Гаплогруппа G-M201
| Гаплогруппа G | |
|---|---|
 | |
| Возможное время происхождения | 48 500 лет назад [ 1 ] |
| Возраст слияния | 25 200 лет назад [ 1 ] |
| Возможное место происхождения | Западная Азия |
| Предок | Гаплогруппа GHIJK |
| Потомки | первичный: G1 , G2 |
| Определение мутаций | M201, PF2957, L116, L154, L204, L240, L269, L402, L520, L521, L522, L523, L605, L769, L770, L836, L837, M201, P257/U6, Страница94/U17, U2, U3, U7, U12, U20, U21, U23, U33 |
Гаплогруппа G ( M201 человека Y-хромосомы ) — гаплогруппа . Это одна из двух ветвей родительской гаплогруппы GHIJK , другая — HIJK .
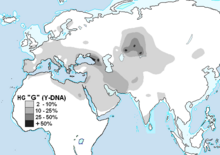
G-M201 чаще всего встречается среди различных этнических групп Кавказа , но также широко распространен с низкой частотой среди этнических групп по всей Европе , Западной Азии , Южной Азии , Центральной Азии и Северной Африке .
Наиболее часто встречающимися субкладами являются G1* (M285) и многие субклады G2 (G-P287), особенно: G2a (P15), G2a1 (G-FGC7535, ранее G-L293), G2a2b2a (G-P303), ранее G2a3b1). ; G2a2b1 (G-M406) ранее G2a3a; G2a2b2a1 (G-L140) ранее G2a3b1a; G2a2b2a1a1b (G-L497) ранее G2a3b1a2; G2a2b2a1a1a1 (G-L13) ранее G2a3b1a1a; G2a2b2a1a1c1a (G-CTS5990 или G-Z1903) ранее G2a3b1a3; G2b (G-M3115) и; G2b1 (G-M377), ранее G2b.
Происхождение
[ редактировать ]Ранее Национальное географическое общество относило свое происхождение к Ближнему Востоку 30 000 лет назад и предполагало, что люди, несущие гаплогруппу, принимали участие в распространении эпохи неолита . [ 2 ] Две научные статьи также предполагают происхождение на Ближнем Востоке, хотя и различаются по дате. Семино и др. (2000) предположили, что это произошло 17 000 лет назад. [ 3 ] Чинниоглу и др. (2004) предположили, что мутация произошла всего 9500 лет назад. [ 4 ]
Для происхождения G-M201 были предложены различные предполагаемые даты и места, большинство из которых находятся в Западной Азии . [ 5 ] Упоминалось также более восточное происхождение, которое, как полагают некоторые, происходит из района Ближнего Востока, недалеко от предгорий Гималаев. [ 6 ] Тем не менее, учитывая наибольшее разнообразие базальных ветвей, в статье Siiri Rootsi et al. предположил, что: «По нашим оценкам, географическое происхождение гаплогруппы G, вероятно, находится где-то недалеко от восточной Анатолии , Армении или западного Ирана ». [ 7 ]
Структура
[ редактировать ]Г* (М201)
- Г1 (М285, М342)
- Г1а (П20.1, П20.2, П20.3)
- Г1а1 (Л201, Л202, Л203)
- G1b (L830, L831, L832, L834, L835)
- Г1а (П20.1, П20.2, П20.3)
- G2 (P287)
- G2a (P15, U5, L31/S149, L149)
- G2a1(L293 ^ )
- G2a1a (П16.1, П16.2)
- Г2а1а1 (П18.1, П18.2, П18.3)
- G2a1a (П16.1, П16.2)
- G2a2(L223 ^ )
- G2a2a (M286)
- G2a2b(L91)
- G2a2b1 (L166, L167)
- G2a3 (L30/S126, L32/S148, L190/M485)
- G2a3a(M406)
- G2a3a1 (L14/Стр.57/S130/U16, L90/S133)
- G2a3a2 (L645)
- G2a3b (L141.1)
- G2a3b1 (P303/S135)
- G2a3b1a (L140)
- G2a3b1a1 (U1)
- G2a3b1a1a (L13/S131/U13, L78/M527)
- G2a3b1a1a1 (L1263 ^ )
- G2a3b1a1b (L1266 ^ )
- G2a3b1a1b1 (L1264 ^ , L1265 ^ , L1268 ^ )
- G2a3b1a1a (L13/S131/U13, L78/M527)
- G2a3b1a2 (L497, L353.1 ^ , L353.2 ^ )
- G2a3b1a2a(Z725 ^ )
- G2a3b1a2a1 (L43/S147)
- G2a3b1a2a1a (L42/S146)
- G2a3b1a2a1 (L43/S147)
- G2a3b1a2a(Z725 ^ )
- G2a3b1a3(Z1903 ^ )
- G2a3b1a3a (Z724 ^ )
- G2a3b1a3a1 (L640)
- G2a3b1a3a (Z724 ^ )
- G2a3b1a4 (Л660, Л662)
- G2a3b1a1 (U1)
- G2a3b1b (L694)
- G2a3b1a (L140)
- G2a3b2 (Л177.1, Л177.2, Л177.3)
- G2a3b1 (P303/S135)
- G2a3a(M406)
- G2a1(L293 ^ )
- G2b (M377, L72, L183)
- G2b1 (M283)
- G2a (P15, U5, L31/S149, L149)
(Здесь субклады соответствуют определениям SNP Y-ДНК, используемым ISOGG. В 2012 году несколько категорий, обнаруженных только у одного человека в ходе научных исследований, были удалены из дерева ISOGG, что привело к некоторому переименованию.)
Географическое распространение
[ редактировать ]Доисторическое и древнее присутствие
[ редактировать ]Древние G-M201 с секвенированием [ самостоятельно опубликованный источник? ] Гаплогруппа G2a (G-P15) была идентифицирована в неолитических человеческих останках в Европе, датируемых между 5000 и 3000 годами до нашей эры. Эти неолитические европейцы были потомками неолитических фермеров из Анатолии, одних из первых народов в мире, занимавшихся сельским хозяйством. G-M201 также был обнаружен в неолитических анатолийских стоянках, таких как Бончуклу, датируемый 8300-7600 годами до нашей эры, и Барцин, датируемый 6419-6238 годами до нашей эры. [ 8 ] [ 9 ]
Более того, большинство всех мужских скелетов периода европейского неолита до сих пор содержали Y-ДНК, принадлежащую этой гаплогруппе. Пять самых старых скелетов, подтвержденных древним тестированием ДНК , содержащих гаплогруппу G2a, были найдены в Авелланер пещерном захоронении , недалеко от Ле-План-д'Остолес , в Каталонии , Испания , и были датированы радиоуглеродным анализом примерно 5000 г. до н.э. [ 10 ]
Скелет, найденный на неолитическом кладбище, известном как Деренбург Мееренштиг II , в Саксонии-Анхальт, Германия , по-видимому, принадлежал G2a3 (G-S126) или субкладу. Он был найден вместе с погребальными артефактами, принадлежащими Linearbandkeramische Kultur (« Линейно-ленточная керамическая культура »; ЛБК). Этот скелет не мог быть датирован радиоуглеродным анализом, но возраст других скелетов был от 5100 до 6100 лет. Наиболее подробной идентифицированной мутацией SNP была S126 (L30), которая определяет G2a3. [ 11 ]
G2a был также обнаружен в 20 из 22 образцов древней Y-ДНК из Трейля , типового места группы фермеров позднего неолита на юге Франции , датируемого примерно 5000 лет назад. [ 12 ] Четвертое место, также относящееся к тому же периоду, — это Эцтальские Альпы мумифицированные останки Эци-Ледяного человека , где были обнаружены . Ледяной человек принадлежит к гаплогруппе G2a2b. [ 13 ] (ранее назывался G2a4). Гаплогруппа G2a2b сегодня является редкой группой в Европе. Авторы испанского исследования указали, что мужчины из Авелланера имели редкие значения маркеров при тестировании их маркеров коротких тандемных повторов (STR).
Во времена энеолита гаплогруппа G считалась повсеместно распространенной в Анатолии, составляя значительное количество местных гаплогрупп Y-ДНК, наряду с гаплогруппой J. Она оставалась распространенной на протяжении всего энеолита, бронзы и доримского периода. [ 14 ] В статье 2004 года была обнаружена значительная корреляция между хаттской и каскской культурами с наличием гаплогруппы G, однако было отмечено, что более высокие различия субклада G2-P15 существуют в направлении западной Анатолии. [ 15 ]
Современное присутствие
[ редактировать ]В России, Украине и Центральной Азии представители различных этнических меньшинств и/или жители определенных населенных пунктов обладают самым высоким уровнем G-M201 в мире, хотя средний уровень на национальном уровне составляет около 1% или меньше. Казахстане Было обнаружено , обладают самым что племена (или кланы) Маджар и Аргын в высоким уровнем G-M201 среди любой современной этнической группы. Среди маджаров G1 встречался в 87%. Отдельное исследование аргынов показало, что 71% мужчин принадлежат к G1. На российском Северном Кавказе кабардинское осетинское и население также отличается высоким уровнем G-M201. Дигора , Северная Осетия, имеет самую высокую известную концентрацию G в одном городе: 74% протестированных мужчин были G. [ 16 ] Гаплогруппа G встречается на востоке, вплоть до северного Китая, в небольших процентах, где G может достигать более значительных процентов в группах меньшинств, таких как уйгуры.
В Турции, на Южном Кавказе и в Иране гаплогруппа G достигает самого высокого процента населения страны. Среди турецких мужчин 11% населения составляют Г. [ 4 ] В Иране гаплогруппа G достигает от 13 до 15% населения в различных частях страны. Хотя он встречается в процентах выше 10% среди бахтиаров , талышей , гилаки , мазандарани и иранских азербайджанцев , он ближе к 5% среди иранских арабов и в некоторых крупных городах. [ 17 ] Среди образцов в базе данных YHRD из стран Южного Кавказа 29% образцов из Абазинии , 31% из Грузии , 18% из Азербайджана и 11% из Армении относятся к образцам G.
В Европе к западу от Черного моря гаплогруппа G встречается в среднем примерно у 5% населения на большей части континента. [ 18 ] Концентрация G падает ниже этого среднего значения в Скандинавии , самых западных бывших советских республиках и Польше , а также в Исландии и на Британских островах . В Европе, похоже, существуют очаги необычной концентрации. В Уэльсе доминирует особый тип G2a3b1 (DYS388=13 и DYS594=11), что приводит к тому, что процент G среди населения выше, чем в Англии.
В Тироле (Тироле) западной Австрии процентное содержание G-M201 может достигать 40% и более; пожалуй, самым известным примером являются древние останки так называемого «Ледяного человека» Эци . В одном исследовании в северных и горных районах острова Сардиния у западной Италии процент G достигает 11% населения. [ 19 ] и, по данным другого исследования, достиг 21% в городе Темпио . На греческом острове Крит примерно 7% [ 20 ] до 11% [ 21 ] мужчин принадлежат к гаплогруппе G. На северо-востоке Хорватии , в городе Осиек , G обнаружен у 14% лиц мужского пола. [ 22 ] Город расположен на берегу реки Драва , которая, в частности, начинается в Тироле/Тирольском регионе Альп, еще одном фокусе гаплогруппы G в Европе. Дальше на север было обнаружено, что 8% этнических венгерских мужчин и 5,1% этнических богемских (чехских) мужчин принадлежат к гаплогруппе G.
В Южной Азии гаплогруппа G встречается редко. [ 23 ] [ 24 ] Некоторые этнические меньшинства обладают им в значительной концентрации, в том числе примерно 18%. [ 25 ] до 20% [ 26 ] Калаша , примерно 16 % Брауи , [ 26 ] и примерно 11,5% опрошенных пуштунов , [ 25 ] все они родом из самых восточных регионов Иранского нагорья , хотя он встречается только примерно у 3% общего населения Пакистана . [ 25 ] В исследовании 936 индийцев гаплогруппа G составляла менее 1% выборки и полностью отсутствовала в протестированной северо-западной индийской популяции. [ 27 ] В одном исследовании около 6% образцов из Шри-Ланки и Малайзии были отнесены к гаплогруппе G, но ни один из них не был обнаружен в других прибрежных землях Индийского или Тихого океана в Азии. [ 28 ] G-M201 был описан как «почти отсутствующий». [ 29 ] и «практически отсутствует» в Индии, причем субклад G2a-P15, в частности, считается незначительным, что указывает на уникальные события распространения из Западной Азии. [ 30 ] [ 31 ]
На Ближнем Востоке гаплогруппа G составляет около 3% населения практически во всех областях. [ 32 ] [ 33 ] Среди друзов, преимущественно жителей Израиля, у 10% обнаружена гаплогруппа G. [ 34 ]
Около 10% еврейских мужчин относятся к гаплогруппе G. [ нужна ссылка ]
В Африке гаплогруппа G редко встречается в странах Африки к югу от Сахары или к югу от Африканского Рога среди коренного населения. В Египте исследования предоставили информацию, согласно которой процент G составляет от 2% до 9%. [ 35 ] [ 36 ] [ 37 ] 3% североафриканских берберов относятся к гаплогруппе G. Было обнаружено, что [ 38 ] 2% марокканцев- арабов и 0,8% марокканцев-берберов также оказались G. [ 39 ]
В Америке процент гаплогруппы G соответствует числу эмигрировавших из стран Старого Света. Он не встречается среди коренных американцев, за исключением случаев, когда имели место смешанные браки с инородцами. Он был обнаружен у мексиканских метисов.
G1 (М285 или М342)
[ редактировать ]Почти все люди с гаплогруппой G1 имеют значение 12 по маркеру короткого тандемного повтора (STR) DYS392, и все будут иметь мутацию SNP M285 или M342 , которая характеризует эту группу. Это значение 12 редко встречается в других категориях G, кроме G1.
существуют субклады G1a, G1a1, G1b.
Самая высокая зарегистрированная концентрация G1 и его субкладов в одной стране наблюдается в Иране , за ней следуют соседние страны на западе.
Существуют отличительные ашкеназские еврейские и казахские субклады, основанные на комбинациях значений маркеров STR.
G2 (P287)
[ редактировать ]
Мужчины, принадлежащие к этой группе, но отрицательные по всем субкладам G2, представляют небольшое количество мужчин гаплогруппы G. P287 был идентифицирован в Университете Аризоны и стал широко известен в конце 2007 года. Его идентификация привела к значительному переименованию категорий G.
G2a (P15)
[ редактировать ]Мужчины с гаплогруппой G, принадлежащие к этой группе, но отрицательные для всех субкладов G2a, редко встречаются в Европе, но могут представлять собой значительную группу в пока плохо исследованных районах к востоку от Турции. P15 был идентифицирован в Университете Аризоны и стал широко известен к 2002 году. Его хромосомная локализация указана как 21653414. G2a был обнаружен в средневековых останках в высокостатусной гробнице VII века нашей эры в Эргольдинге, Бавария , Германия, но субклады G2a не были обнаружены. протестировано. [ 40 ]
Существует несколько SNP, которые на данный момент имеют такое же покрытие, как P15. Это — с сопутствующими местоположениями Y-хромосомы — U5 (rs2178500), L149 (8486380) и L31 (также называемый S149) (rs35617575..12538148). Если какой-либо мужчина с мутацией P15 окажется отрицательным (наследственным) на любой из этих признаков или наоборот, это открытие станет основой новой категории G2a.
G2a1 (FGC7535)
[ редактировать ]Гаплогруппа G2a1 (также известная как G-FGC753, а ранее как G-L293) и ее субклады представляют большинство образцов гаплогруппы G в некоторых частях Кавказских гор . В других местах они встречаются лишь в небольшом количестве. На данный момент все люди G2a1 имеют значение 10 по STR -маркеру DYS392. Люди G2a1a также обычно имеют более высокие значения DYS385b, например 16, 17 или 18, чем у большинства людей G.
Судя по имеющимся образцам, северные осетины среднего Северного Кавказа в России в подавляющем большинстве принадлежат к субкладу G2a1. Южные осетины и сваны , как правило, к югу от Северной Осетии, имеют значительное количество людей G2a1, но процентная доля населения еще не указана.
Наличие мутации SNP P18 характеризует единственный субклад G2a1a, G2a1a. Надежность как P16, так и P18 при идентификации всех лиц в каждой из этих категорий была поставлена под сомнение, и необходимо изучить отдельные компоненты SNP.
Мужчины -ашкенази- евреи G2a1a северо-восточного европейского происхождения образуют отдельный кластер на основе значений маркера STR. Мужчины с Кавказа и мужчины из Восточной Европы также образуют характерные кластеры СПО.
G2a2a(PF3147)
[ редактировать ]G-PF3147 (ранее G-L223 и G-PF3146) характеризуется наличием мутации L223. L223 обнаружен на Y-хромосоме по адресам rs810801 и 6405148 с мутацией от C до G. L223 был впервые идентифицирован в образцах 23andMe в 2009 году, но оказался проблематичным в качестве индивидуального теста, первые успешные результаты были опубликованы в Family Tree DNA в конце 2011 года. под присвоенным ему ярлыком L223. Затем выяснилось, что к L223 относятся несколько субкладов, в том числе:
G-L91 был идентифицирован в 2009 году. В его состав входят « Эци », [ 41 ] так называемый Ледяной человек, умерший по крайней мере 5000 лет назад в Европейских Альпах. G-L91, по-видимому, охватывает значительную часть мужчин, принадлежащих к G. L91 до сих пор встречается в разбросанных частях Европы и Северной Африки, а также в Армении . В группу G-L91 включены некоторые мужчины с двойными значениями STR-маркера DYS19, но есть также мужчины G2a2 с этим показателем, которые не относятся к L91+. Ситуация с двойным значением 19 не наблюдается в субкладах G2a1 и G2a3. Мутация L91 обнаружена по адресам 21327383 и rs35474563 на Y-хромосоме. Передний праймер GTATTGAACTTACAATTCACGTCCC , и обратное CTCTCCAAATCGGGTTTCCT . Мутация включает изменение C на T. [ 42 ] L223 находится на Y-хромосоме по адресу rs13304806.
Субклад G-M286 (M286+) небольшой по сравнению с G-L91. Образцы были обнаружены в Англии, Германии, Черногории (боснийцы), Испании, Кипре (греки), Турции, Армении, Грузии, Ливане, Сирии и Кувейте. Британские образцы во многих случаях имеют противоречивые двойные значения STR-маркера DYS19. M286 был впервые идентифицирован в Стэнфордском университете в положении хромосомы 21151187 и представляет собой мутацию от G к A.
SNP L293, характеризующий третий субклад, был идентифицирован в июне 2010 года в ДНК Семейного древа. Он охватывает небольшую группу латиноамериканских мужчин, которые также на данный момент имеют нечетное значение 13,21 по маркеру YCA. Мутация обнаружена в Y-хромосоме по адресу 10595022 и представляет собой изменение с G на C.
G2a2b (L30, PF3267, S126, U8)
[ редактировать ]G-L30 (также G-PF3267, G-S126 или G-U8; G2a2b, ранее G2a3) Мужчины, принадлежащие к этой группе, но отрицательные ко всем ее субкладам, сегодня составляют небольшое число. Эта гаплогруппа была обнаружена в неолитическом скелете примерно 5000 г. до н.э. на кладбище Деренбург-Мееренштиг II в Германии, которое является частью культуры линейной керамики , известной на немецком языке как Linearbandkeramik (LBK). [ 11 ] но не тестировался на субклады G2a3.
G2a2b1 (M406)
[ редактировать ]G-M406* (G2a2b1*; ранее G2a3a*) и его субклады, по-видимому, чаще всего встречаются в Турции и прибрежных районах восточного Средиземноморья, где они могут составлять до 5% всех марок и 50% образцов гаплогруппы G. G2a2b1 чаще встречается в Южной Европе, чем в Северной Европе. В Европе, за исключением Италии , G2a2b1 составляет менее 20% образцов G. G2a2b1 до сих пор редко появлялся в Северной Африке или Южной Азии, но представляет собой небольшой процент популяции G в регионе Кавказских гор и в Иране .
Относительно высокий процент людей G2a2b1 имеет значение 21 по STR-маркеру DYS390. Маркер DYS391 в основном имеет значение 10, но иногда и 11 у лиц G2a2b1, а DYS392 почти всегда равен 11. Если образец соответствует критериям, указанным для этих трех маркеров, вполне вероятно, что это образец G2a2b1.
G2a2b2 (CTS2488)
[ редактировать ]G-CTS2488 или G2a2b2 (также известный как G-L141.1; ранее G-141 и G2a3b) был идентифицирован только в середине 2009 года в Family Tree DNA . Почти все мужчины L141 принадлежат к субкладам L141. Были идентифицированы образцы от людей с Британскими островами, сицилийского и турецкого происхождения. Лица с L141, которые на данный момент не принадлежат ни к одному субкладу L141, имеют значение 11 по STR-маркеру DYS490 — находка, редкая для других категорий G. Мутация L141 обнаружена на Y-хромосоме по адресу 2948607. Мутация L141 включает вставку. [ 43 ]
G2a2b2a (G-P303)
[ редактировать ]G-P303*, также известный как G2a2b2a* (ранее G2a3b1*), и его субклады в настоящее время сконцентрированы на юге России и Кавказе , а также, на более низких уровнях, в других частях Европы и Юго-Западной Азии , особенно на территории, включающей Турция, Иран и Ближний Восток, где мог возникнуть G2a2b2a. G2a2b2a также встречается в Индии.
Большинство представителей G-P303 принадлежат к одному из его субкладов, а не к G-P303*.
Самый крупный субклад G-P303*, основанный на доступных образцах, - это тот, в котором почти все люди имеют значение 13 по STR-маркеру DYS388. SNP L497 включает этих мужчин, но большинство мужчин G-L497 принадлежат к субкладу G-Z725, также известному как G-DYS388=13. Существуют дополнительные субклады мужчин с DYS388=13, характеризующиеся наличием специфических SNP или необычных странностей маркеров STR. Члены этой группы были обнаружены в Европе и на Ближнем Востоке . [ 7 ]
Следующий по величине субклад G-P303 характеризуется наличием мутации U1. Но высокий процент мужчин U1 принадлежит к двум его субкладам: G-L13/S13 и Z1266 (G2a3b1a1b). Субклад G-L13 наиболее распространен на севере центральной Европы, а G-Z1266 наиболее распространен в горах западного Кавказа.
Последний основной субклад характеризуется наличием SNP Z1903 и значением 9 по маркеру DYS568. Большой процент мужчин G-Z1903 принадлежит к субкладу G-Z724.
Самый высокий процент лиц G-P303 в описанной на данный момент отдельной популяции наблюдается на острове Ибица у восточного побережья Испании. Эта группа была связана с криптоеврейским населением, бежавшим на остров во времена испанской инквизиции , значительная часть которого идентифицируется как G-Z725 (DYS388=13). [ 44 ]
G2a2b2b (PF3359)
[ редактировать ]G-PF3359 (или G2a2b2b; ранее G2a3b2) до 2013 года был известен как G-L177. в качестве идентификатора SNP L177 (также известный как L1771.1/L177_1, L1771.2/L177_2, L177.3/L177_3) был отозван ISOGG в 2013 году, после того как он «был признан ненадежным палиндромным snp». [ 5 ] [ 45 ]
Древняя ДНК, идентифицированная как G-PF3359, была обнаружена на археологических раскопках в: Венгрии (субклад G-F872*), датированная 7500 лет назад (BP); Венгрия (субклад G-F1193*) 7150 лет назад, и; Испания (G-PF3359*) 4700 лет назад. [ 5 ]
Членов G-PF3359, вероятно, меньше, чем мужчин, включенных в G-P303, но соответствующих мутаций было проведено лишь небольшое количество исследований. До сих пор мужчины, позитивно относящиеся к этому, имели ирландское, английское, голландское, ливанское и/или турецкое (армянская фамилия) происхождение. Вероятно, существует несколько субкладов G-PF3359, основанных на общих маркерах STR.
Количество значений маркера STR, разделяющее мужчин в этой группе, позволяет предположить, что G-PF3359 является относительно старой группой, несмотря на небольшое количество вовлеченных мужчин. [ 46 ] [ самостоятельно опубликованный источник? ] Связанные с этим мутации могут быть сложными и трудными для интерпретации.
G2b1 (M377)
[ редактировать ]G-M377, теперь также известный как G2b1, ранее обозначался G2b и G2c. Клада близкородственных евреев-ашкенази представляет практически всех людей G2b , при этом на данный момент зарегистрировано только три других гаплотипа G2b: один турок из Карса на северо-востоке Турции недалеко от Армении, один пуштун и один Бурушо в Пакистане.
Крайняя редкость G-M377 на севере Пакистана может указывать на то, что G2b в этой области берет свое начало за пределами региона и был занесен туда в исторический период, возможно, с дальнего запада (Пакистан входил как в Персидскую империю Ахеменидов , завоеванную Александром Македонским , так и в , а затем вошел в состав Греко-Бактрийского царства ). Эти два зарегистрированных пакистанских гаплотипа G-M377 весьма отличаются от еврейской клады ашкенази и, следовательно, совсем не указывают на недавнее общее происхождение. Турецкий G-M377 несколько ближе, но не идентичен. Еще неизвестно, выявит ли тестирование гаплотипы G-M377 в других популяциях — это является некоторым показателем того, что G-M377 встречается на низких уровнях на Ближнем Востоке.
Все протестированные на данный момент мужчины G-M377 также имеют редкое нулевое значение для DYS425 маркера (отсутствующий аллель «T» палиндромного STR DYS371 ), результат события RecLOH , обнаружение, которое еще не наблюдается среди большинства других гаплотипов G. . По данным одного исследования, среди евреев Израиля, прибывших из многих уголков мира, G-M377 составлял 3,7%. [ 47 ]
Гаплогруппа G-M377 была обнаружена с частотой 60% из выборки пяти пуштунов в регионе Вардак в Афганистане. Вероятно, это связано с эффектом местного основателя. [ 48 ]
Определение SNP
[ редактировать ]Международное общество генетической генеалогии (ISOGG) поддерживает самую актуальную согласованную версию категорий гаплогрупп. [ 49 ] Эти классификации основаны на общих мутациях SNP. Открытие новых SNP может привести к присвоению новых названий категориям гаплогрупп. До 2008 года существовало всего несколько категорий G, когда в категории были внесены серьезные изменения. В 2009–2012 годах было идентифицировано еще больше G-SNP, что привело к еще большим изменениям. До 2008 года о новых G SNP сообщалось из лабораторий Университета Аризоны (обозначения P), Стэнфордского университета (обозначения M) или Университета Центральной Флориды (обозначения U). Начиная с 2008 года, дополнительные G SNP были идентифицированы в генеалогическом древе ДНК (обозначения L) и этнородословной (обозначения S). Эти последние лаборатории также использовали результаты необработанных данных, предоставленные людьми, протестировавшими около 2000 SNP в 23andMe , чтобы предоставить новые тесты SNP, обозначенные L или S. В 2009–2010 годах проект «Прогулка по Y» генеалогического древа ДНК, секвенировавший определенные сегменты Y-хромосомы, предоставил ряд новых G SNP с обозначением L. В 2012 году SNP с обозначением Z, впервые обнаруженные гражданскими исследователями из проекта «1000 геномов» Начали появляться данные .
Поскольку SNP обеспечивают наиболее надежный метод категоризации, каждый из них может представлять официальную категорию G. Категории имеют чередующиеся буквы и цифры. Но необычные значения или необычные комбинации значений, обнаруженные в маркерах коротких тандемных повторов (STR), также могут стать основой для дополнительной таксономизации. Идентификация нового SNP может вызвать необходимость переименования одной или нескольких категорий.
Мутация SNP M201, характеризующая гаплогруппу G, была выявлена в Стэнфордском университете и впервые о ней сообщалось в 2001 году. [ 50 ] Технические характеристики M201 приведены в следующем виде: refSNPid — rs2032636..... Расположение Y-хромосомы 13536923.... прямой праймер — tatgcatttgttgagtatatgtc .....обратный праймер gttctgaatgaaagttcaaacg .....мутация включает изменение G на T.
Был выявлен ряд SNP , по-видимому, с таким же охватом населения, как и M201. Поскольку M201 был идентифицирован первым, это стандартный тест SNP, используемый при тестировании лиц группы G. Чтобы определить, представляет ли один из этих альтернативных SNP субклад M201, альтернативные SNP должны быть протестированы на людях G, которые отрицательны для известных субкладов G. В такой категории имеется лишь небольшое количество людей, и только небольшое количество людей было проверено на наличие G-эквивалентных SNP, отличных от M201.
Следующие SNP на данный момент идентифицированы как эквиваленты M201: L116, L154, L269, L294, L240, P257, L402, L520, L521, L522, L523, L605, страница 94, U2, U3, U6, U7, U12, U17, U20, U21, U23 и У33. P257 впервые был зарегистрирован в 2008 году. [ 51 ] L240 был идентифицирован в 2009 году. [ 52 ] SNP «U» были идентифицированы в 2006 году, но опубликованы только в 2009 году. [ 53 ]
Кроме того, существует множество других SNP, которые, как полагают, имеют такое же покрытие, как M201. Они находятся по адресам: rs9786910, rs9786537, rs2713254, rs35567891 и rs34621155 на Y-хромосоме. Ни одна лаборатория еще не присвоила им сокращенные названия.
Выдающиеся члены G
[ редактировать ]- Человек, известный как « Эци », чьи останки, датируемые 4-м тысячелетием до нашей эры , были найдены на современной границе между Австрией и Италией, является членом G-L91 (G2a2b).
- Многие члены так называемого родства Буре , клановой группы, которая ведет свое происхождение от человека, родившегося в Швеции в конце 14 или начале 15 века, как сообщается, принадлежат к G-Y12970 (также известному как G-Y12971 и G-Y12970). Z30735), также известный как G2a2b2a1a1b1a1a2a1c2a~ (по данным 2017 г.) Дерево ISOGG ; ранее G2a3b1a2). [ 5 ]
- Король Англии Ричард III , эксгумированный в 2012 году , был членом G2 (G-P287). [ 54 ]
- Иосиф Сталин (урожденный Иосеб Бесарионис дзе Джугашвили) был, согласно генетическому тесту одного из его внуков (Александра Бурдонского), членом гаплогруппы Y-ДНК G2a1a (FGC595/Z6553). [ 5 ] [ 55 ]
- Аль Капоне был американским гангстером и бизнесменом, который, по данным Geni.com , был членом гаплогруппы Y-ДНК G-P303. [ 56 ] [ самостоятельно опубликованный источник? ]
- Юя один из Тутанхамона прадедов по материнской линии. Предсказанный гаплотип G2a на основе древних профилей Y-STR. Юя служил ключевым советником Аменхотепа III и занимал такие должности, как «Королевский лейтенант» и «Мастер лошадей»; его титул «Отец бога», возможно, относился конкретно к тому, что он был тестем Аменхотепа. В своем родном городе Ахмин Юя был пророком Мина, главного «бога» этой местности, и служил этому божеству «Управляющим скотом». [ 57 ]
- Ларри Берд - американский профессиональный баскетбольный руководитель, бывший тренер и бывший игрок «Бостон Селтикс». Было показано, что он принадлежит к гаплогруппе G-Z6748 на основании тестирования нескольких родственников, происходящих от Томаса Берда в рамках проекта гаплогруппы G-L497 Y-ДНК. . [ 58 ]
- Другие мужчины, предположительно являющиеся членами гаплогруппы G, включают: немецко-американского пионера и солдата Филиппа Хэммана , пятого правнука Филлипса Шейна Хэммана , американского тяжелоатлета-олимпийца, [ 59 ] Американский политик Линн Бэнкс , физик Джон Г. Крамер , американские актеры Джеймс Франциск и Джейк Джилленхол , американский бизнесмен Наджиб Халаби , американский юрист и администратор Ньютон Миноу , а также американский юрист и подписавший Декларацию независимости Ричард Стоктон . [ 60 ] [ самостоятельно опубликованный источник? ]
См. также
[ редактировать ]- Евреи с гаплогруппой G
- Генетическая история Европы
- Генетическая генеалогия
- Предыстория Анатолии
- Народы Кавказа
- Магдаленский
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК в населении Европы
- Гаплогруппы Y-ДНК у населения Кавказа
- Гаплогруппы Y-ДНК у населения Ближнего Востока
- Гаплогруппы Y-ДНК у населения Северной Африки
- Гаплогруппы Y-ДНК по этническим группам
Ссылки
[ редактировать ]- ^ Перейти обратно: а б "GYTree" .
- ^ «Атлас человеческого пути: гаплогруппа G (M201)» . Архивировано из оригинала 5 февраля 2011 года . Проверено 25 марта 2023 г.
- ^ Семино О, Пассарино Г, Офнер П.Дж., Лин А.А., Арбузова С., Бекман Л.Е., Де Бенедиктис Г., Франкалаччи П., Коуваци А., Лимборска С., Марсикиаэ М., Мика А., Мика Б., Приморак Д., Сантакьяра-Бенеректи А.С., Кавалли- Сфорца Л.Л., Андерхилл, Пенсильвания (ноябрь 2000 г.). «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: взгляд на Y-хромосому». Наука 290 (5494): 1155–9. Бибкод : 2000Sci... 290.1155S дои : 10.1126/science.290.5494.1155 . ПМИД 11073453 .
- ^ Перейти обратно: а б Чинниоглу С и др. (январь 2004 г.). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии». Генетика человека . 114 (2): 127–48. дои : 10.1007/s00439-003-1031-4 . ПМИД 14586639 . S2CID 10763736 .
- ^ Перейти обратно: а б с д и ISOGG, 2017, Гаплогруппа G Y-ДНК и ее субклады - 2017 (17 мая 2017 г.).
- ^ Махал, Дэвид Г.; Мацукас, Янис Г. (23 января 2018 г.). «Географическое происхождение этнических групп на Индийском субконтиненте: исследование древних следов с помощью гаплогрупп Y-ДНК» . Границы генетики . 9 :4. дои : 10.3389/fgene.2018.00004 . ПМЦ 5787057 . ПМИД 29410676 .
- ^ Перейти обратно: а б Роотси, Сиири; Майрес, Натали М; Лин, Алиса А; Ярве, Мари; Кинг, Рой Дж; Кутуев, Ильдус; Кабрера, Висенте М; Хуснутдинова, Эльза К; Варенди, Карт; Саакян, Ованес; Бехар, Дорон М; Хусаинова, Рита; Балановский Олег; Балановская, Елена; Рудан, Павао; Епископосян, Левон; Бахманимер, Ардешир; Фарджадян, Ширин; Кушнеревич Алена; Эррера, Рене Дж; Грюни, Виола; Батталья, Винченца; Ники, Кармела; Кробу, Франческа; Карачанак, Армия; Кашани, Бахарак Хушиар; Хаушманд, Масуд; Санати, Мохаммад Х; Тончева, Драга; Лиза, Антонелла; Семино, Орнелла; Кьярони, Жак; Кристофаро, Джули Ди; Виллемс, Ричард; Кивисилд, Тоомас; Андерхилл, Питер А. (16 мая 2012 г.). «Выявление совместного происхождения Y-хромосом гаплогруппы G в популяциях Европы и Кавказа» . Европейский журнал генетики человека . 20 (12): 1275–1282. дои : 10.1038/ejhg.2012.86 . ПМК 3499744 . ПМИД 22588667 .
- ^ Фельдман, Михал; Фернандес-Домингес, Ева; Рейнольдс, Люк; Бэрд, Дуглас; Пирсон, Джессика; Гершковитц, Израиль; Мэй, Хила; Геринг-Моррис, Найджел; Бенц, Мэрион; Грески, Джулия; Бьянко, Рафаэла А.; Фэйрберн, Эндрю; Мустафаоглу, Гекхан; Стокхаммер, Филипп В.; Пост, Козимо; Хаак, Вольфганг; Чон, Чунгвон; Краузе, Йоханнес (19 марта 2019 г.). «Геном человека позднего плейстоцена предполагает местное происхождение первых земледельцев Центральной Анатолии» . Природные коммуникации . 10 (1): 1218. Бибкод : 2019NatCo..10.1218F . дои : 10.1038/s41467-019-09209-7 . ПМК 6425003 . ПМИД 30890703 . S2CID 83464300 .
- ^ Хофманова, Зузана; Крейцер, Сюзанна; Хеллентал, Гарретт; Продай, Кристиан; Дикманн, Джоан; Диес-дель-Молино, Давид; Дорп, Люси ван; Лопес, Сайоа; Кусафанас, Афанасий; Линк, Вивиан; Кирсанов, Карола; Кэссиди, Лара М.; Мартиниано, Руи; Стробель, Мелани; Шой, Амели; Коцакис, Костас; Холстед, Пол; Триантафиллу, Севи; Кипарисси-Апостолика, Нина; Урем-Коцоу, Душанка-Кристина; Зиота, Кристина; Адактилу, Фотини; Гопалан, Шьямалика; Бобо, Дин М.; Винкельбах, Лаура; Блёхер, Йенс; Унтерлендер, Мартина; Лойенбергер, Кристоф; Чилингироглу, Чилер; Хорейс, Барбара; Герритсен, Фокке; Шеннан, Стивен; Брэдли, Дэниел Г.; Куррат, Матиас; Вирама, Кришна Р.; Вегманн, Дэниел; Томас, Марк Г.; Папагеоргопулу, Кристина; Бургер, Иоахим (25 ноября 2015 г.). «Ранние земледельцы со всей Европы напрямую произошли от неолитических эгейцев» (PDF) . bioRxiv : 032763. doi : 10.1101/032763 . S2CID 196682085 .
- ^ Лакан, Мари; Кейзер, Кристина; Рико, Франсуа-Ксавье; Брукато, Николас; Таррус, Хосеп; Босх, Ангел; Гилейн, Жан; Крубези, Эрик; Людес, Бертран (8 ноября 2011 г.). «Древняя ДНК позволяет предположить ведущую роль, которую играли мужчины в распространении неолита» . Труды Национальной академии наук Соединенных Штатов Америки . 108 (45): 18255–18259. Бибкод : 2011PNAS..10818255L . дои : 10.1073/pnas.1113061108 . ПМК 3215063 . ПМИД 22042855 .
- ^ Перейти обратно: а б Хаак В, Балановский О, Санчес Дж., Кошель С, Запорожченко В, Адлер С.Дж., Дер Саркисян К.С., Брандт Г., Шварц С., Никлиш Н., Дрезели В., Фрич Б., Балановска Е., Виллемс Р., Меллер Х., Альт К.В., Купер А (2010). Пенни Д. (ред.). «Древняя ДНК европейских фермеров раннего неолита раскрывает их ближневосточное родство» . ПЛОС Биология . 8 (11): е1000536. дои : 10.1371/journal.pbio.1000536 . ПМЦ 2976717 . ПМИД 21085689 .
- ^ Мари Лакан, Кристин Кейзер, Франсуа-Ксавье Рико, Николя Брукато, Фрэнсис Дюрантон, Жан Гилейн, Эрик Крубези и Бертран Люд, Древняя ДНК показывает распространение мужчин по неолитическому средиземноморскому маршруту, Труды Национальной академии наук США , онлайн 31 мая 2011 г. перед печатью.
- ^ Интервью с доктором Эдуардом Эгартер-Виглем , руководителем отдела консервации и помощником исследовательских проектов Археологического музея в Боцене. Из документального фильма: «Ötzi, ein Archäologiekrimi» [Эци, археологическое преступление] Кристины Шпрахманн. Телетрансляция по 3 субботы, 10 августа 2011 г. и br-alpha, 13 сентября 2011 г.
- ^ Лазаридис I, Алпаслан-Руденберг С, Ачар А, Ачиккол А, Агеларакис А, Агикян Л, Акюз У, Андреева Д, Андрияшевич Г, Антонович Д, Армит И, Атмаца А, Аветисян П, Айтек А.И., Бадзваров К, Бадалян Р. , Бакарджиев С, Бален Дж, Бейко Л, Бернардос Р, Берцатос А, Бибер Х, Билир А, Бодружич М, Боногофски М, Бонсалл С, Борич Д, Боровинич Н, Браво Моранте Г, Баттингер К, Каллан К, Кандилио Ф, Чарич М, Черонет О, Чохаджиев С, Човалопулу М.Е., Криссулаки С, Чобану И, Чондич Н, Константинеску М, Кристиани Е, Каллетон Б.Дж., Кертис Е, Дэвис Дж, Демченко Т.И., Дергачев В, Дерин З, Дескай С, Деведжян С , Джорджевич В, Даффет Карлсон К.С., Экклс ЛР, Эленски Н, Энгин А, Эрдоган Н, Эрир-Пазарчи С, Фернандес ДМ, Ферри М, Фрейлих С, Фринкулеса А, Галати МЛ, Гамарра Б, Гаспарян Б, Гайдарска Б, Генч Э, Гюльтекин Т, Гундуз С, Хайду Т, Хейд В, Хобосян С, Оганесян Н, Илиев И, Илиев Л, Илиев С, Ивгин И, Янкович И, Йованова Л, Карканас П, Каваз-Кындыгыли Б, Кая Э., Китинг Д, Кеннетт Д.Д., Дениз Кешичи С., Худавердян А., Кисс К., Кылыч С., Клостерманн П., Костак Бока Негра Вальдес С., Ковачевич С., Кренц-Нидбала М., Крзнарич Шкриванко М., Курти Р., Кузман П., Лоусон А.М., Лазар С. , Лештаков К., Леви Т.Е., Лирицис И., Лоренц К.О., Лукасик С., Мах М., Маллик С., Мандл К., Мартиросян-Ольшанский К., Мэтьюз Р., Мэтьюз В., Максуини К., Меликян В., Микко А., Мишель М., Милашинович Л. , Миттник А, Монге Дж.М., Нехризов Г., Николлс Р., Никитин А.Г., Николов В., Новак М., Олальде И., Оппенгеймер Дж., Остерхольц А., Оздемир Дж., Оздоган К.Т., Озтюрк Н., Пападимитриу Н., Папаконстантину Н., Папатанасиу А., Параман Л, Паскари Э.Г., Паттерсон Н., Петракиев И., Петросян Л., Петрова В., Филиппа-Туше А., Пилипосян А., Покуца Кузман Н., Потребица Х., Преда-Баланица Б., Премужич З., Прайс ТД, Цю Л., Радович С., Раеф. Азиз К, Раич Шиканич П, Рашид Рахим К, Разумов С, Ричардсон А, Руденберг Дж, Рука Р, Русева В, Шахин М, Шарбак А, Саваш Э, Шаттке Дж, Шепарц Л, Сельчук Т, Севим-Эрол А, Шамун -Пур М, Шепард ХМ, Сидерис А, Симальчик А, Симонян Х, Синица В, Сирак К, Сирбу Г, Шлаус М, Софикару А, Сёгют Б, Солтисиак А, Сенмез-Сёзер Ч, Стати М, Стескал М, Стюардсон К. , Стокер С, Суата-Альпаслан Ф, Суворов А, Сечени-Надь А, Сеничей Т, Тельнов Н, Темов С, Тодорова Н, Тота У, Тушайс Г, Триантафиллу С, Тюркер А, Угаркович М, Валчев Т, Вельяновска Ф, Видевски З., Вираг Дж., Вагнер А., Уолш С., Влодарчак П., Уоркман Ю.Н., Ярдумян А., Яровой Е., Явуз А.Ю., Йылмаз Х., Залзала Ф., Зеттл А., Чжан З., Чавушоглу Р., Роланд Н., Пинхаси Р., Райх Д. , Давтян Р. Генетическая история Южной дуги: мост между Западной Азией и Европой. Наука. 26 августа 2022 г.; 377 (6609): eabm4247. doi: 10.1126/science.abm4247. Электронная публикация 2022 г., 26 августа. PMID 36007055; PMCID: PMC10064553.
- ^ Чинниоглу, Дженгиз; Кинг, Рой; Кивисилд, Тоомас; Калфоглу, Эрси; Атасой, Севиль; Каваллери, Джанпьеро Л.; Лилли, Анита С.; Роузман, Чарльз К.; Лин, Алиса А.; Принц, Кристина; Офнер, Питер Дж.; Шен, Пейдун; Семино, Орнелла; Кавалли-Сфорца, Л. Лука; Андерхилл, Питер А. (1 января 2004 г.). «Раскопки слоев гаплотипов Y-хромосомы в Анатолии» . Генетика человека . 114 (2): 127–148. дои : 10.1007/s00439-003-1031-4 . ISSN 0340-6717 . ПМИД 14586639 .
- ^ Насидзе Иван; Саркисян Тамара; Керимов, Азер; Стоункинг, Марк (14 декабря 2002 г.). «Проверка гипотезы замены языка на Кавказе: данные Y-хромосомы». Генетика человека . 112 (3): 255–261. дои : 10.1007/s00439-002-0874-4 . ПМИД 12596050 . S2CID 13232436 .
- ^ Насидзе И и др. (2008). «Тесная генетическая связь между семитоязычными и индоевропейскоязычными группами в Иране». Анналы генетики человека . 72 (Часть 2): 241–52. дои : 10.1111/j.1469-1809.2007.00413.x . ПМИД 18205892 . S2CID 5873833 .
- ^ «G-Z7941 — Проект гаплогруппы G» . гугл.com .
- ^ Зей Дж. и др. (2003). «От фамилий к истории Y-хромосом: население Сардинии как парадигма» . Европейский журнал генетики человека . 11 (10): 802–07. дои : 10.1038/sj.ejhg.5201040 . ПМИД 14512971 .
- ^ Мартинес Л. и др. (апрель 2007 г.). «На Критском высокогорном плато преобладает палеолитическое наследие Y-гаплогруппы» . Европейский журнал генетики человека . 15 (4): 485–93. дои : 10.1038/sj.ejhg.5201769 . PMID 17264870 .
- ^ Кинг Р.Дж. и др. (март 2008 г.). «Дифференциальное анатолийское влияние Y-хромосомы на греческий и критский неолит». Анналы генетики человека . 72 (Часть 2): 205–14. дои : 10.1111/j.1469-1809.2007.00414.x . ПМИД 18269686 . S2CID 22406638 .
- ^ Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Лопата, Мэри; Майрес, Натали М; Кинг, Рой Дж; Корни, Сирия; Марьянович, Дамир; Приморак, Драган; Хаджиселимович, Рифат; Видович, Стойко; Дробник, Катя; Дурмиши, Насер; Торрони, Энтони; Сантакьяра-Бенерсетти, Сильвана; Андерхилл, Питер А; Семино, Орнелла (24 декабря 2008 г.). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» . Европейский журнал генетики человека . 17 (6): 820–830. дои : 10.1038/ejhg.2008.249 . ПМК 2947100 . ПМИД 19107149 .
- ^ Сингх, Мугдха; Саркар, Ануджит; Нандинени, Мадхусудан Р. (18 октября 2018 г.). «Полный портрет разнообразия Y-STR индийского населения и сравнение со 129 популяциями по всему миру» . Научные отчеты . 8 (1): 15421. Бибкод : 2018NatSR...815421S . дои : 10.1038/s41598-018-33714-2 . ISSN 2045-2322 . ПМК 6194109 . ПМИД 30337554 .
- ^ Иасвархант, Мутхукришнан; Хак, Икрамул; Равеш, Зейнаб; Ромеро, Ирен Гальего; Меганатан, Пурлин Рамакоди; Дубей, Бхавна; Хан, Файзан Ахмед; Чаубей, Гьянешвар; Кивисилд, Тоомас; Тайлер-Смит, Крис; Сингх, Лалджи; Тангарадж, Кумарасами (март 2010 г.). «Следы происхождения к югу от Сахары и Ближнего Востока в мусульманском населении Индии» . Европейский журнал генетики человека . 18 (3): 354–363. дои : 10.1038/ejhg.2009.168 . ISSN 1018-4813 . ПМЦ 2859343 . ПМИД 19809480 .
- ^ Перейти обратно: а б с Фирасат С., Халик С., Мохьюддин А., Папайоанну М., Тайлер-Смит С., Андерхилл П.А., Аюб К. (2006). «Y-хромосомные доказательства ограниченного вклада греков в патанское население Пакистана» . Европейский журнал генетики человека . 15 (1): 121–26. дои : 10.1038/sj.ejhg.5201726 . ПМЦ 2588664 . ПМИД 17047675 .
- ^ Перейти обратно: а б Сенгупта С. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ Саху, Сангамитра; Сингх, Анамика; Химабинду, Г.; Банерджи, Джилам; Ситалакшими, Т.; Гайквад, Сонали; Триведи, Р.; Эндикотт, Филипп; Кивисилд, Тоомас; Мецпалу, Мейт; Виллемс, Ричард (24 января 2006 г.). «Предыстория индийских Y-хромосом: оценка сценариев демического распространения» . Труды Национальной академии наук . 103 (4): 843–848. Бибкод : 2006PNAS..103..843S . дои : 10.1073/pnas.0507714103 . ISSN 0027-8424 . ПМК 1347984 . ПМИД 16415161 .
- ^ Хаммер М.Ф. и др. (2006). «Двойное происхождение японцев: точки соприкосновения Y-хромосом охотников-собирателей и фермеров» . Журнал генетики человека . 51 (1): 47–58. дои : 10.1007/s10038-005-0322-0 . ПМИД 16328082 .
- ^ Хуснутдинова Е.К.; Екомасова, Н.В.; Джаубермезов, М.А.; Габидуллина, ЛР; Суфьянова З.Р.; Хидиятова И.М.; Казанцева А.В.; Литвинов, С.С.; Нургалиева А.Х.; Прокофьева, ДС (2023). «Распространение гаплогруппы G-P15 Y-хромосомы среди представителей древних культур и современных популяций Северной Евразии» . Опера Медика и Физиология 10 (4): 57–72. ISSN 2500-2295 .
- ^ Сенгупта, Сангамитра; Животовский Лев А.; Кинг, Рой; Мехди, SQ; Эдмондс, Кристофер А.; Чоу, Шерил-Эмилиан Т.; Лин, Алиса А.; Митра, Миташри; Сил, Самир К.; Рамеш, А.; Уша Рани, MV; Тхакур, Читра М.; Кавалли-Сфорца, Л. Лука; Маджумдер, Парта П.; Андерхилл, Питер А. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы с высоким разрешением в Индии определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Американский журнал генетики человека . 78 (2): 202–221. дои : 10.1086/499411 . ISSN 0002-9297 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ Сингх, Сакши; Сингх, Ашиш; Раджкумар, Раджа; Сампат Кумар, Катакам; Кадаркарай Сами, Суббурадж; Низамуддин, шейх; Сингх, Амита; Ахмед Шейх, Шахнаваз; Педдада, Видья; Ханна, Вини; Вирайя, Пандичелвам; Пандит, Аридаман; Чаубей, Гьянешвар; Сингх, Лалджи; Тангарадж, Кумарасами (2016). «Анализ влияния неолитической демической диффузии на индийский пул Y-хромосом через гаплогруппу J2-M172» . Научные отчеты . 6 : 19157. Бибкод : 2016NatSR...619157S . дои : 10.1038/srep19157 . ПМК 4709632 . ПМИД 26754573 .
- ^ Однако в Ливане группа G составляет 6,5% населения, а в Иране - около 10%. Флорес С. и др. (2005). «Изоляты в коридоре миграций: анализ вариаций Y-хромосомы с высоким разрешением в Иордании» . Журнал генетики человека . 50 (9): 435–41. дои : 10.1007/s10038-005-0274-4 . ПМИД 16142507 .
- ^ Каденас А.М. и др. (2008). «Хромосомное разнообразие характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–86. дои : 10.1038/sj.ejhg.5201934 . ПМИД 17928816 .
- ^ Шлуш Л.И. и др. (2008). Геммелл, Нью-Джерси (ред.). «Друзы: популяционно-генетическое убежище Ближнего Востока» . ПЛОС ОДИН . 3 (5): е2105. Бибкод : 2008PLoSO...3.2105S . дои : 10.1371/journal.pone.0002105 . ПМК 2324201 . ПМИД 18461126 .
- ^ Элизабет Т. Вуд, Дарин А. Стовер, Кристофер Эрет и др. , «Контрастные закономерности вариаций Y-хромосомы и мтДНК в Африке: свидетельства демографических процессов с учетом пола», European Journal of Human Genetics (2005) 13, 867–876. (см. Приложение A: Частоты гаплотипов Y-хромосомы)
- ^ Луис Дж.Р. и др. (2004). «Левант против Африканского Рога: свидетельства двунаправленных коридоров миграции людей» . Американский журнал генетики человека . 74 (3): 532–44. дои : 10.1086/382286 . ПМЦ 1182266 . ПМИД 14973781 .
- ^ Эль-Сибай М., Платт Д.Э., Хабер М., Сюэ Ю., Юханна С.С., Уэллс Р.С., Изаабель Х., Санюра М.Ф., Харманани Х., Бонаб М.А., Бехбехани Дж., Хашва Ф., Тайлер-Смит С., Заллуа П.А. (2009). «Географическая структура Y-хромосомного генетического ландшафта Леванта: контраст прибрежных и внутренних территорий» . Анналы генетики человека . 73 (Часть 6): 561–81. дои : 10.1111/j.1469-1809.2009.00538.x . ПМЦ 3312577 . ПМИД 19686289 .
- ^ Алонсо С, Флорес С, Кабрера В, Алонсо А, Мартин П, Альбарран С, Исагирре Н, де ла Руа С, Гарсия О (2005). «Место басков в европейском ландшафте разнообразия Y-хромосомы» . Европейский журнал генетики человека . 13 (12): 1293–1302. дои : 10.1038/sj.ejhg.5201482 . ПМИД 16094307 .
- ^ Кручиани Ф и др. (2002). «Обратная миграция из Азии в страны Африки к югу от Сахары подтверждается анализом гаплотипов Y-хромосомы человека с высоким разрешением» . Американский журнал генетики человека . 70 (5): 1197–1214. дои : 10.1086/340257 . ПМЦ 447595 . ПМИД 11910562 .
- ^ Ванек Д., Саскова Л., Кох Х. (2009). «Анализ родства и Y-хромосомы человеческих останков VII века: новая процедура извлечения ДНК и типирования древнего материала» . Хорватский медицинский журнал . 50 (3): 286–95. дои : 10.3325/cmj.2009.50.286 . ПМК 2702742 . ПМИД 19480023 .
- ^ «Ледяной человек Эци: что мы знаем спустя 3 десятилетия после его открытия» . Премиум . 15 сентября 2021 г. Архивировано из оригинала 27 мая 2023 года . Проверено 21 октября 2023 г.
- ^ «Точечная мутация, которая включает в себя изменение A → G, C → T, C → G и T → A в ДНК» . Топпр Спросите . Проверено 21 октября 2023 г.
- ^ ymap.ftdna.com [ постоянная мертвая ссылка ]
- ^ Адамс С.М., Бош Э., Балареск П.Л., Баллеро С.Дж., Ли А.С., Арройо Е., Лопес-Парра А.М., Алер М., Грифо М.С., Брайон М., Карраседо А., Лавинья Дж., Мартинес-Харрета Б., Кинтана-Мурси Л., Пикорнелл А. , Рамон М., Скорецки К., Бехар Д.М., Калафель Ф., Джоблинг М.А. (декабрь 2008 г.). «Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, евреев и мусульман на Пиренейском полуострове» . Американский журнал генетики человека . 83 (6): 725–36. дои : 10.1016/j.ajhg.2008.11.007 . ПМК 2668061 . ПМИД 19061982 .
- ^ L177, позже отброшенный в пользу PF3359 и эквивалентных SNP, был впервые идентифицирован в ДНК генеалогического древа в 2009 году. Компонент L177.1 обнаружен в положении Y-хромосомы 23397163; Л177.2 по номеру 25030912; L177.3 по адресу 25750264. См.: http://ytree.ftdna.com/index.php?name=Draft&parent=20173662. Архивировано 18 октября 2010 г. на Wayback Machine .
- ^ «..Списки проектов - Проект гаплогруппы G» . гугл.com . Архивировано из оригинала 27 ноября 2013 г. Проверено 9 декабря 2011 г.
- ^ Хаммер М.Ф. и др. (2009). «Расширенные гаплотипы Y-хромосомы определяют многочисленные и уникальные линии еврейского духовенства» . Генетика человека . 126 (5): 707–17. дои : 10.1007/s00439-009-0727-5 . ПМЦ 2771134 . ПМИД 19669163 .
- ^ Хабе, М; и др. (28 марта 2012 г.). «Этнические группы Афганистана разделяют Y-хромосомное наследие, структурированное историческими событиями» . ПЛОС ОДИН . 7 (3): e34288. Бибкод : 2012PLoSO...734288H . дои : 10.1371/journal.pone.0034288 . ПМЦ 3314501 . ПМИД 22470552 .
- ^ Авторские права: ISOGG, 2015 г. «Дерево гаплогрупп Y-ДНК ISOGG 2015» . isogg.org .
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Андерхилл П.А., Пассарино Г., Лин А.А., Шен П., Мирасон Лар М., Фоли Р.А., Офнер П.Дж., Кавалли-Сфорца Л.Л. (2001). «Филогеография бинарных гаплотипов Y-хромосомы и происхождение современных человеческих популяций» . Анналы генетики человека . 65 (Часть 1): 43–62. дои : 10.1046/j.1469-1809.2001.6510043.x . ПМИД 11415522 . S2CID 9441236 .
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогруппы Y-хромосомы человека» . Геномные исследования . 18 (5): 830–38. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ http://ymap.ftdna.com/cgi-bin/gbrowse_details/hs_chrY?name=L240;class=Sequence;ref=ChrY;start=3191153;end=3191153;feature_id=40369 [ постоянная мертвая ссылка ]
- ^ Симс Л.М. и др. (2009). Батцер М.А. (ред.). «Филогения гаплогруппы G улучшенного разрешения в Y-хромосоме, выявленная с помощью набора недавно охарактеризованных SNP» . ПЛОС ОДИН . 4 (6): 1–5. Бибкод : 2009PLoSO...4.5792S . дои : 10.1371/journal.pone.0005792 . ПМК 2686153 . ПМИД 19495413 .
- ^ Кинг Т.Э., Фортес Г.Г., Балареск П., Томас М.Г., Болдинг Д., Майзано Дельсер П., Нойманн Р., Парсон В., Кнапп М., Уолш С., Тонассо Л., Холт Дж., Кайзер М., Эпплби Дж., Форстер П., Эксерджян Д., Хофрейтер М, Шюрер К (2014). «Опознание останков короля Ричарда III» . Нат. Коммун . 5 : 5631. Бибкод : 2014NatCo...5.5631K . дои : 10.1038/ncomms6631 . ПМЦ 4268703 . ПМИД 25463651 .
- ^ «Дневной зверь» .
- ^ «G-P303 (Y-ДНК)» .
- ^ Список древней ДНК
- ^ Масьямо. «Эупедия» . Евпедия . Проверено 7 августа 2022 г.
- ^ «Результаты тестов ДНК Y-хромосомы семьи Хамман» . Архивировано из оригинала 2 февраля 2023 г. Проверено 14 апреля 2022 г.
- ^ «Гаплогруппа G2a (Y-хромосомная ДНК) — Eupedia» .
Внешние ссылки
[ редактировать ]- Сайт проекта гаплогруппы G. Архивировано 31 июля 2014 г. на Wayback Machine.
- Распространение гаплогруппы G по данным National Geographic.
- Учебник по гаплогруппе G от Genebase
- Гаплогруппа G Y-ДНК и ее субклады из гаплодерева ISOGG текущего года
- Y-поиск пользователей с гаплогруппой G [ постоянная мертвая ссылка ]
- Проект ДНК Британских островов
- Проект гаплогруппы G в ДНК генеалогического древа
- Проект G Arabia в ДНК генеалогического древа