Тригонотарбида
| Тригонотарбида Временной диапазон: Поздний силурий – ранняя пермь | |
|---|---|
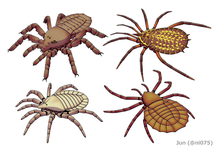 | |
| Palaeocharinus rhyniensis , Eophrynus prestvicii , Trigonotarbus johnsoni и Maiocercus celticus. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Подтип: | Хелицерата |
| Сорт: | Паукообразные |
| Заказ: | † Тригонотарбида Петрункевич , 1949 г. |
| Семьи | |
| Синонимы | |
| |
Отряд , Trigonotarbida — группа вымерших паукообразных которых летопись окаменелостей простирается от позднего силура до ранней перми ( от Придоли до Сакмария ). [1] [2] [3] Эти животные известны из нескольких местонахождений в Европе и Северной Америке , а также единичная находка из Аргентины . Тригонотарбидов можно рассматривать как паукообразных паукообразных производящих шелк , но без фильер, . Они имели размер от нескольких миллиметров до нескольких сантиметров в длину тела и имели сегментированное брюшко ( опистосома ) со спинным экзоскелетом ( тергитами ) на задней части брюшка животных, которые характерно были разделены на три или пять отдельных пластинок. [1] Вероятно, живя в качестве хищников на других членистоногих , некоторые более поздние виды тригонотарбидов имели довольно тяжелую броню и защищали себя шипами и бугорками. [4] В настоящее время известно около семидесяти видов, причем большинство окаменелостей происходит из угольных отложений каменноугольного периода .
Историческая справка
[ редактировать ]
Первый тригонотарбид был описан в 1837 году из угольных залежей Коулбрукдейла в Англии известным английским геологом Дином Уильямом Баклендом . [5] Он считал, что это ископаемый жук , и назвал его Curculoides prestvicii . Гораздо лучше сохранившийся экземпляр был позже обнаружен в Козли недалеко от Дадли ; также в английском агломерации Уэст-Мидлендс . Описан в 1871 году Генри Вудвордом . [6] он правильно идентифицировал его как паукообразного и переименовал его в Eophrynus prestvicii , в результате чего название рода происходит от ἠώς ( eos , что означает «рассвет») и Phrynus , рода живых пауков-хлыстов ( Amblypygi ). Впоследствии Вудворд описал еще один тригонотарбид, Brachypyge Carbonis , из угольных месторождений Монса в Бельгии ; [7] хотя это ископаемое известно только по его брюшку, и первоначально его приняли за окаменелость краба .
Новый отряд паукообразных
[ редактировать ]
В 1882 году немецкий зоолог Фердинанд Карш описал ряд ископаемых паукообразных из угольных залежей Нейроде в Силезии (ныне Польша ), в том числе одного, которого он назвал Anthracomartus voelkelianus в честь герра Фелькеля, мастера шахты, где он был обнаружен. [9] Этот вид был возведен в новый, вымерший отряд паукообразных, который Карш назвал Anthracomarti. Название происходит от ἄνθραξ ( сибирская язва ), греческого слова, обозначающего уголь . Примерно в это же время был обнаружен ряд других окаменелостей, которые в конечном итоге были помещены в Тригонотарбиду. Ханнс Бруно Гейниц описал Kreischeria wiedei из угольных запасов Цвикау в Германии . [10] хотя он интерпретировал его как ископаемый псевдоскорпион . Иоганн Кушта описал Anthracomartus krejcii. [11] из Раковника в Чехии и опубликовал дальнейшие описания в ряде последующих статей. [12] [13] [14] В 1884 году Сэмюэл Хаббард Скаддер описал Anthracomartus trilobitus из Фейетвилля, штат Арканзас — первого тригонотарбида из Северной Америки . [15]
Отношения
[ редактировать ]
| |||||||||||||||
| Внутренняя филогения Pantetrapulmonata , показывающая положение Trigonotarbita и возможную связь с Ricinulei . [16] [17] Вымершие таксоны обозначены знаком «†». |
Ранние исследования имели тенденцию путать тригонотарбидов с другими ныне живущими или вымершими группами паукообразных; особенно сборщики урожая ( Opiliones ). Разделение Петрункевичем тригонотарбидов на два, неродственных порядка, отмечалось выше. Подробно он разделил паукообразных на подотряды в зависимости от ширины разделения между двумя частями тела ( просомой и опистосомой ). Anthracomartida и другой вымерший отряд, Haptopoda , были сгруппированы в подкласс Stethostomata, характеризующийся широким разделением тела и свисающим вниз ротовым аппаратом. Тригонотарбида была помещена в отдельный подкласс Soluta и определена как имеющая деление тела переменной ширины. Схема Петрункевича во многом использовалась в последующих исследованиях ископаемых паукообразных.
Пантетрапульмоната
[ редактировать ]В 1980-е годы Билл Шир и его коллеги [18] провел важное исследование хорошо сохранившихся тригонотарбидов среднего девона из Гильбоа, штат Нью-Йорк . Они задались вопросом, уместно ли определять группу животных по изменчивому состоянию признаков, и провели первый кладистический анализ ископаемых и живых паукообразных. Они показали, что тригонотарбиды тесно связаны с группой паукообразных, ходивших под разными названиями (Caulogastra, Arachnidea и др. название Tetrapulmonata . ), но для которых наибольшее распространение получило [19] К представителям Tetrapulmonata относятся пауки ( Araneae ), пауки-клыки ( Amblypygi ), скорпионы-клыки ( Uropygi ) и короткохвостые скорпионы ( Schizomida ) и вместе с тригонотарбидами имеют общие признаки, подобные двум парам книжных легких и схожим ротовым аппаратам с клыками, работающими скорее как карманный нож. [20] В исследовании взаимоотношений паукообразных, проведенном в 2007 году, Shear et al. Гипотеза была в значительной степени поддержана, и была предложена группа Pantetrapulmonata , включающая Trigonotarbida + Tetrapulmonata. [21] С тех пор это было подтверждено в более поздних кладистических анализах. [22] [23] [24] [25]
Тригонотарбиды и рицинулеиды
[ редактировать ]

В 1892 году Фердинанд Карш предположил, что редкие и довольно причудливого вида рицинулеиды ( Ricinulei ) были последними живыми потомками тригонотарбидов. [26] Аналогичная гипотеза была вновь выдвинута Данлопом. [16] которые указали на явные сходства и возможные сестринские отношения между этими группами паукообразных. Оба имеют опистосомальные тергиты, разделенные на срединную и латеральную пластинки, и оба имеют сложный механизм соединения между просомой и опистосомой, который «скрепляет» две половины тела вместе. Хотя кладистический анализ имел тенденцию обнаруживать рицинулеиды в их традиционном положении, тесно связанном с клещами и клещами, дальнейшие открытия показали, что кончик педипальпы заканчивается небольшим когтем как у тригонотарбидов, так и у рицинулеид. [17] [27] Если гипотеза верна, рицинулеиды, несмотря на отсутствие ключевых признаков тетрапульмоната (например, книжные легкие), могут также представлять часть пантетрапульмонатной клады наряду с тригонотарбидами. [16] [17] [20]
Внутренние отношения
[ редактировать ]Первый кладистический анализ тригонотарбидов был опубликован в 2014 году. [8] Это восстановило семейства Anthracomartidae , Anthracosironidae и Eophrynidae как монофилетические. Напротив, Trigonotarbidae , Aphantomartidae , Palaeocharinidae и Kreischeriidae не были таковыми. Две клады были последовательно восстановлены при сильной поддержке - ( Palaeocharinus (Archaeomartidae + Anthracomartidae)) и Lissomartus как сестринская группа «ассоциация эофринид» ( Aphantomartus ( Alkenia ( Pseudokreischeria ( Kreischeria ( Eophrynus + Pleophrynus )))))).
Описание
[ редактировать ]

Тригонотарбиды внешне напоминают пауков , но их легко узнать по тергитам на дорсальной стороне опистосомы, разделенным на срединную и латеральную пластинки. [28] Этот признак является общим с рицинулеидами ( Ricinulei ) (см. также Ricinulei#Relationships ). Как и у других паукообразных, тело разделено на просому (или головогрудь) и опистосому (или брюшко). Длина тела колеблется от пары миллиметров до примерно 5 см (2,0 дюйма). [29]
Просома
[ редактировать ]Просома покрыта панцирем и всегда имеет пару срединных глаз. [1] В вероятно базальных семействах Palaeocharinidae , Anthracomartidae [30] — и, возможно, также Anthracosironidae — имеется дополнительная пара боковых глазных бугорков, которые, по крайней мере у палеохаринид, [31] похоже, имел ряд отдельных линз. В этом смысле палеохариниды, по-видимому, находятся в процессе уменьшения сложного глаза . [32] Передний край панциря выдается в выступ, называемый наличником. [22]
Хелицеры . типа «карманного ножа», состоящие из базального сегмента и острого изогнутого клыка [1] Хелицеры описываются как палеогнатические : клыки расположены параллельно друг другу, как у мезотелиальных и мигаломорфных пауков, но хелицеры свисают вниз, как у аранеоморфных пауков. [33] В хорошо сохранившихся окаменелостях нет доказательств открытия ядовитой железы, поэтому тригонотарбиды, вероятно, не были ядовитыми . Хелицеры могли быть слегка втянутыми в просому. У хорошо сохранившихся палеохаринид имеется небольшой щелевидный рот с верхней губой ( губой или рострумом) и нижней губой (или губой). [34] Внутри рта имеется своего рода фильтрующая система, образованная волосками или тромбоцитами, что убедительно свидетельствует о том, что тригонотарбиды (подобно паукам и многим другим паукообразным) могли питаться только предварительно переваренной, разжиженной добычей. [34]
Педипальпы . имеют типичное паукообразное строение с тазиком, вертелом, бедренной костью, надколенником, голенью и предплюсной Они ножкообразные , то есть выглядят как маленькие ножки и не сильно видоизменились. [35] Нет никаких доказательств существования специального устройства для переноса спермы, как в модифицированном пальпальном органе самцов пауков. По крайней мере, у палеохаринид и антракомартид кончик педипальпы видоизменился в небольшую челу (коготь), образованную из тарзального когтя (или апотеле) и выступа из предплюсны. Как упоминалось выше, очень похожее расположение наблюдается на конце педипальпы у Ricinulei . [17] [27]
Ходячие ноги снова следуют типичному плану паукообразных с тазиком, вертелом, бедренной костью, надколенником, большой берцовой костью, плюсной и предплюсной. [1] Тазики окружают одну грудину. У хорошо сохранившихся палеохаринид вокруг вертел-бедренного сустава имеется кольцо, или кольцо, которое может быть остатками более раннего сегмента ноги. [35] Ноги практически не изменены, хотя у Anthracosironidae передние ноги довольно крупные и колючие. [36] предположительно, чтобы помочь поймать добычу. Ноги заканчиваются тремя коготками: двумя большими и срединным коготком поменьше. [35]
Опистосома
[ редактировать ]

Опистосома в основном субовальной формы с уплощенной дорсальной поверхностью. [35] Он состоит из 12 сегментов, некоторые из которых подверглись степени слияния или редукции, отсюда и предыдущая неправильная интерпретация примерно от 8 до 11 сегментов. [20] Тергит первого сегмента частично прикрыт задним краем предыдущего панциря, образуя сложный соединительный механизм, известный как «запирающий гребень». [1] [20] Тергиты сегментов 2–8 (у некоторых видов сегмент 9) были разделены по бокам на 3 (одну срединную и две латеральные) пластинки, причем у большинства видов тергиты сегментов 2 и 3 срослись друг с другом. [20] Однако соответствующие тергиты семейства Anthracomartidae подразделяются еще на 5 пластинок. [30] Последние три сегмента обычно видны только с брюшной стороны. [1] два последних сегмента сужены в крошечную кольцеобразную часть, известную как пигидий. [20]
Вентральная сторона 2-9 сегмента опистосомы покрыта рядом легочных крышек (2 и 3) и изогнутыми стернитами (4-9). [1] [20] В первом сегменте, по-видимому, отсутствуют вентральные пластинки. Так же, как и у других легочных паукообразных ( скорпионов и тетрапульмонатов ), книжные легкие тригонотарбид образованы слоями трабекулонесущих пластинок, что является особенностью, приспособленной к наземному , воздушно-дышащему образу жизни. [37] У некоторых видов наблюдалась пара вентральных мешков, расположенных между задней покрышкой и следующим за ней стернитом. [38] [4] [20]
Палеобиология
[ редактировать ]В июле 2014 года ученые использовали компьютерные методы, чтобы воссоздать возможную походку животного. [39] [35] В последующей обзорной статье было предположено, что по сравнению с клещами, предположительно имеющими схожий образ жизни и окружающую среду, более вероятна метахронная, а не попеременная координация ног. [40] Последующая работа исследователей, стоящих за первой публикацией [35] использовали подходы моделирования для оценки эффективности различных походок с использованием обновленной тригонотарбидной модели. [41]
Включенные таксоны
[ редактировать ]70 действующих видов , а именно: По состоянию на 2020 год в Trigonotarbida было включено [3]
- Палеотарбус Данлоп, 1999 г.
- Палеотарбус с соломенной крышей (Данлоп, 1996) - поздний силур, Англия.
- Палеохариниды Херст, 1923 г.
- Aculeatarbus Shear, Selden & Rolfe, 1987 г.
- Aculeatarbus depressus Shear, Selden & Rolfe, 1987 – средний девон, США.
- Геласинотарбус Сдвиг, Selden & Rolfe, 1987 г.
- Gelasinotarbus bifidus Shear, Selden & Rolfe, 1987 – средний девон, США.
- Gelasinotarbus bonamoae Shear, Selden & Rolfe, 1987 – средний девон, США.
- Gelasinotarbus heptops Shear, Selden & Rolfe, 1987 – средний девон, США.
- Gelasinotarbus reticulatus Shear, Selden & Rolfe, 1987 – средний девон, США.
- Гигантохаринус Шир, 2000 г.
- Gigantocharinus szatmaryi Shear, 2000 – поздний девон, США.
- Гилбоарахне Шир, Селден и Рольф, 1987 г.
- Gilboarachne griersoni Shear, Selden & Rolfe, 1987 – средний девон, США.
- Палеохаринус Херст, 1923 г.
- Palaeocharinus Calmani Hirst, 1923 - ранний девон, Шотландия.
- Palaeocharinus Hornei Hirst, 1923 - ранний девон, Шотландия.
- Palaeocharinus kidstoni Hirst, 1923 - ранний девон, Шотландия.
- Palaeocharinus rhyniensis Hirst, 1923 - ранний девон, Шотландия.
- Palaeocharinus scourfieldi Hirst, 1923 - ранний девон, Шотландия.
- Palaeocharinus tuberculatus Fayers, Dunlop & Trewin, 2005 – ранний девон, Шотландия.
- Спинохаринус Пошманн и Данлоп, 2011 г.
- Spinocharinus steinmeyeri Poschman & Dunlop, 2011 - девон, Бюрденбах
- Archaeomartidae Haase, 1890 г.
- Археомартус Стёрмер, 1970 г.
- Archaeomartus levis Størmer, 1970 - девон, Алькен-ан-дер-Мозель
- Антракомартиды Хаазе, 1890 г.
- синонимы
- = Промигалиды Фрич, 1904 г.
- = Brachypygidae Pocock, 1911 г.
- = Корифомартиды Петрункевич, 1945
- = Плеомартиды Петрункевич, 1945
- Антракомартус Карш, 1882 г.
- синонимы
- = Брахиликоза Фрич, 1904 г.
- = Клептомартус Петрункевич, 1949 г.
- = Корифомарт Петрункевич, 1945 г.
- = Криптомартус Петрункевич, 1945 г.
- = Оомартус Петрункевич, 1953 г.
- = Пернерия Фрич, 1904 г.
- = Плеомарт Петрункевич, 1945 г.
- = Промигале Фрич, 1901 г.
- Anthracomartus bohemica (Frič, 1901) - поздний карбон, Чехия.
- Anthracomartus carcinoides (Frič, 1901) - поздний карбон, Чехия.
- синонимы
- = Промигальская ротонда Фрич, 1901 г.
- = Perneria salticoides Frič, 1904 г.
- синонимы
- Anthracomartus elegans Frič, 1901 – поздний карбон, Чехия.
- Anthracomartus hidi Pocock, 1911 – поздний карбон, Англия.
- синонимы
- = Клептомартус хангарди Гутерль, 1965 г.
- = Криптомартус Мейери Гутерль, 1964 г.
- = Cleptomartus planus Петрункевич, 1949 г.
- = Cryptomartus rebskei Brauckmann, 1984 г.
- синонимы
- Anthracomartus granulatus Frič, 1904 - поздний карбон, Польша.
- Anthracomartus janae (Opluštil, 1986) - поздний карбон, Чехия.
- Anthracomartus kustae Petrunkevitch, 1953 – поздний карбон, Чехия.
- Anthracomartus major Kušta, 1884 - поздний карбон, Чехия.
- синоним
- = Anthracomartus socius Kušta, 1888 г.
- синоним
- Anthracomartus nyranensis (Петрункевич, 1953) - поздний карбон, Чехия.
- Anthracomartus palatinus Ammon, 1901 – поздний карбон, Германия.
- Anthracomartus pristi Pocock, 1911 – поздний карбон, Англия.
- синонимы
- = Anthracomartus denuiti Pruvost, 1922 г.
- = Cleptomartus plautus Петрункевич, 1949 г.
- синонимы
- Anthracomartus radvanicensis (Opluštil, 1985) - поздний карбон, Чехия.
- Anthracomartus triangularis Petrunkevitch, 1913 – поздний карбон, Канада.
- Anthracomartus trilobitus Scudder, 1884 - поздний карбон, США.
- Anthracomartus voelkelianus Karsch, 1882 - поздний карбон, Польша.
- синонимы
- Брахипиг Вудворд, 1878 г.
- Brachypyge Carbonis Woodward, 1878 - поздний карбон, Бельгия.
- Майоцерк Покок, 1911 год.
- Maiocercus celticus (Pocock, 1902) - поздний карбон, Европа.
- синоним
- = Maiocercus orbcularis Gill, 1911 г.
- синоним
- Maiocercus celticus (Pocock, 1902) - поздний карбон, Европа.
- Антракосирониды Покок, 1903 г.
- Антракосиро Покок, 1903 год.
- Anthracosiro fritschii Pocock, 1903 – поздний карбон, Европа.
- синоним
- = Anthracosiro elongatus Waterlot, 1934 г.
- синоним
- Anthracosiro woodwardi Pocock, 1903 – поздний карбон, Европа.
- синонимы
- = Антракосиро Корсини Прувост, 1926 г.
- = Anthracosiro latipes Gill, 1909 г.
- синонимы
- Anthracosiro fritschii Pocock, 1903 – поздний карбон, Европа.
- Арианрода Данлоп и Селден, 2004 г.
- Арианрода Беннетти Данлоп и Селден, 2004 г. - ранний девон, Уэльс.
- Вратиславия Фрич, 1904 г.
- Vratislavia silesica (Roemer, 1878) - Каменноугольный период, Силезия.
- Тригонотарбиды Петрункевича, 1949 г.
- Тригонотарбус Покок, 1911 год.
- Trigonotarbus arnoldi Petrunkevitch, 1955 – поздний карбон, Франция.
- Trigonotarbus johnsoni Pocock, 1911 – поздний карбон, Англия.
- Trigonotarbus stoermeri Schultka, 1991 – ранний девон, Германия.
- Лиссомартиды Данлоп, 1995 г.
- Лиссомартус Петрункевич, 1949 г.
- Lissomartus Carbonarius (Петрункевич, 1913) - поздний карбон, США.
- Lissomartus schucherti (Петрункевич, 1913) - поздний карбон, США.
- Афантомартиды Петрункевич, 1945 г.
- синоним
- = Тригономартиды Петрункевич, 1949
- Алкения Стёрмер, 1970 г.
- Alkenia mirabilis Størmer, 1970 - девон, Алкен на Мозеле.
- Афантомартус Покок, 1911 год.
- синонимы
- = Тригономарт Петрункевич, 1913 г.
- = Фриномарт Петрункевич, 1945а
- Aphantomartus areolatus Pocock, 1911 – ранний/поздний карбон, Европа.
- синонимы
- = Aphantomartus pococki Pruvost, 1912 г.
- = Trigonomartus dorlodoti Pruvost, 1930
- = Eophrynus waechteri Guthörl, 1938 г.
- = ? Trigonomartus pruvosti van der Heide, 1951 г.
- = ? Brachylycosa manebachensis Müller, 1957 г.
- синонимы
- Aphantomartus ilfeldicus (Scharf, 1924) - Пермь, Германия.
- Aphantomartus pustulatus (Scudder, 1884) - поздний карбон, Европа, Северная Америка.
- синонимы
- = ? Kreischeria villeti Pruvost, 1912 г.
- = Cleptomartus plötzensis Саймон, 1971 г.
- синонимы
- синонимы
- Kreischeriidae Haase, 1890 г.
- Анзиния Петрункевич, 1953 год.
- Anzinia thevenini (Pruvost, 1919) - поздний карбон, Франция.
- Гондванараче Пинто и Хюникен, 1980 г.
- Gondwanarache argentinensis Pinto & Hünicken, 1980 – поздний карбон, Аргентина.
- Гемикрайшерия Фрич, 1904 г.
- Hemikreischeria geinitzi (Thevenin, 1902) - поздний карбон, Франция.
- Крайшерия Гейниц, 1882 г.
- Kreischeria wiedei Geinitz, 1882 - поздний карбон, Германия.
- Псевдокрейшерия Петрункевич, 1953 г.
- Pseudokreischeria pococki (Gill, 1924) - поздний карбон, Англия.
- синоним
- = Eophrynus varius Петрункевич, 1949 г.
- синоним
- Pseudokreischeria pococki (Gill, 1924) - поздний карбон, Англия.
- Эофриниды Карш, 1882 г.
- синоним
- = Hemiphrynidae Frič, 1904
- Эофрин Вудворд, 1871 г.
- Eophrynus prestvicii (Бакленд, 1837) - поздний карбон, Англия.
- Eophrynus udus Brauckmann, Koch & Kemper, 1985 – поздний карбон, Германия.
- Нюранитарбус Харви и Селден, 1995 г.
- синоним
- Гемифрин Фрич, 1901 г.
- Nyranytarbus hofmanni (Фрич, 1901) - поздний карбон, Чехия.
- Nyranytarbus longipes (Frič, 1901) - поздний карбон, Чехия.
- синоним
- Петровича Фрич, 1904 г.
- Petrovicia proditoria Frič, 1904 - поздний карбон, Чехия.
- Планомартус Петрункевич, 1953 г.
- Planomartus krejcii (Kušta, 1883) - поздний карбон, Чехия.
- синоним
- = Anthracomartus affinis Kušta, 1885 г.
- синоним
- Planomartus krejcii (Kušta, 1883) - поздний карбон, Чехия.
- Плеофрин Петрункевич, 1945а
- Pleophrynus verrucosus (Pocock, 1911) - поздний карбон, Великобритания, США.
- синоним
- = Eophrynus warei Dix & Pringle, 1930 г.
- = Pleophrynus ensifer Petrunkevitch, 1945a
- = Eophrynus jugatus Ambrose & Romano, 1972.
- синоним
- Pleophrynus verrucosus (Pocock, 1911) - поздний карбон, Великобритания, США.
- Поконония Петрункевич, 1953 г.
- Pocononia whitei (Ewing, 1930) - ранний карбон, США.
- Сомаспидион Джукс, 1982 г.
- Сомаспидион хаммаферон Юкс, 1982 г.
- Стенотрогул Фрич, 1904 г.
- синонимы
- = Циклотрогул Фрич, 1904 г.
- = Псевдоэофрин Пршибыл, 1958
- Stenotrogulus salmii (Stur, 1877) - поздний карбон, Чехия.
- синонимы
- = Cyclotrogulus sturii Frič, 1904 [нон Хассе, 1890]
- = Pseudoeophrynus ostraviensis Příbyl, 1958
- синонимы
- синонимы
- Семья неопределенная
- Пошманн , Данлоп, Берту и Галтье, 2016 г.
- Aenigmatarbus rastelli Poschmann, Dunlop, Bértoux & Galtier, 2016 - Каменноугольный период, Грассессак, Франция
- Намуротарбус Пошманн и Данлоп, 2010 г.
- Namurotarbus roessleri (Dunlop & Brauckmann, 2006) - каменноугольный период, Хаген-Ворхалле
- синонимы
- = Archaeomartus roessleri Dunlop & Brauckmann, 2006.
- синонимы
- Namurotarbus roessleri (Dunlop & Brauckmann, 2006) - каменноугольный период, Хаген-Ворхалле
- Пермотарбус Данлоп и Рёсслер, 2013 г.
- Permotarbus schuberti Dunlop & Rößler, 2013 Пермь, Хемниц
- Тынекотарбус Градска и Данлоп, 2013 г.
- Tynecotarbus tichaveki Hradská & Dunlop, 2013 - Каменноугольный период, Тынец
- Антракофрин Андре, 1913 г.
- Anthracophrynus Tuberculatus Andrée, 1913 – поздний карбон, Германия.
- Ареомарт Петрункевич, 1913 г.
- Areomartus ovatus Petrunkevitch, 1913 - Каменноугольный период, Западная Вирджиния.
- ' Eophrynus ' scharfi Scharf, 1924 – ранняя пермь, Германия.
- Афантомартус Покок, 1911 год.
- Aphantomartus woodruffi (Scudder, 1893) - каменноугольный период, Род-Айленд.
- Anthracomartus buchi (Goldenberg, 1873) - поздний карбон, Германия.
- Anthracomartus hageni (Goldenberg, 1873) - поздний карбон, Германия.
- Elaverimartus pococki Petrunkevitch, 1953 – поздний карбон, Шотландия.
- Eurymartus latus Matthew, 1895 - поздний карбон, Канада.
- ?Eurymartus spinulosus Matthew, 1895 – поздний карбон, Канада.
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час Гарвуд, Рассел Дж.; Данлоп, Джейсон А. (2010). «Объяснение окаменелостей: тригонотарбиды» . Геология сегодня . 26 (1): 34–37. дои : 10.1111/j.1365-2451.2010.00742.x . S2CID 247708509 . Проверено 12 июня 2015 г.
- ^ Данлоп, Дж.А.; Рёсслер, Р. (1 августа 2013 г.). «Самый молодой тригонотарбид Permotarbus schuberti n. gen., n. sp. из пермского окаменелого леса Хемница в Германии» . Ископаемый рекорд . 16 (2): 229–243. Бибкод : 2013FossR..16..229D . doi : 10.5194/fr-16-229-2013 . ISSN 2193-0074 .
- ^ Jump up to: а б Данлоп, Дж. А., Пенни, Д. и Джекел, Д. 2020. Сводный список ископаемых пауков и их родственников . Во Всемирном каталоге пауков. Музей естественной истории Берна, онлайн по адресу http://wsc.nmbe.ch , версия 20.5.
- ^ Jump up to: а б Данлоп, Джейсон; Гарвуд, Рассел (2014). «Томографическая реконструкция исключительно сохранившегося тригонотарбидного паукообразного Eophrynus prestvicii» . Acta Palaeontologica Polonica . дои : 10.4202/app.2012.0032 .
- ^ Уильям Бакленд (1837). Трактат IV. Геология и минералогия со ссылкой на естественное богословие . Трактаты Бриджуотера посвящены силе, мудрости и благости Бога, проявленной в творении. (2-е изд.). Лондон: Уильям Пикеринг .
- ^ Х. Вудворд (1871). «Об открытии нового и совершенного паукообразного из железного камня угольного месторождения Дадли» . Геологический журнал . 8 (9): 1–4. Бибкод : 1871GeoM....8..385W . дои : 10.1017/s0016756800192817 . S2CID 128850722 .
- ^ Х. Вудворд (1878 г.). «Обнаружение останков ископаемого краба (Decapoda-Bracyura) в угольных залежах окрестностей Монса, Бельгия» . Геологический журнал . новая серия. 2 (5): 433–436. дои : 10.1017/S0016756800152616 . S2CID 129112886 .
- ^ Jump up to: а б Джонс, Фиона; Данлоп, Джейсон А.; Фридман, Мэтью; Гарвуд, Рассел Дж. (2014). « Trigonotarbus johnsoni Pocock, 1911, выявленный с помощью рентгеновской компьютерной томографии с кладистическим анализом вымерших тригонотарбидных паукообразных». Зоологический журнал Линнеевского общества . 172 (1): 49–70. дои : 10.1111/zoj.12167 .
- ^ Ф. Карш (1882). «О новом паукообразном из Силезского угля и паукообразных вообще». Журнал Немецкого геологического общества (на немецком языке). 34 :556-561.
- ^ Х. Б. Гейниц (1882). « Kreischeria wiedei , псевдоскорпион из угольной формации Цвикау». Журнал Немецкого геологического общества (на немецком языке). 34 :238-242.
- ^ Иоганн Кушта (1883). « Anthracomartus krejcii , новый паукообразный из чешского карбона». Отчеты о заседаниях Королевского чешского общества наук, математики и естественных наук (на немецком языке). 1883 : 7.
- ^ Иоганн Кушта (1884). «Новые паукообразные из угольной формации Ракониц». Отчеты о заседаниях Королевского чешского общества наук, математики и естественных наук (на немецком языке). 1884 : 398–401.
- ^ Иоганн Кушта (1885). «Новые ископаемые членистоногие из сланца Ноеггаратиен в Раконице». Отчеты о заседаниях Королевского чешского общества наук, математики и естественных наук (на немецком языке). 1885 : 1–7.
- ^ Иоганн Кушта (1888). «O nových arachnidech z karbonu Rkovnického. (Новые паукообразные из угольных пластов близ Раконица)». Труды Королевского Богемского общества наук, математики и естественных наук (на чешском языке). 1888 : 194–208.
- ^ Сэмюэл Х. Скаддер (1884). «Вклад в наши знания о палеозойских паукообразных». Труды Американской академии искусств и наук . 20 :13–22. дои : 10.2307/25138764 . hdl : 2027/hvd.32044107176828 . JSTOR 25138764 .
- ^ Jump up to: а б с Джейсон А. Данлоп (1996). «Доказательства родственных отношений между Рицинулеем и Тригонотарбидой» (PDF) . Бюллетень Британского арахнологического общества . 10 (6): 193–204.
- ^ Jump up to: а б с д Джейсон А. Данлоп, Карстен Каменц и Джованни Таларико (2009). «Ископаемое тригонотарбидное паукообразное с педипальной когтем, напоминающим рицинулейд». Зооморфология . 128 (4): 305–313. дои : 10.1007/s00435-009-0090-z . S2CID 6769463 .
- ^ Уильям А. Шир, Пол А. Селден, WDI Рольф, Патрисия М. Бонамо и Джеймс Д. Грирсон (1987). «Новые наземные паукообразные из девона Гильбоа, Нью-Йорк». Американский музей Novitates (2901): 1–74. hdl : 2246/5166 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Джеффри В. Шульц (1990). «Эволюционная морфология и филогения паукообразных» . Кладистика . 6 (1): 1–38. дои : 10.1111/j.1096-0031.1990.tb00523.x . ПМИД 34933471 . S2CID 85410687 .
- ^ Jump up to: а б с д и ж г час Данлоп, Джейсон А.; Ламсделл, Джеймс К. (2017). «Сегментация и тагмос у Chelicerata» . Строение и развитие членистоногих . 46 (3): 395–418. дои : 10.1016/j.asd.2016.05.002 . ISSN 1467-8039 . ПМИД 27240897 .
- ^ Джеффри В. Шульц (2007). «Филогенетический анализ отрядов паукообразных на основе морфологических признаков» . Зоологический журнал Линнеевского общества . 150 (2): 221–265. дои : 10.1111/j.1096-3642.2007.00284.x .
- ^ Jump up to: а б Гарвуд, Рассел Дж.; Данлоп, Джейсон А. (2014). «Трёхмерная реконструкция и филогения вымерших отрядов хелицеровых» . ПерДж . 2 : е641. дои : 10.7717/peerj.641 . ПМЦ 4232842 . ПМИД 25405073 .
- ^ Гарвуд, Рассел Дж.; Данлоп, Джейсон А.; Селден, Пол А.; Спенсер, Алан RT; Этвуд, Роберт С.; Во, Нгиа Т.; Дракопулос, Майкл (2016). «Почти паук: ископаемые паукообразные возрастом 305 миллионов лет и происхождение пауков» . Труды Королевского общества B: Биологические науки . 283 (1827): 20160125. doi : 10.1098/rspb.2016.0125 . ISSN 0962-8452 . ПМЦ 4822468 . ПМИД 27030415 .
- ^ Ван, Бо; Данлоп, Джейсон А.; Селден, Пол А.; Гарвуд, Рассел Дж.; Шир, Уильям А.; Мюллер, Патрик; Лей, Сяоцзе (2018). «Меловой паукообразный Chimerarachne yingi gen. et sp. nov. проливает свет на происхождение пауков» . Экология и эволюция природы . 2 (4): 614–622. Бибкод : 2018NatEE...2..614W . дои : 10.1038/s41559-017-0449-3 . ISSN 2397-334X . ПМИД 29403075 . S2CID 4239867 .
- ^ Гарвуд, Рассел Дж.; Данлоп, Джейсон А.; Кнехт, Брайан Дж.; Хегна, Томас А. (2017). «Филогения ископаемых пауков-хлыстов» . Эволюционная биология BMC . 17 (1): 105. Бибкод : 2017BMCEE..17..105G . дои : 10.1186/s12862-017-0931-1 . ISSN 1471-2148 . ПМЦ 5399839 . ПМИД 28431496 .
- ^ Фердинанд Карш (1892). «О Cryptostemma Guèr. как о единственном недавнем продолжении ископаемого отряда паукообразных Meridogastra Thor» . Берлинский энтомологический журнал (на немецком языке). 37 (1): 25–32. дои : 10.1002/mmnd.18920370108 .
- ^ Jump up to: а б Рассел Гарвуд, Джейсон А. Данлоп и Марк Д. Саттон (2009). «Высокоточная рентгеновская микротомография-реконструкция паукообразных каменноугольных паукообразных, обитающих в сидерите» . Письма по биологии . 5 (6): 841–844. дои : 10.1098/rsbl.2009.0464 . ПМК 2828000 . ПМИД 19656861 .
- ^ Стивен Р. Файерс, Джейсон А. Данлоп и Найджел Х. Тревин (2005). «Новый раннедевонский тригонотарбидный паукообразный из кремня Виндифилд, Райни, Шотландия» . Журнал систематической палеонтологии . 2 (4): 269–284. Бибкод : 2005JSPal...2..269F . дои : 10.1017/S147720190400149X . S2CID 140706387 .
- ^ Ронни Рёсслер и Джейсон А. Данлоп (1997). «Переописание крупнейшего паукообразного тригонотарбида - Kreischeria wiedei Geinitz 1882 из верхнего карбона Цвикау, Германия». Палеонтологический журнал . 71 (3–4): 237–245. Бибкод : 1997PalZ...71..237R . дои : 10.1007/BF02988493 . S2CID 129447249 .
- ^ Jump up to: а б Гарвуд, Рассел Дж.; Данлоп, Джейсон А. (2011). «Морфология и систематика антракомартид (Arachnida: Trigonotarbida)» . Палеонтология . 54 (1): 145–161. Бибкод : 2011Palgy..54..145G . дои : 10.1111/j.1475-4983.2010.01000.x . ISSN 0031-0239 . S2CID 83897592 .
- ^ Стэнли Херст и С. Маулик (1926). «Останки некоторых членистоногих из Райни Черт (Старый красный песчаник)». Геологический журнал . 63 (2): 69–71. Бибкод : 1926GeoM...63...69H . дои : 10.1017/S0016756800083692 . S2CID 129777833 .
- ^ Мизер, Себастьян Т.; Данлоп, Джейсон А. (2016). «Боковая эволюция глаз у паукообразных» . Арахнология . 17 (2): 103–119. дои : 10.13156/арак.2006.17.2.103 . ISSN 2050-9928 . S2CID 89428386 .
- ^ Данлоп, Дж. А. (1997). «Палеозойские паукообразные и их значение для филогении паукообразных» (PDF) . Материалы 16-го Европейского коллоквиума по арахнологии . Седльце: Издательство Wyższa Szkoła Rolniczo-Pedagogiczna. стр. 65–82 . Проверено 19 марта 2016 г.
- ^ Jump up to: а б Джейсон А. Данлоп (1994). «Сравнительная анатомия механизмов фильтрации у четверолегочных паукообразных (Trigonotarbida, Araneae, Amblypygi, Uropygi и Schizomida)». Бюллетень Британского арахнологического общества . 9 : 267–273.
- ^ Jump up to: а б с д и ж Гарвуд, Рассел Дж.; Данлоп, Джейсон А. (июль 2014 г.). «Ходячие мертвецы: Блендер как инструмент для палеонтологов с тематическим исследованием вымерших паукообразных» . Журнал палеонтологии . 88 (4): 735–746. Бибкод : 2014JPal...88..735G . дои : 10.1666/13-088 . ISSN 0022-3360 . S2CID 131202472 . Проверено 21 июля 2015 г.
- ^ Р. И. Покок (1903). «Новый каменноугольный паукообразный» . Геологический журнал . Десятилетие 4. 10 (6): 247–251. Бибкод : 1903GeoM...10..247P . дои : 10.1017/S001675680011252X . S2CID 128408478 .
- ^ Каменц, Карстен; Данлоп, Джейсон А; Шольц, Герхард; Керп, Ганс; Хасс, Хаген (23 апреля 2008 г.). «Микроанатомия книжных легких раннего девона» . Письма по биологии . 4 (2): 212–215. дои : 10.1098/rsbl.2007.0597 . ISSN 1744-9561 . ПМЦ 2429929 . ПМИД 18198139 .
- ^ Файерс, Стивен Р.; Данлоп, Джейсон А.; Тревин, Найджел Х. (2005). «Новый раннедевонский паукообразный тригонотарбид из Виндифилд Черт, Райни, Шотландия» (PDF) . Журнал систематической палеонтологии . 2 (4): 269–284. Бибкод : 2005JSPal...2..269F . дои : 10.1017/S147720190400149X . ISSN 1477-2019 . S2CID 140706387 .
- ^ Джонатан Амос (9 июля 2014 г.). «Древний паукообразный «снова ходит» » . Новости Би-би-си . Проверено 9 июля 2014 г.
- ^ Вейманн, Том; Гетцке, Ханнс Хаген; Гюнтер, Михаэль (ноябрь 2015 г.). «Требования и ограничения прогнозов передвижения наземных членистоногих на основе анатомии с упором на паукообразных». Журнал палеонтологии . 89 (6): 980–990. Бибкод : 2015JPal...89..980W . дои : 10.1017/jpa.2016.33 . S2CID 86908527 .
- ^ МакЛин, Каллум. (2017). Моделирование походки вымершего тригонотарбидного паукообразного (MPhil). Университет Манчестера.
- Тригонотарбиды
- Арахнидские приказы
- Доисторические отряды членистоногих
- Силурийские паукообразные
- Девонские паукообразные
- Каменноугольные паукообразные
- Пермские паукообразные
- Девонские членистоногие Северной Америки
- Каменноугольные членистоногие Северной Америки.
- Пермские членистоногие Северной Америки
- Каменноугольные членистоногие Европы
- Девонские членистоногие Европы
- Пермские членистоногие Европы
- Силурийские членистоногие Европы
- Первые появления силурианцев
- Приуральские вымирания
- Таксоны имени Александра Петрункевича