Бацилла субтилис
В этой статье есть несколько проблем. Пожалуйста, помогите улучшить его или обсудите эти проблемы на странице обсуждения . ( Узнайте, как и когда удалять эти шаблонные сообщения )
|
| Бацилла субтилис | |
|---|---|
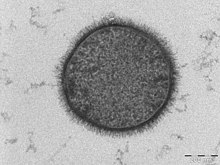
| |
| ПЭМ- микрофотография клетки B. subtilis в поперечном сечении (масштабная линейка = 200 нм ) | |
| Научная классификация | |
| Домен: | Бактерии |
| Тип: | Бациллота |
| Сорт: | Бациллы |
| Заказ: | Бациллы |
| Семья: | Бацилловые |
| Род: | Бацилла |
| Разновидность: | Б. субтилис
|
| Биномиальное имя | |
| Бацилла субтилис ( Эренберг 1835 г.)
Кон 1872 г. | |
| Синонимы | |
| |
Bacillus subtilis ( / b ə ˈ s ɪ l . ə s s u b ˈ t iː . l i s / ), [ 3 ] [ 4 ] как сенная палочка или травяная палочка , представляет собой грамположительную каталазоположительную желудочно известная также бактерию , обнаруженную в почве и -кишечном тракте жвачных животных , человека и морских губок. [ 5 ] [ 6 ] [ 7 ] [ 8 ] Будучи представителем рода Bacillus , B. subtilis имеет палочковидную форму и может образовывать прочную защитную эндоспору , что позволяет ей переносить экстремальные условия окружающей среды. B. subtilis исторически классифицировали как облигатный аэроб , хотя существуют доказательства того, что это факультативный анаэроб . B. subtilis считается наиболее изученной грамположительной бактерией и модельным организмом для изучения репликации бактериальных хромосом и дифференцировки клеток. Это один из бактериальных чемпионов по производству секретируемых ферментов , который используется в промышленных масштабах биотехнологическими компаниями. [ 5 ] [ 6 ] [ 7 ]
Описание
[ редактировать ]Bacillus subtilis — грамположительная бактерия, палочковидная и каталазоположительная . Первоначально он был назван Vibrio subtilis Кристианом Готфридом Эренбергом . [ 9 ] и переименован в Bacillus subtilis Фердинандом Коном в 1872 году. [ 10 ] (subtilis в переводе с латыни означает «тонкий, тонкий, стройный»). Клетки B. subtilis обычно имеют палочковидную форму, длину около 4–10 микрометров (мкм), диаметр 0,25–1,0 мкм и объем клетки в стационарной фазе около 4,6 фл. [ 6 ] [ 11 ]
Как и другие представители рода Bacillus , он может образовывать эндоспоры , чтобы выжить в экстремальных условиях окружающей среды, таких как температура и высыхание. [ 12 ] B. subtilis является факультативным анаэробом. [ 6 ] [ 13 ] считался облигатным аэробом. и до 1998 года B. subtilis имеет большое количество жгутиков , что дает ему способность быстро передвигаться в жидкостях.
B. subtilis оказался легко поддающимся генетическим манипуляциям и получил широкое распространение в качестве модельного организма для лабораторных исследований, особенно споруляции , которая является упрощенным примером клеточной дифференциации . С точки зрения популярности в качестве лабораторного модельного организма, B. subtilis часто считают грамположительным эквивалентом Escherichia coli , широко изученной грамотрицательной бактерии. [ 14 ]
Характеристики
[ редактировать ]Колонии, морфологические, физиологические и биохимические характеристики Bacillus subtilis представлены в таблице ниже. [ 6 ]
| Тип теста | Тест | Характеристики |
|---|---|---|
| Персонажи колонии | Размер | Середина |
| Тип | Круглый | |
| Цвет | Беловатый | |
| Форма | Выпуклый | |
| Морфологические символы | Форма | Стержень |
| Физиологические признаки | Подвижность | + |
| Рост при 6,5% NaCl | + | |
| Биохимические признаки | Окраска по Граму | + |
| Оксидаза | - | |
| Каталаза | + | |
| Окислительно-ферментативный | Ферментативный | |
| Подвижность | - | |
| Метиловый красный | - | |
| Voges-Proskauer | + | |
| Индол | - | |
| H 2 Производство S | + | |
| Уреаза | - | |
| Нитратредуктаза | + | |
| β-галактозидаза | + | |
| Гидролиз | Желатин | + |
| Эскулин | + | |
| Казеин | + | |
| Твин 40 | + | |
| Твин 60 | + | |
| Твин 80 | + | |
| Производство кислоты из | Глицерин | + |
| Галактоза | + | |
| D-глюкоза | + | |
| D-фруктоза | + | |
| D-манноза | + | |
| Маннитол | + | |
| N-ацетилглюкозамин | + | |
| Амигдалин | + | |
| Мальтоза | + | |
| D-Мелибиоза | + | |
| D-трегалоза | + | |
| Гликоген | + | |
| D-Тураноза | + |
Примечание: + = Положительный, – = Отрицательный.
среда обитания
[ редактировать ]Этот вид обычно встречается в верхних слоях почвы, и считается, что B. subtilis является нормальным комменсалом кишечника человека. В исследовании 2009 года сравнивалась плотность спор, обнаруженных в почве (около 10 6 спор на грамм) к тем, которые обнаруживаются в фекалиях человека (около 10 4 спор на грамм). Количество спор, обнаруженных в кишечнике человека, было слишком велико, чтобы его можно было объяснить исключительно употреблением в пищу через загрязнение пищевых продуктов. [ 15 ] В некоторых местах обитания пчел B. subtilis появляется во флоре кишечника медоносных пчел . [ 16 ] B. subtilis также можно встретить в морской среде. [ 6 ] [ 7 ]
Имеются данные о том, что B. subtilis имеет сапрофитную природу. Исследования показали, что бактерия демонстрирует вегетативный рост в почве, богатой органическими веществами, и что споры образуются при истощении питательных веществ. [ 17 ] Кроме того, B. subtilis было показано, что образует биопленки на корнях растений, что может объяснить, почему он часто встречается в микробиомах кишечника. [ 17 ] Возможно, животные, поедающие растения с биопленками B. subtilis, могут способствовать росту этой бактерии в желудочно-кишечном тракте. Было показано, что весь жизненный цикл B. subtilis может проходить в желудочно-кишечном тракте, что подтверждает идею о том, что бактерия попадает в кишечник через употребление растений и остается там благодаря своей способности расти в кишечнике. [ 17 ]
Воспроизведение
[ редактировать ]
Bacillus subtilis может делиться симметрично, образуя две дочерние клетки (бинарное деление), или асимметрично, образуя одну эндоспору , которая может оставаться жизнеспособной в течение десятилетий и устойчива к неблагоприятным условиям окружающей среды, таким как засуха , соленость , экстремальные значения pH , радиация и растворители . Эндоспора образуется во время пищевого стресса и за счет гидролиза, что позволяет организму сохраняться в окружающей среде до тех пор, пока условия не станут благоприятными. Перед процессом споруляции клетки могут стать подвижными , вырабатывая жгутики , поглощая ДНК из окружающей среды или производя антибиотики . [ 6 ] [ 7 ] Эти реакции рассматриваются как попытки найти питательные вещества, ища более благоприятную среду, позволяющую клетке использовать новый полезный генетический материал, или просто уничтожая конкуренцию. [ нужна ссылка ]
В стрессовых условиях, таких как недостаток питательных веществ, B. subtilis подвергается процессу споруляции . Этот процесс очень хорошо изучен и послужил модельным организмом для изучения споруляции. [ 18 ]
Спорообразование
[ редактировать ]Как только B. subtilis начинает спорообразование, сигма-фактор сигма F. секретируется [ 20 ] Этот фактор способствует спорообразованию. Формируется споруляционная перегородка, и хромосома медленно перемещается в предспору. Когда треть одной копии хромосомы находится в предспоре, а остальные две трети - в материнской клетке, фрагмент хромосомы в предспоре содержит локус сигмы F, который начинает экспрессироваться в предспоре. [ 21 ] Чтобы предотвратить экспрессию сигма F в материнской клетке, анти-сигма-фактор, кодируемый spoIIAB, [ 22 ] выражается. Любой остаточный анти-сигма-фактор в предспорах (который в противном случае мешал бы споруляции) ингибируется анти-антисигма-фактором, который кодируется spoIIAA. [ 22 ] SpoIIAA расположен рядом с локусом сигма-фактора, поэтому он постоянно экспрессируется в предспорах. Поскольку локус spoIIAB не расположен рядом с локусами сигма F и spoIIAA, он экспрессируется только в материнской клетке и, следовательно, подавляет споруляцию в этой клетке, позволяя споруляции продолжаться в предспорах. Остаточный spoIIAA в материнской клетке подавляет spoIIAB, но spoIIAB постоянно заменяется, поэтому он продолжает ингибировать споруляцию. Когда полная хромосома локализуется в предспоре, spoIIAB может подавлять сигма F. Следовательно, генетическая асимметрия хромосомы B. subtilis и экспрессия сигма F, spoIIAB и spoIIAA диктуют образование спор у B. subtilis.

Хромосомная репликация
[ редактировать ]Bacillus subtilis — модельный организм , используемый для изучения репликации бактериальных хромосом. Репликация единственной кольцевой хромосомы инициируется в одном локусе — источнике ( oriC ). Репликация происходит в двух направлениях, и две репликационные вилки продвигаются вдоль хромосомы по часовой стрелке и против часовой стрелки. Репликация хромосомы завершается, когда вилки достигают конечной области, которая расположена напротив начала координат на карте хромосомы . Концевая область содержит несколько коротких последовательностей ДНК ( Ter -сайты), которые способствуют остановке репликации. Специфические белки опосредуют все этапы репликации ДНК. Сравнение белков, участвующих в репликации хромосомной ДНК у B. subtilis и Escherichia coli, выявляет сходства и различия. Хотя основные компоненты, способствующие инициации, элонгации и терминации репликации, хорошо консервативны , можно обнаружить некоторые важные различия (например, у одной бактерии отсутствуют белки, необходимые для другой). Эти различия подчеркивают разнообразие механизмов и стратегий, которые различные виды бактерий используют для дублирования своих геномов. [ 25 ]
Геном
[ редактировать ]Bacillus subtilis имеет около 4100 генов. Из них только 192 оказались незаменимыми; еще 79, по прогнозам, также будут необходимы. Подавляющее большинство важнейших генов были отнесены к относительно небольшому числу доменов клеточного метаболизма, при этом около половины участвовало в обработке информации, одна пятая участвовала в синтезе клеточной оболочки и определении формы и деления клеток, а одна десятая часть была связана с клеточными процессами. энергетика. [ 26 ]
Полная последовательность генома субштамма QB928 B. subtilis содержит 4 146 839 пар оснований ДНК и 4 292 гена. Штамм QB928 широко используется в генетических исследованиях благодаря наличию различных маркеров [aroI(aroK)906 purE1 dal(alrA)1 trpC2]. [ 27 ]
было охарактеризовано несколько некодирующих РНК В 2009 году в геноме B. subtilis , включая РНК Bsr . [ 28 ] Сравнительный геномный анализ на основе микрочипов показал, что представители B. subtilis демонстрируют значительное геномное разнообразие. [ 29 ]
FsrA — небольшая РНК, обнаруженная в Bacillus subtilis . Он является эффектором реакции сбережения железа и снижает регуляцию железосодержащих белков в периоды плохой биодоступности железа. [ 30 ] [ 31 ]
Перспективный пробиотик для рыб, штамм Bacillus subtilis WS1A, обладающий антимикробной активностью в отношении Aeromonas veronii и подавляющий подвижную септицемию Aeromonas у Labeo rohita . Сборка de novo привела к получению предполагаемого размера хромосомы 4 148 460 пар оснований с 4 288 открытыми рамками считывания. [ 6 ] [ 7 ] Геном штамма WS1A B. subtilis содержит множество потенциальных генов, например, кодирующих белки, участвующие в биосинтезе рибофлавина , витамина B6 и аминокислот ( ilvD ), а также в утилизации углерода ( pta ). [ 6 ] [ 7 ]
Трансформация
[ редактировать ]Естественная бактериальная трансформация включает перенос ДНК от одной бактерии к другой через окружающую среду. У B. subtilis длина переносимой ДНК превышает 1271 т.п.н. (более 1 млн оснований). [ 32 ] Перенесенная ДНК, вероятно, представляет собой двухцепочечную ДНК и часто составляет более трети общей длины хромосомы, составляющей 4215 т.п.н. [ 33 ] Похоже, что около 7–9% клеток-реципиентов занимают всю хромосому. [ 34 ]
Чтобы бактерия-реципиент связалась, забрала экзогенную ДНК другой бактерии того же вида и рекомбинировала ее в свою хромосому, она должна войти в особое физиологическое состояние, называемое компетентностью . Компетентность B. subtilis индуцируется ближе к концу логарифмического роста, особенно в условиях ограничения аминокислот. [ 35 ] В этих стрессовых условиях полуголодания клетки обычно имеют только одну копию своей хромосомы и, вероятно, имеют повышенное повреждение ДНК. Чтобы проверить, является ли трансформация адаптивной функцией B. subtilis по восстановлению повреждений ДНК, были проведены эксперименты с использованием ультрафиолетового света в качестве повреждающего агента. [ 36 ] [ 37 ] [ 38 ] Эти эксперименты привели к выводу, что компетентность с поглощением ДНК специфически индуцируется условиями, повреждающими ДНК, и что трансформация функционирует как процесс рекомбинационного восстановления повреждений ДНК. [ 39 ]
Хотя естественное компетентное состояние часто встречается среди лабораторных B. subtilis и полевых изолятов , некоторые промышленно значимые штаммы, например B. subtilis (natto), неохотно поглощают ДНК из-за присутствия систем рестрикционной модификации, которые разрушают экзогенную ДНК. Мутанты B. subtilis (natto), дефектные по эндонуклеазе системы рестрикционной модификации I типа, способны выступать в качестве реципиентов конъюгативных плазмид в экспериментах по спариванию, открывая путь для дальнейшей генной инженерии этого конкретного B. subtilis . штамма [ 40 ]
Применяя «зеленую химию» для использования менее опасных материалов и экономя при этом затраты, исследователи имитируют природные методы синтеза химических веществ, которые могут быть полезны для пищевой и фармацевтической промышленности, «соединяя молекулы с короткими нитями ДНК», прежде чем они будут застегнуты на молнию. вместе во время их комплементарного спаривания оснований между двумя цепями. Каждая цепь будет нести определенную интересующую молекулу, которая одновременно подвергнется определенной химической реакции, когда две соответствующие цепи пар ДНК скрепляются, как молния, позволяя другой интересующей молекуле взаимодействовать друг с другом в контролируемой и изолированной реакции между этими молекулами. переносятся в эти комплементарные присоединения ДНК. Используя этот метод с определенными бактериями, которые естественным образом следуют многоэтапному процессу репликации, исследователи могут одновременно продолжать взаимодействие этих добавленных молекул для взаимодействия с ферментами и другими молекулами, используемыми для вторичной реакции, рассматривая их как капсулу. , что похоже на то, как бактерии выполняют свои собственные процессы репликации ДНК. [ 41 ]
Использование
[ редактировать ]20 век
[ редактировать ]
Культуры B. subtilis были популярны во всем мире до появления антибиотиков как иммуностимулирующее средство для лечения заболеваний желудочно- кишечного тракта и мочевыводящих путей . Он использовался на протяжении 1950-х годов в качестве альтернативного лекарства , которое, как было обнаружено, при переваривании значительно стимулирует иммунную активность широкого спектра , включая активацию секреции специфических антител IgM , IgG и IgA. [ 42 ] и высвобождение динуклеотидов CpG, индуцирующих интерферон IFN-α / IFNγ, продуцирующий активность лейкоцитов и цитокинов, важных в развитии цитотоксичности по отношению к опухолевым клеткам . [ 43 ] Он продавался по всей Америке и Европе с 1946 года как иммуностимулирующее средство при лечении заболеваний кишечника и мочевыводящих путей, таких как ротавирус и шигеллез . В 1966 году армия США сбрасывала Bacillus Subtilis на решетки станций нью-йоркского метро на пять дней, чтобы наблюдать за реакцией людей, покрывшихся странной пылью. [ 44 ] Считается, что из-за его способности выживать он до сих пор там присутствует. [ 45 ]
Антибиотик бацитрацин был впервые выделен из разновидности Bacillus licheniformis под названием «Tracy I». [ 46 ] в 1945 г. тогда считался частью вида B. subtilis . Его до сих пор коммерчески производят путем выращивания сорта в контейнере с жидкой питательной средой . Со временем бактерии синтезируют бацитрацин и секретируют антибиотик в среду. Затем бацитрацин экстрагируют из среды с помощью химических процессов. [ 47 ]
С 1960-х годов B. subtilis использовался в качестве экспериментального вида в экспериментах по космическим полетам. Его эндоспоры могут выжить в космосе до 6 лет, если покрыты частицами пыли, защищающими их от солнечных ультрафиолетовых лучей. [ 48 ] Он использовался в качестве индикатора выживания экстремофилов в космическом пространстве , например, в Exobiology Radiation Assembly . [ 49 ] [ 50 ] ЭКЗОСТАК , [ 51 ] [ 52 ] и EXPOSE орбитальных миссий. [ 53 ] [ 54 ] [ 55 ]
дикого типа С природными изолятами B. subtilis труднее работать по сравнению с лабораторными штаммами, прошедшими процессы одомашнивания, мутагенеза и селекции. Эти штаммы часто обладают улучшенными способностями к трансформации (поглощение и интеграция ДНК окружающей среды), росту и утрате способностей, необходимых «в дикой природе». И хотя существуют десятки различных штаммов, соответствующих этому описанию, наиболее широко используется штамм, обозначенный как «168». Штамм 168 является триптофана ауксотрофом , выделенным в результате рентгеновского мутагенеза штамма B. subtilis Marburg и широко используется в исследованиях благодаря высокой эффективности трансформации. [ 56 ]

Bacillus globigii , близкородственный, но филогенетически различный вид, ныне известный как Bacillus atropaeus. [ 57 ] [ 58 ] использовался в качестве симулятора биологической войны во время проекта SHAD (он же Проект 112 ). [ 59 ] Последующий геномный анализ показал, что штаммы, использованные в этих исследованиях, были продуктами преднамеренного обогащения штаммами, демонстрировавшими аномально высокий уровень споруляции . [ 60 ]
Штамм B. subtilis, ранее известный как Bacillus natto, используется в коммерческом производстве японской еды натто , а также аналогичного корейского блюда чхонггукчжан .
21 век
[ редактировать ]- В качестве модельного организма B. subtilis обычно используется в лабораторных исследованиях, направленных на обнаружение фундаментальных свойств и характеристик грамположительных спорообразующих бактерий. [ 29 ] В частности, в результате исследований спорообразования у B. subtilis были выведены основные принципы и механизмы, лежащие в основе образования прочных эндоспор .
- Его свойства связывания с поверхностью играют роль в безопасном удалении радионуклидных отходов [например, тория (IV) и плутония (IV)]. [ нужна ссылка ]
- Благодаря своим превосходным ферментационным свойствам и высокому выходу продукта (от 20 до 25 граммов на литр) его используют для производства различных ферментов, таких как амилаза и протеазы. [ 61 ]
- B. subtilis используется в качестве инокулянта почвы в садоводстве и сельском хозяйстве . [ 62 ] [ 63 ] [ 64 ]
- Это может принести некоторую пользу производителям шафрана , ускоряя рост клубнелуковиц и увеличивая выход биомассы рыльцев. [ 65 ]
- Он используется в качестве «организма-индикатора» во время процедур газовой стерилизации, чтобы гарантировать успешное завершение цикла стерилизации. В частности, эндоспоры B. subtilis используются для проверки того, что цикл достиг условий уничтожения спор. [ 66 ] [ 67 ]
- B. subtilis Было обнаружено, что действует как полезный биопродукт-фунгицид, который предотвращает рост Monilinia vaccinii-corymbosi , также известного как мумие-ягодный гриб, не влияя при этом на опыление или качество плодов. [ 68 ]
- как метаболически активные, так и неметаболически активные клетки B. subtilis Было показано, что восстанавливают золото (III) до золота (I) и золота (0) в присутствии кислорода. Это биотическое восстановление играет роль в круговороте золота в геологических системах и потенциально может быть использовано для извлечения твердого золота из этих систем.
Новые и искусственные субштаммы
[ редактировать ]- Были выделены новые штаммы B. subtilis , которые могли использовать для размножения 4-фтортриптофан (4FTrp), но не канонический триптофан (Trp). Поскольку Trp кодируется только одним кодоном, есть свидетельства того, что Trp может быть заменен 4FTrp в генетическом коде. Эксперименты показали, что канонический генетический код может быть изменчивым. [ 69 ]
- Рекомбинантные штаммы pBE2C1 и pBE2C1AB были использованы в производстве полигидроксиалканоатов (ПГА), а солодовые отходы могут быть использованы в качестве источника углерода для более дешевого производства ПГА. [ нужна ссылка ]
- Он используется для производства гиалуроновой кислоты , которая используется в секторе ухода за суставами в здравоохранении. [ 70 ] и косметика.
- Компания Monsanto выделила из B. subtilis ген , экспрессирующий белок холодового шока B, и включила его в свой засухоустойчивый гибрид кукурузы MON 87460, который был одобрен для продажи в США в ноябре 2011 года. [ 71 ] [ 72 ]
- Новый штамм был модифицирован для преобразования нектара в мед путем секреции ферментов. [ 73 ]
Безопасность
[ редактировать ]У других животных
[ редактировать ]Bacillus subtilis FDA США была проверена Центром ветеринарной медицины и не выявила проблем с безопасностью при использовании в микробных продуктах прямого кормления, поэтому Ассоциация американских чиновников по контролю за кормами включила ее в список одобренных для использования в качестве ингредиента кормов для животных в соответствии с разделом 36.14. «Микроорганизмы прямого питания». [ нужна ссылка ] Секция здоровья животных и производственных кормов Канадского агентства по контролю качества пищевых продуктов классифицировала Bacillus, одобренные кормовые ингредиенты, обезвоженные культуры как добавку для силоса в соответствии с Приложением IV, часть 2, класс 8.6, и присвоила международный номер кормовым ингредиентам IFN 8-19-119. [ нужна ссылка ] С другой стороны, несколько кормовых добавок, содержащих жизнеспособные споры B. subtilis , получили положительную оценку Европейского агентства по безопасности пищевых продуктов в отношении их безопасного использования для увеличения веса в животноводстве.
У людей
[ редактировать ]Споры Bacillus subtilis могут пережить очень высокую температуру, возникающую во время приготовления пищи. Некоторые штаммы B. subtilis ответственны за появление вязкости или порчи веревки – липкой, тягучей консистенции, вызванной выработкой бактериями длинноцепочечных полисахаридов – в испорченном хлебном тесте и хлебобулочных изделиях. [ 74 ] Долгое время B. subtilis биохимические тесты связывали хлебную вязкость исключительно с видами . Молекулярные анализы (ПЦР со случайной амплификацией полиморфной ДНК, анализ электрофореза в денатурирующем градиентном геле и секвенирование области V3 16S рибосомальной ДНК ) выявили большее разнообразие видов Bacillus в рыхлом хлебе, который, по-видимому, обладает положительной амилазной активностью и высокой термостойкостью. [ 75 ]
B. subtilis CU1 (2 × 10 9 спор в день) оценивалось в 16-недельном исследовании (10-дневное введение пробиотика с последующим 18-дневным периодом вымывания каждый месяц; повторение той же процедуры в течение всего 4 месяцев) здоровым субъектам. Было обнаружено, что B. subtilis CU1 безопасен и хорошо переносится субъектами без каких-либо побочных эффектов. [ 76 ]
Bacillus subtilis и полученные из нее вещества были оценены различными авторитетными органами на предмет их безопасного и полезного использования в пищевых продуктах. В Соединенных Штатах в письме-заключении, опубликованном в начале 1960-х годов Управлением по контролю за продуктами и лекарствами (FDA), некоторые вещества, полученные из микроорганизмов, были признаны безопасными (GRAS), включая ферменты карбогидразу и протеазу из B. subtilis . Мнения основывались на использовании непатогенных и нетоксигенных штаммов соответствующих организмов, а также на использовании современной надлежащей производственной практики. [ 77 ] FDA заявило, что ферменты, полученные из штамма B. subtilis , широко использовались в пищевых продуктах до 1 января 1958 года и что нетоксигенные и непатогенные штаммы B. subtilis широко доступны и безопасно используются в различных пищевых целях. . Сюда входит потребление японских ферментированных соевых бобов в форме натто , которые обычно едят в Японии и содержат до 10 8 жизнеспособных клеток на грамм. Ферментированные бобы признаны за их вклад в здоровую кишечную флору и витамина К2 ; потребление За время своей долгой истории широкого использования натто не вызывал побочных эффектов, потенциально связанных с присутствием B. subtilis . [ нужна ссылка ] Продукт натто и натто B. subtilis в качестве его основного компонента представляют собой FOSHU (продукты для специального медицинского использования), одобренные Министерством здравоохранения, труда и социального обеспечения Японии как эффективные для сохранения здоровья. [ 78 ]
Bacillus subtilis получила статус «Квалифицированной презумпции безопасности» Европейского агентства по безопасности пищевых продуктов . [ 79 ]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Эзеби Дж.П. (2008). «Бацилла» . Список названий прокариот, стоящих в номенклатуре . Проверено 18 ноября 2008 г.
- ^ Амброзиано Н (30 июня 1999 г.). «Лабораторные биодетекторные тесты должны быть безопасными, общественность должна быть хорошо информирована» . Пресс-релиз . Лос-Аламосские национальные лаборатории. Архивировано из оригинала 21 сентября 2008 года . Проверено 18 ноября 2008 г.
- ^ «Как произносится бацилла» . Кембриджский словарь .
- ^ «тонкость» . Викисловарь . 10 марта 2023 г.
- ^ Jump up to: а б Эррингтон Дж., Аарт Л.Т. (май 2020 г.). «Профиль микроба: Bacillus subtilis : модельный организм для клеточного развития и промышленная рабочая лошадка» . Микробиология . 166 (5): 425–427. дои : 10.1099/mic.0.000922 . ПМЦ 7376258 . ПМИД 32391747 .
- ^ Jump up to: а б с д и ж г час я Пол С.И., Рахман М.М., Салам М.А., Хан М.А., Ислам М.Т. (15 декабря 2021 г.). «Идентификация бактерий, связанных с морскими губками, на острове Сен-Мартен в Бенгальском заливе с акцентом на профилактику подвижной сепсиса Aeromonas в Лабеорохите». Аквакультура . 545 : 737156. doi : 10.1016/j.aquacultural.2021.737156 . ISSN 0044-8486 .
- ^ Jump up to: а б с д и ж Рахман М.М., Пол С.И., Актер Т., Тай AC, Фойсал MJ, Ислам М.Т. (сентябрь 2020 г.). «Полногеномная последовательность Bacillus subtilis WS1A, многообещающего рыбного пробиотического штамма, выделенного из морской губки Бенгальского залива» . Объявления о ресурсах по микробиологии . 9 (39). дои : 10.1128/mra.00641-20 . ПМЦ 7516141 . ПМИД 32972930 .
- ^ Пол С.И., Рахман М.М. (октябрь 2022 г.). Гилл С.Р. (ред.). «Проект геномной последовательности Bacillus subtilis YBS29, потенциального рыбного пробиотика, который предотвращает септицемию подвижных Aeromonas у Labeo rohita» . Объявления о ресурсах по микробиологии . 11 (10): e0091522. дои : 10.1128/mra.00915-22 . ПМЦ 9583808 . ПМИД 36154193 .
- ^ Эренберг К.Г. (1835). Физические трактаты Королевской академии наук в Берлине за 1833–1835 годы . стр. 145–336.
- ^ Кон Ф. (1872 г.). «Исследования о бактериях» . Вклад в биологию растений . Том 1. стр. 127–224.
- ^ Ю А.С., Лу Дж.Ф., Ю С., Конг С.К., Чан Т.Ф. (январь 2014 г.). «Мониторинг роста бактерий с использованием настраиваемого резистивного импульсного зондирования с использованием метода пор». Прикладная микробиология и биотехнология . 98 (2): 855–62. дои : 10.1007/s00253-013-5377-9 . ПМИД 24287933 . S2CID 2956197 .
- ^ Мэдиган М., Мартинко Дж., ред. (2005). Брок Биология микроорганизмов (11-е изд.). Прентис Холл. ISBN 978-0-13-144329-7 . [ нужна страница ]
- ^ Накано М.М., Зубер П. (1998). «Анаэробный рост «строгих аэробов» ( Bacillus subtilis )». Ежегодный обзор микробиологии . 52 (1): 165–90. дои : 10.1146/аннурев.микро.52.1.165 . ПМИД 9891797 .
- ^ Руис Н., Силхави Т.Дж. (сентябрь 2022 г.). «Как Escherichia coli стала флагманской бактерией молекулярной биологии» . Журнал бактериологии . 204 (9): e0023022. дои : 10.1128/jb.00230-22 . ПМЦ 9487582 . ПМИД 35916528 . S2CID 251254431 .
- ^ Хонг Х.А., Ханеджа Р., Там Н.М., Каццато А., Тан С., Урдачи М., Бриссон А., Гасбаррини А., Барнс И., Каттинг С.М. (март 2009 г.). « Bacillus subtilis, выделенная из желудочно-кишечного тракта человека» . Исследования в области микробиологии . 160 (2): 134–43. дои : 10.1016/j.resmic.2008.11.002 . ПМИД 19068230 .
- ^ Судхагар С., Редди П.Р., Нагалакшми Г. (апрель 2017 г.). «Влияние высоты на структурирование кишечных бактериальных сообществ Apis cerana Fab» (PDF) . Журнал энтомологии и зоологии . 5 (3): 434–440.
- ^ Jump up to: а б с Тан И.С., Рамамурти К.С. (июнь 2014 г.). «Спорообразование у Bacillus subtilis: спорообразование Bacillus subtilis» . Отчеты по экологической микробиологии . 6 (3): 212–225. дои : 10.1111/1758-2229.12130 . ПМК 4078662 . ПМИД 24983526 .
- ^ Маккенни П.Т., Дрикс А., Айхенбергер П. (январь 2013 г.). «Эндоспора Bacillus subtilis: сборка и функции многослойной оболочки» . Обзоры природы. Микробиология . 11 (1): 33–44. дои : 10.1038/nrmicro2921 . ПМК 9910062 . ПМИД 23202530 . S2CID 205498395 .
- ^ Стивенс С. (январь 1998 г.). «Бактериальная споруляция: вопрос обязательств?» . Современная биология . 8 (2): С45–Р48. дои : 10.1016/S0960-9822(98)70031-4 . ПМИД 9427639 . S2CID 14126998 .
- ^ Эрл А.М., Лосик Р., Колтер Р. (июнь 2008 г.). «Экология и геномика Bacillus subtilis» . Тенденции в микробиологии . 16 (6): 269–275. дои : 10.1016/j.tim.2008.03.004 . ПМК 2819312 . ПМИД 18467096 .
- ^ Хиггинс Д., Дворкин Дж. (январь 2012 г.). «Недавний прогресс в спорообразовании Bacillus subtilis» . Обзоры микробиологии FEMS . 36 (1): 131–148. дои : 10.1111/j.1574-6976.2011.00310.x . ПМЦ 3237856 . ПМИД 22091839 .
- ^ Jump up to: а б Слончевски Дж., Джон Уоткинс Фостер, Зинсер Э.Р. 2020. Микробиология: развивающаяся наука. Нью-Йорк: WW Norton & Company.
- ^ Пиггот П.Дж., Кут Дж.Г. (декабрь 1976 г.). «Генетические аспекты образования бактериальных эндоспор» . Бактериологические обзоры . 40 (4): 908–962. дои : 10.1128/MMBR.40.4.908-962.1976 . ПМК 413989 . ПМИД 12736 .
- ^ Джаббари С., Хип Дж.Т., Кинг-младший (январь 2011 г.). «Математическое моделирование сети инициации споруляции у Bacillus subtilis, раскрывающее двойную роль предполагаемой сигнальной молекулы PhrA, воспринимающей кворум» (PDF) . Бюллетень математической биологии . 73 (1): 181–211. дои : 10.1007/s11538-010-9530-7 . ПМИД 20238180 . S2CID 9875633 .
- ^ Нуаро П. (2007). «Репликация хромосомы Bacillus subtilis » . В Грауманне П. (ред.). Бациллы : клеточная и молекулярная биология . Кайстер Академик Пресс. ISBN 978-1-904455-12-7 . [ нужна страница ]
- ^ Кобаяши К., Эрлих С.Д., Альбертини А., Амати Г., Андерсен К.К., Арно М. и др. (апрель 2003 г.). «Основные гены Bacillus subtilis» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (8): 4678–83. Бибкод : 2003PNAS..100.4678K . дои : 10.1073/pnas.0730515100 . JSTOR 3144001 . ПМК 153615 . ПМИД 12682299 .
- ^ Ю К.С., Йим К.Ю., Цуй С.К., Чан Т.Ф. (ноябрь 2012 г.). «Полная последовательность генома штамма Bacillus subtilis QB928, штамма, широко используемого в генетических исследованиях B. subtilis» . Журнал бактериологии . 194 (22): 6308–9. дои : 10.1128/JB.01533-12 . ПМЦ 3486399 . ПМИД 23105055 .
- ^ Сайто С., Какешита Х., Накамура К. (январь 2009 г.). «Новые гены, кодирующие малые РНК, в межгенных областях Bacillus subtilis». Джин . 428 (1–2): 2–8. дои : 10.1016/j.gene.2008.09.024 . ПМИД 18948176 .
- ^ Jump up to: а б Эрл А.М., Лосик Р., Колтер Р. (июнь 2008 г.). «Экология и геномика Bacillus subtilis» . Тенденции в микробиологии . 16 (6): 269–75. дои : 10.1016/j.tim.2008.03.004 . ПМК 2819312 . ПМИД 18467096 .
- ^ Габалла А., Антельманн Х., Агилар С., Ках С.К., Сонг КБ, Смальдоне Г.Т., Хельманн Дж.Д. (август 2008 г.). «Реакция сохранения железа Bacillus subtilis опосредуется небольшой РНК, регулируемой Fur, и тремя небольшими основными белками» . Труды Национальной академии наук Соединенных Штатов Америки . 105 (33): 11927–32. Бибкод : 2008PNAS..10511927G . дои : 10.1073/pnas.0711752105 . ПМК 2575260 . ПМИД 18697947 .
- ^ Смальдоне Г.Т., Антельманн Х., Габалла А., Хельманн Дж.Д. (май 2012 г.). «МРНК FsrA и белок FbpB опосредуют железозависимую индукцию железосеросодержащих оксидаз Bacillus subtilis lutABC» . Журнал бактериологии . 194 (10): 2586–93. дои : 10.1128/JB.05567-11 . ПМК 3347220 . ПМИД 22427629 .
- ^ Сайто Ю., Тагучи Х., Акамацу Т. (март 2006 г.). «Судьба трансформации бактериального генома после включения в компетентные клетки Bacillus subtilis: непрерывная длина включенной ДНК». Журнал бионауки и биоинженерии . 101 (3): 257–62. дои : 10.1263/jbb.101.257 . ПМИД 16716928 .
- ^ Сайто Ю., Тагучи Х., Акамацу Т. (апрель 2006 г.). «ДНК, попадающая в компетентные клетки Bacillus subtilis путем трансформации лизированных протопластов, представляет собой не оцДНК, а дцДНК». Журнал бионауки и биоинженерии . 101 (4): 334–39. дои : 10.1263/jbb.101.334 . ПМИД 16716942 .
- ^ Акамацу Т., Тагучи Х. (апрель 2001 г.). «Включение всей хромосомной ДНК в лизаты протопластов в компетентные клетки Bacillus subtilis» . Бионауки, биотехнологии и биохимия . 65 (4): 823–29. дои : 10.1271/bbb.65.823 . ПМИД 11388459 . S2CID 30118947 .
- ^ Анагностопулос С., Спизизен Дж. (май 1961 г.). «Требования к трансформации в Bacillus Subtilis» . Журнал бактериологии . 81 (5): 741–46. дои : 10.1128/JB.81.5.741-746.1961 . ПМК 279084 . ПМИД 16561900 .
- ^ Хельцер М.А., Мишод Р.Э. (июнь 1991 г.). «Репарация ДНК и эволюция трансформации Bacillus subtilis. III. Секс с поврежденной ДНК» . Генетика . 128 (2): 215–23. дои : 10.1093/генетика/128.2.215 . ПМК 1204460 . ПМИД 1906416 .
- ^ Мишод Р.Э., Войцеховский М.Ф., Хельцер М.А. (январь 1988 г.). «Репарация ДНК и эволюция трансформации бактерии Bacillus subtilis» . Генетика . 118 (1): 31–39. дои : 10.1093/генетика/118.1.31 . ПМЦ 1203263 . ПМИД 8608929 .
- ^ Войцеховский М.Ф., Хельцер М.А., Мишод Р.Э. (март 1989 г.). «Репарация ДНК и эволюция трансформации Bacillus subtilis. II. Роль индуцируемой репарации» . Генетика . 121 (3): 411–22. дои : 10.1093/генетика/121.3.411 . ПМЦ 1203629 . ПМИД 2497048 .
- ^ Мишод Р.Э., Бернштейн Х., Недельку А.М. (май 2008 г.). «Адаптационное значение пола у микробных патогенов». Инфекция, генетика и эволюция . 8 (3): 267–85. дои : 10.1016/j.meegid.2008.01.002 . ПМИД 18295550 .
- ^ Итая М., Нагасаку М., Шимада Т., Отани Н., Шива Ю., Ёсикава Х. и др. (февраль 2019 г.). «Стабильная и эффективная доставка ДНК в Bacillus subtilis (натто) с использованием плазмид конъюгационного переноса pLS20». Письма FEMS по микробиологии . 366 (4). дои : 10.1093/femsle/fnz032 . ПМИД 30726909 .
- ^ «Химия AU naturall: подражание чистым и эффективным способам природы. - Бесплатная онлайн-библиотека» . www.thefreelibrary.com . Проверено 29 апреля 2023 г.
- ^ Чипранди Дж., Скордамалья А., Венути Д., Кария М., Каноника Г.В. (декабрь 1986 г.). «Влияние Bacillus subtilis in vitro на иммунный ответ». Химиотерапия 5 (6): 404–07. ПМИД 3100070 .
- ^ Шилаховенко В.А. (июнь 2003 г.). «Противораковое и иммуностимулирующее действие нуклеопротеиновой фракции Bacillus subtilis» . Экспериментальная онкология . 25 : 119–23.
- ^ Исследование уязвимости пассажиров метро в Нью-Йорке к тайным действиям с биологическими агентами . Разное издание. Департамент армии, Форт-Детрик. 1968.
- ^ Розофф С., Понтелл Х., Тиллман Р. (2020). Прибыль без чести: преступления «белых воротничков» и грабеж Америки . Пирсон. стр. 352–3. ISBN 9780134871486 .
- ^ Подставка А. «Bacillus licheniformis Tracy I | DSM 603, ATCC 10716, CCM 2181, IFO 12199, NBRC 12199, NCIB 8874, FDA BT1 | BacDiveID:686» . bacdive.dsmz.de .
- ^ Джонсон Б.А., Анкер Х., Мелени Флорида (октябрь 1945 г.). «Бацитрацин: новый антибиотик, производимый членом группы B. subtilis». Наука . 102 (2650): 376–7. Бибкод : 1945Sci...102..376J . дои : 10.1126/science.102.2650.376 . ПМИД 17770204 . S2CID 51066 .
- ^ Хорнек Дж., Клаус Д.М., Манчинелли Р.Л. (март 2010 г.). «Космическая микробиология» . Обзоры микробиологии и молекулярной биологии . 74 (1): 121–56. Бибкод : 2010MMBR...74..121H . дои : 10.1128/mmbr.00016-09 . ПМЦ 2832349 . ПМИД 20197502 .
- ^ Доза К, Бигер-Доза А, Диллманн Р., Гилл М., Керц О., Кляйн А. и др. (1995). «ЭРА-эксперимент «Космическая биохимия» ». Достижения в космических исследованиях . 16 (8): 119–29. Бибкод : 1995АдСпР..16х.119Д . дои : 10.1016/0273-1177(95)00280-Р . ПМИД 11542696 .
- ^ Вайсберг О., Федоров А., Дунюшкин Ф., Кожуховский А., Смирнов В., Аванов Л. и др. (1995). «Ионные популяции в хвосте Венеры». Достижения в космических исследованиях . 16 (4): 105–18. Бибкод : 1995АдСпР..16д.105В . дои : 10.1016/0273-1177(95)00217-3 .
- ^ Клэнси П. (23 июня 2005 г.). В поисках жизни, в поисках Солнечной системы . Издательство Кембриджского университета. [ нужна страница ]
- ^ Хорнек Дж., Клаус Д.М., Манчинелли Р.Л. (март 2010 г.). «Космическая микробиология» . Обзоры микробиологии и молекулярной биологии . 74 (1): 121–56. Бибкод : 2010MMBR...74..121H . дои : 10.1128/MMBR.00016-09 . ПМЦ 2832349 . ПМИД 20197502 .
- ^ Фахардо-Кавасос П., Линк Л., Мелош Х.Дж., Николсон В.Л. (декабрь 2005 г.). «Споры Bacillus subtilis на искусственных метеоритах выдерживают вход в атмосферу на сверхскорости: последствия для литопанспермии». Астробиология . 5 (6): 726–36. Бибкод : 2005AsBio...5..726F . дои : 10.1089/ast.2005.5.726 . ПМИД 16379527 .
- ^ Брандштеттер Ф., Брэк А., Бальони П., Кокелл К.С., Деметс Р., Эдвардс Х.Г. и др. (2008). «Минералогическое изменение искусственных метеоритов при входе в атмосферу. Эксперимент STONE-5». Планетарная и космическая наука . 56 (7): 976–84. Бибкод : 2008P&SS...56..976B . CiteSeerX 10.1.1.549.4307 . дои : 10.1016/j.pss.2007.12.014 .
- ^ Вассманн М., Мёллер Р., Раббоу Э., Паниц С., Хорнек Г., Райц Г. и др. (май 2012 г.). «Выживание спор устойчивого к УФ-излучению штамма Bacillus subtilis MW01 после пребывания на низкой околоземной орбите и смоделированных марсианских условиях: данные космического эксперимента ADAPT на EXPOSE-E». Астробиология . 12 (5): 498–507. Бибкод : 2012AsBio..12..498W . дои : 10.1089/ast.2011.0772 . ПМИД 22680695 .
- ^ Зейглер Д.Р., Прагай З., Родригес С., Шеврё Б., Глушитель А., Альберт Т. и др. (ноябрь 2008 г.). «Происхождение 168, W23 и других устаревших штаммов Bacillus subtilis» . Журнал бактериологии . 190 (21): 6983–95. дои : 10.1128/JB.00722-08 . ПМК 2580678 . ПМИД 18723616 .
- ^ Накамура Л.К. (1989). «Таксономическая связь чернопигментированных штаммов Bacillus subtilis и предложение по Bacillus atropaeus sp. nov» . Международный журнал систематической бактериологии . 39 (3): 295–300. дои : 10.1099/00207713-39-3-295 .
- ^ Берк С.А., Райт Дж.Д., Робинсон М.К., Бронк Б.В., Уоррен Р.Л. (май 2004 г.). «Обнаружение молекулярного разнообразия в Bacillus aтрофеус с помощью анализа полиморфизма длин амплифицированных фрагментов» . Прикладная и экологическая микробиология . 70 (5): 2786–90. Бибкод : 2004ApEnM..70.2786B . дои : 10.1128/АЕМ.70.5.2786-2790.2004 . ПМК 404429 . ПМИД 15128533 .
- ^ «Проект 112/ШАД – Корабельная защита и защита» . Министерство по делам ветеранов США. Архивировано из оригинала 21 февраля 2015 года . Проверено 25 февраля 2015 г.
- ^ Гиббонс Х.С., Брумолл С.М., МакНью Л.А., Далигот Х., Чепмен С., Брюс Д., Каравис М., Креппс М., МакГрегор П.А., Хонг С., Парк К.Х., Акмал А., Фельдман А., Лин Дж.С., Чанг В.Е., Хиггс Б.В., Демирев П. , Линдквист Дж., Лием А., Фоклер Э., Рид Т.Д., Тапиа Р., Джонсон С., Бишоп-Лилли К.А., Деттер С., Хан С., Сожаманнан С., Розенцвейг К.Н., Сковронски Е.В. (март 2011 г.). «Геномные признаки отбора и улучшения штаммов Bacillus atropaeus var. globigii, исторического симулятора биологической войны» . ПЛОС ОДИН . 6 (3): e17836. Бибкод : 2011PLoSO...617836G . дои : 10.1371/journal.pone.0017836 . ПМК 3064580 . ПМИД 21464989 .
- ^ ван Дейл Дж. М., Хекер М. (январь 2013 г.). «Bacillus subtilis: от почвенной бактерии к фабрике сверхсекретирующих клеток» . Заводы по производству микробных клеток . 12 (3): 3. дои : 10.1186/1475-2859-12-3 . ПМК 3564730 . ПМИД 23311580 .
- ^ «Монилиния фруктовая» (PDF) . Паспорта карантинных вредных организмов . Европейская прокуратура (EPPO). Архивировано из оригинала (PDF) 4 июня 2015 г. Проверено 21 июля 2015 г.
- ^ Суэйн М.Р., Рэй Р.К. (2009). «Биоконтроль и другие полезные действия Bacillus subtilis, выделенных из микрофлоры коровьего навоза». Микробиологические исследования . 164 (2): 121–30. дои : 10.1016/j.micres.2006.10.009 . ПМИД 17320363 .
- ^ Янез-Мендисабал V (2011). «Биологический контроль коричневой гнили персика (Monilinia spp.) с помощью Bacillus subtilis CPA-8 основан на выработке фенгициноподобных липопептидов». Европейский журнал патологии растений . 132 (4): 609–19. дои : 10.1007/s10658-011-9905-0 . S2CID 15761522 .
- ^ Шараф-Элдин М., Элхоли С., Фернандес Х.А., Юнге Х., Читам Р., Гвардиола Дж., Уэзерс П. (август 2008 г.). «Bacillus subtilis FZB24 влияет на количество и качество цветков шафрана (Crocus sativus)» . Планта Медика . 74 (10): 1316–20. дои : 10.1055/s-2008-1081293 . ПМЦ 3947403 . ПМИД 18622904 .
- ^ «Международная фармакопея – Четвертое приложение: Методы анализа: 5. Фармацевтические технические процедуры: 5.8 Методы стерилизации» . Архивировано из оригинала 8 декабря 2008 года.
- ^ «Биологический индикатор ЭО АН-2203 (25 шт. в коробке)» . Продукция Андерсен . Архивировано из оригинала 6 октября 2013 г. Проверено 24 мая 2013 г.
- ^ Нгуги Х.К., Дедей С., Делаплейн К.С., Савелль А.Т., Шерм Х. (01.04.2005). «Влияние биофунгицида Серенада (Bacillus subtilis), применяемого для цветов, на переменные, связанные с опылением голубики кроличьего глаза». Биологический контроль . 33 (1): 32–38. doi : 10.1016/j.biocontrol.2005.01.002 . ISSN 1049-9644 .
- ^ Ю А.К., Йим А.К., Мат В.К., Тонг А.Х., Лок С., Сюэ Х., Цуй С.К., Вонг Дж.Т., Чан Т.Ф. (март 2014 г.). «Мутации, позволяющие заменить триптофан на 4-фтортриптофан как каноническую аминокислоту генетического кода» . Геномная биология и эволюция . 6 (3): 629–41. дои : 10.1093/gbe/evu044 . ПМЦ 3971595 . ПМИД 24572018 .
- ^ «Часто задаваемые вопросы по гиалуронату натрия, часто задаваемые вопросы по гиалуроновой кислоте, HA – Hyasis® | Novozymes Biopharma» . Архивировано из оригинала 28 августа 2013 г. Проверено 13 августа 2013 г.
- ^ Харриган Г.Г., Ридли В.П., Миллер К.Д., Сорбет Р., Риордан С.Г., Немет М.А. и др. (октябрь 2009 г.). «Корм и зерно MON 87460, засухоустойчивого гибрида кукурузы, по составу эквивалентны фуражу обычной кукурузы». Журнал сельскохозяйственной и пищевой химии . 57 (20): 9754–63. дои : 10.1021/jf9021515 . ПМИД 19778059 .
- ^ Министерство сельского хозяйства США: Определение нерегулируемого статуса кукурузы MON 87460 ( Zea mays L)
- ^ Блюм Б (17 ноября 2019 г.). «Израильские студенты получили награду за приготовление меда без пчел» . Израиль21c . Проверено 24 ноября 2019 г.
- ^ «Порча веревки | Процессы выпечки» . БЕЙКЕРпедия . 21 июня 2016 года . Проверено 7 февраля 2021 г.
- ^ Пепе О, Блайотта Дж, Москетти Дж, Греко Т, Виллани Ф (апрель 2003 г.). «Штаммы Bacillus spp., продуцирующие веревку из пшеничного хлеба, и стратегия борьбы с ними с помощью молочнокислых бактерий» . Прикладная и экологическая микробиология . 69 (4): 2321–9. Бибкод : 2003ApEnM..69.2321P . дои : 10.1128/АЕМ.69.4.2321-2329.2003 . ПМК 154770 . ПМИД 12676716 .
- ^ Лефевр М., Раседо С.М., Денайроль М., Риперт Г., Дефужер Т., Лобах А.Р. и др. (февраль 2017 г.). «Оценка безопасности Bacillus subtilis CU1 для использования в качестве пробиотика у людей» . Нормативная токсикология и фармакология . 83 : 54–65. дои : 10.1016/j.yrtph.2016.11.010 . ПМИД 27825987 .
- ^ «Неполный список микроорганизмов FDA» . Управление по контролю за продуктами и лекарствами. 2002.
- ^ Шортт С. (сентябрь 2005 г.). «Перспективы пищевых продуктов для конкретного использования в здравоохранении (FOSHU)». В Гибсоне GR (ред.). Бюллетень пищевой науки и технологий: Функциональные продукты питания . Том. 1. Чтение: Издательство IFIS. стр. 7–1. ISBN 978-0-86014-193-8 .
- ^ Группа экспертов EFSA по биологическим опасностям (BIOHAZ) (2010). «Научное заключение о ведении списка микроорганизмов QPS, намеренно добавляемых в пищу или корм (обновление 2010 г.)» . Журнал EFSA . 8 (12): 1944. doi : 10.2903/j.efsa.2010.1944 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Bacillus subtilis, на Викискладе?
СМИ, связанные с Bacillus subtilis, на Викискладе? - Subti Wiki «актуальная информация обо всех генах Bacillus subtilis »
- Окончательная оценка риска Bacillus subtilis на сайте EPA.gov. Архивировано из оригинала 9 сентября 2015 г.
- Bacillus subtilis Браузер генома
- Типовой штамм Bacillus subtilis в Bac Dive - база метаданных бактериального разнообразия