РНК-полимераза
| ДНК-направленная РНК-полимераза | |||
|---|---|---|---|
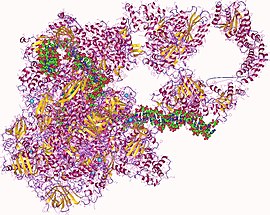 Гетеро27мер РНК-полимеразы, человека | |||
| Идентификаторы | |||
| Номер ЕС. | 2.7.7.6 | ||
| Номер CAS. | 9014-24-8 | ||
| Базы данных | |||
| ИнтЭнк | вид IntEnz | ||
| БРЕНДА | БРЕНДА запись | ||
| Экспаси | Просмотр NiceZyme | ||
| КЕГГ | КЕГГ запись | ||
| МетаЦик | метаболический путь | ||
| ПРЯМОЙ | профиль | ||
| PDB Структуры | RCSB PDB PDBe PDBsum | ||
| Генная онтология | АмиГО / QuickGO | ||
| |||

В молекулярной биологии РНК -полимераза (сокращенно РНКП или РНКпол ), или, более конкретно, ДНК-направленная/зависимая РНК-полимераза ( DdRP ), представляет собой фермент , который катализирует химические реакции, в ходе которых синтезируется РНК из матрицы ДНК .
Используя фермент хеликазу , РНКП локально открывает двухцепочечную ДНК, так что одну цепь обнаженных нуклеотидов можно использовать в качестве матрицы для синтеза РНК — процесса, называемого транскрипцией . Фактор транскрипции и связанный с ним комплекс медиатора транскрипции должны быть прикреплены к участку связывания ДНК, называемому промоторной областью, прежде чем РНКП сможет инициировать раскручивание ДНК в этом положении. РНКП не только инициирует транскрипцию РНК, но также направляет нуклеотиды в нужное положение, облегчает прикрепление и удлинение , обладает собственными возможностями корректуры и замены, а также способностью распознавания терминации. У эукариот РНКП может строить цепи длиной до 2,4 миллиона нуклеотидов.
РНКП продуцирует РНК, которая функционально предназначена либо для кодирования белка , то есть информационной РНК (мРНК); или некодирующие (так называемые «гены РНК»). Примерами четырех функциональных типов генов РНК являются:
- Транспортная РНК (тРНК)
- Переносит определенные аминокислоты в растущие полипептидные цепи в рибосомальном сайте синтеза белка во время трансляции ;
- Рибосомальная РНК (рРНК)
- Встраивается в рибосомы;
- Микро РНК (миРНК)
- Регулирует активность генов; и подавление РНК
- Каталитическая РНК ( рибозим )
- Функционирует как ферментативно активная молекула РНК.
РНК-полимераза необходима для жизни и обнаружена во всех живых организмах и во многих вирусах . В зависимости от организма РНК-полимераза может представлять собой белковый комплекс (многосубъединичная РНКП) или состоять только из одной субъединицы (односубъединичная РНКП, оцРНКП), каждая из которых представляет независимую линию. Первый обнаружен как у бактерий , архей и эукариот , имеющих схожую основную структуру и механизм. [1] Последний обнаружен в фагах , а также в эукариотических хлоропластах и митохондриях и родственен современным ДНК-полимеразам . [2] Эукариотические и архейные РНКП имеют больше субъединиц, чем бактериальные, и контролируются по-разному.
Бактерии и археи имеют только одну РНК-полимеразу. Эукариоты имеют несколько типов ядерной РНКП, каждый из которых отвечает за синтез определенного подмножества РНК:
- РНК-полимераза I синтезирует пре-рРНК 45S ( 35S у дрожжей ), которая созревает и образует основные участки РНК рибосомы.
- РНК-полимераза II синтезирует предшественники мРНК и большинство мРНК и микроРНК.
- РНК-полимераза III синтезирует тРНК, рРНК 5S и другие малые РНК, находящиеся в ядре и цитозоле .
- РНК-полимеразы IV и V , обнаруженные в растениях, менее изучены; они производят миРНК . Помимо оцРНКП, хлоропласты также кодируют и используют бактериоподобную РНКП.
Структура
[ редактировать ]

2006 года Нобелевская премия по химии была присуждена Роджеру Д. Корнбергу за создание детальных молекулярных изображений РНК-полимеразы на различных стадиях процесса транскрипции. [3] [4]
У большинства прокариот один вид РНК-полимеразы транскрибирует все типы РНК. «Ядро» РНК-полимеразы E. coli состоит из пяти субъединиц: двух альфа-(α)-субъединиц массой 36 кДа , бета-(β)-субъединицы массой 150 кДа, бета-субъединицы (β') массой 155 кДа и небольшой омега-субъединицы. (ω) субъединица. Сигма-фактор (σ) связывается с ядром, образуя голофермент. После начала транскрипции фактор может отсоединиться и позволить основному ферменту продолжить свою работу. [5] [6] Комплекс коровой РНК-полимеразы образует структуру «крабовой клешни» или «зажимной челюсти» с внутренним каналом, проходящим по всей длине. [7] Эукариотические и архейные РНК-полимеразы имеют схожую основную структуру и работают сходным образом, хотя у них есть много дополнительных субъединиц. [8]
Все РНКП содержат кофакторы металлов , в частности катионы цинка и магния , которые способствуют процессу транскрипции. [9] [10]
Функция
[ редактировать ]Контроль процесса транскрипции генов влияет на характер экспрессии генов и тем самым позволяет клетке адаптироваться к изменяющейся среде, выполнять специализированные роли внутри организма и поддерживать основные метаболические процессы, необходимые для выживания. Поэтому неудивительно, что активность РНКП продолжительна, сложна и строго регулируется. У бактерий Escherichia coli более 100 факторов транскрипции , модифицирующих активность РНКП. идентифицировано [11]
РНКП может инициировать транскрипцию на определенных последовательностях ДНК, известных как промоторы . Затем он производит цепь РНК, которая комплементарна цепи матрицы ДНК. Процесс добавления нуклеотидов к цепи РНК известен как элонгация; у эукариот РНКП может строить цепи длиной до 2,4 миллиона нуклеотидов (полная длина гена дистрофина ). РНКП будет преимущественно высвобождать свой транскрипт РНК в определенных последовательностях ДНК, кодируемых на концах генов, которые известны как терминаторы .
Продукты RNAP включают:
- Информационная РНК (мРНК) — матрица для белков рибосомами синтеза .
- Некодирующая РНК или «гены РНК» — широкий класс генов, кодирующих РНК, не транслируемую в белок. Наиболее яркими примерами генов РНК являются транспортная РНК (тРНК) и рибосомальная РНК (рРНК), обе из которых участвуют в процессе трансляции . Однако с конца 1990-х годов было обнаружено множество новых генов РНК, и, таким образом, гены РНК могут играть гораздо более важную роль, чем считалось ранее.
- Транспортная РНК (тРНК) — переносит определенные аминокислоты к растущим полипептидным цепям в рибосомальном сайте синтеза белка во время трансляции.
- Рибосомальная РНК (рРНК) — компонент рибосом.
- Микро РНК — регулирует активность генов.
- Каталитическая РНК ( рибозим ) — ферментативно активные молекулы РНК.
РНКП осуществляет de novo синтез . Он способен на это, поскольку специфические взаимодействия с инициирующим нуклеотидом жестко удерживают РНКП на месте, облегчая химическую атаку на входящий нуклеотид. Такие специфические взаимодействия объясняют, почему RNAP предпочитает начинать транскрипты с АТФ (за которым следуют GTP, UTP и затем CTP). В отличие от ДНК-полимеразы , РНКП обладает геликазной активностью, поэтому для раскручивания ДНК не требуется отдельный фермент.
Действие
[ редактировать ]Инициация
[ редактировать ]Связывание РНК-полимеразы у бактерий включает сигма-фактор , распознающий основную область промотора, содержащую элементы -35 и -10 (расположенные перед началом транскрибируемой последовательности), а также, на некоторых промоторах, С-концевой домен α-субъединицы, распознающий промотор выше по ходу транскрипции. элементы. [12] Существует множество взаимозаменяемых сигма-факторов, каждый из которых распознает отдельный набор промоторов. Например, у E. coli σ 70 экспрессируется в нормальных условиях и распознает промоторы генов, необходимых в нормальных условиях (« гены домашнего хозяйства »), тогда как σ 32 распознает промоторы генов, необходимых при высоких температурах (« гены теплового шока »). У архей и эукариот функции бактериального общего фактора транскрипции сигма выполняются несколькими общими факторами транскрипции , которые работают вместе. Закрытый комплекс РНК-полимераза-промотор обычно называют « комплексом преинициации транскрипции ». [13] [14]
После связывания с ДНК РНК-полимераза переключается с закрытого комплекса на открытый. Это изменение включает разделение нитей ДНК с образованием раскрученного участка ДНК длиной примерно 13 пар оснований, называемого « пузырем транскрипции ». Суперспирализация играет важную роль в активности полимеразы из-за раскручивания и перемотки ДНК. Поскольку участки ДНК перед РНКП раскручены, возникают компенсаторные положительные суперспирали. Области позади РНКП перемотаны и присутствуют отрицательные суперспирали. [14]
Побег промоутера
[ редактировать ]Затем РНК-полимераза начинает синтезировать исходный гетеродуплекс ДНК-РНК, в котором основания рибонуклеотидов спарены с цепью матричной ДНК в соответствии с взаимодействиями спаривания оснований Уотсона-Крика. Как отмечалось выше, РНК-полимераза вступает в контакт с промоторной областью. Однако эти стабилизирующие контакты ингибируют способность фермента получать доступ к ДНК в дальнейшем и, следовательно, синтез полноразмерного продукта. Чтобы продолжить синтез РНК, РНК-полимераза должна покинуть промотор. Он должен поддерживать контакты с промотором, одновременно раскручивая больше нижестоящей ДНК для синтеза, «сжимая» больше нижестоящей ДНК в инициирующий комплекс. [15] Во время перехода от промотора РНК-полимераза считается «стрессовым промежуточным продуктом». С термодинамической точки зрения стресс накапливается в результате действий по раскручиванию и уплотнению ДНК. Как только гетеродуплекс ДНК-РНК становится достаточно длинным (~ 10 п.н.), РНК-полимераза освобождает свои верхние контакты и эффективно достигает перехода от промотора в фазу элонгации. Гетеродуплекс в активном центре стабилизирует комплекс элонгации.
Однако бегство промоутера — не единственный результат. РНК-полимераза также может снять стресс, освобождая свои нижележащие контакты и останавливая транскрипцию. У приостановленного транскрипционного комплекса есть два варианта: (1) высвободить возникающий транскрипт и начать заново с промотора или (2) восстановить новый 3'-ОН на возникающем транскрипте в активном сайте посредством каталитической активности РНК-полимеразы и возобновить сжатие ДНК для добиться побега промотора. Абортивная инициация , непродуктивный цикл РНК-полимеразы перед переходом, ускользающим от промотора, приводит к образованию коротких фрагментов РНК длиной около 9 п.н. в процессе, известном как абортивная транскрипция. Степень абортивной инициации зависит от присутствия факторов транскрипции и силы контактов промотора. [16]
Удлинение
[ редактировать ]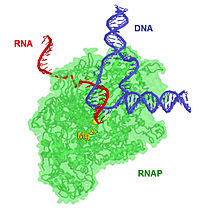
Транскрипционный комплекс длиной 17 п.н. представляет собой гибрид ДНК-РНК длиной 8 п.о., то есть 8 пар оснований включают транскрипт РНК, связанный с цепью матрицы ДНК. [17] По мере прогрессирования транскрипции к 3'-концу транскрипта РНК добавляются рибонуклеотиды, и комплекс РНКП перемещается по ДНК. Характерные скорости элонгации у прокариот и эукариот составляют около 10–100 нт/сек. [18]
Остатки аспартила ( asp ) в РНКП удерживают Mg. 2+ ионы, которые, в свою очередь, координируют фосфаты рибонуклеотидов. Первый мг 2+ будет удерживать α-фосфат добавляемого NTP. Это позволяет нуклеофильно атаковать 3'-ОН транскрипта РНК, добавляя к цепи еще один NTP. Второй мг 2+ будет держаться пирофосфат НТП. [19] Общее уравнение реакции:
- (НМП) n + НТП → (НМП) n+1 + ПП i
Верность
[ редактировать ]В отличие от механизмов корректуры ДНК-полимеразы, механизмы РНКП были исследованы лишь недавно. Корректура начинается с отделения неправильно включенного нуклеотида от матрицы ДНК. Это приостанавливает транскрипцию. Затем полимераза возвращается на одну позицию и расщепляет динуклеотид, содержащий несовпадающий нуклеотид. В РНК-полимеразе это происходит в том же активном сайте, который используется для полимеризации, и поэтому заметно отличается от ДНК-полимеразы, где корректура происходит в отдельном активном сайте нуклеазы. [20]
Общий коэффициент ошибок составляет около 10. −4 до 10 −6 . [21]
Прекращение действия
[ редактировать ]У бактерий терминация транскрипции РНК может быть ро-зависимой или ро-независимой. Первый основан на факторе rho , который дестабилизирует гетеродуплекс ДНК-РНК и вызывает высвобождение РНК. [22] Последнее, также известное как внутренняя терминация , основано на палиндромном участке ДНК. Транскрипция этой области вызывает образование «шпильки» из зацикливания транскрипционной РНК и ее связывания сама с собой. Эта шпильчная структура часто богата парами оснований GC, что делает ее более стабильной, чем сам гибрид ДНК-РНК. В результате гибрид ДНК-РНК длиной 8 п.н. в транскрипционном комплексе превращается в гибрид длиной 4 п.н. Эти последние 4 пары оснований являются слабыми парами оснований AU, и весь транскрипт РНК отпадет от ДНК. [23]
Терминация транскрипции у эукариот менее изучена, чем у бактерий, но включает расщепление нового транскрипта с последующим независимым от матрицы добавлением аденинов к его новому 3'-концу в процессе, называемом полиаденилированием . [24]
Другие организмы
[ редактировать ]Учитывая, что ДНК- и РНК-полимеразы осуществляют матрично-зависимую полимеризацию нуклеотидов, можно было бы ожидать, что эти два типа ферментов будут структурно родственными. Однако рентгеновские кристаллографические исследования обоих типов ферментов показывают, что, помимо содержания критического Mg 2+ ион в каталитическом центре практически не связан друг с другом; действительно, зависимые от матрицы ферменты, полимеризующие нуклеотиды, по-видимому, возникали независимо дважды в ходе ранней эволюции клеток. Одна линия привела к современным ДНК-полимеразам и обратным транскриптазам, а также к нескольким односубъединичным РНК-полимеразам (оцРНКП) из фагов и органелл. [2] Другая мультисубъединичная линия РНКП сформировала все современные клеточные РНК-полимеразы. [25] [1]
Бактерии
[ редактировать ]У бактерий один и тот же фермент катализирует синтез мРНК и некодирующей РНК (нкРНК) .
РНКП представляет собой большую молекулу. Основной фермент состоит из пяти субъединиц (~ 400 кДа ): [26]
- б'
- Субъединица β' является самой крупной субъединицей и кодируется геном rpoC. [27] Субъединица β' содержит часть активного центра, ответственного за синтез РНК, и содержит некоторые детерминанты неспецифичных для последовательности взаимодействий с ДНК и возникающей РНК. У цианобактерий он разделен на две субъединицы и хлоропласты. [28]
- б
- Субъединица β является второй по величине субъединицей и кодируется геном rpoB . Субъединица β содержит остальную часть активного центра, ответственного за синтез РНК, и остальные детерминанты неспецифичных для последовательности взаимодействий с ДНК и образующейся РНК.
- а (а я и α II )
- В молекуле РНКП присутствуют две копии α-субъединицы, третьей по величине субъединицы: α я и α II (один и два). Каждая субъединица α содержит два домена: αNTD (N-концевой домен) и αCTD (С-концевой домен). αNTD содержит детерминанты сборки РНКП. αCTD (С-концевой домен) содержит детерминанты для взаимодействия с промоторной ДНК, обеспечивающие неспецифичные для последовательности взаимодействия на большинстве промоторов и специфичные для последовательности взаимодействия на промоторах, содержащих вышерасположенные элементы, а также содержит детерминанты для взаимодействия с регуляторными факторами.
- ой
- Субъединица ω является наименьшей субъединицей. Субъединица ω облегчает сборку РНКП и стабилизирует собранную РНКП. [29]
Чтобы связать промоторы, ядро РНКП связывается с фактором инициации транскрипции сигма (σ), образуя голофермент РНК-полимеразы. Sigma снижает сродство РНКП к неспецифической ДНК, одновременно повышая специфичность к промоторам, позволяя инициировать транскрипцию в правильных сайтах. Таким образом, полный голофермент имеет 6 субъединиц: β'βα. я и α II ос (~450 кДа).
Эукариоты
[ редактировать ]
Эукариоты имеют несколько типов ядерной РНКП, каждый из которых отвечает за синтез определенного подмножества РНК. Все они структурно и механически связаны друг с другом и с бактериальной РНКП:
- РНК-полимераза I синтезирует пре- рРНК 45S (35S у дрожжей), которая созревает в 28S, 18S и 5,8S рРНК, которые образуют основные участки РНК рибосомы . [30]
- РНК-полимераза II синтезирует предшественники мРНК и большинство мяРНК и микроРНК . [31] ряд транскрипционных факторов . Это наиболее изученный тип, и из-за высокого уровня контроля, необходимого для транскрипции, для его связывания с промоторами необходим
- РНК-полимераза III синтезирует тРНК , рРНК 5S и другие малые РНК, находящиеся в ядре и цитозоле . [32]
- РНК-полимераза IV синтезирует миРНК в растениях. [33]
- РНК-полимераза V синтезирует РНК, участвующие в siRNA -направленном образовании гетерохроматина у растений. [34]
Эукариотические хлоропласты содержат РНКП, очень похожую на бактериальную РНКП («кодируемая пластидами полимераза, PEP»). Они используют сигма-факторы, закодированные в ядерном геноме. [35]
Хлоропласты также содержат вторую, структурно и механически неродственную, односубъединичную РНКП («кодируемая ядром полимераза, NEP»). Эукариотические митохондрии используют POLRMT (человеческий), односубъединичную РНКП, кодируемую ядром. [2] Такие фагоподобные полимеразы у растений называются RpoT. [35]
Архея
[ редактировать ]Археи имеют один тип РНКП, отвечающий за синтез всех РНК. Архейный РНКП структурно и механически подобен бактериальному РНКП и эукариотическому ядерному РНКП IV и особенно тесно структурно и механически связан с эукариотическим ядерным РНКП II. [8] [36] История открытия архейной РНК-полимеразы сравнительно недавняя. Первый анализ РНКП археи был выполнен в 1971 г., когда РНКП крайнего галофила Halobacterium Cutirubrum . была выделена и очищена [37] Кристаллические структуры РНКП Sulfolobus solfataricus и Sulfolobus shibatae определяют общее количество идентифицированных архейных субъединиц в тринадцать. [8] [38]
У архей субъединица, соответствующая эукариотическому Rpb1, разделена на две части. нет гомолога эукариотического Rpb9 ( POLR2I В комплексе S. shibatae ) , хотя TFS (гомолог TFIIS) был предложен как гомолог на основании сходства. Существует дополнительная субъединица, получившая название Rpo13; вместе с Rpo5 он занимает пространство, заполненное вставкой, обнаруженной в бактериальных β'-субъединицах (1377–1420 в Taq ). [8] Более раннее исследование структуры S. solfataricus с более низким разрешением не обнаружило Rpo13, а место было отнесено только к Rpo5/Rpb5. Rpo3 примечателен тем, что это железо-серный белок . Субъединица AC40 РНКП I/III, обнаруженная у некоторых эукариот, имеет схожие последовательности. [38] но не связывает железо. [39] В любом случае эта область выполняет структурную функцию. [40]
Субъединица РНКП архей ранее использовала номенклатуру «RpoX», в которой каждой субъединице присвоена буква способом, не связанным с какими-либо другими системами. [1] В 2009 году была предложена новая номенклатура, основанная на нумерации субъединицы эукариотического Pol II «Rpb». [8]
Вирусы
[ редактировать ]
Ортопоксвирусы и некоторые другие нуклеоцитоплазматические крупные ДНК-вирусы синтезируют РНК с использованием многосубъединичной РНКП, кодируемой вирусом. Они наиболее похожи на эукариотические РНКП, но некоторые субъединицы минимизированы или удалены. [41] Вопрос о том, на какой именно RNAP они больше всего похожи, является предметом споров. [42] Большинство других вирусов, синтезирующих РНК, используют несвязанную механику.
Многие вирусы используют односубъединичную ДНК-зависимую РНКП (оцРНКП), которая структурно и механически родственна односубъединичной РНКП эукариотических хлоропластов (RpoT) и митохондрий ( POLRMT ) и, в более отдаленной степени, ДНК-полимеразам и обратным транскриптазам . Вероятно, наиболее широко изученной такой односубъединичной РНКП является РНК-полимераза бактериофага Т7 . ssRNAP не могут корректироваться. [2]
SPβ B. subtilis Профаг использует YonO, гомолог субъединиц β+β' мсРНКП, для формирования мономерной (оба ствола на одной цепи) РНКП, отличной от обычной «правой» оцРНКП. Вероятно, он очень давно отошел от канонической пятизвенной мсРНКП, еще до появления последнего универсального общего предка . [43] [44]
Другие вирусы используют РНК-зависимую РНКП (РНКП, в которой в качестве матрицы используется РНК вместо ДНК). Это происходит у РНК-вирусов с отрицательной цепью и дцРНК-вирусов , которые часть своего жизненного цикла существуют в виде двухцепочечной РНК. Однако некоторые вирусы с положительной цепью РНК , такие как полиовирус , также содержат РНК-зависимую РНКП. [45]
История
[ редактировать ]RNAP был открыт независимо Чарльзом Ло, Одри Стивенс и Джерардом Гурвицем в 1960 году. [46] К этому времени половина Нобелевской премии по медицине 1959 года была присуждена Северо Очоа за открытие того, что считалось РНКП. [47] но вместо этого оказалась полинуклеотидфосфорилаза .
Очистка
[ редактировать ]РНК-полимеразу можно выделить следующими способами:
- На колонке с фосфоцеллюлозой . [48]
- Центрифугированием в градиенте глицерина . [49]
- По колонке ДНК .
- На колонке ионной хроматографии . [50]
А также комбинации вышеперечисленных техник.
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д Вернер Ф., Громанн Д. (февраль 2011 г.). «Эволюция многосубъединичных РНК-полимераз в трех доменах жизни». Обзоры природы. Микробиология . 9 (2): 85–98. дои : 10.1038/nrmicro2507 . ПМИД 21233849 . S2CID 30004345 . См. также Крамер 2002: Крамер П. (февраль 2002 г.). «Мультисубъединичные РНК-полимеразы». Современное мнение в области структурной биологии . 12 (1): 89–97. дои : 10.1016/s0959-440x(02)00294-4 . ПМИД 11839495 .
- ^ Jump up to: а б с д Чермакян Н., Икеда Т.М., Мирамонтес П., Ланг Б.Ф., Грей М.В., Седергрен Р. (декабрь 1997 г.). «Об эволюции односубъединичных РНК-полимераз». Журнал молекулярной эволюции . 45 (6): 671–681. Бибкод : 1997JMolE..45..671C . CiteSeerX 10.1.1.520.3555 . дои : 10.1007/PL00006271 . ПМИД 9419244 . S2CID 1624391 .
- ^ Нобелевская премия по химии 2006 г.
- ^ Стоддарт, Шарлотта (1 марта 2022 г.). «Структурная биология: как белки оказались крупным планом» . Знающий журнал . doi : 10.1146/knowable-022822-1 . Проверено 25 марта 2022 г.
- ^ Гриффитс AJF, Миллер Дж.Х., Сузуки Д.Т. и др. Введение в генетический анализ. 7-е издание. Нью-Йорк: WH Freeman; 2000. Глава 10.
- ^ Финн Р.Д., Орлова Е.В., Гоуэн Б., Бак М., ван Хил М. (декабрь 2000 г.). «Ядро РНК-полимеразы Escherichia coli и структуры голоферментов» . Журнал ЭМБО . 19 (24): 6833–6844. дои : 10.1093/emboj/19.24.6833 . ПМК 305883 . ПМИД 11118218 .
- ^ Чжан Г., Кэмпбелл Е.А., Минахин Л., Рихтер С., Северинов К., Дарст С.А. (сентябрь 1999 г.). «Кристаллическая структура коровой РНК-полимеразы Thermus aquaticus при разрешении 3,3 А» . Клетка . 98 (6): 811–824. дои : 10.1016/S0092-8674(00)81515-9 . ПМИД 10499798 .
- ^ Jump up to: а б с д и Корхин Ю., Унлигил У.М., Литтлфилд О., Нельсон П.Дж., Стюарт Д.И., Сиглер П.Б. и др. (май 2009 г.). «Эволюция сложных РНК-полимераз: полная структура РНК-полимеразы архей» . ПЛОС Биология . 7 (5): e1000102. дои : 10.1371/journal.pbio.1000102 . ПМЦ 2675907 . ПМИД 19419240 .
- ^ Альбертс Б (18 ноября 2014 г.). Молекулярная биология клетки (Шестое изд.). Нью-Йорк, штат Нью-Йорк. ISBN 9780815344322 . OCLC 887605755 .
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ) - ^ Марков Д., Нарышкина Т., Мустаев А., Северинов К. (сентябрь 1999 г.). «Участок связывания цинка в крупнейшей субъединице ДНК-зависимой РНК-полимеразы участвует в сборке фермента» . Гены и развитие . 13 (18): 2439–2448. дои : 10.1101/gad.13.18.2439 . ПМК 317019 . ПМИД 10500100 .
- ^ Исихама А (2000). «Функциональная модуляция РНК-полимеразы Escherichia coli». Ежегодный обзор микробиологии . 54 : 499–518. дои : 10.1146/annurev.micro.54.1.499 . ПМИД 11018136 .
- ^ ИнтерПро : IPR011260
- ^ Редер Р.Г. (ноябрь 1991 г.). «Сложности инициации транскрипции эукариот: регуляция сборки преинициативного комплекса». Тенденции биохимических наук . 16 (11): 402–408. дои : 10.1016/0968-0004(91)90164-Q . ПМИД 1776168 .
- ^ Jump up to: а б Уотсон Дж.Д., Бейкер Т.А., Белл С.П., Ганн А.А., Левин М., Лосик Р.М. (2013). Молекулярная биология гена (7-е изд.). Пирсон.
- ^ Ревякин А., Лю С., Эбрайт Р.Х., Стрик Т.Р. (ноябрь 2006 г.). «Абортивная и продуктивная инициация с помощью РНК-полимеразы включает в себя сжатие ДНК» . Наука . 314 (5802): 1139–1143. Бибкод : 2006Sci...314.1139R . дои : 10.1126/science.1131398 . ПМЦ 2754787 . ПМИД 17110577 .
- ^ Goldman SR, Ebright RH , Nickels BE (май 2009 г.). «Прямое обнаружение абортивных транскриптов РНК in vivo» . Наука . 324 (5929): 927–928. Бибкод : 2009Sci...324..927G . дои : 10.1126/science.1169237 . ПМЦ 2718712 . ПМИД 19443781 .
- ^ Кеттенбергер Х., Армаш К.Дж., Крамер П. (декабрь 2004 г.). «Полная структура комплекса элонгации РНК-полимеразы II и ее взаимодействие с NTP и TFIIS» . Молекулярная клетка . 16 (6): 955–965. doi : 10.1016/j.molcel.2004.11.040 . hdl : 11858/00-001M-0000-0015-84E1-D . ПМИД 15610738 .
- ^ Майло Р., Филипс Р. «Клеточная биология в цифрах: что быстрее: транскрипция или трансляция?» . book.bionumbers.org . Архивировано из оригинала 20 апреля 2017 года . Проверено 8 марта 2017 г.
- ^ Светлов В., Нудлер Э. (январь 2013 г.). «Основной механизм транскрипции РНК-полимеразой II» . Biochimica et Biophysica Acta (BBA) - Механизмы регуляции генов . 1829 (1): 20–28. дои : 10.1016/j.bbagrm.2012.08.009 . ПМК 3545073 . ПМИД 22982365 .
- ^ Сюдов Дж. Ф., Крамер П. (декабрь 2009 г.). «Правильность РНК-полимеразы и корректура транскрипции». Современное мнение в области структурной биологии . 19 (6): 732–739. дои : 10.1016/j.sbi.2009.10.009 . hdl : 11858/00-001M-0000-0015-837E-8 . ПМИД 19914059 .
- ^ Филипс Р., Майло Р. «Какова частота ошибок при транскрипции и переводе?» . Проверено 26 марта 2019 г.
- ^ Ричардсон Дж. П. (сентябрь 2002 г.). «Rho-зависимое терминирование и АТФазы в терминации транскрипта». Biochimica et Biophysical Acta (BBA) – Структура и экспрессия генов . 1577 (2): 251–260. дои : 10.1016/S0167-4781(02)00456-6 . ПМИД 12213656 .
- ^ Порруа О, Будвиллен М, Либри Д (август 2016 г.). «Завершение транскрипции: вариации на общие темы». Тенденции в генетике . 32 (8): 508–522. дои : 10.1016/j.tig.2016.05.007 .
- ^ Ликке-Андерсен С., Йенсен Т.Х. (октябрь 2007 г.). «Перекрывающиеся пути диктуют прекращение транскрипции РНК-полимеразы II». Биохимия . 89 (10): 1177–1182. дои : 10.1016/j.biochi.2007.05.007 . ПМИД 17629387 .
- ^ Стиллер Дж.В., Даффилд Э.К., Холл Б.Д. (сентябрь 1998 г.). «Амитохондриальные амебы и эволюция ДНК-зависимой РНК-полимеразы II» . Труды Национальной академии наук Соединенных Штатов Америки . 95 (20): 11769–11774. Бибкод : 1998PNAS...9511769S . дои : 10.1073/pnas.95.20.11769 . ПМК 21715 . ПМИД 9751740 .
- ^ Эбрайт Р.Х. (декабрь 2000 г.). «РНК-полимераза: структурное сходство между бактериальной РНК-полимеразой и эукариотической РНК-полимеразой II». Журнал молекулярной биологии . 304 (5): 687–698. дои : 10.1006/jmbi.2000.4309 . ПМИД 11124018 .
- ^ Монастырская Г.С., Губанов В.В., Гурьев С.О., Саломатина И.С., Шуваева Т.М., Липкин В.М., Свердлов Э.Д. (июль 1982 г.). «Первичная структура РНК-полимеразы E. coli , нуклеотидная последовательность гена rpoC и аминокислотная последовательность бета'-субъединицы» . Исследования нуклеиновых кислот . 10 (13): 4035–4044. дои : 10.1093/нар/10.13.4035 . ПМК 320776 . ПМИД 6287430 .
- ^ Бергсланд К.Дж., Хазелькорн Р. (июнь 1991 г.). «Эволюционные взаимоотношения между эубактериями, цианобактериями и хлоропластами: данные гена rpoC1 штамма Anabaena sp. PCC 7120» . Журнал бактериологии . 173 (11): 3446–3455. дои : 10.1128/jb.173.11.3446-3455.1991 . ПМК 207958 . ПМИД 1904436 .
- ^ Мэтью Р., Чаттерджи Д. (октябрь 2006 г.). «Развивающаяся история омега-субъединицы бактериальной РНК-полимеразы». Тенденции в микробиологии . 14 (10): 450–455. дои : 10.1016/j.tim.2006.08.002 . ПМИД 16908155 .
- ^ Груммт I (1999). Регуляция транскрипции рибосомальных генов млекопитающих с помощью РНК-полимеразы I. Прогресс в исследованиях нуклеиновых кислот и молекулярной биологии. Том. 62. стр. 109–54. дои : 10.1016/S0079-6603(08)60506-1 . ISBN 9780125400626 . ПМИД 9932453 .
- ^ Ли Ю, Ким М, Хан Дж, Ём К. Х., Ли С., Пэк Ш., Ким В. Н. (октябрь 2004 г.). «Гены микроРНК транскрибируются РНК-полимеразой II» . Журнал ЭМБО . 23 (20): 4051–4060. дои : 10.1038/sj.emboj.7600385 . ПМК 524334 . ПМИД 15372072 .
- ^ Уиллис И.М. (февраль 1993 г.). «РНК-полимераза III. Гены, факторы и транскрипционная специфичность» . Евро. Дж. Биохим . 212 (1): 1–11. дои : 10.1111/j.1432-1033.1993.tb17626.x . ПМИД 8444147 .
- ^ Герр А.Дж., Дженсен М.Б., Далмей Т., Баулкомб, округ Колумбия (апрель 2005 г.). «РНК-полимераза IV управляет молчанием эндогенной ДНК» . Наука . 308 (5718): 118–120. Бибкод : 2005Sci...308..118H . дои : 10.1126/science.1106910 . ПМИД 15692015 . S2CID 206507767 .
- ^ Вежбицкий А.Т., Реам Т.С., Хааг Дж.Р., Пикаард К.С. (май 2009 г.). «Транскрипция РНК-полимеразы V направляет ARGONAUTE4 к хроматину» . Природная генетика . 41 (5): 630–634. дои : 10.1038/ng.365 . ПМЦ 2674513 . ПМИД 19377477 .
- ^ Jump up to: а б Швеер Дж., Тюркери Х., Колпак А., Линк Дж. (декабрь 2010 г.). «Роль и регуляция пластидных сигма-факторов и их функциональных взаимодействий во время транскрипции хлоропластов - недавние уроки Arabidopsis thaliana». Европейский журнал клеточной биологии . 89 (12): 940–946. дои : 10.1016/j.ejcb.2010.06.016 . ПМИД 20701995 .
- ^ Вернер Ф (сентябрь 2007 г.). «Структура и функции РНК-полимераз архей» . Молекулярная микробиология . 65 (6): 1395–1404. дои : 10.1111/j.1365-2958.2007.05876.x . ПМИД 17697097 .
- ^ Луи Б.Г., Фитт П.С. (февраль 1971 г.). «Ферментология нуклеиновых кислот чрезвычайно галофильных бактерий. Halobacterium Cutirubrum, зависимая от дезоксирибонуклеиновой кислоты, полимераза рибонуклеиновой кислоты» . Биохимический журнал . 121 (4): 621–627. дои : 10.1042/bj1210621 . ПМЦ 1176638 . ПМИД 4940048 .
- ^ Jump up to: а б Хирата А., Кляйн Б.Дж., Мураками К.С. (февраль 2008 г.). «Рентгенокристаллическая структура РНК-полимеразы архей» . Природа . 451 (7180): 851–854. Бибкод : 2008Natur.451..851H . дои : 10.1038/nature06530 . ПМК 2805805 . ПМИД 18235446 .
- ^ Фернандес-Торнеро С., Морено-Морсильо М., Рашид У.Дж., Тейлор Н.М., Руис Ф.М., Груин Т. и др. (октябрь 2013 г.). «Кристаллическая структура 14-субъединичной РНК-полимеразы I» . Природа . 502 (7473): 644–649. Бибкод : 2013Natur.502..644F . дои : 10.1038/nature12636 . ПМИД 24153184 . S2CID 205235881 .
- ^ Дженнингс М.Э., Лесснер Ф.Х., Карр Э.А., Лесснер DJ (февраль 2017 г.). «Кластеры [4Fe-4S] Rpo3 являются ключевыми детерминантами формирования гетеродимера РНК-полимеразы после Rpo3/Rpo11 у Methanosarcina acetivorans» . МикробиологияОткрыть . 6 (1): e00399. дои : 10.1002/mbo3.399 . ПМК 5300874 . ПМИД 27557794 .
- ^ Мирзаханян Ю., Гершон П.Д. (сентябрь 2017 г.). «Мультисубъединичные ДНК-зависимые РНК-полимеразы вируса коровьей оспы и других нуклеоцитоплазматических вирусов с большой ДНК: впечатления от эпохи структуры» . Обзоры микробиологии и молекулярной биологии . 81 (3). дои : 10.1128/MMBR.00010-17 . ПМЦ 5584312 . ПМИД 28701329 .
- ^ Гульельмини Дж., Ву А.С., Крупович М., Фортерр П., Гайя М. (сентябрь 2019 г.). «Диверсификация гигантских и крупных эукариотических вирусов дцДНК предшествовала возникновению современных эукариот» . Труды Национальной академии наук Соединенных Штатов Америки . 116 (39): 19585–19592. Бибкод : 2019PNAS..11619585G . дои : 10.1073/pnas.1912006116 . ПМК 6765235 . ПМИД 31506349 .
- ^ Форрест Д., Джеймс К., Юзенкова Ю., Зенкин Н. (июнь 2017 г.). «Однопептидная ДНК-зависимая РНК-полимераза, гомологичная многосубъединичной РНК-полимеразе» . Природные коммуникации . 8 : 15774. Бибкод : 2017NatCo...815774F . дои : 10.1038/ncomms15774 . ПМК 5467207 . ПМИД 28585540 .
- ^ Соге Л (сентябрь 2019 г.). «Расширенное суперсемейство двухствольных полимераз: структура, функции и эволюция» . Журнал молекулярной биологии . 431 (20): 4167–4183. дои : 10.1016/j.jmb.2019.05.017 . ПМИД 31103775 .
- ^ Алквист П. (май 2002 г.). «РНК-зависимые РНК-полимеразы, вирусы и подавление РНК». Наука . 296 (5571): 1270–1273. Бибкод : 2002Sci...296.1270A . дои : 10.1126/science.1069132 . ПМИД 12016304 . S2CID 42526536 .
- ^ Гурвиц Дж. (декабрь 2005 г.). «Открытие РНК-полимеразы» . Журнал биологической химии . 280 (52): 42477–42485. дои : 10.1074/jbc.X500006200 . ПМИД 16230341 .
- ^ Нобелевская премия 1959 г.
- ^ Келли Дж.Л., Lehman IR (август 1986 г.). «Дрожжевая митохондриальная РНК-полимераза. Очистка и свойства каталитической субъединицы» . Журнал биологической химии . 261 (22): 10340–10347. дои : 10.1016/S0021-9258(18)67529-5 . ПМИД 3525543 .
- ^ Хонда А., Мукаигава Дж., Ёкоияма А., Като А., Уэда С., Нагата К. и др. (апрель 1990 г.). «Очистка и молекулярная структура РНК-полимеразы вируса гриппа A/PR8». Журнал биохимии . 107 (4): 624–628. doi : 10.1093/oxfordjournals.jbchem.a123097 . ПМИД 2358436 .
- ^ Хагер Д.А., Джин DJ, Берджесс Р.Р. (август 1990 г.). «Использование ионообменной хроматографии высокого разрешения Mono Q для получения высокочистой и активной РНК-полимеразы Escherichia coli». Биохимия . 29 (34): 7890–7894. дои : 10.1021/bi00486a016 . ПМИД 2261443 .
Внешние ссылки
[ редактировать ]- DNAi - DNA Interactive, включая информацию и флэш-ролики о РНК-полимеразе.
- РНК + полимераза в Национальной медицинской библиотеке США по медицинским предметным рубрикам (MeSH)
- ЭК 2.7.7.6
- РНК-полимераза – синтез РНК из матрицы ДНК
| Базы данных органов управления : Национальные |
|---|