Гаплогруппа C-M130
| Гаплогруппа С | |
|---|---|
 | |
| Возможное время происхождения | 53 000 лет назад [1] |
| Возможное место происхождения | Юго-Западная Азия , через миграцию из Африки. [ 1 ] [ 2 ] [ 3 ] |
| Предок | CF |
| Потомки | C1 F3393/Z1426 (ранее CxC3) C2 (ранее C3*) M217 [ 4 ] |
| Определение мутаций | М130/РПС4И 711 , П184, П255, П260 |
Гаплогруппа C — это основная Y-хромосомы гаплогруппа , определяемая UEPs M130/RPS4Y 711 , P184, P255 и P260, которые все являются мутациями SNP . Это одна из двух основных ветвей гаплогруппы CF наряду с гаплогруппой F. Гаплогруппа C встречается у древних популяций на всех континентах, кроме Африки, и является преобладающей гаплогруппой Y-ДНК среди мужчин, принадлежащих ко многим народам, коренным народам Восточной Азии , Центральной Азии , Сибири , Северной Америки и Австралии, а также у некоторых популяций в Европе . Левант , а позднее Япония . [ 1 ]
Гаплогруппа также встречается с умеренной и низкой частотой среди многих современных популяций Юго-Восточной Азии , Южной Азии и Юго-Западной Азии .
Помимо базальной парагруппы C*, эта гаплогруппа теперь имеет две основные ветви: C1 (F3393/Z1426; ранее CxC3, т.е. старый C1, старый C2, старый C4, старый C5 и старый C6) и C2 (M217; бывший C3). ).
Происхождение
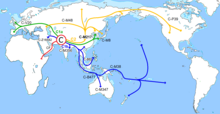
[ редактировать ]Гаплогруппа C-M130, вероятно, возникла в результате исхода современных людей из Африки , которые распространились на восток из Юго-Западной Азии и постепенно колонизировали Южную Азию, Восточную Азию и Океанию. Исследования относительно того, как произошла эта миграция, разделились; большинство исследований поддерживают Северный путь через Сибирь, в то время как другие поддерживают гипотезу Южного пути, согласно которой носители гаплогруппы C мигрировали вдоль побережий Индии и Юго-Восточной Азии, чтобы добраться до Китая. [ 2 ]
Гаплогруппа C-M130, по-видимому, возникла вскоре после того, как мутация SNP впервые произошла M168, что привело к появлению современной гаплогруппы CT , от которой произошли гаплогруппа CF и, в свою очередь, гаплогруппа C. Вероятно, это было по крайней мере 60 000 лет назад.
Гаплогруппа C-M130 достигает наибольшей частоты среди коренного населения Казахстана , Монголии , Дальнего Востока России , Полинезии , некоторых групп Австралии , а также умеренной частоты у коренного населения Кореи и маньчжуров . Поэтому предполагается, что гаплогруппа C-M130 либо возникла, либо прошла самый длительный период эволюции в регионе Большой Центральной Азии или в регионах Юго-Восточной Азии. Предполагается, что его распространение в Восточной Азии началось примерно 40 000 лет назад. [ 2 ]
Считается, что самцы-носители C-M130 мигрировали в Америку около 6000–8000 лет назад и были занесены народами, говорящими на языке на-дене, на северо-западное тихоокеанское побережье Северной Америки .
Азия также является территорией гаплогруппы D-M174 концентрации . Однако D-M174 более тесно связан с гаплогруппой E, чем с C-M130, а географическое распространение гаплогрупп C-M130 и D-M174 полностью и совершенно различно, причем различные подтипы гаплогруппы C-M130 встречаются с высокой частотой среди современные казахи и монголы , а также у некоторых коренных народов Америки , маньчжуры . Он также встречается со средней частотой у корейцев , коренных жителей Дальнего Востока России , некоторых групп австралийских аборигенов и с умеренной частотой в других местах по всей Азии и Океании. Носители гаплогруппы C среди более позднего народа Дзёмон в Японии и некоторых европейцев эпохи палеолита и неолита несли C1a, C1b и C1a2. Тогда как гаплогруппа D встречается с высокими частотами только среди тибетцев, японцев и жителей Андаманских островов и не обнаружена ни в Индии, ни среди коренных жителей Америки или Океании. [ 1 ]
По мнению Сакитани и др., гаплогруппа C-M130 возникла в Центральной Азии и распространилась оттуда в другие части Евразии и в части Австралии. Предполагается, что С-М130 был обнаружен у охотников-собирателей Восточной Евразии, а также в древних образцах Восточной и Юго-Восточной Азии и Европы. [ 1 ]
Структура
[ редактировать ]С* (M130/Страница51/RPS4Y711, M216)
- С1 (F3393)
- С1а (CTS11043)
- С1б (F1370)
- С2 (ранее С3) — (М217)
- С2а(М93)
- C2b (L1373/F1396)
- C2b1
- C2b1a
- C2b1a1
- C2b1a1a (P39)
- C2b1a2 (ранее C3c) — (M48)
- C2b1a1
- C2b1a
- C2b1
- C2c (C-F1067)
- C2c1 (F2613/Z1338)
- C2c1a (Z1300)
- C2c1a1
- C2c1a1a
- C2c1a1a1 (M407)
- C2c1a1a
- C2c1a1
- C2c1a (Z1300)
- C2c1 (F2613/Z1338)
- Другие нетаксономизированные субклады :
(Приведенная выше филогенетическая структура субкладов гаплогруппы C-M130 основана на дереве ISOGG 2015, дереве YCC 2008 и последующих опубликованных исследованиях. [ 7 ] [ 8 ] )
Распределение
[ редактировать ]
Распространение гаплогруппы C-M130 обычно ограничивается населением Сибири, некоторых частей Восточной Азии, Океании и Америки . Из-за огромного возраста гаплогруппы C успели накопиться многочисленные вторичные мутации, и было идентифицировано множество регионально важных подветвей гаплогруппы C-M130.
до 46% австралийских аборигенов -мужчин были носителями базального C* (C-M130*), C1b2b* (C-M347*) или C1b2b1 (C-M210) до контакта Согласно исследованию 2015 года, с европейцами и значительной иммиграции со стороны них. Нэгл и др. [ 10 ] То есть 20,0% Y-хромосом 657 современных людей, раньше 56% этих выборок были исключены как «некоренные». До колонизации C-M130*, по-видимому, носили до 2,7% мужчин-аборигенов; 43% имели C-M347, который не был обнаружен за пределами Австралии. Остальные гаплогруппы аборигенов Австралии сходны с папуасами и другими негритосами ( гаплогруппы S-M230 и M-P256 ). [ 10 ] [ 11 ]
Низкие уровни C-M130* переносят самцы:
- из Индийского субконтинента , Шри-Ланки и Юго-Восточной Азии , [ 7 ] и;
- в Европе среди мужчин с фамилией Ллах, происходящих из Гаррочи , Каталония , Испания (но не мужчин с такой же фамилией из других регионов). [ 12 ]
Базальный C1a* (CTS11043) был обнаружен у европейцев верхнего палеолита ( ориньякцев ), GoyetQ116-1 и Pestera Muerii2. [ 13 ]
C1b был идентифицирован в доисторических останках, датируемых 34 000 лет назад, найденных в России и известных как « Костенки 14 ». [ 14 ]
Гаплогруппа C2 (M217) — самая многочисленная и широко распространенная линия C — когда-то считалась, что она возникла в Центральной Азии , распространилась оттуда в Северную Азию и Америку, в то время как по другой теории она возникла из Восточной Азии. [ 7 ] C-M217 простирается в продольном направлении от Центральной Европы и Турции до народа вайю в Колумбии и Венесуэле , а в широтном направлении от атабаскских народов Аляски до Вьетнама и Малайского архипелага . Встречается в небольших концентрациях в Восточной Европе , где может быть наследием вторжений/миграций гуннов , тюрков и /или монголов в средние века. Особенно часто встречается у бурятов , дауров , хазарейцев , ительменов , калмыков , коряков , маньчжуров , монголов , орокенцев и сибесов , с умеренным распространением среди других тунгусских народов , корейцев , айнов , нивхов , алтайцев , тувинцев , узбеков , ханьских китайцев. , Туцзя , Хани и Хуэй . [ 15 ] [ 16 ] [ 17 ] [ 18 ] [ 19 ] [ 20 ] [ 21 ] Самые высокие частоты гаплогруппы C-M217 встречаются среди населения Монголии и Дальнего Востока России , где она является модальной гаплогруппой. Гаплогруппа C-M217 — единственная разновидность гаплогруппы C-M130, встречающаяся среди коренных американцев , среди которых она достигает наибольшей частоты в популяциях На-Дене . Также имело бы смысл, что эта линия, возможно, возникла в Америке, поскольку это единственная разновидность, встречающаяся среди аборигенного населения.
Другие субклады специфичны для определенных популяций в пределах ограниченного географического ареала; даже там, где эти другие ветви встречаются, они имеют тенденцию проявляться как очень низкочастотный, второстепенный компонент палитры разнообразия Y-хромосомы на этих территориях:
- C-M8 (C1a1) в настоящее время регулярно встречается только с низкой частотой (около 5%; диапазон 3,3% — 10% всех образцов) в Японии. Он также был обнаружен в ходе академического исследования у одного человека на острове Чеджу и в ходе коммерческих испытаний у одного человека, который сообщил о своем происхождении из провинции Ляонин в Китае, и у одного человека, который сообщил о своем происхождении из Сеула. У древнего народа Дзёмон частота C1a1 составляла около 30%.
- C-V20 (C1a2; ранее C6) встречается с низкой частотой среди европейцев. [ 22 ] Его несли 7000-летние останки охотника-собирателя из Ла-Бранья (современная Астурия , Испания), [ 23 ] также присутствовал в Венгрии . и C1a2 примерно в то же время [ 7 ] [ 24 ] В 2016 году здесь были найдены 35 000-летние останки охотника-собирателя из пещер Гойет ( Намюр , Бельгия) и 30 000-летние останки охотника-собирателя из Долни-Вестонице (Вестонице16, Моравия , Чехия). гаплогруппа. [ 25 ]
- C-B66/Z16458 (C1b1a) встречается с низкой частотой в Южной Азии , Центральной Азии и Юго-Западной Азии . [ 18 ] [ 26 ] [ 27 ] [ 28 ] [ 29 ] [ 30 ]
- C-M356 (C1b1a1; ранее C5) с низкой частотой обнаруживался в образцах из Индии , Непала , [ 26 ] [ 27 ] Пакистан , Афганистан , Аравия , [ 28 ] [ 29 ] и Северный Китай . [ 2 ] [ 7 ]
- C-M38 (C1b2a; ранее C2) среди некоторых местных популяций в Индонезии , Меланезии (особенно Новой Гвинее ), Микронезии и некоторых островах Полинезии C-M38 стала модальной гаплогруппой, вероятно, из-за серьезных эффектов основателя и генетического дрейфа. . [ 7 ]
- C-P33 (C1b2a1a): часто встречается среди полинезийских мужчин. [ 16 ] [ 31 ]
- C-P55 (C1b3) встречается с низкой частотой в высокогорьях Новой Гвинеи . [ 6 ]
- C-M93 (C2a) спорадически встречается у японцев . [ 18 ] [ 32 ]
- C-L1373/F1396 (C2b) идентифицирован в Центральной Азии. [ нужна ссылка ]
- C-P39 (C2b1a1a) встречается у нескольких коренных народов Северной Америки, включая некоторые группы населения, говорящие на языке на-дене , алгонкинском языке и сиуанском языке. [ 7 ] [ 33 ]
- C-M48 (C2b1b; ранее C3c) с высокой частотой встречается у северных тунгусских народов , казахов , ойратов , калмыков , монголов , юкагиров , нивхов , коряков , ительменов и удэгейцев , с умеренным распространением среди южно -тунгусских народов , бурятов , тувинцев. , якуты , чукчи , киргизы , уйгуры , узбеки , каракалпаки и таджики . [ 19 ] [ 34 ] [ 35 ]
- C-P53.1 (C2c) переносится примерно 10% мужчин Синьцзян- Сибе , с низкой частотой в Монголии и среди эвенков , Нинся- Хуэй , Сизан- Тибетцев , Синьцзян- Уйгуров и Ганьсу- Хань . [ 2 ]
- C-F2613/Z1338 (C2e): как Центральная Азия , так и Восточная Азия . [ 7 ]
- C-M407 (C2e1a1a) с высокой частотой обнаруживался среди баргутов , бурятов , сойотов и хамниганцев , с умеренной частотой среди других монголов и калмыков и с низкой частотой у армян , баев , камбоджийцев , эвенков , [ 36 ] Ханьцы , японцы , казахи , корейцы , маньчжуры , телеуты , [ 36 ] Туджа , тувинец , [ 36 ] Уйгурское и якутское население. [ 2 ] [ 18 ]
- C-P343 встречается с высокой частотой среди мужчин из Лембаты (17,4% из 92 образцов), с более низкой частотой во Флоресе , Пантаре и Тиморе . [ 5 ] P343 находится за пределами C1a1 (M8), C1b2a (M38), C2 (бывший C3; M217), C1b2b (ранее C4; M347) или C1b1a1 (ранее C5; M356), но его связь с другими ветвями еще не проверена. [ 5 ]
- Неуказанные экземпляры C-M130, которые, возможно, могут принадлежать к одному из вышеуказанных субкладов, включают:
- Кайзер и др. (2006) обнаружили C-M130(xM38, 390.1del, M217) в 10,3% (4/39) выборки групп этнических меньшинств из Филиппин, 10,0% (6/60) выборки из Арнемленда , 6,5% (2/31) выборки с Нуса-Тенгары , 5,3% (3/57) выборки с Суматры, 3,0% (1/33) выборки с побережья Папуа-Новой Гвинеи, 2,9% (1/34) образец с Молуккских островов , 1,9% (1/53) образца с Явы и 0,9% (1/107) образца с Фиджи . [ 37 ]
- C-RPS4Y (сейчас C-RPS4Y 711 ) (xM38) Y-ДНК довольно распространена среди населения индонезийской провинции Восточная Нуса-Тенгара и независимого Восточного Тимора : 13/31 = 41,9% Лембата , 16/71 = 22,5% Флорес , 5/43 = 11,6% Солор , 10/96 = 10,4% Адонара , 3/39 = 7,7% Восточный Тимор , 1/26 = 3,8% Алор , 1/38 = 2,6% Пантар . Все особи C-RPS4Y(xM38), за исключением одиночки из Алора, были описаны как говорящие на австронезийском языке . [ 38 ]
- C-RPS4Y (теперь C-RPS4Y 711 ) (xM38, M217) Y-ДНК встречается, согласно исследованию, опубликованному в 2010 году, с довольно высокой частотой в большинстве популяций центральной Индонезии (115/394 = 29,2% Флорес, 21/92). = 22,8% Лембата, 19/86 = 22,1% Борнео, 6/54 = 11,1% Мандар , 1/9 = 11,1% Тимор, но только 1/350 = 0,3% Сумба). C-RPS4Y(xM38, M217) Y-ДНК обычно становится редкой по направлению к западу (2/61 = 3,3% Явы, 1/32 = 3,1% Малайзии, 9/641 = 1,4% Балийцев, 0/38 Батак Тоба, 0/ 60 Ниас, но 10/74 = 13,5% Ментавай) и на восток (1/28 = 3,6% Алор, 0/30 Молуккские острова, 1/15 = 6,7% Побережье PNG, 0/33 Хайленд PNG, 0/10 Насиой, 0/44 Маэво (Вануату), 1/16 = 6,3% Микронезия, 0/64 Полинезия). [ 39 ]
- C-RPS4Y 711 (xM8, M217) Y-ДНК была обнаружена в 17% (6/35) выборки Яо из Бамы , Гуанси на юге центрального Китая, 4/35 = 11% выборки Хуэй и 2/70 = 3% пары выборок уйгуров из северо-западного Китая, 3/45 = 7% выборки хэчжэ и 1/26 = 4% выборки эвэньков из северо-восточного Китая. [ 15 ]
- C-RPS4Y 711 (xM8, M38, M217) обнаружен у 48,5% (16/33) выборки коренных австралийцев , 20% (12/60) выборки Яо , 6,1% (3/49) выборки выборка туджиа , 5,9% (1/17) выборки микронезийцев , 5,5% (3/55) выборки восточных индонезийцев , 4,0% (1/25) выборки западных индонезийцев, 3,3% (3 /91) выборки шриланкийцев , 3,1% (1/32) выборки малайцев , 2,5% (10/405) выборки индийцев , 2,2% (1/46) выборки жителей Папуа-Новой Гвинеи , 1,7% (1/58) выборки мяо и 1,5% (1/67) выборки уйгуров . [ 16 ]
- Чжун и др. (2010) обнаружили Y-ДНК C-M130(xM8, M38, M217, M347, M356, P55) в 9,09% (1/11) образца Мулао из Гуанси, 8,20% (5/61) образца. хуэй из Нинся, 7,96% (9/113) образца Яо из Гуанси, 7,69% (2/26) образца Туцзя из Хубэя, 6,90% (2/29) образца шуй из Гуйчжоу, 3,28% (2/61) выборки Сибе из Синьцзяна, 1,45% (1/69) выборки Чжуан из Гуанси, 0,92% (1/109) выборки Буи из Гуйчжоу, 0,87% (2/ 231) образца Хань из Гуйчжоу и 0,74% (1/136) образца Хань из Юньнани. [ 2 ] Большинство этих людей имеют одинаковый гаплотип Y-STR из 8 локусов: DYS19=15, DYS389I=12, DYS389b=16, DYS388=13, DYS390=21, DYS391=10, DYS392=11, DYS393=12. [ 2 ]
- Ди Кристофаро и др. (2013) обнаружили C-M130(xC2-PK2/M386, C1b1a1-M356) у 1,9% (1/53) выборки пуштунов из Кундуза, Афганистан, и у 1,4% (1/69) выборки хазарейцев. из Бамиана, Афганистан. [ 40 ]
- Ван и др. (2014) обнаружили C-M130(xM105, M38, M217, M347, M356) в 5,6% (1/18) выборки Хорпа Цян из округа Данба провинции Сычуань, Китай, и в 2,2% (1/46) образец тибетцев Кхамса из округа Синьлун провинции Сычуань, Китай. [ 41 ] Гаплотипы Y-STR этих двух людей соответствуют модальному гаплотипу C* из исследования Zhong et al. (2010) в каждом сопоставимом локусе. [ 41 ]
- «C-M216», SNP, который теперь считается синонимом C-M130 - (xM8, M38, M217, M210, M356), был обнаружен у 3,9% (3/77) выборки общего населения Катманду. , Непал. [ 26 ] Другие примеры C-M216 были зарегистрированы среди монгольских и тюркских народов , в том числе: джалаиров , казахов , меркитов , джучидов и узбеков ; [ нужна ссылка ]
Филогенетика
[ редактировать ]Филогенетическая история
[ редактировать ]До 2002 года в научной литературе существовало как минимум семь систем наименования филогенетического дерева Y-хромосомы. Это привело к значительной путанице. В 2002 году основные исследовательские группы объединились и сформировали Консорциум Y-хромосомы (YCC). Они опубликовали совместный документ, в котором было создано единое новое дерево, которое все согласились использовать. В таблице ниже собраны все эти работы, относящиеся к знаковому дереву YCC 2002 года. Это позволяет исследователю, просматривающему старую опубликованную литературу, быстро перемещаться между номенклатурами.
| YCC 2002/2008 (сокращенно) | (а) | (б) | (с) | (г) | (е) | (г) | (или) | YCC 2002 (от руки) | YCC 2005 (от руки) | YCC 2008 (от руки) | YCC 2010r (от руки) | ИСОГГ 2006 г. | ИСОГГ 2007 | ИСОГГ 2008 | ИСОГГ 2009 | ИСОГГ 2010 | ИЗОГГ 2011 | ИЗОГГ 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| С-М216 | 10 | V | 1F | 16 | Еу6 | H1 | С | С* | С | С | С | С | С | С | С | С | С | С |
| С-М8 | 10 | V | 1F | 19 | Еу6 | H1 | С | С1 | С1 | С1 | С1 | С1 | С1 | С1 | С1 | С1 | С1 | С1 |
| С-М38 | 10 | V | 1F | 16 | Еу6 | H1 | С | С2* | С2 | С2 | С2 | С2 | С2 | С2 | С2 | С2 | С2 | С2 |
| C-P33 | 10 | V | 1F | 18 | Еу6 | H1 | С | C2a | C2a | C2a1 | C2a1 | C2a | C2a | C2a1 | C2a1 | C2a1 | удаленный | удаленный |
| C-P44 | 10 | V | 1F | 17 | Еу6 | H1 | С | С3* | С3 | С3 | С3 | С3 | С3 | С3 | С3 | С3 | С3 | С3 |
| С-М93 | 10 | V | 1F | 17 | Еу6 | H1 | С | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a | C3a1 |
| С-М208 | 10 | V | 1F | 17 | Еу6 | H1 | С | C3b | C2b | C2a | C2a | C2b | C2b | C2a | C2a | C2a | C2a | C2a |
| С-М210 | 36 | V | 1F | 17 | Еу6 | H1 | С | C3c | C2c | C4a | C4a | C4b | C4b | C4a | C4a | C4a | C4a | C4a |
Научные публикации
[ редактировать ]Следующие исследовательские группы в соответствии с их публикациями были представлены в создании Дерева YCC.
Известные участники
[ редактировать ]Один конкретный гаплотип в гаплогруппе C-M217 привлек большое внимание из-за возможности того, что он может представлять прямое отцовское происхождение от Чингисхана .
Опубликованная в 2017 году исследовательская работа - «Генетический след ранних миграций Айсин Гиоро, императорского дома династии Цин». [ 42 ] подтвердил, что клан Айсин Гиоро принадлежит к гаплогруппе C3b1a3a2-F8951, братской ветви C3*-Звездного скопления (в настоящее время называемого C3b1a3a1-F3796, когда-то связанного с Чингисханом ).
См. также
[ редактировать ]Генетика
[ редактировать ]- генетическая генеалогия
- Гаплогруппа
- Гаплотип
- Гаплогруппа ДНК Y-хромосомы человека
- молекулярная филогения
- Парагруппа
- Субклад
- Гаплогруппы Y-хромосомы в популяциях мира
- Гаплогруппы Y-ДНК по этническим группам
- Гаплогруппы Y-ДНК в популяциях Восточной и Юго-Восточной Азии
- Гаплогруппы Y-ДНК в популяциях Океании
Субклады Y-ДНК C
[ редактировать ]Основополагающее дерево Y-ДНК
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Мицуру Сакитани, «Новая история Японского архипелага, раскрытая в результате междисциплинарных исследований ДНК, археологии и языка» (Bensei Publishing, 2009) (на японском языке)
- ^ Jump up to: а б с д и ж г час Чжун Х., Ши Х., Ци XB и др. (июль 2010 г.). «Глобальное распространение гаплогруппы Y-хромосомы C-M130 раскрывает доисторические миграционные пути африканского исхода и раннего расселения в Восточной Азии» . Дж. Хум. Жене . 55 (7): 428–35. дои : 10.1038/jhg.2010.40 . ПМИД 20448651 .
- ^ «В настоящее время большая часть археологических и генетических данных подтверждает, что самый ранний исход африканцев произошел из Африки через Красное море, а затем быстро мигрировал в материковую часть Юго-Восточной Азии через береговую линию Индии и в конечном итоге достиг Океании.36, 37, 38, 39 Недавний анализ Y-хромосомы и митохондриальной ДНК в Австралии и Новой Гвинее показал, что Hg C, вероятно, является одним из самых ранних типов-основателей за пределами Африки,12 что также было предложено в другом исследовании6, и что линии митохондриальной ДНК, состоящие из типы-основатели (M и N) датируются приблизительно 50–70 годами тыс. лет назад.12" ... "Мы предполагаем, что Hg C возникла в результате исхода из Африки и постепенно колонизировала Южную Азию, Юго-Восточную Азию, Океанию и Восточную Азию одним Палеолитическая миграция из Африки в Азию и Океанию, произошедшая более 40 тыс. лет назад».
- ^ «ISOGG 2018 Гаплогруппа C Y-ДНК» .
- ^ Jump up to: а б с Тумонггор, Мерианн К; Карафет Татьяна М; Дауни, Шон; и др. (2014). «Изоляция, контакты и социальное поведение сформировали генетическое разнообразие в Западном Тиморе» . Журнал генетики человека . 59 (9): 1–10. дои : 10.1038/jhg.2014.62 . ПМЦ 4521296 . ПМИД 25078354 .
- ^ Jump up to: а б Шейнфельдт, Л.; Фридлендер, Ф; Фридлендер, Дж; Лэтэм, К; Коки, Г; Карафет, Т; Хаммер, М; Лоренц, Дж (2006). «Неожиданная хромосомная вариация NRY на Северном острове Меланезии» . Молекулярная биология и эволюция . 23 (8): 1628–41. дои : 10.1093/molbev/msl028 . ПМИД 16754639 .
- ^ Jump up to: а б с д и ж г час ISOGG, 2015 «Гаплогруппа C Y-ДНК и ее субклады – 2015» (15 сентября 2015 г.).
- ^ Карафет Т.М., Мендес Ф.Л., Мейлерман М.Б., Андерхилл П.А., Зегура С.Л., Хаммер М.Ф. (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Ван, CC; Ли, Х (2013). «Выводы об истории человечества в Восточной Азии на основе Y-хромосом» . Расследование Жене . 4 (1): 11. дои : 10.1186/2041-2223-4-11 . ПМЦ 3687582 . ПМИД 23731529 .
- ^ Jump up to: а б Нэгл, Н.; и др. (2015). «Древность и разнообразие Y-хромосом аборигенов Австралии». Американский журнал физической антропологии . 159 (3): 367–81. дои : 10.1002/ajpa.22886 . ПМИД 26515539 . S2CID 2225529 .
- ^ Худжашов Г., Кивисилд Т., Андерхилл П.А. и др. (май 2007 г.). «Выявление доисторического поселения Австралии с помощью Y-хромосомы и анализа мтДНК» . Учеб. Натл. акад. наук. США . 104 (21): 8726–30. Бибкод : 2007PNAS..104.8726H . дои : 10.1073/pnas.0702928104 . ПМК 1885570 . ПМИД 17496137 .
- ^ Фамилии каталонцев, без даты, Результат (15 сентября 2015 г.). (Проект Cognoms Catalans, который исследует «генетические фамилии» в Каталонии, Валенсии и на Балеарских островах, базируется в Университете Помпеу Фабра в Барселоне.)
- ^ Фу, К.; Пост, К.; Хайдиняк, М.; Петр, М.; Маллик, С.; Фернандес, Д.; Фуртвенглер, А.; Хаак, В.; Мейер, М.; Миттник, А.; Никель, Б.; Пельтцер, А.; Роланд, Н.; Слон, В.; Таламо, С.; Лазаридис, И.; Липсон, М.; Мэтисон, И.; Шиффельс, С.; Скоглунд, П.; Деревянко А.П.; Дроздов Н.; Славинский, В.; Цыбанков А. ; Кремонези, RG ; Маллени, Ф.; Гелли, Б.; Вакка, Э.; Гонсалес Моралес, MR; и др. (2016). «Генетическая история Европы ледникового периода» . Природа 534 (7606): 200–205. Бибкод : 2016Nature.534..200F . дои : 10.1038/nature17993 . ПМЦ 4943878 . ПМИД 27135931 .
- ^ Сеген-Орландо, А.; и др. (2014). «Геномная структура европейцев насчитывает не менее 36 200 лет» (PDF) . Наука . 346 (6213): 1113–1118. Бибкод : 2014Sci...346.1113S . дои : 10.1126/science.aaa0114 . ПМИД 25378462 . S2CID 206632421 . Архивировано из оригинала (PDF) 29 августа 2016 г.
- ^ Jump up to: а б Сюэ Ю, Зержал Т, Бао В и др. (апрель 2006 г.). «Мужская демография в Восточной Азии: контраст между севером и югом во времени роста населения» . Генетика . 172 (4): 2431–9. doi : 10.1534/genetics.105.054270 . ПМЦ 1456369 . ПМИД 16489223 .
- ^ Jump up to: а б с Хаммер М.Ф., Карафет Т.М., Парк Х. и др. (2006). «Двойное происхождение японцев: общая основа Y-хромосом охотников-собирателей и фермеров» . Дж. Хум. Жене . 51 (1): 47–58. дои : 10.1007/s10038-005-0322-0 . ПМИД 16328082 .
- ^ Тадзима, Ацуши; Хаями, Масанори; Токунага, Кацуши; Джуджи, Т; Мацуо, М; Марзуки, С; Омото, К; Хорай, С (2004). «Генетическое происхождение айнов выведено на основе комбинированного анализа ДНК материнских и отцовских линий» . Журнал генетики человека . 49 (4): 187–193. дои : 10.1007/s10038-004-0131-x . ПМИД 14997363 .
- ^ Jump up to: а б с д Сенгупта С., Животовский Л.А., Кинг Р. и др. (февраль 2006 г.). «Полярность и временность распределения Y-хромосомы в Индии с высоким разрешением определяют как местное, так и экзогенное распространение и выявляют незначительное генетическое влияние скотоводов Центральной Азии» . Являюсь. Дж. Хум. Жене . 78 (2): 202–21. дои : 10.1086/499411 . ПМЦ 1380230 . ПМИД 16400607 .
- ^ Jump up to: а б Лелль Дж.Т., Сукерник Р.И., Стариковская Ю.Б. и др. (январь 2002 г.). «Двойное происхождение и сибирское сходство Y-хромосом коренных американцев» . Являюсь. Дж. Хум. Жене 70 (1): 192–206. дои : 10.1086/338457 . ПМЦ 384887 . ПМИД 11731934 .
- ^ Уэллс Р.С., Юлдашева Н., Рузибакиев Р. и др. (август 2001 г.). «Центр Евразии: континентальный взгляд на разнообразие Y-хромосомы» . Учеб. Натл. акад. наук. США . 98 (18): 10244–9. Бибкод : 2001PNAS...9810244W . дои : 10.1073/pnas.171305098 . ПМК 56946 . ПМИД 11526236 .
- ^ Насидзе И., Куинке Д., Дюпанлуп И., Кордо Р., Кокшунова Л., Стоункинг М. (декабрь 2005 г.). «Генетические доказательства монгольского происхождения калмыков». Являюсь. Дж. Физ. Антрополь . 128 (4): 846–54. дои : 10.1002/ajpa.20159 . ПМИД 16028228 . S2CID 27115596 .
- ^ Скоццари Р., Массаиа А., Д'Атанасио Э., Майрес Н.М., Перего Ю.А. и др. (2012). «Молекулярное рассечение базальных клад филогенетического дерева Y-хромосомы человека» . ПЛОС ОДИН . 7 (11): e49170. Бибкод : 2012PLoSO...749170S . дои : 10.1371/journal.pone.0049170 . ПМЦ 3492319 . ПМИД 23145109 .
- ^ «Блог антропологии Диенекеса: смуглый, голубоглазый, европейский охотник-собиратель с Y-гаплогруппой C из Испании (Olalde et al. 2014)» . 26 января 2014 г.
- ^ http://biorxiv.org/content/biorxiv/early/2015/02/10/013433.full.pdf [ только URL-адрес PDF ]
- ^ Фу, Цяомэй; и др. (2016). «Генетическая история Европы ледникового периода» . Природа . 534 (7606): 200–5. Бибкод : 2016Natur.534..200F . дои : 10.1038/nature17993 . ПМЦ 4943878 . ПМИД 27135931 .
- ^ Jump up to: а б с Гайден, Тензин; Каденас, Алисия М.; Регейру, Мария; Сингх, Северная Каролина; Животовский, Л.А.; Андерхилл, Пенсильвания; Кавалли-Сфорца, LL; Эррера, Р.Дж. (2007). «Гималаи как направленный барьер на пути потока генов» . Американский журнал генетики человека . 80 (5): 884–894. дои : 10.1086/516757 . ПМЦ 1852741 . ПМИД 17436243 .
- ^ Jump up to: а б Форнарино, Симона; Пала, Мария; Батталья, Винченца; и др. (2009). «Митохондриальное и Y-хромосомное разнообразие Таруса (Непал): резервуар генетических вариаций» . Эволюционная биология BMC . 9 (1): 154. Бибкод : 2009BMCEE...9..154F . дои : 10.1186/1471-2148-9-154 . ПМК 2720951 . ПМИД 19573232 .
- ^ Jump up to: а б Каденас, Алисия М; Животовский Лев А; Кавалли-Сфорца, Лука Л; Андерхилл, Пенсильвания; Эррера, Р.Дж. (2008). «Разнообразие Y-хромосом характеризует Оманский залив» . Европейский журнал генетики человека . 16 (3): 374–386. дои : 10.1038/sj.ejhg.5201934 . ПМИД 17928816 .
- ^ Jump up to: а б Абу-Амеро, Халед К.; Хеллани, Али; Гонсалес, Ана М; Ларруга, Хосе М; Кабрера, Висенте М; Андерхилл, Питер А. (2009). «Разнообразие Y-хромосомы Саудовской Аравии и его связь с близлежащими регионами» . БМК Генетика . 10:59 . дои : 10.1186/1471-2156-10-59 . ПМК 2759955 . ПМИД 19772609 .
- ^ Карафет, ТМ; Мендес, Флорида; Мейлерман, МБ; Андерхилл, Пенсильвания; Зегура, СЛ; Хаммер, МФ (2008). «Новые бинарные полиморфизмы изменяют форму и увеличивают разрешение дерева гаплогрупп Y-хромосомы человека» . Геномные исследования . 18 (5): 830–8. дои : 10.1101/гр.7172008 . ПМК 2336805 . ПМИД 18385274 .
- ^ Кокс М.П., Редд А.Дж., Карафет Т.М. и др. (октябрь 2007 г.). «Полинезийский мотив на Y-хромосоме: структура населения в отдаленной Океании». Хм. Биол . 79 (5): 525–35. дои : 10.1353/hub.2008.0004 . hdl : 1808/13585 . ПМИД 18478968 . S2CID 4834817 .
- ^ Андерхилл П.А., Шен П., Лин А.А. и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Нат. Жене . 26 (3): 358–61. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
- ^ Зегура С.Л., Карафет Т.М., Животовский Л.А., Хаммер М.Ф. (январь 2004 г.). «SNP высокого разрешения и микросателлитные гаплотипы указывают на единственное недавнее проникновение Y-хромосом коренных американцев в Америку» . Мол. Биол. Эвол . 21 (1): 164–75. дои : 10.1093/molbev/msh009 . ПМИД 14595095 .
- ^ Пакендорф Б, Новгородов И.Н., Осаковский В.Л., Данилова А.П., Протодьаконов А.П., Стоункинг М. (октябрь 2006 г.). «Исследование последствий доисторических миграций в Сибири: генетическая изменчивость и происхождение якутов». Хм. Жене . 120 (3): 334–53. дои : 10.1007/s00439-006-0213-2 . ПМИД 16845541 . S2CID 31651899 .
- ^ Харьков В.Н., Степанов В.А., Медведев О.Ф. и др. (2008). «[Происхождение якутов: анализ гаплотипов Y-хромосомы]». Мол. Биол. (Моск.) (на русском языке). 42 (2): 226–37. ПМИД 18610830 .
- ^ Jump up to: а б с Борис Малярчук, Мирослава Деренко, Галина Денисова и др. , «Филогеография гаплогруппы С Y-хромосомы в Северной Евразии». Анналы генетики человека (2010) 74 539–546. doi: 10.1111/j.1469–1809.2010.00601.x.
- ^ Манфред Кайзер, Силке Брауэр, Ришар Кордо и др. (2006), «Меланезийское и азиатское происхождение полинезийцев: градиенты мтДНК и Y-хромосомы в Тихом океане». Мол. Биол. Эвол. 23(11):2234–2244. doi:10.1093/molbev/msl093
- ^ Мона, Стефано; Грунц, Катарина Э.; Брауэр, Силке; и др. (2009). «История генетических примесей в Восточной Индонезии, выявленная с помощью анализа Y-хромосомы и митохондриальной ДНК» . Мол. Биол. Эвол . 26 (8): 1865–1877. дои : 10.1093/molbev/msp097 . ПМИД 19414523 .
- ^ Карафет Татьяна Михайловна; Отличительный Брайан; Кокс Мюррей П.; и др. (2010). «Основное разделение Восток-Запад лежит в основе стратификации Y-хромосомы в Индонезии» . Мол. Биол. Эвол . 27 (8): 1833–1844. дои : 10.1093/molbev/msq063 . ПМИД 20207712 . S2CID 4819475 .
- ^ JD Cristofaro et al., 2013, «Афганский Гиндукуш: где сходятся потоки генов на Евразийском субконтиненте», http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0076748
- ^ Jump up to: а б Ван CC, Ван LX, Шреста Р, Чжан М, Хуан XY и др. (2014), «Генетическая структура цянских популяций, проживающих в Западном Сычуаньском коридоре». PLoS ONE 9(8): e103772. doi:10.1371/journal.pone.0103772
- ^ Вэй, Лань-Хай; Ю, Ге; Яо, Да-Ли, Цзинь, Ли; Хуэй (2017). Айсин Гиоро, императорского дома династии Цин». Hum Genet . 62 (62 марта (3)): 407–411. doi : 10.1038/jhg.2016.142 . PMID 27853133. J S2CID 7685248 .
Источники таблиц преобразования
[ редактировать ]- Капелли, Кристиан; Уилсон, Джеймс Ф.; Ричардс, Мартин; Штумпф, Майкл П.Х.; и др. (февраль 2001 г.). «Отцовское наследие преимущественно коренных народов австронезийскоязычных народов островной Юго-Восточной Азии и Океании» . Американский журнал генетики человека . 68 (2): 432–443. дои : 10.1086/318205 . ПМЦ 1235276 . ПМИД 11170891 .
- Хаммер, Майкл Ф.; Карафет Татьяна М.; Редд, Алан Дж.; Джарджанази, Хамди; и др. (1 июля 2001 г.). «Иерархические закономерности глобального разнообразия Y-хромосом человека» . Молекулярная биология и эволюция . 18 (7): 1189–1203. doi : 10.1093/oxfordjournals.molbev.a003906 . ПМИД 11420360 .
- Джоблинг, Марк А.; Тайлер-Смит, Крис (2000), «Новое использование новых гаплотипов», Trends in Genetics , 16 (8): 356–62, doi : 10.1016/S0168-9525(00)02057-6 , PMID 10904265
- Каладжиева, Люба; Калафель, Франческ; Джоблинг, Марк А; Ангеличева, Дора; и др. (февраль 2001 г.). «Закономерности меж- и внутригруппового генетического разнообразия у влакс-рома, выявленные с помощью Y-хромосомы и линий митохондриальной ДНК» . Европейский журнал генетики человека . 9 (2): 97–104. дои : 10.1038/sj.ejhg.5200597 . ПМИД 11313742 . S2CID 21432405 .
- Карафет, Татьяна; Сюй, Липин; Ду, Руофу; Ван, Уильям; и др. (сентябрь 2001 г.). «История отцовского населения Восточной Азии: источники, закономерности и микроэволюционные процессы» . Американский журнал генетики человека . 69 (3): 615–628. дои : 10.1086/323299 . ПМЦ 1235490 . ПМИД 11481588 .
- Семино, О.; Пассарино, Дж; Оефнер, П.Дж.; Лин, А.А.; и др. (2000), «Генетическое наследие палеолитического Homo sapiens sapiens у современных европейцев: хромосомная перспектива AY», Science , 290 (5494): 1155–9, Бибкод : 2000Sci...290.1155S , doi : 10.1126/science.290.5494. 1155 , ПМИД 11073453
- Су, Бинг; Сяо, Цзюньхуа; Андерхилл, Питер; Дека, Ранджан; и др. (декабрь 1999 г.). «Y-хромосома свидетельствует о миграции современных людей на север в Восточную Азию во время последнего ледникового периода» . Американский журнал генетики человека . 65 (6): 1718–1724. дои : 10.1086/302680 . ПМЦ 1288383 . ПМИД 10577926 .
- Андерхилл, Питер А.; Шен, Пейдун; Лин, Алиса А.; Джин, Ли; и др. (ноябрь 2000 г.). «Вариации последовательности Y-хромосомы и история человеческих популяций». Природная генетика . 26 (3): 358–361. дои : 10.1038/81685 . ПМИД 11062480 . S2CID 12893406 .
Внешние ссылки
[ редактировать ]- C3-M217 ФТДНА
- Распространение гаплогруппы C по данным National Geographic.
- https://groups.google.com/forum/#!topic/mintamil/k1GofGgPfXY