттл, Винсент; Кутуев, Ильдус; Ярве, Мари; Чаубей, Гьянешвар; Аюб, Касим; Мохьюддин, Аиша; Мехди, С. Касим; Сенгупта, Сангамитра; Рогаев Евгений Иванович; Хуснутдинова Эльза К.; Пшеничнов Андрей; Балановский Олег; Балановская, Елена; Джеран, Нина; Огюстен, Дубравка Гавас; Бальдович, Мариан; Эррера, Рене Дж.; Тангарадж, Кумарасами; Сингх, Виджай; Сингх, Лалджи; и др. (2010). «Разделение постледникового происхождения европейских и азиатских y-хромосом внутри гаплогруппы R1a» . Европейский журнал генетики человека . 18 (4): 479–84. дои : 10.1038/ejhg.2009.194 . ПМЦ 2987245 . ПМИД 19888303 .
- ^ Филогенетическое древо Y-хромосомы, 2005 г. , с сайта FamilyTreeDNA.com.
- ^ Система номенклатуры Древа гаплогрупп Y-хромосомы человека , Genome.org
Дальнейшее чтение
[ редактировать ]- Мендес, Фернандо; Кран, Томас; Шрак, Бонни; Кран, Астрид-Мария; Вирама, Кришна; Вернер, август; Фомине, Форка Лейпей Мэтью; Брэдман, Нил; Томас, Марк; Карафет Татьяна М.; Хаммер, Майкл Ф. (7 марта 2013 г.). «Афроамериканская отцовская линия добавляет чрезвычайно древний корень к филогенетическому дереву Y-хромосомы человека» (PDF) . Американский журнал генетики человека . 92 (3): 454–59. дои : 10.1016/j.ajhg.2013.02.002 . ПМК 3591855 . ПМИД 23453668 . Архивировано из оригинала (PDF) 24 сентября 2019 года . Проверено 31 марта 2013 г.
- «Y-гаплогруппа А Филогенетическое древо» . Март 2013 г. Архивировано из оригинала 18 августа 2018 г. Проверено 30 марта 2013 г. (диаграмма, показывающая новые ветви, добавленные в филодерево A в марте 2013 г.)
Внешние ссылки
[ редактировать ]- Дерево гаплогрупп ISOGG Y-ДНК
- Генеалогическое древо ДНК Public Haplotree
- График скорости различных мутаций STR в Y-хромосоме
- Карта Y-гаплогрупп
- Атлас человеческого пути , Генографический проект , National Geographic.
- Карта Y-гаплогруппы DNA Heritage. Архивировано 18 февраля 2006 г. в Wayback Machine.
- Видеоурок по выявлению отцовского происхождения с помощью Y-хромосомы
- Предсказатель гаплогруппы
- Семино О., Пассарино Г., Офнер П.Дж. и др. (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука . 290 (5494): 1155–59. Бибкод : 2000Sci...290.1155S . дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 . В формате PDF- документа, в котором определены гаплогруппы «Eu».
- Гаплогруппа Y-ДНК и проекты субкладов
- Описания, проекты и ссылки гаплогруппы YDNA Кершнера
- Сравнительная таблица маркеров STR компании по тестированию Y-ДНК
- Y-ДНК Этнографический и генографический атлас и сборник данных из открытых источников
- Консорциум Y-хромосомы