Абортивное инициирование
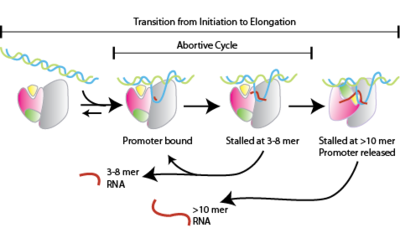
Абортивная инициация , также известная как абортивная транскрипция , представляет собой ранний процесс генетической транскрипции , при котором РНК-полимераза связывается с промотором ДНК и вступает в циклы синтеза коротких транскриптов мРНК , которые высвобождаются до того, как транскрипционный комплекс покинет промотор. Этот процесс происходит как у эукариот , так и у прокариот . Абортивную инициацию обычно изучают на РНК-полимеразах Т3 и Т7 в бактериофагах и E. coli .
Общий процесс
[ редактировать ]Абортивная инициация происходит до клиренса промотора . [1]
- РНК-полимераза связывается с ДНК промотора, образуя закрытый комплекс РНК-полимераза-промотор.
- Затем РНК-полимераза раскручивает один виток ДНК, окружающий сайт начала транскрипции, с образованием открытого комплекса РНК-полимераза-промотор.
- РНК-полимераза вступает в абортивные циклы синтеза и высвобождает короткие продукты РНК (содержит менее 10 нуклеотидов)
- РНК-полимераза покидает промотор и вступает на стадию элонгации транскрипции.
Механизм
[ редактировать ]Абортивная инициация является нормальным процессом транскрипции и происходит как in vitro , так и in vivo . [2] После каждого этапа добавления нуклеотидов в начальной транскрипции РНК-полимераза стохастически может двигаться по пути ухода с промотора (продуктивная инициация) или может высвободить продукт РНК и вернуться к открытому комплексу РНК-полимераза-промотор (абортивная инициация). На этой ранней стадии транскрипции РНК-полимераза вступает в фазу, во время которой диссоциация транскрипционного комплекса энергетически конкурирует с процессом элонгации. Прерывание цикла не вызвано сильным связыванием между инициирующим комплексом и промотором. [3]
ДНК сжимается
[ редактировать ]
В течение многих лет механизм, с помощью которого РНК-полимераза движется по цепи ДНК во время абортивной инициации, оставался неясным. Было замечено, что РНК-полимераза не выходит из промотора во время инициации транскрипции, поэтому было неизвестно, как фермент может читать цепь ДНК, чтобы транскрибировать ее, не перемещаясь вниз по ходу транскрипции . Исследования, проведенные за последнее десятилетие, показали, что абортивная инициация включает сжатие ДНК, при котором РНК-полимераза остается неподвижной, пока она раскручивается и втягивает ДНК в транскрипционный комплекс, чтобы пройти нуклеотиды через активный сайт полимеразы, тем самым транскрибируя ДНК без движения. Это приводит к тому, что раскрученная ДНК накапливается внутри фермента, отсюда и название ДНК «скручивание». При прерывающейся инициации РНК-полимераза перематывает и выбрасывает нижнюю часть раскрученной ДНК, высвобождая РНК и возвращаясь к открытому комплексу РНК-полимераза-промотор; Напротив, при продуктивной инициации РНК-полимераза перематывает и выбрасывает расположенную выше часть раскрученной ДНК, разрывая взаимодействия РНК-полимеразы с промотором, выходя из промотора и образуя комплекс элонгации транскрипции. [1] [4]
В статье 2006 года, которая продемонстрировала участие сжатия ДНК в начальной транскрипции, была предложена идея о том, что стресс, возникающий во время сжатия ДНК, обеспечивает движущую силу как для абортивной и продуктивной инициации. [4] Сопутствующая статья, опубликованная в том же году, подтвердила, что обнаруживаемое сжатие ДНК происходит в 80% циклов транскрипции, а фактически оценивается в 100%, учитывая ограничение способности обнаруживать быстрое сжатие (20% сжатий имеют продолжительность менее 1 секунда). [1]
Статья 2016 года показала, что сжатие ДНК также происходит перед синтезом РНК во время выбора сайта начала транскрипции. [5]
Функция
[ редактировать ]Не существует общепринятых функций полученных усеченных транскриптов РНК. Однако исследование 1981 года обнаружило доказательства того, что существует связь между количеством абортивных транскриптов и временем, необходимым для успешного образования длинных цепей РНК. Когда РНК-полимераза претерпевает абортивную транскрипцию в присутствии АТФ, УТФ и ГТФ, образуется комплекс, который имеет гораздо меньшую способность к абортивному рециклингу и гораздо более высокую скорость синтеза полноразмерного транскрипта РНК. [6] Исследование 2010 года обнаружило доказательства того, что эти укороченные транскрипты ингибируют терминацию синтеза РНК с помощью шпильки РНК , зависимого от внутреннего терминатора . [7]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с Ревякин А, Лю С, Эбрайт Р.Х. , Стрик Т.Р. (2006). «Абортивная и продуктивная инициация с помощью РНК-полимеразы включает в себя сжатие ДНК» . Наука . 314 (5802): 1139–43. Бибкод : 2006Sci...314.1139R . дои : 10.1126/science.1131398 . ПМЦ 2754787 . ПМИД 17110577 .
- ^ Голдман С., Эбрайт Р.Х. , Никельс Б. (2009). «Прямое обнаружение абортивных транскриптов РНК in vivo » . Наука . 324 (5929): 927–928. Бибкод : 2009Sci...324..927G . дои : 10.1126/science.1169237 . ПМЦ 2718712 . ПМИД 19443781 .
- ^ Мартин К.Т., Мюллер Д.К., Коулман Дж.Э. (1988). «Процессивность на ранних стадиях транскрипции РНК-полимеразой Т7». Биохимия . 27 (11): 3966–74. дои : 10.1021/bi00411a012 . ПМИД 3415967 .
- ^ Перейти обратно: а б Капанидис А.Н., Марджат Э., Хо СО, Кортхонджиа Э., Вайс С., Эбрайт Р.Х. (2006). «Первоначальная транскрипция РНК-полимеразой осуществляется посредством механизма сжатия ДНК» . Наука . 314 (5802): 1144–7. Бибкод : 2006Sci...314.1144K . дои : 10.1126/science.1131399 . ПМЦ 2754788 . ПМИД 17110578 .
- ^ Винкельман Дж.Т., Введенская И.О., Чжан Ю., Чжан Ю., Бёрд Дж.Г., Тейлор Д.М., Гурс Р.Л., Эбрайт Р.Х., Никелс Б.Е. (2016). «Мультиплексное сшивание белок-ДНК: затруднения при выборе места начала транскрипции» . Наука . 351 (6277): 1090–3. Бибкод : 2016Sci...351.1090W . doi : 10.1126/science.aad6881 . ПМЦ 4797950 . ПМИД 26941320 .
- ^ Мансон Л.М., Резников В.С. (1981). «Абортивное инициирование и синтез длинной рибонуклеиновой кислоты». Биохимия . 20 (8): 2081–5. дои : 10.1021/bi00511a003 . ПМИД 6165380 .
- ^ Ли С., Нгуен Х.М., Канг С. (2010). «Крошечные транскрипты абортивной инициации оказывают антитерминирующую активность на зависимом от шпильки внутреннем терминаторе РНК» . Нуклеиновые кислоты Рез . 38 (18): 6045–53. дои : 10.1093/nar/gkq450 . ПМЦ 2952870 . ПМИД 20507918 .