Сумчатый
| Сумчатые Временной диапазон: Возможные записи позднего мела | |
|---|---|
 | |
| По часовой стрелке слева: восточный серый кенгуру , вирджинский опоссум , длинноносый бандикут , монито дель монте и тасманский дьявол, представляющие отряды Diprotodontia , Didelphimorphia , Peramelemorphia , Microbiotheria и Dasyuromorphia соответственно. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Клэйд : | Сумчатообразные |
| Инфракласс: | Сумчатые Иллигер , 1811 г. |
| Заказы | |
 | |
| Современное распространение сумчатых. Представлено Родной | |
Сумчатые — разнообразная группа млекопитающих , относящаяся к инфраклассу Marsupialia . Они родом из Австралазии , Уоллесии и Америки . Одной из определяющих особенностей сумчатых является их уникальная репродуктивная стратегия, при которой детеныши рождаются в относительно неразвитом состоянии, а затем вынашиваются в сумке на животе матери.
К современным сумчатым относятся широкий спектр видов, в том числе кенгуру , коалы , опоссумы , тасманские дьяволы , вомбаты , валлаби и бандикуты , среди других.
Сумчатые составляют кладу, происходящую от последнего общего предка современных метатерий , которая охватывает всех млекопитающих, более близких к сумчатым, чем к плацентарным . Этот эволюционный раскол между плацентарными и сумчатыми произошел по меньшей мере 125 миллионов лет назад, возможно, более 160 миллионов лет назад, в период средней юры — раннего мела .
В настоящее время около 70% из 334 существующих видов сумчатых сосредоточены на австралийском континенте, включая материковую Австралию, Тасманию, Новую Гвинею и близлежащие острова. Остальные 30% распространены по всей Америке, в первую очередь в Южной Америке, при этом тринадцать видов обитают в Центральной Америке, а один вид, опоссум Вирджиния, населяет Северную Америку к северу от Мексики.
Размер сумчатых варьируется от нескольких граммов у длиннохвостого планигала . [1] до нескольких тонн у вымершего дипротодона . [2]
Слово «сумчатые» происходит от «marsupium» , технического термина, обозначающего брюшную сумку. Оно, в свою очередь, заимствовано от латинского marsupium и, в конечном итоге, от древнегреческого μάρσιππος mársippos , что означает «мешочек».
Анатомия [ править ]

( Phascolarctos cinereus )
Сумчатые имеют типичные характеристики млекопитающих — например, молочные железы, три косточки среднего уха (и уши, которые обычно имеют траги, [3] разный порог слышимости [4] ) и настоящие волосы . [5] Однако существуют поразительные различия, а также ряд анатомических особенностей, которые отличают их от плаунов .
У большинства самок сумчатых есть передняя сумка , в которой находится несколько сосков для пропитания детенышей. Сумчатые имеют и другие общие черты строения. Оссифицированные надколенники отсутствуют у большинства современных сумчатых (хотя сообщается о небольшом количестве исключений). [6] и надлобковые кости присутствуют. У сумчатых (и однопроходных ) также отсутствует грубая связь ( мозолистое тело ) между правым и левым полушариями мозга. [7]
Череп и зубы [ править ]
Сумчатые имеют отличительные черты черепа по сравнению с плацентарными млекопитающими. Как правило, их черепа относительно маленькие и компактные. Примечательно, что у них есть лобные отверстия, известные как слезное отверстие, расположенные в передней части глазницы. У сумчатых также увеличены скулы, которые простираются дальше назад, а угловое расширение их нижней челюсти (processus angularis) изогнуто внутрь к центру. Твердое небо сумчатых содержит больше отверстий по сравнению с плацентарными млекопитающими.
Зубы сумчатых также существенно отличаются от зубов плацентарных млекопитающих. Например, у большинства австралийских сумчатых, не относящихся к отряду Diprotodontia, между верхней и нижней челюстями разное количество резцов. Ранние сумчатые имели зубную формулу 5.1.3.4/4.1.3.4 на квадрант, состоящую из пяти (верхних) или четырех (нижнечелюстных) резцов, одного клыка, трех премоляров и четырех коренных зубов, всего 50 зубов. В то время как некоторые таксоны, такие как опоссум, сохраняют исходное количество зубов, у других их количество уменьшилось.
Например, представители семейства Macropodidae, включая кенгуру и валлаби, имеют зубную формулу 3/1 – (0 или 1)/0 – 2/2 – 4/4. Многие сумчатые обычно имеют от 40 до 50 зубов, что значительно больше, чем у большинства плацентарных млекопитающих. Примечательно, что у сумчатых второй набор зубов растет только на месте третьего премоляра и сзади; все зубы перед этим прорезываются первоначально как постоянные зубы.
Торс [ править ]
Их скелет описывают несколько общих характеристик. Помимо уникальных деталей строения голеностопного сустава, наблюдаются надлобковые кости ( ossa epubica ), выступающие вперед из лобковой кости таза. Поскольку они присутствуют у самцов и видов, не имеющих мешков, считается, что изначально они не имели ничего общего с размножением, а служили мускулистому подходу к движению задних конечностей. Это можно объяснить своеобразной особенностью млекопитающих, поскольку эти надлобковые кости встречаются и у однопроходных . Репродуктивные органы сумчатых отличаются от плацентарных млекопитающих. У них репродуктивный тракт удваивается. У самок две матки и два влагалища, и перед рождением между ними формируются родовые пути — срединное влагалище. [7] У большинства видов самцы имеют разделенный или двойной половой член, лежащий перед мошонкой. [8] который не гомологичен мошонке плацентарных млекопитающих. [9]
Сумка имеется у большинства, но не у всех видов. У многих сумчатых имеется постоянная сумка, тогда как у других сумка развивается во время беременности, как у опоссума землеройки , у которого детеныши скрыты только складками кожи или в шерсти матери. Расположение сумки варьируется, чтобы обеспечить максимальную защиту потомству. У локомотивных кенгуру отверстие в сумке расположено спереди, в то время как у многих других кенгуру, которые ходят или лазают на четвереньках, отверстие расположено сзади. Обычно сумка есть только у самок, но у самца водного опоссума есть сумка, в которой помещаются его гениталии во время плавания или бега.
Общие сведения и конвергенции [ править ]


Сумчатые приспособились ко многим средам обитания, что отражается в большом разнообразии их телосложения. Самое крупное из ныне живущих сумчатых, красный кенгуру , вырастает до 1,8 метра (5 футов 11 дюймов) в высоту и до 90 килограммов (200 фунтов) в весе, но вымершие виды, такие как Diprotodon , были значительно крупнее и тяжелее. Самыми маленькими представителями этой группы являются сумчатые мыши , длина тела которых часто достигает всего 5 сантиметров (2,0 дюйма).
Некоторые виды напоминают плацентарных млекопитающих и являются примерами конвергентной эволюции . Эта конвергенция очевидна как в эволюции мозга, так и в эволюции мозга. [10] и поведение. [11] Вымерший тилацин сильно напоминал плацентарного волка, отсюда и одно из его прозвищ «тасманский волк». Способность планировать развилась как у сумчатых (как у сахарных планеров ), так и у некоторых плацентарных млекопитающих (как у белок-летяг ), которые развились независимо. Однако другие группы, такие как кенгуру, не имеют явных плацентарных аналогов, хотя в образе жизни и экологических нишах они имеют сходство со жвачными животными .
Температура тела [ править ]
Сумчатые, наряду с однопроходными ( утконосы и ехидны ), обычно имеют более низкую температуру тела, чем плацентарные млекопитающие аналогичного размера ( ейтерианцы ). [12] средняя температура составляет 35 ° C (95 ° F) для сумчатых и 37 ° C (99 ° F) для плацентарных млекопитающих. [13] [14] Некоторые виды будут греться, чтобы сохранить энергию. [15]
Репродуктивная система [ править ]

Репродуктивная система сумчатых заметно отличается от таковой у плацентарных млекопитающих . [16] [17] В процессе эмбрионального развития хориовителлиновая плацента у всех сумчатых формируется . У бандикутов образуется дополнительная хориоаллантоисная плацента , хотя в ней отсутствуют ворсинки хориона, встречающиеся в плацентах плаценты .
Эволюция размножения сумчатых и предположения об изначальном состоянии размножения млекопитающих вызывают дискуссии с конца XIX века. Оба пола обладают клоакой . [17] который связан с мочеполовым мешком, используемым для хранения отходов перед их изгнанием. Мочевой пузырь сумчатых функционирует как место концентрирования мочи и впадает в общий мочеполовой синус как у самок, так и у самцов. [17]
Мужская репродуктивная система [ править ]

Большинство самцов сумчатых, за исключением макроподов. [18] и сумчатые кроты , [19] имеют раздвоенный пенис, разделенный на две колонны, так что пенис имеет два конца, соответствующие двум влагалищам самок. [7] [17] [20] [21] [8] [22] [23] Пенис используется только во время совокупления и отделен от мочевыводящих путей . [8] [17] В вертикальном положении он изгибается вперед, [24] а когда он не выпрямлен, он втягивается в тело по S-образной кривой. [8] Ни сумчатые, ни однопроходные не имеют бакулума . [7] Форма головки полового члена различается у разных видов сумчатых. [8] [25] [26] [27]
У самца тилацина была сумка, которая служила защитной оболочкой, закрывающей его внешние репродуктивные органы во время движения через густые заросли. [28]
Форма уретральных бороздок гениталий самцов используется для различения Monodelphis brevicaudata , Monodelphis Domestica и Monodelphis americana . Борозды образуют 2 отдельных канала, образующих вентральные и дорсальные складки эректильной ткани. [29] Несколько видов сумчатых дасюрид также можно отличить по морфологии пениса. [30] Единственные дополнительные половые железы, которыми обладают сумчатые, — это предстательная и бульбоуретральные железы . [31] Самцы сумчатых имеют 1-3 пары бульбоуретральных желез. [32] отсутствуют . Ампулы семявыносящих протоков , семенных пузырьков и свертывающих желез [33] [20] Простата у сумчатых пропорционально больше, чем у плацентарных млекопитающих. [8] В период размножения простата и бульбоуретральная железа самца таммарского валлаби увеличиваются. Однако, по-видимому, сезонной разницы в весе семенников не наблюдается. [34]
Женская репродуктивная система [ править ]
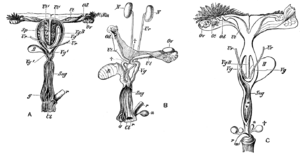
У самок сумчатых есть два боковых влагалища , которые ведут к отдельным маткам , но оба открываются наружу через одно и то же отверстие. Третий канал, срединное влагалище, используется для родов. Этот канал может быть временным или постоянным. [7] Некоторые виды сумчатых способны хранить сперму в яйцеводе после спаривания. [35]
Сумчатые рожают на очень ранней стадии развития; после рождения новорожденные сумчатые ползают вверх по телу своих матерей и прикрепляются к соске, которая находится на нижней стороне матери либо внутри сумки, называемой сумчатым , либо открытой для окружающей среды. Матери часто вылизывают свою шерсть, чтобы оставить запах новорожденного, чтобы увеличить шансы попасть в сумчатое животное. Там они остаются прикрепленными к соске в течение нескольких недель. Потомство в конечном итоге может покидать сумку на короткие периоды времени, возвращаясь к ней за теплом, защитой и питанием. [36] [37]
Раннее развитие [ править ]

различается Пренатальное развитие сумчатых и плацентарных млекопитающих . Ключевые аспекты первых стадий развития эмбрионов плацентарных млекопитающих, такие как внутренняя клеточная масса и процесс уплотнения, у сумчатых не обнаружены. [38] Стадии расщепления развития сумчатых очень различаются между группами, и аспекты раннего развития сумчатых еще не до конца изучены.
Детёныш сумчатого известен как Джоуи . У сумчатых период беременности очень короткий — обычно от 12,5 до 33 дней. [39] но всего 10,7 дней в случае полосатого даннарта и целых 38 дней для длинноносого потору . [40] Джоуи рождается, по существу, в состоянии эмбриона , эквивалентного 8–12-недельному человеческому плоду, слепому, лишенному шерсти и маленькому по сравнению с плацентарными новорожденными, размеры которых варьируются от 4 г до более 800 г. [39] Новорожденного сумчатого можно отнести к одной из трех степеней сложности развития. Наименее развитые при рождении встречаются у дасюридов , промежуточные — у дидельфид и перамелид , наиболее развитые — у макропод . [41] Несмотря на недостаточное развитие, он ползает по шерсти матери, чтобы пробраться в сумку , которая действует как внешняя матка . [42] где он цепляется за соску в поисках еды. Он не появится снова в течение нескольких месяцев, в течение которых он полностью зависит от материнского молока в плане получения необходимых питательных веществ, факторов роста и иммунологической защиты. [43] Гены, экспрессируемые в плаценте плаценты и важные для более поздних стадий развития плода, у самок сумчатых экспрессируются в их молочных железах во время периода лактации. [44] По истечении этого периода джоуи начинает проводить все больше времени вне сумки, питаясь и изучая навыки выживания. Однако он возвращается в сумку, чтобы заснуть, и если ему угрожает опасность, в целях безопасности он ищет убежища в сумке матери.
При раннем рождении развивающееся сумчатое животное удаляется из тела матери гораздо раньше, чем у плацентарных млекопитающих; таким образом, у сумчатых не развилась сложная плацента , защищающая эмбрион матери от иммунной системы . Хотя ранние роды подвергают крошечного новорожденного сумчатого большему экологическому риску, они значительно снижают опасности, связанные с длительной беременностью, поскольку нет необходимости вынашивать крупный плод до полного срока в плохие времена года. Сумчатые — чрезвычайно незрелые животные, о которых необходимо тщательно заботиться сразу после рождения ( см. преждевременное развитие ). У новорожденных сумчатых отсутствуют гистологически зрелые иммунные ткани. [45] [46] [47] и в значительной степени зависят от иммунной системы своей матери в вопросах иммунологической защиты. [48] а также молоко. [36] [37]
Новорожденным сумчатым приходится подниматься к соскам матери, а их передние конечности и лицевые структуры гораздо более развиты, чем остальные части тела на момент рождения. [49] [50] [45] Утверждалось, что это требование привело к ограничению диапазона локомоторных адаптаций у сумчатых по сравнению с плацентарными. У сумчатых в раннем детстве должны развиться хватательные передние лапы, что затрудняет эволюционный переход от этих конечностей к копытам , крыльям или ластам , как это сделали некоторые группы плацентарных млекопитающих. Однако некоторые сумчатые действительно обладают атипичной морфологией передних конечностей, например, копытные передние конечности свиноногого бандикута , что позволяет предположить, что диапазон специализации передних конечностей не так ограничен, как предполагалось. [51]
У некоторых видов Джоуи остаются в сумке до года или до рождения следующего Джоуи. Сумчатый джой не способен регулировать температуру своего тела и полагается на внешний источник тепла. Пока у детеныша не появится хорошая шерсть и он не станет достаточно взрослым, чтобы покинуть сумку, необходимо постоянно поддерживать температуру в сумке 30–32 ° C (86–90 ° F).
Джоуи рождаются с «ротовыми щитками», состоящими из мягких тканей, которые сужают ротовое отверстие до круглого отверстия, достаточно большого, чтобы вместить соску матери. Попав в рот, выпуклая опухоль на конце соска прикрепляет его к потомству, пока оно не вырастет достаточно большим, чтобы его можно было отпустить. У видов без мешочков или с рудиментарными мешочками они более развиты, чем у форм с хорошо развитыми мешочками, что подразумевает повышенную роль в поддержании прикрепления детенышей к соску матери. [52] [53]
География [ править ]
В Австралазии сумчатые встречаются в Австралии, Тасмании и Новой Гвинее; на всей территории Малуккских островов , Тимора и Сулавеси к западу от Новой Гвинеи, а также на архипелаге Бисмарка (включая острова Адмиралтейства ) и Соломоновых островах к востоку от Новой Гвинеи.
В Америке сумчатые встречаются по всей Южной Америке, за исключением центральных/южных Анд и некоторых частей Патагонии ; и через Центральную Америку и южно-центральную Мексику, с единственным видом ( опоссум Вирджинии Didelphis Virginiana ), широко распространенным в восточной части Соединенных Штатов и вдоль побережья Тихого океана.
Взаимодействие с европейцами [ править ]
Первым американским сумчатым (и сумчатым вообще), с которым столкнулся европеец, был обыкновенный опоссум . Висенте Яньес Пинсон , командир корабля « Нинья» во время Христофора Колумба в первого путешествия конце пятнадцатого века, поймал самку опоссума с детенышами в сумке у берегов Южной Америки. Он подарил их испанским монархам, но к тому времени детеныши пропали, а самка умерла. Животное было известно своей странной сумкой или «вторым животом», и то, как потомство достигло сумки, оставалось загадкой. [54] [55]
С другой стороны, именно португальцы первыми описали австралийских сумчатых. Антониу Гальван , португальский администратор в Тернате (1536–1540), написал подробный отчет о северном обыкновенном кускусе ( Phalanger orientalis ): [54]
Некоторые животные напоминают хорьков, только немного крупнее. Их называют Кусу. У них длинный хвост, которым они постоянно свешиваются с деревьев, на которых живут, обматывая его один или два раза вокруг ветки. На брюхе у них имеется карман наподобие промежуточного балкона; как только они рожают детеныша, они выращивают его там же, на соске, до тех пор, пока он не перестанет нуждаться в уходе. Как только она родила и вскормила его, мать снова забеременела.
С начала 17 века поступило больше сообщений о сумчатых. Например, в записи 1606 года о животном, убитом на южном побережье Новой Гвинеи, оно описывается как «имеющее форму собаки, размером меньше борзой», со змееподобным «голым чешуйчатым хвостом» и свисающими яичками. Мясо по вкусу напоминало оленину , а в желудке были листья имбиря. Это описание, по-видимому, очень похоже на темный падебуз ( Thylogale brunii ), и в этом случае это будет самая ранняя европейская запись о члене семейства кенгуру ( Macropodidae ). [56] [54]
Таксономия [ править ]
Сумчатые таксономически идентифицируются как члены млекопитающих инфракласса Marsupialia, впервые описанные как семейство под отрядом Pollicata немецким зоологом Иоганном Карлом Вильгельмом Иллигером в его работе 1811 года Prodromus Systematis Mammalium et Avium . Однако Джеймс Ренни, автор книги «Естественная история обезьян, опоссумов и лемуров» (1838), указывал, что размещение пяти различных групп млекопитающих — обезьян , лемуров , долгопятов , ай-ай и сумчатых (за исключением кенгуру, которые были размещены под приказом Salientia) – под одним приказом (Pollicata), по-видимому, не имели веского обоснования. В 1816 году французский зоолог Жорж Кювье отнес всех сумчатых к отряду Marsupialia. [57] [58] В 1997 году исследователь Дж. А. Кирш и другие присвоили сумчатым ранг инфракласса. [58]
Классификация [ править ]
Сумчатые подразделяются следующим образом: [59] † - Вымерший
- Надотряд Ameridelphia (американские сумчатые)
- Отряд Didelphimorphia (93 вида) - см. список дидельфиморфов.
- Семейство Didelphidae : опоссумы.
- Отряд Paucituberculata (семь видов).
- Семейство Caenolestidae : землероековые опоссумы.
- Отряд Didelphimorphia (93 вида) - см. список дидельфиморфов.
- Надотряд Australidelphia (австралийские сумчатые)
- Отряд Microbiotheria (один современный вид)
- Семейство Microbiotheriidae : monitos del monte.
- Отряд † Yalkaparidontia ( место не определено )
- Грандердер Агреодонтия
- Отряд Dasyuromorphia (73 вида) - см. список дасюроморфов.
- Семейство † Thylacinidae : тилацин.
- Семейство Dasyuridae : антехинусы , кволлы , даннарты , тасманский дьявол и родственники.
- Семейство Myrmecobiidae : намбат.
- Отряд Notoryctemorphia (два вида)
- Семейство Notoryctidae : сумчатые кроты.
- Отряд Peramelemorphia (27 видов).
- Семейство Thylacomyidae : билби.
- Семейство † Chaeropodidae: свиноногие бандикуты.
- Семейство Peramelidae : бандикуты и союзники.
- Отряд Dasyuromorphia (73 вида) - см. список дасюроморфов.
- Отряд Diprotodontia (136 видов) - см. Список дипротодонтов.
- Подотряд Vombatiformes
- Семейство Vombatidae : вомбаты.
- Семейство Phascolarctidae : коалы.
- Семейство † Diprotodontidae.
- Семейство † Palorchestidae : сумчатые тапиры.
- Семейство † Thylacoleonidae : сумчатые львы.
- Подотряд Фалангерида
- Инфраотряд Phalangeriformes - см. список фаланжериобразных.
- Семейство Acrobatidae : планер с перьями и опоссум с перьями.
- Семейство Burramyidae : карликовые опоссумы.
- Семейство † Ektopodontidae : опоссумы-спрайты.
- Семейство Petauridae : полосатый опоссум , опоссум Ледбитера , желтобрюхий планер , сахарный планер , планер красного дерева , беличий планер.
- Семейство Phalangeridae : опоссумы и кускусы.
- Семейство Pseudocheiridae : кольчатые опоссумы и родственники.
- Семейство Tarsipedidae : медовый тростник.
- Инфраотряд Macropodiformes - см. список макроподиформ.
- Семейство Macropodidae : кенгуру , валлаби и их родственники.
- Family Potoroidae : potoroos , rat kangaroos, bettongs
- Семейство Hypsiprymnodontidae : мускусная крыса-кенгуру.
- Семейство † Balbaridae : базальные четвероногие кенгуру.
- Инфраотряд Phalangeriformes - см. список фаланжериобразных.
- Подотряд Vombatiformes
- Отряд Microbiotheria (один современный вид)
Эволюционная история [ править ]
Насчитывая более 300 существующих видов, было предпринято несколько попыток точно интерпретировать филогенетические отношения между различными отрядами сумчатых. Исследования расходятся во мнениях относительно того, является ли Didelphimorphia или Paucituberculata сестринской группой всех других сумчатых. [60] Хотя отряд Microbiotheria (который имеет только один вид, monito del monte ) встречается в Южной Америке, морфологическое сходство позволяет предположить, что он тесно связан с австралийскими сумчатыми. [61] Молекулярный анализ в 2010 и 2011 годах выявил Microbiotheria как родственную группу всем австралийским сумчатым. Однако отношения между четырьмя отрядами австралидельфид не так хорошо изучены.
| Кладограмма Marsupialia Upham et al. 2019 год [62] [63] и Альварес-Карретеро и др. 2022 год [64] [65] |
| Кладограмма Marsupialia Галла и др. 2015 год [60] | ||
|---|---|---|
|

Данные ДНК подтверждают южноамериканское происхождение сумчатых, причем австралийские сумчатые произошли в результате единственной миграции сумчатых из Гондваны из Южной Америки через Антарктиду в Австралию. [66] [67] В каждой группе много мелких древесных видов. Термин « опоссум » используется для обозначения американских видов (хотя «опоссум» является общепринятой аббревиатурой), в то время как аналогичные австралийские виды правильно называются «опоссумами».
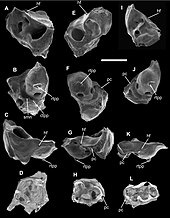

Отношения между тремя существующими отделами млекопитающих ( однопроходные , сумчатые и плацентарные ) долгое время были предметом споров среди систематиков . [69] Большинство морфологических данных, сравнивающих такие черты, как количество и расположение зубов , а также структуру репродуктивной системы и систем удаления отходов , а также большинство генетических и молекулярных данных, свидетельствуют в пользу более тесной эволюционной связи между сумчатыми и плацентарными млекопитающими, чем с однопроходными. [70]

Предки сумчатых, входящие в более крупную группу, называемую метатериями , вероятно, отделились от предков плацентарных млекопитающих ( эутерий ) в середине юрского периода, хотя никаких ископаемых свидетельств существования самих метатерий с этого времени не известно. [71] По данным анализа ДНК и белков, время расхождения двух линий оценивается примерно в 100–120 млн лет назад . [54] Ископаемые метатерии отличаются от эвтерий формой зубов; метатерии обладают четырьмя парами коренных зубов на каждой челюсти, тогда как у плацентарных млекопитающих (включая настоящие плацентарные) никогда не бывает более трех пар. [72] Используя этот критерий, самым ранним известным метатерием считался Sinodelphys szalayi , который жил в Китае около 125 млн лет назад. [73] [74] [75] Однако позже Синодельфис был интерпретирован как один из первых членов Эутерии . Самыми древними из известных метатерий в настоящее время являются окаменелости возрастом 110 миллионов лет из западной части Северной Америки. [76] Метатерийцы были широко распространены в Северной Америке и Азии в позднем меловом периоде, но их численность резко сократилась во время вымирания в конце мелового периода. [77]
Кладограмма от Wilson et al. (2016) [78]
| Метатерия |
| ||||||||||||
В 2022 году исследование предоставило убедительные доказательства того, что самым ранним из известных сумчатых был Deltatheridium, известный по образцам кампанского периода позднего мела в Монголии. [79] В этом исследовании Deltatheridium и Pucadelphys были отнесены к родственным таксонам современных крупных американских опоссумов .
Сумчатые распространились в Южную Америку из Северной Америки в палеоцене , возможно, через хребет Авес . [80] [81] [82] Метатерии Северного полушария, имевшие низкое морфологическое и видовое разнообразие по сравнению с современными плацентарными млекопитающими, в конечном итоге вымерли в эпоху миоцена . [83]
В Южной Америке опоссумы эволюционировали и получили широкое распространение, а в палеогене также наблюдалась эволюция землероек-опоссумов (Paucituberculata) наряду с несумчатыми метатерийными хищниками, такими как borhyaenids и саблезубый Thylacosmilus . В южноамериканских нишах хищных млекопитающих доминировали эти сумчатые и спарассодонтные метатерии, которые, по-видимому, конкурентно исключили южноамериканских плацентарных животных из развития хищных животных. [84] В то время как плацентарные хищники отсутствовали, метатериям приходилось бороться с конкуренцией среди птиц ( террористических птиц ) и наземных крокодиломорфов. Сумчатые, в свою очередь, были исключены из крупных ниш травоядных в Южной Америке из-за присутствия местных плацентарных копытных (ныне вымерших) и ксенартранов (самые крупные формы которых также вымерли). Южная Америка и Антарктида оставались соединенными до 35 млн лет назад, о чем свидетельствуют найденные там уникальные окаменелости. Северная и Южная Америка были разъединены примерно три миллиона лет назад, когда образовался Панамский перешеек . Это привело к Великому американскому обмену . Спарассодонты исчезли по неясным причинам – опять же, классически это предполагалось как конкуренция со стороны плацентарных хищников, но последние спарассодонты сосуществовали с несколькими мелкими хищниками, такими как проциониды и собаки, и исчезли задолго до появления макрохищных форм, таких как кошачьи. [85] в то время как дидельфиморфы (опоссумы) вторглись в Центральную Америку, а опоссумы из Вирджинии достигли севера вплоть до Канады.
Сумчатые прибыли в Австралию через Антарктиду в раннем эоцене, около 50 млн лет назад, вскоре после того, как Австралия отделилась. [n 1] [n 2] Это предполагает единичное распространение только одного вида, скорее всего, родственника южноамериканского монито дель монте ( микробиотера , единственного австралидельфии Нового Света ). Этот прародитель, возможно, пересек расширяющуюся, но все еще узкую пропасть между Австралией и Антарктидой. Путешествие, должно быть, было нелегким; Южноамериканское копытное [89] [90] [91] и ксенартран [92] останки были найдены в Антарктиде, но эти группы не достигли Австралии.
В Австралии сумчатые превратились в широкое разнообразие, наблюдаемое сегодня, включая не только всеядных и плотоядных форм, которые присутствовали в Южной Америке, но и крупных травоядных. Современные сумчатые, судя по всему, относительно недавно достигли островов Новой Гвинеи и Сулавеси через Австралию. [93] [94] [95] Анализ ретропозонов сайтов вставки в ядерной ДНК различных сумчатых в 2010 году подтвердил, что все ныне живущие сумчатые имеют южноамериканских предков. Последовательность ветвления отрядов сумчатых, указанная в исследовании, ставит Didelphimorphia в самое базальное положение, за ней следует Paucituberculata, затем Microbiotheria и заканчивается радиация австралийских сумчатых. Это указывает на то, что австралидельфия возникла в Южной Америке и достигла Австралии после отделения микробиотерии. [66] [67]
В Австралии наземные плацентарные млекопитающие исчезли в начале кайнозоя ( их самые последние известные окаменелости представляют собой зубы возрастом 55 миллионов лет, напоминающие зубы мыщелков ) по неясным причинам, что позволило сумчатым доминировать в австралийской экосистеме. [93] Существующие местные австралийские наземные плацентарные млекопитающие (такие как прыгающие мыши ) являются относительно недавними иммигрантами, прибывшими через острова из Юго-Восточной Азии. [94]
Генетический анализ предполагает, что расхождение сумчатых и плацентарных произошло 160 миллионов лет назад . [96] Предковое число хромосом оценивается в 2n = 14.
Недавняя гипотеза предполагает, что южноамериканские микробиотеры возникли в результате обратного расселения из восточной Гондваны. Эта интерпретация основана на новых черепных и посткраниальных окаменелостях сумчатых Djarthia murgonensis из раннеэоценовой местной фауны Тингамарры в Австралии, которые указывают на то, что этот вид является наиболее плезиоморфным предком, старейшим однозначным австралидельфийцем и может быть предковым морфотипом австралийских сумчатых. радиация. [68]
В 2023 году визуализация частичного скелета, найденного в Австралии палеонтологами из Университета Флиндерса, привела к идентификации Ambulator keanei , первого пешехода на длинные дистанции в Австралии. [97]
См. также [ править ]
- Сумчатый газон
- Метатерия
- Список родов млекопитающих
- Список недавно вымерших млекопитающих
- Список доисторических млекопитающих
Примечания [ править ]
- ^ Это подтверждается находкой эоценовых ископаемых останков австралидельфии, микробиотерия Woodburnodon Casei , на Антарктическом полуострове . [86]
- ^ Ратиты , возможно, также путешествовали по суше из Южной Америки, чтобы колонизировать Австралию; [87] ископаемый бескилевой известен из Антарктиды, [88] и южноамериканские реи являются более базальными внутри группы, чем австралийско-тихоокеанские бескилевые. [87]
Ссылки [ править ]
- ^ Бейкер, Эндрю М. (27 апреля 2021 г.). «Познакомьтесь с пятью мельчайшими млекопитающими Австралии, которые каждую ночь идут по канату между жизнью и смертью» . Разговор . Проверено 29 мая 2024 г.
- ^ Роу, С.; Кроутер, М.; Дортч, Дж.; Чонг, Дж. (2004). «Размер самого крупного сумчатого животного и почему это важно» . Труды Королевского общества B: Биологические науки . 271 (Приложение 3): S34–S36. дои : 10.1098/rsbl.2003.0095 . ПМК 1810005 . ПМИД 15101412 .
- ^ Стэннард Х.Дж., Деннингтон К., Олд Дж.М. (2020). «Морфология наружного уха и наличие траги у австралийских сумчатых» . Экология и эволюция . 10 : 9853–9866. дои : 10.1002/ece3.6634 . ПМК 7520188 .
- ^ Олд Дж. М., Парсонс С. Л., Талк, М. Л. (2020). «Порог слуха мелких местных австралийских млекопитающих — краснохвостых фаскогалов ( Phascogale Calura ), культарров ( Antechinomys laniger ) и прыгающих мышей-спинифексов ( Notomys Alexis )». Зоологический журнал Линнеевского общества . 190 : 342–351. doi : 10.1093/zoolinnean/zlaa003 .
- ^ Фельдхамер Г.А., Дрикамер Л.С., Весси С.Х., Мерритт Дж.Ф., Краевски С. (2007). Маммология: адаптация, разнообразие, экология (3-е изд.). Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-8695-9 . OCLC 124031907 .
- ^ Сэмюэлс М.Э., Рено С., Хатчинсон-младший (2017). «Эволюция сесамовидной кости надколенника у млекопитающих» . ПерДж . 5 : е3103. дои : 10.7717/peerj.3103 . ПМЦ 5363259 . ПМИД 28344905 .
- ^ Jump up to: Перейти обратно: а б с д и Новак 1999 .
- ^ Jump up to: Перейти обратно: а б с д и ж Ренфри М., Тиндейл-Биско Х. (1987). Репродуктивная физиология сумчатых . Издательство Кембриджского университета. ISBN 9780521337922 .
- ^ Армати, Патрисия Дж.; Дикман, Крис Р.; Хьюм, Ян Д. (17 августа 2006 г.). Сумчатые . Издательство Кембриджского университета. ISBN 978-1-139-45742-2 .
- ^ Тодоров О.С., Бломберг С.П., Госвами А., Сирс К., Дрлик П., Петерс Дж., Вайсбекер В. (март 2021 г.). «Проверка гипотез об изменении размера мозга сумчатых с использованием множественных филогенетических вычислений и байесовской сравнительной структуры» . Слушания. Биологические науки . 288 (1947): 20210394. doi : 10.1098/rspb.2021.0394 . ПМК 8059968 . ПМИД 33784860 .
- ^ Тодорова ОС (2019). «Сумчатое познание». В Вонк Дж, Шакелфорд Т. (ред.). Энциклопедия познания и поведения животных . Чам: Международное издательство Springer. стр. 1–8. дои : 10.1007/978-3-319-47829-6_1167-1 . ISBN 978-3-319-47829-6 . S2CID 242256517 .
- ^ Гоган, Джон Б.; Хоган, Линдси А.; Уолледж, Андреа (2015). Аннотация: Терморегуляция у сумчатых и однопроходных, глава Сумчатые и однопроходные: загадочные млекопитающие природы . Издательство Nova Science, Incorporated. ISBN 9781634834872 . Проверено 20 апреля 2022 г.
- ^ Белый (1999). «Тепловая биология утконоса» . Дэвидсон Колледж. Архивировано из оригинала 6 марта 2012 года . Проверено 14 сентября 2006 г.
- ^ «Системы управления, часть 2» (PDF) . Архивировано из оригинала (PDF) 8 октября 2016 года . Проверено 6 июля 2016 г.
- ^ Стэннард, Хейли Дж.; Фабиан, Меган; Олд, Джули М. (2015). «Гряться или не греться: поведенческая терморегуляция у двух видов дасюрид, Phascogale Calura и Antechinomys laniger ». Журнал термической биологии . 53 : 66–71. дои : 10.1016/j.jtherbio.2015.08.012 . ПМИД 26590457 .
- ^ Короткометражный автофургон, Балабан Э (1994). Различия между полами . Издательство Кембриджского университета. ISBN 978-0-521-44878-9 .
- ^ Jump up to: Перейти обратно: а б с д и Король А (2001). «Открытия о размножении сумчатых» . Факультет биологии Университета штата Айова. Архивировано из оригинала 5 сентября 2012 года . Проверено 22 ноября 2012 г.
- ^ Стакер Л (30 июня 2014 г.). Разведение макропод, здравоохранение и лекарственные препараты – тома первый и второй . Линда Стейкер. ISBN 978-0-9775751-2-1 .
- ^ О привычках и сходстве нового австралийского млекопитающего Notoryctes typlops. ЭД Коуп Американский натуралист Том. 26, № 302 (февраль 1892 г.), стр. 121–128.
- ^ Jump up to: Перейти обратно: а б Хунсакер II D (1977). Биология сумчатых . Эльзевир Наука. ISBN 978-0-323-14620-3 .
- ^ Сетчелл Б.П. (1977). «Размножение самцов сумчатых». Биология сумчатых . стр. 411–457. дои : 10.1007/978-1-349-02721-7_24 . ISBN 978-1-349-02723-1 .
- ^ Шарман ГБ, Пилтон П.Е. (1964). «История жизни и размножение рыжего кенгуру ( Megaleia rufa )». Труды Лондонского зоологического общества . 142 (1): 29–48. дои : 10.1111/j.1469-7998.1964.tb05152.x .
- ^ Садлер Р.М. (1965). «Размножение двух видов кенгуру ( Macropusrobustus и Megaleia rufa в засушливом регионе Пилбара в Западной Австралии». Труды Лондонского зоологического общества . 145 (2): 239–261. doi : 10.1111/j.1469-7998.1965. tb02016.x .
- ^ Садлер Р. (1973). Размножение позвоночных . Эльзевир Наука. ISBN 978-0-323-15935-7 .
- ^ Австралийское общество млекопитающих (1978). Австралийское общество млекопитающих . Австралийское общество млекопитающих. стр. 73–.
- ^ Осгуд WH, Херрик CJ (1921). Монографическое исследование американского сумчатого Caenolestes ... Чикагский университет. стр. 64–.
- ^ Урологический и кожный обзор . Урологическая и кожная пресса. 1920. стр. 677–.
- ^ Весло Р. (2002). Последний тасманский тигр: история и исчезновение тилацина (изд. В мягкой обложке). Порт Мельбурн, Вика: Издательство Кембриджского университета. ISBN 978-0-521-53154-2 .
- ^ Ногейра Дж., Кастро А.С., Камара ЕС, Камара Б.О. (2004). «Морфология мужской половой системы Chironectes minimus и сравнение с другими дидельфидными сумчатыми». Журнал маммологии . 85 (5): 834–841. дои : 10.1644/207 . S2CID 85595933 .
- ^ Вулли П.А., Вестерман М., Краевски С. (декабрь 2007 г.). «Межвидовое сходство внутри рода Sminthopsis (Dasyuromorphia: Dasyuridae) на основе морфологии полового члена: соответствие с другими анатомическими и молекулярными данными» . Журнал маммологии . 88 (6): 1381–1392. doi : 10.1644/06-мамм-а-443р.1 . ISSN 0022-2372 .
- ^ Роджер Дж.К., Хьюз Р.Л. (1973). «Исследование дополнительных желез самцов сумчатых» . Австралийский журнал зоологии . 21 (3): 303. doi : 10.1071/ZO9730303 . hdl : 1959.4/70011 .
- ^ Фогельнест Л., Портас Т. (1 мая 2019 г.). Современная терапия в медицине австралийских млекопитающих . Издательство Csiro. ISBN 978-1-4863-0753-1 .
- ^ Роджер Дж. К. (январь 1976 г.). «Сравнительные аспекты дополнительных половых желез и биохимии семенной жидкости млекопитающих». Сравнительная биохимия и физиология. Б. Сравнительная биохимия . 55 (1): 1–8. дои : 10.1016/0305-0491(76)90164-4 . ПМИД 780045 .
- ^ Иннс RW (ноябрь 1982 г.). «Сезонные изменения в дополнительной репродуктивной системе и уровнях тестостерона в плазме самцов таммарского валлаби Macropus eugenii в дикой природе» . Журнал репродукции и фертильности . 66 (2): 675–680. дои : 10.1530/jrf.0.0660675 . ПМИД 7175821 .
- ^ Завод ТМ, Железник А.Я. (15 ноября 2014 г.). Физиология репродукции Кнобила и Нила . Академическая пресса. ISBN 978-0-12-397769-4 .
- ^ Jump up to: Перейти обратно: а б Стэннард, Хейли Дж.; Олд, Джули М. (2023). «Валлаби-джои и мопсы-утконосы рождаются крошечными и неразвитыми. Но молоко их матери почти волшебное» . Разговор.
- ^ Jump up to: Перейти обратно: а б Стэннард, Хейли Дж.; Миллер, Роберт Д.; Олд, Джули М. (2020). «Молоко сумчатых и однопроходных – обзор его питательных веществ и иммунных свойств» . ПерДж . 8 : е9335. дои : 10.7717/peerj.9335 . ПМК 7319036 . ПМИД 32612884 .
- ^ Франкенберг С.Р., де Баррос Ф.Р., Россант Дж., Ренфри М.Б. (2016). «Бластоциста млекопитающих». Междисциплинарные обзоры Wiley. Биология развития . 5 (2): 210–232. дои : 10.1002/wdev.220 . ПМИД 26799266 . S2CID 22001725 .
- ^ Jump up to: Перейти обратно: а б Смит, Кэтлин К.; Кейте, Анна Л. (2020). «Адаптации новорожденного сумчатого: рождение как экстремальная среда» . Анатомическая запись . 303 (2): 235–249. дои : 10.1002/ar.24049 . ПМИД 30548826 . S2CID 56484546 .
- ^ Дрюс, Барбара; Руллиг, Кэтлин; Мензис, Брэндон Р.; Шоу, Джефф; Буентхен, Ина; Герберт, Кэтрин А.; Хильдебрандт, Томас Б.; Ренфри, Мэрилин Б. (15 марта 2013 г.). «Ультрасонография пренатального развития валлаби показывает, что восхождение к сумке начинается внутриутробно» . Научные отчеты . 3 (1458): 1458. Бибкод : 2013NatSR...3E1458D . дои : 10.1038/srep01458 . ПМЦ 3597997 . ПМИД 23492830 .
- ^ Шнайдер, Нью-Йорк; Гурович Ю. (2017). «Морфология и эволюция ротового щитка у новорожденных сумчатых, включая новорожденного мешка монито дель монте (Dromiciops gliroides, Marsupialia Microbiotheria) молодых - PMC» . Журнал анатомии . 231 (1): 59–83. дои : 10.1111/joa.12621 . ПМЦ 5472534 . ПМИД 28620997 .
- ^ Докинз, Ричард; Вонг, Ян (9 февраля 2005 г.). Рассказ предка: паломничество на зарю эволюции . Хоутон Миффлин Харкорт. ISBN 9780618619160 – через Google Книги.
- ^ Стэннард Х.Дж., Миллер Р.Д., Олд Дж.М. (2020). «Молоко сумчатых и однопроходных – обзор его питательных веществ и иммунных свойств» . ПерДж . 8 : е9335. дои : 10.7717/peerj.9335 . ПМК 7319036 . ПМИД 32612884 .
- ^ [электронная почта защищена] , <img src=' https://med stanford edu/news/media-contacts/krista_conger/_jcr_content/image img 620 high jpg/conger-krista-90 jpg' alt='Криста Конгер'> Криста Конгер Криста Конгер — старший научный обозреватель в Управлении по связям с общественностью. Напишите ей по адресу. «Детеныши сумчатых «пьют» плаценту для ускорения развития» . Центр новостей .
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ) - ^ Jump up to: Перейти обратно: а б Старый Дж.М., Дин Э.М. (2003). «Лимфоидная и иммуногематопоэтическая ткань эмбрионального опоссума ( Trichosurus vulpecula )». Анатомия и эмбриология . 206 (3): 193–197. дои : 10.1007/s00429-002-0285-2 . ПМИД 12592570 . S2CID 546795 .
- ^ Олд Дж. М., Селвуд Л., Дин Э. М. (2003). «Гистологическое исследование лимфоидной и иммуногемопоэтической тканей взрослого полосатого даннарта ( Sminthopsis macroura )». Клетки Ткани Органы . 173 (2): 115–121. дои : 10.1159/000068946 . ПМИД 12649589 . S2CID 46354564 .
- ^ Олд Дж. М., Селвуд Л., Дин Э. М. (2003). «Развитие лимфоидной ткани полосатого даннарта ( Sminthopsis macroura )» . Клетки Ткани Органы . 175 (4): 192–201. дои : 10.1111/j.0021-8782.2004.00310.x . ПМК 1571326 . ПМИД 15255959 .
- ^ Старый Дж. М., Дин Э. М. (2000). «Развитие иммунной системы и иммунологической защиты у молодняка сумчатой сумки». Развивающая и сравнительная иммунология . 24 (5): 445–454. дои : 10.1016/S0145-305X(00)00008-2 . ПМИД 10785270 .
- ^ Sears KE (август 2009 г.). «Различия во времени прехондрогенного развития конечностей у млекопитающих: решена сумчато-плацентарная дихотомия» . Эволюция; Международный журнал органической эволюции . 63 (8): 2193–2200. дои : 10.1111/j.1558-5646.2009.00690.x . ПМИД 19453378 . S2CID 42635687 .
- ^ Смит К.К. (2001). «Раннее развитие нервной пластинки, нервного гребня и лицевой области сумчатых» . Журнал анатомии . 199 (Части 1–2): 121–131. дои : 10.1046/j.1469-7580.2001.19910121.x . ПМК 1594995 . ПМИД 11523813 .
- ^ Ларри Фогельнест, Грэм Аллан, Радиология австралийских млекопитающих
- ^ Шнайдер, штат Нью-Йорк (август 2011 г.). «Развитие органов обоняния у только что вылупившихся однопроходных и новорожденных сумчатых» . Журнал анатомии . 219 (2): 229–242. дои : 10.1111/j.1469-7580.2011.01393.x . ПМК 3162242 . ПМИД 21592102 .
- ^ Фернер, К.; Шульц, Дж. А.; Целлер, У. (2017). «Сравнительная анатомия новорожденных трех основных групп млекопитающих (однопроходные, сумчатые, плацентарные) и значение морфотипа предкового новорожденного млекопитающих - PMC» . Журнал анатомии . 231 (6): 798–822. дои : 10.1111/joa.12689 . ПМК 5696127 . ПМИД 28960296 .
- ^ Jump up to: Перейти обратно: а б с д Тиндейл-Биско Х (2004). Жизнь сумчатых . Коллингвуд, Австралия: CSIRO. ISBN 978-0-643-06257-3 .
- ^ Краузе В.Дж., Краузе В.А. (2006). Опоссум: его удивительная история . Колумбия, США: Кафедра патологии анатомических наук, Медицинский факультет Университета Миссури. п. 6. ISBN 978-0-9785999-0-4 .
- ^ Доусон Т.Дж. (2012). Кенгуру (2-е изд.). Коллингвуд, США: Издательство CSIRO. п. 181. ИСБН 978-0-643-10625-3 .
- ^ Мартин В.К. (1841 г.). Общее введение в естественную историю млекопитающих . Лондон, Великобритания: Принтеры Wright and Co. стр. 182–4.
- ^ Jump up to: Перейти обратно: а б Джексон С., Гроувс С. (2015). Систематика австралийских млекопитающих . Австралия: Издательство CSIRO. стр. 82–3. ISBN 978-1-4863-0014-3 .
- ^ Гарднер, А. (2005). Уилсон, Делавэр ; Ридер, Д.М. (ред.). Виды млекопитающих мира: таксономический и географический справочник (3-е изд.). Издательство Университета Джонса Хопкинса. стр. 3–21. ISBN 978-0-8018-8221-0 . OCLC 62265494 .
- ^ Jump up to: Перейти обратно: а б Галлус С., Янке А., Кумар В., Нильссон М.А. (март 2015 г.). «Распутывание взаимоотношений австралийских отрядов сумчатых с использованием анализа ретротранспозонов и эволюционных сетей» . Геномная биология и эволюция . 7 (4): 985–992. дои : 10.1093/gbe/evv052 . ПМЦ 4419798 . ПМИД 25786431 .
- ^ Салай Ф (1982). Арчер М. (ред.). «Новая оценка филогении и классификации сумчатых». Плотоядные сумчатые . 2 : 621–40.
- ^ Апхэм, Натан С.; Эссельстин, Джейкоб А.; Джетц, Уолтер (2019). «Вывод о дереве млекопитающих: наборы филогений на уровне видов для вопросов экологии, эволюции и сохранения» . ПЛОС Биол . 17 (12): e3000494. дои : 10.1371/journal.pbio.3000494 . ПМК 6892540 . ПМИД 31800571 .
- ^ Апхэм, Натан С.; Эссельстин, Джейкоб А.; Джетц, Уолтер (2019). «DR_on4phylosCompared_linear_richCol_justScale_ownColors_withTips_80in» (PDF) . ПЛОС Биология . 17 (12). дои : 10.1371/journal.pbio.3000494 . ПМИД 31800571 .
- ^ Альварес-Карретеро, Сандра; Тамури, Асиф У.; Баттини, Маттео; Насименто, Фабрисия Ф.; Карлайл, Эмили; Ашер, Роберт Дж.; Ян, Цзыхэн; Донохью, Филип CJ; дос Рейс, Марио (2022). «Хронология эволюции млекопитающих на уровне вида, объединяющая филогеномные данные» . Природа . 602 (7896): 263–267. Бибкод : 2022Natur.602..263A . дои : 10.1038/s41586-021-04341-1 . hdl : 1983/de841853-d57b-40d9-876f-9bfcf7253f12 . ПМИД 34937052 . S2CID 245438816 .
- ^ Альварес-Карретеро, Сандра; Тамури, Асиф У.; Баттини, Маттео; Насименто, Фабрисия Ф.; Карлайл, Эмили; Ашер, Роберт Дж.; Ян, Цзыхэн; Донохью, Филип CJ; дос Рейс, Марио (2022). "4705sp_colours_mammal-time.tree" . Природа . 602 (7896): 263–267. Бибкод : 2022Natur.602..263A . дои : 10.1038/s41586-021-04341-1 . hdl : 1983/de841853-d57b-40d9-876f-9bfcf7253f12 . ПМИД 34937052 . S2CID 245438816 .
- ^ Jump up to: Перейти обратно: а б Шиве Дж. (28 июля 2010 г.). «Австралийские сумчатые произошли на территории нынешней Южной Америки, говорится в исследовании» . Лос-Анджелес Таймс . Архивировано из оригинала 1 августа 2010 года . Проверено 1 августа 2010 г.
- ^ Jump up to: Перейти обратно: а б с Нильссон М.А., Чураков Г., Зоммер М., Тран Н.В., Земанн А., Брозиус Дж., Шмитц Дж. (июль 2010 г.). «Отслеживание эволюции сумчатых с помощью архаичных геномных вставок ретропозонов» . ПЛОС Биология . 8 (7): e1000436. дои : 10.1371/journal.pbio.1000436 . ПМЦ 2910653 . ПМИД 20668664 .
- ^ Jump up to: Перейти обратно: а б Бек Р.М., Годтелп Х., Вайсбекер В., Арчер М., Хэнд С.Дж. (март 2008 г.). Хоукс Дж. (ред.). «Старейшие окаменелости сумчатых в Австралии и их биогеографическое значение» . ПЛОС ОДИН . 3 (3): e1858. Бибкод : 2008PLoSO...3.1858B . дои : 10.1371/journal.pone.0001858 . ПМК 2267999 . ПМИД 18365013 .
- ^ Мойал А.М. (2004). Утконос: Необыкновенная история о том, как любопытное существо поставило мир в тупик . Балтимор: Издательство Университета Джона Хопкинса. ISBN 978-0-8018-8052-0 .
- ^ ван Рид Т., Бастианс Т., Бун Д.Н., Хеджес С.Б., де Йонг В.В., Мэдсен О. (март 2006 г.). «Утконос на своем месте: ядерные гены и инделы подтверждают родство сестринской группы однопроходных и терианцев» . Молекулярная биология и эволюция . 23 (3): 587–597. дои : 10.1093/molbev/msj064 . ПМИД 16291999 .
- ^ Луо ZX, Юань CX, Мэн QJ, Цзи Q (август 2011 г.). «Юрское плацентарное млекопитающее и дивергенция сумчатых и плацентарных». Природа . 476 (7361): 442–445. Бибкод : 2011Natur.476..442L . дои : 10.1038/nature10291 . ПМИД 21866158 . S2CID 205225806 .
- ^ Бентон М.Дж. (1997). Палеонтология позвоночных . Лондон: Чепмен и Холл. п. 306. ИСБН 978-0-412-73810-4 .
- ^ Ринкон П. (12 декабря 2003 г.). «Найден древнейший предок сумчатых» . Новости Би-би-си . Проверено 16 марта 2010 г.
- ^ Луо ZX, Цзи Q, Wible JR, Юань CX (декабрь 2003 г.). «Раннемеловое трибосфеническое млекопитающее и метатерийская эволюция». Наука . 302 (5652): 1934–1940. Бибкод : 2003Sci...302.1934L . дои : 10.1126/science.1090718 . ПМИД 14671295 . S2CID 18032860 .
- ^ Ху Ю, Мэн Дж, Ли С, Ван Ю (январь 2010 г.). «Новое базальное млекопитающее из раннемеловой биоты Джехоль, Ляонин, Китай» . Слушания. Биологические науки . 277 (1679): 229–236. дои : 10.1098/rspb.2009.0203 . ПМЦ 2842663 . ПМИД 19419990 .
- ^ Би С., Чжэн Икс, Ван Икс, Чигнетти Н.Э., Ян С., Вибл-младший (июнь 2018 г.). «Раннемеловой эвтерий и плацентарно-сумчатая дихотомия». Природа . 558 (7710): 390–395. Бибкод : 2018Natur.558..390B . дои : 10.1038/s41586-018-0210-3 . ПМИД 29899454 . S2CID 49183466 .
- ^ Беннетт, К. Верити; Апчерч, Пол; Гойн, Франциско Дж.; Госвами, Анджали (6 февраля 2018 г.). «Глубинное временное разнообразие метатерийных млекопитающих: значение для истории эволюции и качества летописи окаменелостей» . Палеобиология . 44 (2): 171–198. Бибкод : 2018Pbio...44..171B . дои : 10.1017/pab.2017.34 . hdl : 11336/94590 . ISSN 0094-8373 . S2CID 46796692 .
- ^ Уилсон, врач общей практики; Экдейл, Э.Г.; Хогансон, Дж.В.; Каледе, Джей Джей; Линден, А.В. (2016). « Крупное хищное млекопитающее позднего мела и североамериканского происхождения сумчатых ». Природные коммуникации. 7. дои : 10.1038/ncomms13734 .
- ^ Веласко, Пол М; Бучек, Александра Дж; Хоффман, Ева; Хоффман, Девин К; О'Лири, Морин А; Новачек, Майкл Дж. (30 января 2022 г.). «Комбинированный анализ данных ископаемых и живых млекопитающих: родственный палеогеновый таксон Placentalia и древность Marsupialia» . Кладистика . 38 (3): 359–373. дои : 10.1111/cla.12499 . ПМИД 35098586 . S2CID 246429311 .
- ^ Кемп, Томас Стейнфорт (2005). Происхождение и эволюция млекопитающих (PDF) . Оксфорд: Издательство Оксфордского университета. п. 217. ИСБН 0-19-850760-7 .
- ^ Бошман Л.М., ван Хинсберген DJ, Торсвик Т.Х., Спакман В., Пинделл Дж.Л. (23 августа 2014 г.). «Кинематическая реконструкция Карибского региона начиная с ранней юры». Обзоры наук о Земле . 138 : 102–136. Бибкод : 2014ESRv..138..102B . CiteSeerX 10.1.1.727.4858 . doi : 10.1016/j.earscirev.2014.08.007 .
- ^ Али, Джейсон Р.; Хеджес, С. Блэр (ноябрь 2021 г.). Хорн, Карина (ред.). «Колонизация Карибского бассейна: новые геологические данные и обновленные данные о колонизации наземных позвоночных бросают вызов гипотезе сухопутного моста GAARlandia» . Журнал биогеографии . 48 (11): 2699–2707. дои : 10.1111/jbi.14234 . ISSN 0305-0270 . S2CID 238647106 .
- ^ Элдридж, Марк Д.Б.; Бек, Робин, доктор медицины; Крофт, Дарин А; Травуийон, Кенни Дж; Фокс, Барри Дж. (23 мая 2019 г.). «Новый консенсус в эволюции, филогении и систематике сумчатых и их ископаемых родственников (Metatheria)» . Журнал маммологии . 100 (3): 802–837. дои : 10.1093/jmammal/gyz018 . ISSN 0022-2372 .
- ^ Симпсон Г.Г. (июль 1950 г.). «История фауны Латинской Америки» . Американский учёный . 38 (3): 361–389, см. с. 368. JSTOR 27826322 . Проверено 21 января 2020 г.
- ^ Превости Ф.Дж., Форасепи А., Зимич Н. (2011). «Эволюция кайнозойской гильдии хищников наземных млекопитающих в Южной Америке: конкуренция или замена?». Журнал эволюции млекопитающих . 20 :3–21. дои : 10.1007/s10914-011-9175-9 . hdl : 11336/2663 . S2CID 15751319 .
- ^ Гоин Ф.Дж., Зимич Н., Регеро М.А., Сантильяна С.Н., Маренси С.А., Моли Дж.Дж. (2007). «Новые сумчатые (млекопитающие) эоцена Антарктиды, а также происхождение и родство микробиотерий » Журнал Аргентинской геологической ассоциации . 62 (4): 597–603 . Получено 17 июля.
- ^ Jump up to: Перейти обратно: а б Ёнезава Т., Сегава Т., Мори Х., Кампос П.Ф., Хонго Ю., Эндо Х. и др. (январь 2017 г.). «Филогеномика и морфология вымерших палеогнат раскрывают происхождение и эволюцию бескилевых» . Современная биология . 27 (1): 68–77. дои : 10.1016/j.cub.2016.10.029 . ПМИД 27989673 .
- ^ Тамбусси КП, Норьега Дж.И., Гадзицки А., Татур А., Регуэро М.А., Вискайно С.Ф. (1994). «Ратитовая птица из палеогеновой формации Ла-Месета, остров Сеймур, Антарктида» (PDF) . Польские полярные исследования . 15 (1–2): 15–20. Архивировано из оригинала (PDF) 28 декабря 2019 года . Проверено 28 декабря 2019 г.
- ^ Бонд М., Регеро М.А., Вискайно С.Ф., Маренси С.А. (2006). «Новое южноамериканское копытное животное (Mammalia: Litopterna) из эоцена Антарктического полуострова» . У Фрэнсиса Дж. Э., Пирри Д., Крейма Дж. А. (ред.). Мел-третичные высокоширотные палеообстановки: бассейн Джеймса Росса, Антарктида . Геологическое общество, Лондон, Специальные публикации. Том. 258. Лондонское геологическое общество. стр. 163–176. Бибкод : 2006GSLSP.258..163B . дои : 10.1144/ГСЛ.СП.2006.258.01.12 . S2CID 140546667 .
- ^ Бонд М., Крамарц А., Макфи Р.Д., Регеро М. (2011). «Новый астрапотер (Mammalia, Meridiungulata) из формации Ла-Месета, остров Сеймур (Марамбио), и переоценка предыдущих записей антарктических астрапотеров» . Американский музей Novitates (3718): 1–16. дои : 10.1206/3718.2 . hdl : 11336/98139 . S2CID 58908785 . Архивировано из оригинала (PDF) 17 сентября 2012 года.
- ^ Гельфо Х.Н., Мёрс Т., Лоренте М., Лопес Г.М., Регеро М. (16 июля 2014 г.). «Самые старые млекопитающие Антарктиды, ранний эоцен формации Ла-Месета, остров Сеймур» . Палеонтология . 58 (1): 101–110. дои : 10.1111/пала.12121 . Архивировано из оригинала 5 апреля 2023 года.
- ^ Дэвис С.Н., Торрес С.Р., Массер Г.М., Проффитт Дж.В., Крауч Н.М., Лунделиус Э.Л. и др. (2020). «Новые находки млекопитающих и птиц из позднеэоценовых формаций Ла-Месета и Субмесета на острове Сеймур, Антарктида» . ПерДж . 8 : е8268. дои : 10.7717/peerj.8268 . ПМК 6955110 . ПМИД 31942255 .
- ^ Jump up to: Перейти обратно: а б Докинз Р. (2005). Рассказ предка: Паломничество на заре эволюции . Бостон: Mariner Books. п. 223. ИСБН 978-0-618-61916-0 .
- ^ Jump up to: Перейти обратно: а б Хэнд С.Дж., Лонг Дж., Арчер М., Фланнери Т.Ф. (2002). Доисторические млекопитающие Австралии и Новой Гвинеи: сто миллионов лет эволюции . Балтимор: Издательство Университета Джонса Хопкинса. ISBN 978-0-8018-7223-5 .
- ^ Кемп, Т.С. (2005). Происхождение и эволюция млекопитающих . Оксфорд [Оксфордшир]: Издательство Оксфордского университета. ISBN 978-0-19-850761-1 .
- ^ Грейвс Дж. А., Ренфри М.Б. (2013) Сумчатые в эпоху геномики . Анну Рев Геном Хум Генет
- ^ Лу, Донна (30 мая 2023 г.). «Познакомьтесь с первым в Австралии ходоком на длинные дистанции: сумчатым весом 250 кг с «пятками на руках» » . Хранитель . Проверено 31 мая 2023 г.
Дальнейшее чтение [ править ]
- Остин CR, Short RV, ред. (21 марта 1985 г.). Размножение млекопитающих: Том 4, Репродуктивная пригодность . Издательство Кембриджского университета. стр. 4–. ISBN 978-0-521-31984-3 .
- Бронсон Ф.Х. (1989). Репродуктивная биология млекопитающих . Издательство Чикагского университета. ISBN 978-0-226-07559-4 .
- Доусон Т.Дж. (1995). Кенгуру: Биология крупнейших сумчатых . Издательство Корнельского университета. ISBN 978-0-8014-8262-5 .
- Фланнери Т.Ф. (2002). Будущие пожиратели: экологическая история австралийских земель и людей . Гроув Пресс. стр. 67–75. ISBN 978-0-8021-3943-6 .
- Фланнери Т.Ф. (2008). В погоне за кенгуру: континент, ученый и поиск самого необычного существа в мире (1-е американское изд.). Нью-Йорк: Гроув. ISBN 9780802143716 .
- Фланнери Т.Ф. (2005). Страна: континент, учёный и кенгуру (2-е изд.). Мельбурн: Текстовый паб. ISBN 978-1-920885-76-2 .
- Фрит, Х.Дж. и Дж.Х. Калаби. Кенгуру. Нью-Йорк: Humanities Press, 1969.
- Маккей Дж. (2006). Энциклопедия МЛЕКОПИТАЮЩИХ . Уэлдон Оуэн. ISBN 978-1-74089-352-7 .
- Хансакер Д. (1977). Биология сумчатых . Нью-Йорк: Академическая пресса.
- Джонсон М.Х., Эверит Б.Дж. (1988). Необходимая репродукция . Блэквелл Сайентифик. ISBN 978-0-632-02183-3 .
- Джонс М., Дикман С., Арчер (2003). Хищники с сумками: биология хищных сумчатых . Коллингвуд, Виктория: Австралия). ISBN 9780643066342 .
- Нобилл Э., Нил Дж.Д., ред. (1998). Энциклопедия репродукции . Том. 3. Нью-Йорк: Академик Пресс.
- Маккалоу Д.Р., Маккалоу Ю. (2000). Кенгуру в глубинке Австралии: сравнительная экология и поведение трех сосуществующих видов . Издательство Колумбийского университета. ISBN 978-0-231-11916-0 .
- Новак Р.М. (7 апреля 1999 г.). Млекопитающие мира Уокера . Джу Пресс. ISBN 978-0-8018-5789-8 .
- Тейлор AC, Тейлор П (1997). «Пол детенышей сумки связан с весом матери у Macropus eugeni и M. parma ». Австралийский журнал зоологии . 45 (6): 573–578. дои : 10.1071/ZO97038 .
Внешние ссылки [ править ]
- «Виды млекопитающих Западной Австралии» . Members.iinet.net.au . Проверено 28 июня 2021 г.
- «Исследователи опубликовали первую последовательность генома сумчатых» . Genome.gov . Проверено 28 июня 2021 г.
- Опубликован первый геном сумчатого. Большинство различий между опоссомами и плацентарными млекопитающими связаны с некодирующей ДНК. Архивировано 4 февраля 2011 г. в Wayback Machine.



