Радиорезистентность
Эта статья нуждается в дополнительных цитатах для проверки . ( май 2017 г. ) |
Радиорезистентность – это уровень ионизирующего излучения , который организмы способны выдержать .
Организмы, устойчивые к ионизирующему излучению (IRRO), были определены как организмы, для которых доза острой ионизирующей радиации (IR), необходимая для достижения 90% снижения (D10), превышает 1000 грей (Гр). [1]
Радиорезистентность у многих организмов удивительно высока, в отличие от ранее существовавших представлений. Например, исследование окружающей среды, животных и растений вокруг зоны чернобыльской катастрофы выявило неожиданное выживание многих видов, несмотря на высокий уровень радиации. Бразильское исследование , проведенное на холме в штате Минас-Жерайс , где наблюдается высокий уровень естественной радиации от месторождений урана, также выявило множество радиорезистентных насекомых , червей и растений. [2] [3] Некоторые экстремофилы , такие как бактерии Deinococcus radiodurans и тихоходки , могут выдерживать большие дозы ионизирующей радиации , порядка 5000 Гр . [4] [5] [6]
Индуцированная радиорезистентность
[ редактировать ]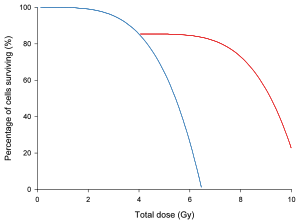
На графике слева кривая доза/выживаемость для гипотетической изображена группы клеток с временем отдыха для восстановления клеток и без него. Если не считать времени восстановления в середине облучения, с клетками обращались бы одинаково.
Радиорезистентность может быть вызвана воздействием небольших доз ионизирующего излучения. Несколько исследований задокументировали этот эффект на дрожжах , бактериях , простейших , водорослях , растениях, насекомых, а также в in vitro клетках млекопитающих и человека и на животных моделях . Могут быть задействованы несколько механизмов клеточной радиозащиты, такие как изменения уровней некоторых цитоплазматических и ядерных белков и усиление экспрессии генов , репарация ДНК и другие процессы. Также биофизические модели представили общие основы этого явления. [7]
Было обнаружено, что многие организмы обладают механизмом самовосстановления, который в некоторых случаях может быть активирован под воздействием радиации. Ниже описаны два примера этого процесса самовосстановления у людей.
Деваир Алвес Феррейра получил большую дозу (7,0 Гр ) во время аварии в Гоянии и выжил, тогда как его жена, получившая дозу 5,7 Гр, умерла. Наиболее вероятное объяснение [ нужна ссылка ] заключается в том, что его доза была разделена на множество меньших доз, которые поглощались в течение определенного периода времени, в то время как его жена больше оставалась дома и подвергалась непрерывному облучению без перерыва, что давало механизмам самовосстановления в ее организме меньше времени на восстановление некоторых из них. ущерб, причиненный радиацией. Это привело к ее смерти. В конце концов он также умер в 1994 году. Точно так же, как некоторые люди, работавшие в подвалах разрушенной Чернобыля, накопили дозы в 10 Гр, эти рабочие получали эти дозы небольшими фракциями, чтобы избежать острых эффектов.
было обнаружено В экспериментах по радиационной биологии , что если облучить группу клеток, то с увеличением дозы количество выживших клеток уменьшается. Также было обнаружено, что если популяция клеток получает дозу перед тем, как ее отложить (без облучения) на некоторое время перед повторным облучением, то радиация будет иметь меньшую способность вызывать гибель клеток . Человеческое тело содержит множество типов клеток , и человек может погибнуть в результате потери одной ткани жизненно важного органа. [ нужна ссылка ] . Для многих краткосрочных радиационных смертей (от 3 до 30 дней) гибель клеток, образующих клетки крови ( костный мозг ) и клеток пищеварительной системы (стенка кишечника ) , приводит к смерти.
Наследование радиорезистентности
[ редактировать ]Имеются убедительные доказательства того, что радиорезистентность может быть генетически детерминирована и наследственна, по крайней мере, у некоторых организмов. Генрих Нётель, генетик из Свободного университета Берлина, провел самое обширное исследование мутаций радиорезистентности с использованием обыкновенной плодовой мушки Drosophila melanogaster в серии из 14 публикаций.
Эволюция радиорезистентности
[ редактировать ]С точки зрения истории эволюции и причинно-следственных связей, радиорезистентность не является адаптивным признаком, поскольку не существует документально подтвержденного естественного давления отбора, которое могло бы дать преимущество в приспособленности способности организмов противостоять дозам ионизирующей радиации в диапазоне, превышающем несколько единиц. Было замечено, что экстремофильные виды способны выживать. [8] Это связано прежде всего с тем, что магнитное поле Земли защищает всех ее обитателей от солнечной космической радиации и галактических космических лучей. [9] которые являются двумя основными источниками ионизирующего излучения в нашей Солнечной системе, [10] и даже включая все документально подтвержденные наземные источники ионизирующего излучения, такие как газ радон и первичные радионуклиды в географических точках, считающихся естественными местами с высоким уровнем радиации, годовая доза естественного радиационного фона [11] остается в десятки тысяч раз меньшими, чем уровни ионизирующей радиации, которые могут выдержать многие высокорадиорезистентные организмы.
Одним из возможных объяснений существования радиорезистентности является то, что она является примером кооптированной адаптации или экзаптации, где радиорезистентность может быть косвенным следствием эволюции другой, связанной адаптации, которая была положительно отобрана в ходе эволюции. Например, гипотеза адаптации к высыханию предполагает, что экстремальные температуры, присутствующие в местах обитания гипертермофилов, таких как Deinococcus radiodurans, вызывают повреждение клеток, практически идентичное повреждению, обычно вызываемому ионизирующей радиацией, и что механизмы восстановления клеток, которые развились для восстановления этого тепла, или повреждение от высыхания также можно обобщить на радиационное повреждение, что позволяет D. radiodurans выдерживать экстремальные дозы ионизирующей радиации. [12] Воздействие гамма-излучения приводит к повреждению клеточной ДНК, включая изменения в спаривании азотистых оснований, повреждение сахаро-фосфатного остова и двухцепочечные повреждения ДНК. [13] Чрезвычайно эффективные механизмы восстановления клеток, которые Deinoccocus виды , такие как D. radiodurans, развили для восстановления тепловых повреждений, вероятно, также способны обращать вспять последствия повреждения ДНК, вызванного ионизирующим излучением, например, путем объединения воедино любых компонентов их генома, которые были повреждены. фрагментированные радиацией. [14] [15] [16]
Бацилла сп. производящие необычно устойчивые к радиации (и перекиси) споры, были выделены из сборочных цехов космических кораблей и рассматриваются как кандидаты, которые могли бы путешествовать на космических кораблях посредством межпланетного перемещения. [17] [18] [19] [20] [21] Анализ генома некоторых из этих производителей радиационно-устойчивых спор пролил некоторый свет на генетические особенности, которые могут быть ответственны за наблюдаемую устойчивость. [22] [23] [24] [25]
Радиорезистентность в радиационной онкологии
[ редактировать ]Радиорезистентность – это также термин, который иногда используется в медицине ( онкологии ) для обозначения раковых клеток, которые трудно лечить лучевой терапией . Радиорезистентность раковых клеток может быть внутренней или индуцированной самой лучевой терапией.
Сравнение радиорезистентности
[ редактировать ]Сравнение в таблице ниже предназначено только для того, чтобы дать приблизительные данные о радиорезистентности различных видов, и к нему следует относиться с большой осторожностью. Как правило, между экспериментами существуют большие различия в радиорезистентности одного вида из-за воздействия радиации на живые ткани и разных условий эксперимента. Например, нам следует учитывать, что, поскольку радиация препятствует делению клеток, незрелые организмы менее устойчивы к радиации, чем взрослые, и взрослые особи стерилизуются дозами, намного меньшими, чем те, которые необходимы для их уничтожения. Например, для насекомого-паразитоида Habrobracon hebetor ЛД 50 для гаплоидного зародыша в период дробления (1-3-часовой возраст) составляет 200 Р , а примерно через 4 часа - 7000 Р (при интенсивности рентгеновского излучения 110 Р/ минуту), а гаплоидные (= мужские) эмбрионы более устойчивы, чем диплоидные (= женские) эмбрионы. [26] Смертность взрослых H. hebetor , подвергшихся дозе 180 250 Р , такая же, как и в необлученной контрольной группе (ни одной из групп питание не предоставлялось) (при 6 000 Р/мин). [27] [28] Однако более низкой дозы в 102 000 Р (за 6 000 Р/мин) достаточно, чтобы вызвать у H. hebetor состояние летаргии , проявляющееся полным прекращением активности, в том числе прекращением питания, и эти особи в конечном итоге позволяют себе голодать до смерть. [28] А еще меньшая доза 4858 Р (при 2650 Р/мин) достаточна для стерилизации взрослой самки H. hebetor (стерильность возникает через 3 дня после заражения). [29] Другие важные факторы, влияющие на уровень радиорезистентности, включают: продолжительность времени, в течение которого доставляется доза радиации — дозы, доставляемые в течение более длительных периодов времени или через определенные промежутки времени, связаны со значительно меньшими отрицательными эффектами; [29] [30] Состояние кормления особей: особи, накормленные до и после кормления, более устойчивы к радиации по сравнению с голодающими особями; [29] [30] тип используемого излучения (например, тихоходки Milnesium tardigradum , облученные тяжелыми ионами, имеют более высокую выживаемость, чем при облучении гамма-лучами, при той же дозе облучения); [31] Физиологическое состояние особей (например, тихоходки Richtersius coronifer и Milnesium tardigradum более устойчивы к гамма-излучению в гидратированном состоянии, а Macrobiotus areolatus более устойчивы к рентгеновскому излучению в ангидробиотическом состоянии). [31] Способ измерения летальности также является источником вариаций предполагаемой радиорезистентности вида. Облученные образцы не погибают мгновенно, если только они не подвергаются очень высокой дозе (острая доза). [32] Следовательно, облученные особи погибают в течение определенного периода времени, а более низкие дозы облучения соответствуют большей выживаемости. Это означает, что доза облучения LD 50 колеблется в зависимости от времени ее измерения. Например, доза β-излучения, вызывающая 50% смертность американского таракана через 25 дней после воздействия, составляет 5700 Р, но для достижения 50% смертности через 3 дня после воздействия необходимо 45610 Р. [30] 25 дней могут представлять собой длительный период выживания для короткоживущих видов, таких как насекомые, но будут представлять собой очень короткое время выживания для долгоживущих видов, таких как млекопитающие, поэтому сравнение выживаемости различных видов через такое же количество времени после воздействия также создает некоторые проблемы интерпретации. Эти примеры иллюстрируют множество проблем, связанных со сравнением радиорезистентности разных видов, и необходимость проявлять при этом осторожность.
| Организм | Смертельная доза | ЛД 50 | ЛД 100 | Класс/Королевство |
|---|---|---|---|---|
| Собака | 3,5 (ЛД 50/30 дней ) [33] | Млекопитающие | ||
| Человек | 4–10 [34] | 4.5 [35] | 10 [36] | Млекопитающие |
| Крыса | 7.5 | Млекопитающие | ||
| Мышь | 4.5–12 | 8.6–9 | Млекопитающие | |
| Кролик | 8 (LD 50/30 дней ) [33] | Млекопитающие | ||
| Черепаха | 15 (LD 50/30 дней ) [33] | Рептилия | ||
| Золотая рыбка | 20 (LD 50/30 дней ) [33] | Рыба | ||
| кишечная палочка | 60 | 60 | Бактерии | |
| Немецкий таракан | 64 [34] | Насекомые | ||
| Моллюски | 200 (LD 50/30 дней ) [33] | - | ||
| Обыкновенная плодовая мушка | 640 [34] | Насекомые | ||
| К. Элеганс ∗ | 160-200 [37] | ≫ 500-800 [38] [39] | Нематода | |
| Амеба | 1000 ( 50/30 дней LD ) [33] | - | ||
| Хабробракон гебетор | 1,800 [27] [28] | Насекомые | ||
| Милнезиум тихоходка | 5,000 [31] | Евтардиградный | ||
| Дейнококк радиодуранс | 15,000 [34] | Бактерии | ||
| Термококк гамматолеранс | 30,000 [34] | Архея |
о ЛД 50 дикого типа сообщалось * Хотя для особей C. elegans , верхний предел летальности не установлен, скорее «почти все животные были живы без признаков избыточной летальности вплоть до 800 Гр, самой высокой дозы... измеренной». ." [39]
См. также
[ редактировать ]- Ex-Rad - препарат радиозащитный , изученный на предмет его способности защищать от острого лучевого синдрома.
- CBLB502 аналогичный радиозащитный препарат, защищающий от острого лучевого синдрома во время лучевой терапии .
- Радиочувствительность
- Фоновое излучение
- Радиационный гормезис
- Радиотрофный гриб
- Койевая кислота
Примечания и ссылки
[ редактировать ]- ^ Сгайер, Хайтам; Гедира, Кайс; Бенкахла, Алия; Баркалла, Инсаф (2008). «Механизм базальной репарации ДНК подлежит положительному отбору у бактерий, устойчивых к ионизирующему излучению» . БМК Геномика . 9 : 297. дои : 10.1186/1471-2164-9-297 . ПМК 2441631 . ПМИД 18570673 .
- ^ Кордейро, Арканзас; Маркес, ЕК; Вейга-Нето, AJ (1973). «Радиорезистентность природной популяции Drosophila willistoni, обитающей в радиоактивной среде». Мутационные исследования . 19 (3): 325–9. дои : 10.1016/0027-5107(73)90233-9 . ПМИД 4796403 .
- ^ Мустакки, Э (1965). «Индукция физическими и химическими агентами мутаций радиорезистентности у Saccharomyces cerevisiae». Мутационные исследования . 2 (5): 403–12. дои : 10.1016/0027-5107(65)90052-7 . ПМИД 5878261 .
- ^ Мозли БЭБ; Маттингли А. (1971). «Репарация облученной трансформирующей дезоксирибонуклеиновой кислоты дикого типа и радиационно-чувствительного мутанта Micrococcus radiodurans» . Дж. Бактериол. 105 (3): 976–83. дои : 10.1128/JB.105.3.976-983.1971 . ПМК 248526 . ПМИД 4929286 .
- ^ Мюррей RGE. 1992. Семейство Deinococcaceae. В «Прокариотах» под ред. А. Баллоуз, Х. Г. Трупер, М. Дворкин, В. Хардер, К. Х. Шляйфер 4: 3732–44. Нью-Йорк: Спрингер-Верлаг
- ^ Ито Х; Ватанабэ Х; Такешия М; Иидзука Х (1983). «Выделение и идентификация радиационно-устойчивых кокков рода Deinococcus из осадков сточных вод и кормов для животных» . Сельскохозяйственная и биологическая химия . 47 (6): 1239–47. дои : 10.1271/bbb1961.47.1239 .
- ^ Форнальски К.В. (2019). «Радиационная адаптивная реакция и рак: с точки зрения статистической физики». Физический обзор E . 99 (2): 022139. Бибкод : 2019PhRvE..99b2139F . дои : 10.1103/PhysRevE.99.022139 . ПМИД 30934317 . S2CID 91187501 .
- ^ Анитори, Роберто Пол (2012). Экстремофилы: микробиология и биотехнология . Горизонт Научная Пресса. ISBN 9781904455981 .
- ^ Мукерджи, Саумитра (3 декабря 2008 г.). «Космическое влияние на солнечно-земную среду» . Датчики (Базель, Швейцария) . 8 (12): 7736–7752. Бибкод : 2008Senso...8.7736M . дои : 10.3390/s8127736 . ISSN 1424-8220 . ПМК 3790986 . ПМИД 27873955 .
- ^ Кеннеди, Энн Р. (1 апреля 2014 г.). «Биологическое действие космической радиации и разработка эффективных мер противодействия» . Науки о жизни в космических исследованиях . 1 :10–43. Бибкод : 2014ЛССР....1...10К . дои : 10.1016/j.lssr.2014.02.004 . ISSN 2214-5524 . ПМК 4170231 . ПМИД 25258703 .
- ^ Шахбази-Гахруи, Дариуш; Голами, Мехрдад; Сетаянде, Самане (01 января 2013 г.). «Обзор естественного радиационного фона» . Передовые биомедицинские исследования . 2 (1): 65. дои : 10.4103/2277-9175.115821 . ISSN 2277-9175 . ПМЦ 3814895 . ПМИД 24223380 .
- ^ Маттимор, В.; Баттиста-младший (февраль 1996 г.). «Радиорезистентность Deinococcus radiodurans: функции, необходимые для выживания при ионизирующей радиации, также необходимы для выживания при длительном высыхании» . Журнал бактериологии . 178 (3): 633–637. дои : 10.1128/jb.178.3.633-637.1996 . ISSN 0021-9193 . ПМК 177705 . ПМИД 8550493 .
- ^ Фридберг, Эррол К.; Фридберг, ЕС; Уокер, GC; Уокер, Грэм К.; Сиде, Вольфрам; Вольфрам, Сиде (1995). Репарация ДНК и мутагенез . АСМ Пресс. ISBN 9781555810887 .
- ^ Минтон, КВ (июль 1994 г.). «Репарация ДНК чрезвычайно радиорезистентной бактерии Deinococcus radiodurans» . Молекулярная микробиология . 13 (1): 9–15. дои : 10.1111/j.1365-2958.1994.tb00397.x . ISSN 0950-382X . ПМИД 7984097 .
- ^ Слэйд, Деа; Радман, Мирослав (март 2011 г.). «Устойчивость к окислительному стрессу у Deinococcus radiodurans» . Обзоры микробиологии и молекулярной биологии . 75 (1): 133–191. дои : 10.1128/MMBR.00015-10 . ISSN 1098-5557 . ПМК 3063356 . ПМИД 21372322 .
- ^ Агапов А.А.; Кульбачинский А.В. (октябрь 2015 г.). «Механизмы стрессоустойчивости и генной регуляции радиорезистентной бактерии Deinococcus radiodurans». Биохимия. Биохимия . 80 (10): 1201–1216. дои : 10.1134/S0006297915100016 . ISSN 1608-3040 . ПМИД 26567564 . S2CID 14981740 .
- ^ Ла Дюк М.Т., Николсон В., Керн Р., Венкатешваран К. (2003). «Микробная характеристика космического корабля Mars Odyssey и его инкапсуляционной установки». Энвайрон Микробиол . 5 (10): 977–85. дои : 10.1046/j.1462-2920.2003.00496.x . ПМИД 14510851 .
- ^ Линк Л., Сойер Дж., Венкатешваран К., Николсон В. (февраль 2004 г.). «Чрезвычайная устойчивость спор к УФ-излучению изолятов Bacillus pumilus , полученных на сверхчистом объекте сборки космических кораблей». Микроб Экол . 47 (2): 159–163. дои : 10.1007/s00248-003-1029-4 . ПМИД 14502417 . S2CID 13416635 .
- ^ Кемпф М.Дж., Чен Ф., Керн Р., Венкатешваран К. (июнь 2005 г.). «Периодическое выделение устойчивых к перекиси водорода спор Bacillus pumilus на сборочном предприятии космических кораблей». Астробиология . 5 (3): 391–405. Бибкод : 2005AsBio...5..391K . дои : 10.1089/ast.2005.5.391 . ПМИД 15941382 .
- ^ Ньюкомб Д.А., Шуергер А.С., Бенардини Дж.Н., Дикинсон Д., Таннер Р., Венкатешваран К. (декабрь 2005 г.). «Выживание микроорганизмов, связанных с космическим кораблем, в условиях искусственного марсианского УФ-облучения» . Appl Environ Microbiol . 71 (12): 8147–8156. Бибкод : 2005ApEnM..71.8147N . дои : 10.1128/АЕМ.71.12.8147-8156.2005 . ПМЦ 1317311 . ПМИД 16332797 .
- ^ Гош С., Осман С., Вайшампаян П., Венкатешваран К. (2010). «Периодическое выделение экстремотолерантных бактерий из чистой комнаты, где собирались компоненты космического корабля Феникс» (PDF) . Астробиология . 10 (3): 325–35. Бибкод : 2010AsBio..10..325G . дои : 10.1089/ast.2009.0396 . hdl : 2027.42/85129 . ПМИД 20446872 .
- ^ Джоя Дж., Йеррапрагада С., Цинь X и др. (сентябрь 2007 г.). «Парадоксальная репарация ДНК и сохранение генов устойчивости к перекиси у Bacillus pumilus SAFR-032» . ПЛОС ОДИН . 2 (9:e928): e928. Бибкод : 2007PLoSO...2..928G . дои : 10.1371/journal.pone.0000928 . ПМК 1976550 . ПМИД 17895969 .
- ^ Тирумалай М.Р., Растоги Р., Замани Н., О'Брайант Уильямс Э., Аллен С., Диуф Ф., Квенде С., Вайнсток ГМ, Венкатешваран К.Дж., Фокс Дж.Е. (июнь 2013 г.). «Гены-кандидаты, которые могут быть ответственны за необычную устойчивость, проявляемую спорами Bacillus pumilus SAFR-032» . ПЛОС ОДИН . 8 (6:e66012): e66012. Бибкод : 2013PLoSO...866012T . дои : 10.1371/journal.pone.0066012 . ПМЦ 3682946 . ПМИД 23799069 .
- ^ Тирумалай М.Р., Fox GE (сентябрь 2013 г.). «Элемент, подобный ICEBs1, может быть связан с чрезвычайной устойчивостью спор Bacillus pumilus SAFR-032 к радиации и высыханию». Экстремофилы . 17 (5): 767–774. дои : 10.1007/s00792-013-0559-z . ПМИД 23812891 . S2CID 8675124 .
- ^ Тирумалай М.Р., Степанов В.Г., Вюнше А., Монтазари С., Гонсалес Р.О., Венкатешваран К., Фокс Г.Е. (июнь 2018 г.). « Б. Safensis FO-36b Т и B. pumilus SAFR-032: Сравнение всего генома двух изолятов на объекте сборки космического корабля» . BMC Microbiol . 18 (57): 57. : 10.1186 /s12866-018-1191-y . PMC 5994023. . PMID 29884123 doi
- ^ Кларк, AM; Митчелл, CJ (1952). «Воздействие рентгеновских лучей на гаплоидные и диплоидные эмбрионы хабробракона » . Биологический бюллетень . 103 (2): 170–177. дои : 10.2307/1538443 . JSTOR 1538443 .
- ^ Jump up to: а б Салливан, Р; Грош, Д. (1953). «Радиационная толерантность взрослой осы». Нуклеотики . 11 : 21–23.
- ^ Jump up to: а б с Грош, Д.С. (1954). «Вызванная летаргия и радиационный контроль насекомых». Журнал экономической энтомологии . 49 (5): 629–631. дои : 10.1093/джи/49.5.629 .
- ^ Jump up to: а б с Грош, Д.С.; Салливан, Р.Л. (1954). «Количественные аспекты постоянной и временной стерильности, вызванной у самок Хабробракона рентгеновскими лучами и β-излучением». Радиационные исследования . 1 (3): 294–320. Бибкод : 1954РадР....1..294Г . дои : 10.2307/3570374 . JSTOR 3570374 . ПМИД 13167339 .
- ^ Jump up to: а б с Уортон, ДРА; Уортон, М.Л. (1959). «Влияние радиации на продолжительность жизни таракана Periplaneta americana в зависимости от дозы, возраста, пола и приема пищи». Радиационные исследования . 11 (4): 600–615. Бибкод : 1959РадР...11..600Вт . дои : 10.2307/3570814 . JSTOR 3570814 . ПМИД 13844254 .
- ^ Jump up to: а б с Хорикава Д.Д.; Сакашита Т; Катагири С; Ватанабэ М; и др. (2006). «Радиационная толерантность тихоходок Milnesium tardigradum». Международный журнал радиационной биологии . 82 (12): 843–8. дои : 10.1080/09553000600972956 . ПМИД 17178624 . S2CID 25354328 .
- ^ Хейденталь, Г (1945). «Возникновение доминантных летальных мутаций, вызванных рентгеновским излучением, у Хабробракона » . Генетика . 30 (2): 197–205. дои : 10.1093/генетика/30.2.197 . ПМК 1209282 . ПМИД 17247153 .
- ^ Jump up to: а б с д и ж Радиохимия и ядерная химия, Г. Чоппин, Дж.О. Лильензин и Дж. Ридберг, издание третье, стр. 481, ISBN 0-7506-7463-6
- ^ Jump up to: а б с д и «Тараканы и радиация» . Австралийская радиовещательная корпорация . 23 февраля 2006 г. Проверено 13 мая 2006 г.
- ^ «Заметки о радиации: радиационное повреждение и измерение дозы» . Проверено 16 июня 2018 г.
- ^ «Радиационные чрезвычайные ситуации CDC, острый радиационный синдром: информационный бюллетень для врачей» . Архивировано из оригинала 16 июля 2006 г.
- ^ Хартман, П; Гольдштейн, П; Альгарра, М; Хаббард, Д; Мабери, Дж (1996). «Нематода Caenorhabditis elegans в 39 раз более чувствительна к гамма-излучению, генерируемому 137Cs, чем 60Co». Мутат Рес . 363 (3): 201–208. дои : 10.1016/0921-8777(96)00012-2 . ПМИД 8765161 .
- ^ Вайдхас, Дж.Б.; Эйзенманн, DM; Голуб, Дж. М.; Наллур, СВ (2006). «Тканевая модель Caenorhabditis elegans радиационно-индуцированной гибели репродуктивных клеток» . Proc Natl Acad Sci США . 103 (26): 9946–51. Бибкод : 2006PNAS..103.9946W . дои : 10.1073/pnas.0603791103 . ПМК 1502559 . ПМИД 16788064 .
- ^ Jump up to: а б Криско, А.; Магали, Л.; Радман, М.; Мезельсон, М. (2012). «Чрезвычайная антиоксидантная защита бделлоидных коловраток от ионизирующего излучения» . Proc Natl Acad Sci США . 109 (7): 2354–2357. Бибкод : 2012PNAS..109.2354K . дои : 10.1073/pnas.1119762109 . ПМЦ 3289372 . ПМИД 22308443 .
Дальнейшее чтение
[ редактировать ]- Столяр, MC (1994). «Индуцированная радиорезистентность: обзор и историческая перспектива». Международный журнал радиационной биологии . 65 (1): 79–84. дои : 10.1080/09553009414550111 . ПМИД 7905914 .
- Клифтон Линг, К.; Эндлич, Б. (1989). «Радиорезистентность, вызванная онкогенной трансформацией». Радиационные исследования . 120 (2): 267–79. Бибкод : 1989РадР..120..267Л . дои : 10.2307/3577713 . JSTOR 3577713 . ПМИД 2694214 .
- Нотель, Х. (1987). «Адаптация популяций Drosophila melanogaster к высокому давлению мутаций: эволюционная корректировка скорости мутаций» . Труды Национальной академии наук . 84 (4): 1045–9. Бибкод : 1987PNAS...84.1045N . дои : 10.1073/pnas.84.4.1045 . ПМК 304358 . ПМИД 3103121 .
- Форнальски, К.В. (2016). «Радиация и эволюция: от уравнения Лотки-Вольтерра к уравнению баланса». Международный журнал низкой радиации . 10 (3): 222–33. дои : 10.1504/IJLR.2016.10002388 .
| Базы данных органов управления : Национальные |
|---|