Онихофора
| Онихофора Временной диапазон: стволовая группа из кембрия | |
|---|---|
 | |
| Ороперипатус сп. (Перипатиды) | |
 | |
| Opera patellus sp. (Перипатопсиды) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Subkingdom: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | билатерия |
| Клэйд : | Нефрозоа |
| (без рейтинга): | протостомия |
| Супертип: | Экдисозоа |
| (без рейтинга): | панартроподы |
| Тип: | Онихофора Толстый , 1850 год. |
| Подгруппы | |
| |
 | |
| Глобальный ареал онихофоры существуют Перипатиды окаменелости | |
Онихофоры / ɒ n ɪ ˈ k ɒ f ə r ə / (от древнегреческого : ονυχής , onyches , «когти»; и φέρειν , pherein , «нести»), широко известные как бархатные черви (из-за их бархатистой текстуры и несколько червеобразный вид) или более двусмысленно как перипат / pə ˈ r ɪ ( p ə t ə s / по первому описанному роду, Peripatus ), — тип удлиненных, мягкотелых, многоногих животных . [1] [2] По внешнему виду их сравнивали с червями на ногах, гусеницами и слизняками. [3] Они охотятся на других беспозвоночных, которых ловят, выбрасывая липкую слизь. Описано около 200 видов бархатных червей, хотя истинное количество видов, вероятно, больше. Два современных семейства бархатных червей — Peripatidae и Peripatopsidae . Они демонстрируют своеобразное распространение: перипатиды преимущественно экваториальные и тропические, тогда как все перипатопсиды встречаются к югу от экватора. Это единственный тип Animalia , полностью эндемичный для наземной среды обитания, по крайней мере, среди ныне живущих представителей. [4] [5] Бархатных червей обычно считают близкими родственниками Arthropoda и Tardigrada , вместе с которыми они образуют предполагаемый таксон Panarthropoda . [6] Это делает их интересными для палеонтологии, поскольку они могут помочь реконструировать предков членистоногих. Лишь два ископаемых вида уверенно относят к онихофорам: Antennipatus из позднего карбона и Cretoperipatus из позднего мела , причем последний принадлежит к Peripatidae. [5] [7] В современной зоологии они особенно известны своим любопытным брачным поведением и вынашиванием живых детенышей у некоторых видов.
Анатомия
[ редактировать ]Бархатные черви — это сегментированные животные с уплощенным цилиндрическим поперечным сечением тела и рядами неструктурированных придатков тела , известные как онкоподы или лобоподы (неофициально: короткие ноги). Они достигают длины от 0,1 до 22 см (0,04–8,66 дюйма) в зависимости от вида, самый маленький из известных — Ooperipatellus nanus , а самый крупный — Mongeperipatus solorzanoi . [8] [9] Число пар ног колеблется от 13 (у Ooperipatellus nanus ) до 43 (у Plicatoperipatus jamaicensis ). [10] Их кожа состоит из многочисленных тонких поперечных колец и часто имеет незаметный оранжевый, красный или коричневый цвет, но иногда также ярко-зеленый, синий, золотой или белый, а иногда и украшена узорами других цветов. Сегментация внешне незаметна и определяется регулярным расстоянием между парами ног, правильным расположением пор кожи, органов выделения и концентраций нервных клеток . Отдельные части тела в значительной степени неспециализированы ; даже голова развивается лишь несколько иначе, чем брюшные сегменты. Сегментация, по-видимому, задается тем же геном , что и у других групп животных, и активируется в каждом случае в процессе эмбрионального развития у заднего края каждого сегмента и в зоне роста коротких ног. Хотя онихофоры относятся к группе протостомов , их раннее развитие имеет траекторию вторичного развития (рот и анус формируются отдельно); эта траектория скрыта довольно сложными процессами, происходящими на ранних стадиях развития. [11]
Придатки
[ редактировать ]Короткие лапки, характерные для бархатных червей, представляют собой конические , мешковатые придатки тела, полые внутри и не имеющие суставов. Хотя количество ног может значительно различаться у разных видов, их строение в основном очень похоже. Жесткость обеспечивается гидростатическим давлением их жидкого содержимого, а движение обычно достигается пассивно за счет растяжения и сокращения всего тела животного. Однако каждая нога также может быть укорочена и согнута за счет внутренних мышц . [12] Из-за отсутствия суставов этот изгиб может происходить в любой точке по бокам ноги. У некоторых видов в ступнях находятся два разных органа:
- Голенные железы расположены на плечах ног и заходят в полость тела. Они открываются наружу в сосочках голени — небольших бородавчатых бугорках на брюшной стороне ноги — и выделяют химические вещества-переносчики, называемые феромонами . Их название происходит от латинского cruralis , что означает «ноги». [13]
- Коксальные везикулы представляют собой мешочки, расположенные на брюшной стороне голени, которые могут выворачиваться и, вероятно, служат для поглощения воды. Они принадлежат к семейству Peripatidae и названы от coxa , латинского слова, означающего «бедро». [14]

На каждой стопе имеется пара втягивающихся, затвердевших (склеротизированных) хитиновых когтей, которые дали таксону научное название: Onychophora происходит от древнегреческого : ονυχής , onyches , «когти»; и φέρειν , здесь , «нести». У основания когтей находятся от трех до шести колючих «подушек», на которых нога сидит в положении покоя и на которых животное ходит по гладкому субстрату. Когти используются в основном для прочного закрепления на неровной местности. Каждая клешня состоит из трех сложенных друг на друга элементов, как у русской матрешки . Самый внешний элемент отслаивается во время шелушения, что обнажает следующий элемент, который полностью сформирован, поэтому ему не требуется время для затвердевания перед использованием. [15] Эта отличительная конструкция идентифицирует многие окаменелости раннего кембрия как ранние ответвления линии онихофоров. [15] Помимо пар ног, есть еще три придатка тела, которые находятся в голове и состоят из трех сегментов:
- На первом сегменте головы имеется пара тонких усиков , служащих для сенсорного восприятия . Вероятно, они не соответствуют непосредственно усикам членистоногих. [16] но, возможно, скорее «губами» или губами . У их основания находится пара простых глаз , за исключением некоторых слепых видов. Перед ними у многих австралийских видов имеются различные ямочки, функция которых пока не ясна. Похоже, что по крайней мере у некоторых видов они служат для переноса пакетов сперматозоидов ( сперматофоров ). [ нужна ссылка ]
- На брюшной стороне второго сегмента головы находится верхняя губа — ротовое отверстие, окруженное чувствительными «губами». У бархатных червей эта структура представляет собой мускульный вырост горла , поэтому, несмотря на свое название, она, вероятно, не гомологична верхней губе членистоногих и используется для питания. Глубоко внутри ротовой полости лежат острые серповидные «челюсти» или мандибулы , сильно затвердевшие и напоминающие когти ног, которым они серийно гомологичны; [17] На ранних этапах развития придатки челюстей имеют положение и форму, аналогичные последующим ногам. [18] Челюсти разделены на внутреннюю и внешнюю челюсти, их вогнутая поверхность покрыта мелкими зубчиками. Они движутся вперед и назад в продольном направлении, разрывая добычу, перемещаясь, по-видимому, в одну сторону за счет мускулатуры, а в другую за счет гидростатического давления. [17] Когти состоят из склеротизированного α-хитина, армированного фенолами и хинонами, и имеют однородный состав, за исключением того, что ближе к кончику находится более высокая концентрация кальция, что, по-видимому, обеспечивает большую прочность. [17]
Поверхность мандибул гладкая, без орнамента. [19] Кутикула жвал (и когтей) отличается от остальной части тела. У него есть внутренний и внешний компонент; внешний компонент имеет всего два слоя (тогда как кутикула тела имеет четыре), и эти внешние слои (в частности, внутренняя эпикутикула) обезвожены и сильно загорели, что обеспечивает прочность. [19]
Слизистые железы
[ редактировать ]
На третьем сегменте головы, слева и справа от рта, расположены два отверстия, называемые «ротовыми сосочками». Внутри них находится пара больших, сильно разветвленных внутри слизистых желез. Они лежат примерно в центре тела и выделяют своего рода молочно-белую слизь, которая используется для ловли добычи и в защитных целях. [20] Иногда соединительный «проводник слизи» расширяется до резервуара, который может буферизовать уже произведенную слизь. Сами слизевые железы, вероятно, представляют собой модифицированные железы голеней. [21] Все три структуры соответствуют эволюционному происхождению в парах ног других сегментов. [ нужна ссылка ]
Кожа и мышцы
[ редактировать ]В отличие от членистоногих, бархатные черви не обладают жестким экзоскелетом . Вместо этого заполненная жидкостью полость тела действует как гидростатический скелет, подобно многим отдаленно родственным мягкотелым животным, имеющим цилиндрическую форму, например морским анемонам и различным червям . Давление их несжимаемой внутренней телесной жидкости на стенки тела обеспечивает жесткость, и мышцы способны противодействовать этому. Стенка тела состоит из неклеточной внешней оболочки, кутикулы ; один слой клеток эпидермиса, образующий внутреннюю кожу; а под ним обычно три слоя мышц, погруженных в соединительные ткани. Кутикула толщиной около микрометра покрыта мелкими ворсинками . По составу и строению он напоминает кутикулу членистоногих, состоящую из α-хитина и различных белков . [17] хотя и не содержит коллагена . Ее можно разделить на наружную эпикутикулу и внутреннюю прокутикулу, которые сами состоят из экзо- и эндокутикулы. Эта многоуровневая структура отвечает за высокую гибкость внешней кожицы, что позволяет бархатному червю протискиваться в самые узкие щели. Хотя внешне водоотталкивающая , кутикула не способна предотвратить потерю воды при дыхании , в результате чего бархатные черви могут жить только в микроклимате с повышенной влажностью во избежание высыхания . Поверхность кутикулы усеяна многочисленными тонкими сосочками, более крупные из которых несут видимые ворсинчатые чувствительные щетинки. Сами сосочки покрыты крошечными чешуйками , придающими коже бархатистый вид (отсюда, вероятно, и произошло общее название). На ощупь он также похож на сухой бархат, за что отвечает его водоотталкивающая природа. Линька кожи ( шелушение ) происходит регулярно, примерно каждые 14 дней. [22] индуцируется гормоном экдизоном . Внутренняя поверхность кожи имеет шестиугольный рисунок. [23] При каждой линьке сброшенная кожа заменяется эпидермисом, лежащим непосредственно под ней; в отличие от кутикулы, она состоит из живых клеток. Под ним находится толстый слой соединительной ткани, который состоит в основном из коллагеновых волокон, расположенных параллельно или перпендикулярно продольной оси тела. Окраска Onychophora создается рядом пигментов. [ нужны разъяснения ] Растворимость этих пигментов является полезным диагностическим признаком: у всех членистоногих и тихоходок пигмент тела растворим в этаноле. Это справедливо и для Peripatidae, но у Peripatopsidae пигмент тела нерастворим в этаноле. [24]
Внутри соединительной ткани лежат три непрерывных слоя неспециализированной гладкой мышечной ткани. Относительно толстый внешний слой состоит из кольцевых мышц, а такой же объемистый внутренний слой — из продольных мышц. извиваются вперед и назад вдоль оси тела Между ними лежат тонкие диагональные мышцы, которые по спирали . Между кольцевыми и диагональными мышцами существуют тонкие кровеносные сосуды , которые лежат под поверхностно распознаваемыми поперечными кольцами кожи и отвечают за псевдосегментированные отметины. [12] Под внутренним мышечным слоем находится полость тела. В поперечном сечении он разделен на три отдела так называемыми дорсо-вентральными мышцами, которые проходят от середины нижней части живота к краям верхней стороны: центральный срединный отдел, а слева и справа - два боковых отдела. это также включает в себя ноги. [ нужна ссылка ]
Тираж
[ редактировать ]Полость тела известна как «псевдоцель», или гемоцель . В отличие от настоящего целома , псевдоцель не полностью окружен клеточным слоем, происходящим из эмбриональной мезодермы . , формируется целом Однако вокруг гонад , выводящих отходы и нефридиев . [12] Как следует из названия гемокоэль , полость тела заполнена кровеподобной жидкостью, в которой заключены все органы; таким образом, их можно легко снабжать питательными веществами , циркулирующими в крови. Эта жидкость бесцветна, так как не содержит пигментов ; по этой причине он играет лишь ограниченную роль в транспорте кислорода .В жидкости циркулируют два разных типа клеток крови (или гемоцитов): амебоциты и нефроциты. Амебоциты, вероятно, защищают от бактерий и других инородных тел; у некоторых видов они также играют роль в размножении . Нефроциты поглощают токсины или преобразуют их в форму, пригодную для выведения нефридиями. [ нужна ссылка ]
Гемоцель разделен горизонтальной перегородкой — диафрагмой — на две части: перикардиальный синус на спине и перивисцеральный синус на брюшке. Первая окружает трубчатое сердце, а вторая — другие органы. Диафрагма перфорирована во многих местах, что обеспечивает обмен жидкостями между двумя полостями. [ нужна ссылка ] Само сердце представляет собой трубку кольцевых мышц, состоящую из эпителиальных тканей, с двумя боковыми отверстиями ( остиями ) на каждый сегмент. Хотя неизвестно, открыта или закрыта задняя часть, спереди она открывается прямо в полость тела.
Поскольку кровеносных сосудов нет, за исключением мелких сосудов, проходящих между мышечными слоями стенки тела, и пары артерий, кровоснабжающих усики, это называется открытым кровообращением . [ нужна ссылка ] Время процедуры сцеживания можно разделить на две части: диастолу и систолу . Во время диастолы кровь течет через отверстие из синуса перикарда (полости, содержащей сердце) в сердце. Когда начинается систола, отверстия закрываются, и сердечные мышцы сокращаются внутрь, уменьшая объем сердца. При этом кровь перекачивается из передней части сердца в перивисцеральный синус, содержащий органы. Таким образом, различные органы снабжаются питательными веществами, прежде чем кровь наконец вернется в синус перикарда через перфорации диафрагмы. Помимо насосной функции сердца, на кровообращение также влияют движения тела. [ нужна ссылка ]
Дыхание
[ редактировать ]Поглощение кислорода в некоторой степени происходит за счет простой диффузии через всю поверхность тела, при этом у некоторых видов, возможно, участвуют коксальные пузырьки на ногах. Однако наиболее важным является газообмен через тонкие неразветвленные трубки, трахеи , которые переносят кислород с поверхности глубоко в различные органы, особенно в сердце.
Стенки этих структур, общая толщина которых составляет менее трех микрометров, состоят только из чрезвычайно тонкой мембраны , через которую может легко диффундировать кислород. Трахеи берут начало в крошечных отверстиях, дыхалцах , которые сами сгруппированы в вмятинчатых углублениях внешней кожи, предсердиях . Число образующихся таким образом «пучков трахеи» составляет в среднем около 75 пучков на сегмент тела; наиболее плотно они скапливаются на задней части организма. [ нужна ссылка ]
В отличие от членистоногих, бархатные черви не способны контролировать отверстия своих трахей; трахеи всегда открыты, что приводит к значительной потере воды в засушливых условиях. Вода теряется в два раза быстрее, чем у дождевых червей, и в сорок раз быстрее, чем у гусениц. [25] По этой причине бархатные черви зависят от мест обитания с высокой влажностью воздуха. [26] [27]
Переносу кислорода способствует гемоцианин, переносчик кислорода . [28]
Пищеварение
[ редактировать ]
Пищеварительный тракт начинается немного позади головы, рот лежит на нижней стороне, немного дальше от самой передней точки тела. Здесь добычу можно механически расчленить челюстями, покрытыми тонкими зубцами. Две слюнные железы отходят по общему проводнику в последующее «горло», составляющее первый отдел передней кишки . Слюна, которую они производят, содержит слизь и гидролитические ферменты , которые инициируют пищеварение во рту и за его пределами.
Горло само по себе очень мускулистое, служит для поглощения частично разжиженной пищи и перекачивания ее через пищевод , который образует заднюю часть передней кишки, в центральную кишку. В отличие от передней кишки, она не выстлана кутикулой, а состоит только из одного слоя эпителиальной ткани, на которой нет заметных углублений, как у других животных.
При попадании в центральную кишку частицы пищи покрываются перитрофической мембраной на основе слизи , которая защищает слизистую оболочку кишечника от повреждения частицами с острыми краями. Кишечный эпителий выделяет дополнительные пищеварительные ферменты и поглощает высвободившиеся питательные вещества, хотя большая часть пищеварения уже произошла снаружи или во рту. Неперевариваемые остатки попадают в задний отдел кишечника, или прямую кишку , которая снова покрыта кутикулой и открывается в анус , расположенный на нижней стороне рядом с задним концом. [ нужна ссылка ]
Почти в каждом сегменте имеется пара органов выделения, называемых нефридиями, которые происходят из ткани целома. Каждый состоит из небольшого мешочка, который соединен жгутиковым проводником, называемым нефридиодуктом, с отверстием в основании ближайшей ножки, известным как нефридиопор. Мешок заполнен специальными клетками, называемыми подоцитами , которые способствуют ультрафильтрации крови через перегородку между гемоцелом и нефридием.
Состав мочевого раствора модифицируется в нефридиодукте путем избирательного восстановления питательных веществ и воды, а также выделения ядов и отходов, прежде чем он выводится во внешний мир через нефридиопор. Важнейшим азотистым продуктом экскреции является нерастворимая в воде мочевая кислота ; он может выводиться из организма в твердом состоянии с очень небольшим количеством воды. Этот так называемый урикотелический экскреторный режим представляет собой приспособление к жизни на суше и связанную с этим необходимость экономного обращения с водой. [ нужна ссылка ]
Пара бывших нефридиев в голове вторично превратилась в слюнные железы, а другая пара в последнем сегменте самцов теперь служит железами, которые, по-видимому, играют роль в размножении. [29]
ощущение
[ редактировать ]Все тело, включая короткие ступни, усеяно многочисленными сосочками: бородавчатыми выступами, реагирующими на прикосновение, которые несут на кончике механорецептивную щетину, каждая из которых также связана с другими чувствительными нервными клетками, лежащими под ней. Ротовые сосочки, выходы слизистых желез, вероятно, также выполняют некоторую функцию в сенсорном восприятии . Сенсорные клетки, известные как «сенсиллы» на «губах» или верхней губе, реагируют на химические стимулы и известны как хеморецепторы . Они также находятся на двух усиках, которые, по-видимому, являются наиболее важными органами чувств бархатного червя. [ нужна ссылка ]
За исключением некоторых видов (обычно подземных ), один глаз простой конструкции (глазок) расположен позади каждой антенны, сбоку, прямо под головой. [30] Он состоит из хитинового шарика -хрусталика , роговицы и сетчатки и связан с центром мозга посредством зрительного нерва . [30] Сетчатка состоит из многочисленных пигментных клеток и фоторецепторов; последние представляют собой легко модифицируемые жгутиковые клетки, мембраны жгутиков которых несут на своей поверхности светочувствительный пигмент.
глаза Считается, что рабдомерные Onychophora гомологичны срединным глазкам членистоногих; это позволяет предположить, что у последнего общего предка членистоногих могли быть только срединные глазки. [30] Однако иннервация показывает, что гомология ограничена: глаза Onychophora формируются за антенной, тогда как у членистоногих верно обратное. [31]
Воспроизведение
[ редактировать ]Оба пола обладают парами гонад , открывающихся через канал, называемый гонодуком, в общее половое отверстие, гонопору , расположенное на задней брюшной стороне. И гонады, и гонодук происходят из истинной ткани целома. [ нужна ссылка ]
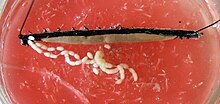
У самок два яичника соединены посередине и с горизонтальной диафрагмой. Гонодук появляется по-разному в зависимости от того, является ли вид живородящим или яйцекладущим . У живородящих видов каждый выходной канал делится на тонкий яйцевод и вместительную «матку» — матку , в которой развиваются зародыши. Единственное влагалище , к которому соединены обе матки, выходит наружу к гонопору. У яйцекладущих видов, гонодук которых устроен равномерно, половое отверстие лежит на кончике длинного яйцекладочного аппарата — яйцеклада . У самок многих видов также есть хранилище спермы, называемое резервуаром Seminis , в котором сперматозоиды самцов могут храниться временно или в течение более длительных периодов времени. [ нужна ссылка ]
У самцов имеются два отдельных семенника , а также соответствующий сперматозоидный пузырек ( семенной пузырь ) и выходной канал ( выносящая трубка ). Два эфферентных протока соединяются в общий семявыносящий проток, который , в свою очередь, расширяется через семявыбрасывающий канал и открывается в гонопоре. Непосредственно рядом или позади него лежат две пары особых желез, выполняющих, вероятно, какую-то вспомогательную репродуктивную функцию; самые задние железы также известны как анальные железы. [32] Пенисоподобная но структура пока обнаружена только у самцов рода Paraperipatus, в действии еще не наблюдалась. [ нужна ссылка ]
Существуют разные процедуры спаривания: у некоторых видов самцы вносят свой сперматофор непосредственно в отверстие гениталий самки, в то время как другие помещают его на тело самки, где кутикула разрушается и позволяет сперматозоидам мигрировать в самку. Есть также австралийские виды, у которых самцы помещают сперматофор на макушку головы, который затем прижимается к гениталиям самки. У этих видов голова имеет сложные структуры, такие как шипы, шипы, полые стилеты, ямки и впадины, цель которых — либо удерживать сперму, либо способствовать передаче спермы самке. Самцы большинства видов также выделяют феромоны из желез на нижней стороне ног, чтобы привлечь самок. [33]
Распространение и среда обитания
[ редактировать ]Распределение
[ редактировать ]Бархатные черви обитают во всех тропических средах обитания и в умеренной зоне Южного полушария , демонстрируя циркумтропическое и околоюжное распространение. Отдельные виды встречаются в Центральной и Южной Америке ; Карибские острова ; экваториальная Западная Африка и Южная Африка ; северо-восток Индии ; [34] [35] Таиланд ; [36] Индонезия и часть Малайзии ; Новая Гвинея ; Австралия ; и Новая Зеландия . [ нужна ссылка ]
Окаменелости были найдены в балтийском янтаре , что указывает на то, что раньше они были более распространены в Северном полушарии, когда условия были более подходящими. [37]
среда обитания
[ редактировать ]Бархатные черви всегда редко занимают места обитания: они редки среди фауны, частью которой являются. [ нужна ссылка ]
Все современные бархатные черви являются наземными (наземными) и предпочитают темную среду с высокой влажностью воздуха. Они встречаются особенно в тропических лесах тропиков и умеренных зон, где живут среди мховых подушек и опавших листьев , под стволами деревьев и камнями, в гниющей древесине или в термитных туннелях. Они также встречаются на незалесенных лугах имеется достаточно расщелин , в которые они могут укрыться в течение дня. , если в почве [ нужна ссылка ] и в пещерах. [38] Два вида обитают в пещерах , среде обитания, к которой их способность протискиваться в мельчайшие щели делает их исключительно хорошо приспособленными и в которой гарантированы постоянные условия жизни. Поскольку основные требования для пещерной жизни, вероятно, существовали еще до заселения этих мест обитания, это можно назвать экзаптацией .
Некоторые виды бархатных червей способны заселять измененные человеком земли, такие как какао и бананов плантации в Южной Америке и Карибском бассейне, но для других преобразование тропических лесов, вероятно, является одной из наиболее важных угроз их выживанию. [ нужна ссылка ] (см. Сохранение ). [ нужны разъяснения ]
Бархатные черви фотофобны: их отпугивают яркие источники света. [39] Поскольку опасность высыхания наиболее велика днем и в сухую погоду, неудивительно, что бархатные черви обычно наиболее активны ночью и в дождливую погоду. В холодных или сухих условиях они активно ищут расщелины, в которых переводят свое тело в состояние покоя. [40]
Слизь
[ редактировать ]
Онихофора с силой выплевывает клейкую слизь. [41] из их оральных сосочков; они делают это либо для защиты от хищников, либо для поимки добычи. [42] Отверстия желез, вырабатывающих слизь, находятся в сосочках — паре сильно видоизмененных конечностей по бокам головы под усиками. Внутри у них есть система, похожая на шприц, которая с помощью геометрического усилителя позволяет быстро впрыскивать, используя медленное мышечное сокращение. [43] На высокоскоростных пленках видно, как животное выбрасывает два потока клейкой жидкости через небольшое отверстие (от 50 до 200 микрон) со скоростью от 3 до 5 м/с (от 10 до 20 футов/с). [43] Взаимодействие между эластичностью сосочков полости рта и быстрым нестационарным потоком приводит к пассивному колебательному движению (30–60 Гц) сосочков полости рта. [43] Колебания заставляют потоки пересекаться в воздухе, сплетая беспорядочную сеть; бархатные черви могут контролировать только общее направление, в котором забрасывается сеть. [44]
Сами слизевые железы находятся глубоко внутри полости тела, каждая на конце трубки длиной более половины длины тела. Трубка одновременно проводит жидкость и сохраняет ее до тех пор, пока она не понадобится. Расстояние, на которое животное может переместить слизь, варьируется; обычно он брызгает примерно на сантиметр, [45] но по разным сообщениям максимальная дальность составляет десять сантиметров, [46] или даже почти фут, [47] хотя точность падает с дальностью. [48] Неясно, в какой степени ареал варьируется в зависимости от вида и других факторов. Одной струи обычно достаточно, чтобы поймать добычу, хотя более крупная добыча может быть дополнительно обездвижена более мелкими струями, направленными на конечности; кроме того, иногда целью становятся клыки пауков. [48] При выбросе он образует сеть нитей диаметром около двадцати микрон, по длине которых равномерно расположены капли вязкой клейкой жидкости. [45] Впоследствии он высыхает, сморщивается, теряет липкость и становится ломким. [45] Онихофоры поедают высушенную слизь, когда могут, и это вполне уместно, поскольку онихофорану требуется около 24 дней, чтобы пополнить истощенное хранилище слизи. [48]
Слизь может составлять до 11% сухого веса организма. [48] и на 90% состоит из воды; его сухой остаток состоит в основном из белков — преимущественно белка коллагенового типа. [45] 1,3% сухого веса слизи составляют сахара, главным образом галактозамин . [45] Слизь также содержит липиды и поверхностно-активное вещество нонилфенол . Онихофоры — единственные известные организмы, производящие это последнее вещество. [45] На вкус он «слегка горький и в то же время несколько вяжущий». [49] и эластичность слизи Белковый состав обеспечивает высокую прочность . [45] Липидные и нонилфенольные компоненты могут служить одной из двух целей. Они могут выстилать канал выброса, предотвращая прилипание слизи к организму во время ее выделения; или они могут замедлить процесс высыхания на время, достаточное для того, чтобы слизь достигла своей цели. [45]
Поведение
[ редактировать ]Передвижение
[ редактировать ]
Бархатные черви/онихофоры движутся медленно и постепенно, поэтому добыче их трудно заметить. [48] Их туловище поднято сравнительно высоко над землей, и они ходят неперекрывающимися шагами. [50] Чтобы передвигаться с места на место, бархатный червь ползет вперед, используя ноги; в отличие от членистоногих, обе ноги пары двигаются одновременно. Когти ног используются только на твердой, пересеченной местности, где необходим надежный захват; на мягких субстратах, например мхе, бархатный червь ходит по подушечкам лапок у основания когтей. [ нужна ссылка ]
Фактическое передвижение достигается не столько за счет напряжения мышц ног, сколько за счет локальных изменений длины тела. Управлять этим можно с помощью кольцевых и продольных мышц. Если кольцевые мышцы сокращаются, поперечное сечение тела уменьшается, а соответствующий сегмент удлиняется; это обычный режим работы гидростатического скелета, который применяется и у других червей. За счет растяжения ноги рассматриваемого сегмента приподнимаются и поворачиваются вперед. Локальное сокращение продольных мышц затем укорачивает соответствующий сегмент, и ноги, соприкасающиеся теперь с землей, перемещаются назад. Эта часть локомотивного цикла представляет собой собственно ход ноги, который отвечает за движение вперед. Отдельные растяжения и сокращения сегментов координируются нервной системой, так что волны сокращения проходят по всей длине тела, каждая пара ног последовательно качается вперед, а затем вниз и назад. Macroperipatus может развивать скорость до четырех сантиметров в секунду. [48] хотя более типична скорость около 6 длин тела в минуту. [51] Тело становится длиннее и уже по мере того, как животное набирает скорость; длина каждой ноги также меняется во время каждого шага. [51]
Социальность
[ редактировать ]Мозг Onychophora хоть и небольшой, но очень сложен; следовательно, организмы способны к довольно сложным социальным взаимодействиям. [52] Поведение может варьироваться от рода к роду, поэтому в этой статье отражен наиболее изученный род Euperipatoides . [52]
Euperipatoides образуют социальные группы численностью до пятнадцати особей, обычно тесно связанных между собой, которые обычно живут и охотятся вместе. Группы обычно живут вместе; в более засушливых регионах примером общего дома может служить влажная внутренняя часть гниющего бревна. Члены группы крайне агрессивны по отношению к особям из других логов. [52] Доминирование достигается посредством агрессии и поддерживается посредством покорного поведения. [52] После добычи первой всегда кормится доминирующая самка, за ней по очереди следуют другие самки, затем самцы и потомство. [52]
Оценивая других людей, люди часто измеряют друг друга, проводя усиками по длине другого человека. [52] Как только иерархия установлена, пары людей часто собираются вместе, образуя «агрегат»; быстрее всего это происходит в парах самец-самка, за ними следуют пары самок, а затем пары самцов. [52]
Социальная иерархия устанавливается посредством ряда взаимодействий: люди более высокого ранга будут преследовать и кусать своих подчиненных, в то время как последние пытаются залезть на них сверху. [52] Молодые особи никогда не ведут себя агрессивно, а забираются на взрослых, которые терпят их присутствие на спине. [52] Иерархия быстро устанавливается среди особей одной группы, но не среди организмов из разных групп; они значительно более агрессивны и очень редко взбираются друг на друга или образуют скопления. [52] Лица в отдельном журнале обычно тесно связаны; особенно с мужчинами. Это может быть связано с сильной агрессией между неродственными самками. [52]
Кормление
[ редактировать ]
Бархатные черви — хищники-засадники , охотящиеся только ночью . [48] и способны ловить животных, по крайней мере, своего размера, хотя для поимки крупной добычи может потребоваться почти вся их слизь . способность выделять [53] Они питаются практически любыми мелкими беспозвоночными, включая мокриц ( Isopoda ), термитов ( Isoptera ), сверчков ( Gryllidae ), книжных/корых вшей ( Psocoptera ), тараканов ( Blattidae ), многоножек и многоножек ( Myriapoda ), пауков ( Araneae ), [53] различные черви и даже крупные улитки ( Gastropoda ). В зависимости от размера они едят в среднем раз в одну-четыре недели. [48] Они считаются экологически эквивалентными многоножкам ( Cilopada ).
Наиболее энергетически выгодная добыча составляет две пятых размера охотничьего онихофорана. [48] Девяносто процентов времени, потраченного на поедание добычи, тратится на ее проглатывание; повторное проглатывание слизи, используемой для ловли насекомого, происходит в то время, когда онихофоран находит подходящее место для прокалывания добычи, и на эту фазу приходится около 8% времени кормления, а оставшееся время равномерно распределяется между осмотром, впрыскиванием и введение добычи. [48] В некоторых случаях куски добычи откусываются и проглатываются; Неперевариваемым компонентам требуется около 18 часов, чтобы пройти через пищеварительный тракт. [17]
Онихофоры, вероятно, в первую очередь не используют зрение для обнаружения своей добычи; хотя их крошечные глаза обладают хорошей способностью формировать изображения, их переднее зрение закрыто усиками; [48] их ночной образ жизни также ограничивает полезность зрения. Считается, что воздушные потоки, образующиеся в результате движения добычи, являются основным способом обнаружения добычи; роль запаха, если таковая имеется, неясна. [48] Поскольку проглатывание добычи занимает очень много времени, охота в основном происходит в сумерках; онихофоры бросают свою добычу на восходе солнца. [48] Такой хищнический образ жизни, вероятно, является следствием потребности бархатного червя оставаться влажным. Из-за постоянного риска высыхания на поиски еды часто выделяется всего несколько часов в день. Это приводит к сильному отбору в пользу низкого соотношения затрат и выгод, чего невозможно достичь с помощью травоядной диеты. [ нужна ссылка ]
Бархатные черви буквально подкрадываются к своей добыче, их плавные, плавные и плавные движения ускользают от обнаружения. [48] Достигнув добычи, они очень мягко прикасаются к ней усиками, чтобы оценить ее размер и пищевую ценность. После каждого тыкания антенну поспешно убирают, чтобы не насторожить добычу. [48] Это расследование может длиться более десяти секунд, пока бархатный червь не примет решение, нападать ли на него, или пока он не потревожит жертву и она не убежит. [48] Голодные онихофоры тратят меньше времени на исследование добычи и быстрее применяют слизь. [48] После того как слизь распылена, онихофоры полны решимости преследовать и пожирать свою добычу, чтобы окупить затраты энергии. Было замечено, что они тратят до десяти минут на поиск удаленной добычи, после чего возвращаются в свою слизь, чтобы съесть ее. [48] В случае с добычей меньшего размера они могут вообще не использовать слизь. [48] Впоследствии мягкая часть объекта добычи (обычно суставная оболочка у добычи членистоногих) идентифицируется, прокалывается укусом челюстей и впрыскивается слюной. Это очень быстро убивает добычу и запускает более медленный процесс пищеварения. [48] Пока онихофоран ждет, пока добыча переварится, он выделяет слюну на ее слизь и начинает поедать ее (и все, что к ней прикреплено). Впоследствии он растягивает и разрезает раннюю перфорацию, чтобы получить доступ к теперь уже разжиженной внутренней части своей добычи. [48] Челюсти действуют, перемещаясь вперед и назад вдоль оси тела (а не зажимающими движениями из стороны в сторону, как у членистоногих), предположительно используя сочетание мускулатуры и гидростатического давления. [17] Глотка специально приспособлена для сосания, для извлечения разжиженных тканей; Расположение челюстей вокруг сосочков языка и губ обеспечивает плотное прилегание и установление присасывания. [17] В социальных группах доминирующая самка кормится первой, не позволяя конкурентам получить доступ к добыче в течение первого часа кормления. Впоследствии начинают питаться подчиненные особи. Численность самцов достигает пика после того, как самки начинают покидать объект добычи. [52] После кормления особи чистят свои усики и ротовую часть, прежде чем снова присоединиться к остальной группе. [52]
Размножение и жизненный цикл
[ редактировать ]Почти все виды бархатного червя размножаются половым путем. Единственное исключение — Epiperipatus imthurni , у которого самцов не обнаружено; вместо этого размножение происходит путем партеногенеза . [54] Все виды в принципе различны по половому признаку и во многих случаях имеют выраженный половой диморфизм : самки обычно крупнее самцов , а у видов с переменным числом ног их больше. Самки многих видов оплодотворяются только один раз в жизни, что приводит к совокуплению, которое иногда происходит до того, как репродуктивные органы самок полностью разовьются. В таких случаях, например в трехмесячном возрасте у Macroperipatus torquatus , перенесенные сперматозоиды сохраняются в специальном резервуаре , где они могут сохранять жизнеспособность в течение более длительного периода времени. Оплодотворение происходит внутри организма , хотя способы передачи сперматозоидов широко варьируются. У большинства видов, например у рода Peripatus пакет сперматозоидов, называемый сперматофором , в половое отверстие самки помещается .
Подробный процесс, с помощью которого это достигается, в большинстве случаев до сих пор неизвестен, настоящий пенис наблюдался только у видов рода Paraperipatus . имеются ямочки или особые образования в форме кинжала или топора У многих австралийских видов на голове ; Самец Florelliceps stutchburyae прижимает длинный шип к половому отверстию самки и, вероятно, таким образом размещает там свой сперматофор. Во время этого процесса самка поддерживает самца, сжимая его когтями своей последней пары ног. брачное поведение двух видов рода Peripatopsis Особенно любопытно . Здесь самец размещает двухмиллиметровые сперматофоры на спине или боках самки. Амебоциты самки из крови собираются на внутренней стороне места отложения как оболочка сперматофора, так и стенка тела, на которой он находится, разлагаются посредством секреции ферментов , и . При этом сперматозоиды высвобождаются, которые затем свободно перемещаются через гемоцель , проникают через внешнюю стенку яичников и, наконец, оплодотворяют яйцеклетка . Почему это нанесенное самому себе повреждение кожи не приводит к бактериальным инфекциям, пока не понятно (хотя, вероятно, это связано с ферментами, используемыми для разрушения кожи или облегчения передачи жизнеспособного генетического материала от мужчины к женщине). Бархатные черви встречаются в яйцекладущей ( яйцеродящей ), яйцеживородящей ( яйцеживородящей ) и живородящей ( живородящей ) формах.
В недавней рецензируемой статье, опубликованной в «Журнале зоологии», исследователи обнаружили, что некоторые виды Peripatus демонстрируют уникальную форму родительской заботы. В отличие от большинства беспозвоночных, у которых участие родителей минимально, самки Peripatus активно охраняют свои яйца и даже обеспечивают защиту потомству после вылупления. Это открытие бросает вызов традиционному пониманию репродуктивного поведения беспозвоночных и подчеркивает разнообразие родительских стратегий в животном мире. [55]
- Яйцекладок встречается исключительно у Peripatopsidae, часто в регионах с неустойчивым снабжением пищей или неустойчивым климатом. В этих случаях желтком, , богатые яйца имеют размер от 1,3 до 2,0 мм и покрыты защитной хитиновой оболочкой. Материнский уход неизвестен.
- Большинство видов яйцеживородящие: яйца среднего размера, покрытые только двойной оболочкой, остаются в матке . Эмбрионы называют не получают питание непосредственно от матери, а вместо этого получают умеренное количество желтка, содержащегося в яйцах, поэтому их лецитотрофными . Молодняк выходит из яиц незадолго до рождения. Вероятно, это отражает первоначальный способ размножения бархатного червя, т. е. как яйцекладущие, так и живородящие виды произошли от яйцеживородящих видов.
- Настоящие живородящие виды встречаются в обоих семействах, особенно в тропических регионах со стабильным климатом и регулярным снабжением пищей в течение всего года. Эмбрионы развиваются из яиц размером всего несколько микрометров и питаются матерью в матке, отсюда и название « матротрофные ». Поставка пищи происходит либо через выделения матери непосредственно в матку, либо через настоящую тканевую связь между эпителием матки и развивающимся эмбрионом, известную как плацента . Первый встречается только за пределами американских континентов, а второй встречается преимущественно в Америке и странах Карибского бассейна и реже в Старом Свете . Период беременности может составлять до 15 месяцев, по истечении которых потомство появляется на продвинутой стадии развития. Эмбрионы, обнаруженные в матке одной самки, не обязательно должны быть одного возраста; вполне возможно, что потомство находится на разных стадиях развития и происходит от разных самцов. У некоторых видов молодь, как правило, выпускается только в определенные моменты года. [56]
Самка может иметь от 1 до 23 потомков в год; Развитие от оплодотворенной яйцеклетки до взрослой особи занимает от 6 до 17 месяцев и не имеет личиночной стадии. Вероятно, это также первоначальный способ развития. Известно, что бархатные черви живут до шести лет.
Экология
[ редактировать ]Важными хищниками бархатного червя являются в первую очередь различные пауки и многоножки, а также грызуны и птицы , такие как в Центральной Америке дрозд глинистого цвета ( Turdus Grayi ). В Южной Америке коралловая змея Хемпричи ( Micrurus hemprichii ) питается почти исключительно бархатными червями. [57] Для защиты некоторые виды рефлекторно сворачиваются в спираль, а также могут отбиваться от более мелких противников, выбрасывая слизь. Известно, что различные клещи ( Acari ) являются эктопаразитами, поражающими кожу бархатного червя. Повреждения кожи обычно сопровождаются бактериальными инфекциями, которые почти всегда приводят к летальному исходу. [ нужна ссылка ]
Южноафриканский вид Peripatopsis capensis был случайно завезен на остров Санта-Крус на Галапагосских островах , где он встречается вместе с местными бархатными червями. [58]
Сохранение
[ редактировать ]Глобальный статус сохранения видов бархатных червей оценить трудно; Известно, что многие виды существуют только в своей типовой местности (месте, где они впервые были обнаружены и описаны). Сбору надежных данных также препятствуют низкая плотность населения, их типичное ночное поведение и, возможно, также еще недокументированные сезонные влияния и половой диморфизм. На сегодняшний день единственными онихофорами, оцененными МСОП, являются:
- Mesoperipatus tholloni (недостаточно данных)
- Plicatooperipatus jamaicensis (под угрозой исчезновения)
- Peripatoides indigo (уязвимый)
- Peripatoides suteri (Уязвимый)
- Peripatopsis alba (Уязвимый)
- Peripatopsis clavigera (Уязвимый)
- Macroperipatus insularis (находится под угрозой исчезновения)
- Leucopatus anophthalmus (находится под угрозой исчезновения)
- Opisthopatus roseus (находящийся под угрозой исчезновения)
- Peripatopsis leonina (находящийся под угрозой исчезновения)
- Speleoperipatus spelaeus (находящийся под угрозой исчезновения) [59]
Основная угроза исходит от разрушения и фрагментации среды обитания бархатного червя из-за индустриализации , осушения водно-болотных угодий и подсечно-огневого земледелия. Многие виды также имеют естественную низкую плотность населения и строго ограниченный географический ареал; в результате относительно небольшие локальные нарушения важных экосистем могут привести к исчезновению целых популяций или видов. Сбор образцов для университетов или исследовательских институтов также играет роль в местном масштабе. [60] Существует очень выраженная разница в защите бархатных червей между регионами: в некоторых странах, таких как Южная Африка, существуют ограничения как на сбор, так и на экспорт , тогда как в других, таких как Австралия, существуют только ограничения на экспорт. Многие страны вообще не предлагают никаких конкретных гарантий. В Тасмании действует уникальная во всем мире программа защиты: в одном регионе леса имеется собственный план сохранения бархатных червей, адаптированный к конкретному виду бархатных червей. [60]
Филогения
[ редактировать ]
В своей нынешней форме бархатные черви, вероятно, очень тесно связаны с членистоногими — очень обширным таксоном, включающим, например, ракообразных, насекомых и паукообразных. У них, среди прочего, общий экзоскелет, состоящий из α-хитина и неколлагеновых белков; гонады и органы удаления отходов, заключенные в настоящую ткань целома; открытая кровеносная система с трубчатым сердцем, расположенным сзади; брюшная полость разделена на перикардиальную и перивисцеральную полости; дыхание через трахеи; и подобное эмбриональное развитие. Сегментация с двумя придатками тела на сегмент также является общей чертой.
Однако усики, мандибулы и ротовые сосочки бархатных червей, вероятно, не гомологичны соответствующим признакам членистоногих; т.е. они, вероятно, развивались независимо.
Другая близкородственная группа — сравнительно малоизвестные водяные медведи ( Tardigrada ); однако из-за своего очень маленького размера водяные медведи не нуждаются в кровообращении и отдельных дыхательных структурах и, следовательно, не имеют их: общие характеристики, которые подтверждают общее происхождение бархатных червей и членистоногих.
Вместе бархатные черви, членистоногие и водяные медведи образуют монофилетический таксон Panarthropoda , т. е. три группы в совокупности охватывают всех потомков их последнего общего предка. Из-за определенного сходства формы бархатных червей обычно группировали с водяными медведями, образуя таксон Protoarthropoda. Это обозначение означало бы, что и бархатные черви, и водяные медведи еще не так высоко развиты, как членистоногие. Современные систематические теории отвергают такие представления о «примитивных» и «высокоразвитых» организмах и вместо этого рассматривают исключительно исторические связи между таксонами. Эти взаимоотношения еще не до конца изучены, но считается вероятным, что сестринские группы бархатных червей образуют таксон, обозначенный как Tactopoda , таким образом:
| панартроподы |
| ||||||||||||


Долгое время родственными кольчатым червям считали и бархатных червей . Помимо всего прочего, у них общее червеобразное тело; тонкая и гибкая внешняя оболочка; многослойная мускулатура; парные органы удаления отходов; а также просто устроенный мозг и простые глаза. Решающим, однако, было наличие сегментации в обеих группах, причем сегменты демонстрировали лишь незначительную специализацию. Таким образом, придатки параподий, обнаруженные у кольчатых червей, соответствуют культям ног бархатных червей. В рамках гипотезы Articulata , разработанной Жоржем Кювье , бархатные черви, таким образом, образовали эволюционное звено между кольчатыми червями и членистоногими: у червеобразных предшественников сначала развились параподии, которые затем развились в короткие лапки как промежуточное звено в окончательном развитии членистоногих. 'придатки. Из-за структурного консерватизма бархатных червей считали «живыми ископаемыми». Парадигматично эта точка зрения выражена в высказывании французского зоолога А. Ванделя:
- Онихофорами можно считать высокоразвитых кольчатых червей, приспособленных к наземной жизни, которые пророчески объявили членистоногими. Это боковая ветвь, которая сохранилась с древних времен до наших дней без существенных изменений.
Современная систематика не изучает такие критерии, как «высшая» и «низшая» стадии развития или различия между «главными» и «побочными» ветвями — только семейные отношения, обозначенные кладистическими актуальными считаются методами. С этой точки зрения, несколько общих характеристик все еще поддерживают гипотезу Articulata — сегментированное тело; парные придатки на каждом сегменте; попарное расположение органов удаления отходов в каждом сегменте; и, прежде всего, нервная система, похожая на веревочную лестницу, основанная на двойной нервной нити, лежащей вдоль живота. Альтернативная концепция, наиболее широко распространенная сегодня, — это так называемая Ecdysozoa гипотеза . Это помещает кольчатых червей и Panarthropoda в две совершенно разные группы: первые - в Lophotrochozoa , а вторые - в Ecdysozoa. Последовательности митохондриальных генов также подтверждают эту гипотезу. [61] Сторонники этой гипотезы предполагают, что вышеупомянутые сходства между кольчатыми червями и бархатными червями либо развились конвергентно, либо были примитивными характеристиками, переданными в неизмененном виде от общего предка как Lophotrochozoa, так и Ecdysozoa. Например, в первом случае веревочно-лестничная нервная система развилась бы в обеих группах независимо, а во втором случае это очень старый признак, не предполагающий особо тесного родства кольчатых червей и панартропод. Концепция Ecdysozoa делит таксон на два: Panarthropoda, к которому относятся бархатные черви, и сестринскую группу Cycloneuralia , включающую острицов ( Nematoda ), червей из конского волоса ( Nematomorpha ) и три довольно малоизвестные группы: грязевых драконов ( Kinorhyncha ); половые глисты ( Priapulida ); и кисточки ( Loricifera ).
| протостомия |
| |||||||||||||||


Особенно характерным для Cycloneuralia является кольцо «околоморальных» нервов вокруг ротового отверстия, которое сторонники гипотезы Ecdysozoa в измененной форме узнают и в деталях нервных узоров Panarthropoda. Обе группы также имеют общий механизм шелушения кожи ( шелушение ) и молекулярно-биологическое сходство. Одной из проблем гипотезы Ecdysozoa является субтерминальное положение рта бархатных червей: в отличие от Cycloneuralia, рот находится не на переднем конце тела, а лежит дальше назад, под брюхом. Однако исследования биологии их развития, особенно в отношении развития головных нервов, позволяют предположить, что это было не всегда так и что изначально рот был конечным (расположен на кончике тела). Это подтверждается летописью окаменелостей .
Гипотеза о «стволовой группе членистоногих» очень широко принята, но некоторые деревья предполагают, что онихофоры могут занимать другое положение; анатомия их мозга более тесно связана с анатомией хелицератов, чем с анатомией любого другого членистоногого. [62] Современные бархатные черви образуют монофилетическую группу, включающую всех потомков их общего предка. Важные общие производные характеристики ( синапоморфии ) включают, например, нижние челюсти второго сегмента тела, ротовые сосочки и связанные с ними слизевые железы третьего; нервные тяжи, идущие вдоль нижней стороны, с многочисленными поперечными связями на сегмент; и особая форма трахеи. около 180 современных видов, включающих 49 родов ; К 2011 г. было описано [63] фактическое количество видов, вероятно, примерно вдвое больше. Согласно более недавнему исследованию, на данный момент описано 82 вида Peripatidae и 115 видов Peripatopsidae. Однако из 197 видов 20 относятся к nomina dubia из-за серьезных таксономических несоответствий. [64] Наиболее известным является типовой род Peripatus , который был описан еще в 1825 году и который в англоязычных странах является репрезентативным для всех бархатных червей. Все роды отнесены к одному из двух семейств, ареалы которых не перекрываются, а разделены засушливыми территориями или океанами:
- Peripatopsidae обладают относительно многими характеристиками, которые воспринимаются как оригинальные или «примитивные». Число пар ног в этом семействе колеблется от 13 (у Ooperipatellus nanus [10] ) до 29 (у Paraperipatus papuensis [65] [66] ). [67] Позади или между последней парой ног находится половое отверстие ( гонопор ). [67] [68] Существуют как яйцекладущие, так и яйцеживородящие, а также истинно живородящие виды, хотя у перипатопсид практически отсутствует плацента. Их распространение носит околоюжный характер и охватывает Австралазию , Южную Африку и Чили . [64]
- Peripatidae обладают рядом производных особенностей. В среднем они длиннее, чем Peripatopsidae, а также у них больше ног. Число пар ног в этом семействе колеблется от 19 (у Typhloperipatus williamsoni [69] ) до 43 (у Plicatoperipatus jamaicensis [10] ). [67] [68] Гонопор всегда находится между предпоследней парой ног. [67] Ни один из видов перипатид не является яйцекладущим, а подавляющее большинство — живородящими. У самок многих живородящих видов развивается плацента, с помощью которой растущий эмбрион получает питательные вещества. Распространение перипатид ограничено тропическим и субтропическим поясом; в частности, они населяют Центральную Америку , север Южной Америки , Габон , Северо-Восточную Индию и Юго-Восточную Азию . [64]
Эволюция
[ редактировать ]
Некоторые окаменелости раннего кембрия имеют поразительное сходство с бархатными червями. Эти окаменелости, известные под общим названием лобоподии , были морскими и представляют собой класс, от которого произошли членистоногие, тихоходки и онихофоры. [70] [71] Возможные окаменелости онихофоров обнаружены в кембрии . [72] ордовик (возможно), [73] силурийский [74] и Пенсильванский [5] [75] периоды.

Исторически все ископаемые Onychophora и lobopodians были объединены в таксон Xenusia , который некоторые авторы далее подразделяли на палеозойские Udeonychophora и мезозойские/третичные Ontonychophora; живые онихофоры были названы Euonychophora. [76] Важно отметить, что лишь немногие из кембрийских окаменелостей имеют черты, которые отчетливо объединяют их с Onychophora; ни один из них не может быть с уверенностью отнесен к коронке онихофора или даже группе стеблей. [77] Возможными исключениями являются Hallucigenia и родственные таксоны, такие как Collinsium ciliosum , которые имеют когти, явно похожие на онихофораны. [70] Неясно, когда произошел переход к наземному существованию, но считается правдоподобным, что он произошел между ордовиком и поздним силурием — примерно 490–430 миллионов лет назад — через приливную зону. [24]
Низкий потенциал сохранности неминерализованных онихофор означает, что у них скудная летопись окаменелостей. лобоподия Helenodora из каменноугольного периода Северной Америки является представителем Onychophora. Предполагается, что [78] но другие исследования считают его более близким родственником других лобоподий. Было высказано предположение, что окаменелость позднего карбона из Монсо-ле-Мин , Франция, Antennipatus имеет явное сходство с онихофорами, вероятно, с первым наземным онихофораном, но ее сохранность не позволяет дифференцировать между ее размещением на стебле или кроне двух существующих семейств или на стволовая группа онихофорана. [5] идентификацией Antennipatus В 2018 году Гирибет и его коллеги согласились с как старейшего онихофорана, которые предположили, что минимальный возраст Antennipatus будет приходиться на гжельскую эпоху около 298,75 миллиона лет назад , и консервативно включили этот таксон для филогенетического анализа онцихофоров на основе о неопределенности его размещения в заказе. [7] В 2021 году Бейкер и его коллеги провели анализ расхождений, используя молекулярное датирование и консервативно рассматривая Antennipatus как онихофоран стволовой группы с минимальным возрастом 300 миллионов лет назад , в результате чего дата расхождения составила 376 миллионов лет назад . для онихофора кроновой группы [79] Представители кроновой группы известны только по янтарю, старейшим из которых является этапа позднего мела , около возрастом Cretoperipatus из бирманского янтаря сеноман-туронского 100-90 млн лет, отнесенный к семейству Peripatidae. [76] Родство янтарных пластинок из кайнозойской эры, таких как Tertiapatus и , Succinipatopsis , которые образуют предполагаемое надсемейство, названное Tertiapatoidea [78] было признано сомнительным другими авторами. [5]
Капризы процесса сохранения могут затруднить интерпретацию окаменелостей. Эксперименты по распаду и уплотнению онихофоры демонстрируют трудности интерпретации окаменелостей; определенные части живых онихофор видны только в определенных условиях:
- Рот может сохраниться, а может и не сохраниться;
- Когти могут быть переориентированы или потеряны;
- Ширина штанин может увеличиваться или уменьшаться; и
- Грязь можно принять за органы. [80]
Что еще более важно, особенности, наблюдаемые в окаменелостях, могут быть артефактами процесса консервации: например, «подплечники» могут быть просто вторым рядом ног, соосно сжатых с телом; ветвящиеся «усики» на самом деле могут возникнуть в результате распада. [80]
Ссылки
[ редактировать ]- ^ Холм, Э.; Диппенаар-Шуман, А. (2010). Членистоногие Южной Африки . ISBN 978-0-7993-4689-3 . [ нужна страница ]
- ^ Протеро, ДР; Бьюэлл, компакт-диск (2007). Эволюция: о чем говорят окаменелости и почему это важно . Нью-Йорк: Издательство Колумбийского университета. п. 193 . ISBN 978-0-231-13962-5 .
- ^ Руперт, Э.Э.; Фокс, РС; Барнс, Р.Д. (2004). Зоология беспозвоночных: функционально-эволюционный подход (7-е изд.). Бельмонт: Томсон-Брукс / Коул. п. 505 . ISBN 978-0-03-025982-1 .
Потому что они напоминают червей с ногами... Внешне они напоминают гусениц, но их сравнивают и со слизняками.
- ^ Пайпер, Росс (2007). «Бархатные черви» . Необыкновенные животные: Энциклопедия любопытных и необычных животных . Гринвуд Пресс. стр. 109–11 . ISBN 978-0-313-33922-6 .
- ^ Jump up to: а б с д и Гарвуд, Рассел Дж.; Эджкомб, Грегори Д.; Шарбонье, Сильвен; Шабар, Доминик; Сотти, Дэниел; Гирибет, Гонсало (2016). «Карбоновые онихофоры из Монсо-ле-Мин, Франция, и натеррестриализация онихофора» . Биология беспозвоночных . 135 (3): 179–190. дои : 10.1111/ivb.12130 . ISSN 1077-8306 . ПМК 5042098 . ПМИД 27708504 .
- ^ Фишелсон, Л. (1978). Зоология . Том. 1 (3-е изд.). Израиль: Издательство Хакибуц Хамеухад. п. 430. [ нужна проверка ]
- ^ Jump up to: а б Гирибет, Гонсало; Бакман-Янг, Ребекка С.; Коста, Криштиану Сампайо; Бейкер, Кейтлин М.; Бенавидес, Лигия Р.; Бранстеттер, Майкл Г.; Дэниэлс, Сэвел Р.; Пинто-да-Роча, Рикардо (2018). «Перипаты» в Еврогондване? - Отсутствие доказательств того, что онихофоры Юго-Восточной Азии ходили по Европе». Систематика беспозвоночных . 32 (4): 840–863. дои : 10.1071/IS18007 .
- ^ Оллвуд, Джулия; Глисон, Дайанна; Майер, Георг; Дэниелс, Савел; Беггс, Жаклин Р.; Бакли, Томас Р. (2010). «Поддержка викариантного происхождения новозеландской онихофоры». Журнал биогеографии . 37 (4): 669–681. дои : 10.1111/j.1365-2699.2009.02233.x . S2CID 55395265 .
- ^ Морера-Бренес, Б.; Монж-Нахера, Дж. (2010). «Новый гигантский вид плацентарных червей и механизм, с помощью которого онихофоры плетут свои сети (Onychophora: Peripatidae)». Обзор тропической биологии . 58 (4): 1127–1142. arXiv : 1511.00983 . дои : 10.15517/rbt.v58i4.5398 . ПМИД 21246983 .
- ^ Jump up to: а б с Ян, Цзе; Ортега-Эрнандес, Хавьер; Гербер, Сильвен; Баттерфилд, Николас Дж.; Хоу, Джин-бо; Лан, Тиан; Чжан, Си-гуан (14 июля 2015 г.). «Сверхбронированный лобопод из кембрия Китая и раннее неравенство в эволюции Onychophora» . Труды Национальной академии наук . 112 (28): 8678–8683. Бибкод : 2015PNAS..112.8678Y . дои : 10.1073/pnas.1505596112 . ISSN 0027-8424 . ПМК 4507230 . PMID 26124122 – через ResearchGate.
- ^ Янссен, Р.; Йоргенсен, М.; Лагебро, Л.; Бадд, GE (18 марта 2015 г.). «Судьба и природа рото-анальной борозды онихофора и ее вклад в бластопор» . Труды Королевского общества B: Биологические науки . 282 (1805): 20142628. doi : 10.1098/rspb.2014.2628 . ПМК 4389607 . ПМИД 25788603 .
- ^ Jump up to: а б с Будро, Х. Брюс (1979). Филогения членистоногих с особым упором на насекомых . Уайли. ISBN 9780471042907 .
- ^ «cruralis/crurale, cruralis M» . Латынь – это просто . Питер Вальдерт . Проверено 21 февраля 2021 г.
- ^ «Кокс, тазики, [ф]» . Латынь – это просто . Питер Вальдерт . Проверено 21 февраля 2021 г.
- ^ Jump up to: а б Смит, Мартин Р.; Ортега-Эрнандес, Хавьер (октябрь 2014 г.). « Когти галлюцигении , похожие на онихофоры, и случай Tactopoda» (PDF) . Природа . 514 (7522): 363–6. Бибкод : 2014Natur.514..363S . дои : 10.1038/nature13576 . ПМИД 25132546 . S2CID 205239797 . Архивировано (PDF) из оригинала 10 октября 2022 г.
- ^ Эрикссон, Бо Йоаким; Тейт, Ноэль Н.; Бадд, Грэм Э.; Янссен, Ральф; Акам, Майкл (сентябрь 2010 г.). «Формирование головы и экспрессия гена Hox у онихофора и его значение для проблемы головы членистоногих» . Гены развития и эволюция . 220 (3–4): 117–22. дои : 10.1007/s00427-010-0329-1 . ПМИД 20567844 . S2CID 6755763 .
- ^ Jump up to: а б с д и ж г Майер, Г.; Оливейра, Исландия; Баер, А.; Хаммель, Ю.; Галлант, Дж.; Хохберг, Р. (2015). «Поимка добычи, питание и функциональная анатомия челюстей бархатных червей (Onychophora)» . Интегративная и сравнительная биология . 55 (2): 217–227. дои : 10.1093/icb/icv004 . ПМИД 25829018 .
- ^ Эрикссон, Б. Йоаким; Тейт, Ноэль Н.; Бадд, Грэм Э. (январь 2003 г.). «Развитие головы у онихофора Euperipatoides kanangrensis с особым упором на центральную нервную систему» . Журнал морфологии . 255 (1): 1–23. дои : 10.1002/jmor.10034 . ПМИД 12420318 . S2CID 42865895 .
- ^ Jump up to: а б Райт, Джонатан С.; Люк, Барбара М. (1989). «Ультраструктурные и гистохимические исследования покровов перипатуса ». Ткани и клетки . 21 (4): 605–25. дои : 10.1016/0040-8166(89)90012-8 . ПМИД 18620280 .
- ^ Морера-Бренес, Б.; Монж-Наджера, Дж. (2010). «Новый гигантский вид плацентарных червей и механизм, с помощью которого онихофоры плетут свои сети (onychophora: Peripatidae)» . Обзор тропической биологии . 58 : 1127–1142. arXiv : 1511.00983 . Архивировано из оригинала 02 апреля 2015 г.
- ^ Конча, Андрес; Мелладо, Паула; Морера-Бренес, Б.; Сампайо Коста, Криштиану; Махадеван, Л.; Монге-Нахера, Хулиан (17 марта 2015 г.). «Колебания струи слизи бархатного червя вследствие пассивной гидродинамической неустойчивости» . Природные коммуникации . 6 : 6292. Бибкод : 2015NatCo...6.6292C . дои : 10.1038/ncomms7292 . ПМЦ 4382676 . ПМИД 25780995 .
- ^ Кампилья, Сильвия; Лаваллар, Роджер (1990). «О шелушении при рождении и в периоде между линькой беременных и молодых Peripatus acacioi (Onycophora, Peripatidae)» . В Минелли, Алессандро (ред.). Материалы 7-го Международного конгресса мириаподологов . БРИЛЛ. стр. 461–4. ISBN 978-90-04-08972-3 .
- ^ Маас, Андреас; Майер, Георг; Кристенсен, Рейнхардт М.; Валошек, Дитер (декабрь 2007 г.). «Кембрийский микролобопод и эволюция передвижения и размножения членистоногих». Китайский научный бюллетень . 52 (24): 3385–92. Бибкод : 2007ЧСБу..52.3385М . дои : 10.1007/s11434-007-0515-3 . S2CID 83993887 .
- ^ Jump up to: а б Монж-Наджера, Джулиан (май 1995 г.). «Филогения, биогеография и репродуктивные тенденции онихофоры». Зоологический журнал Линнеевского общества . 114 (1): 21–60. дои : 10.1111/j.1096-3642.1995.tb00111.x .
- ^ Мэнтон, С.М. (20 июня 1949 г.). «Исследования онихофор. VII. Ранние эмбриональные стадии Peripatopsis и некоторые общие соображения, касающиеся морфологии и филогении членистоногих» . Философские труды Лондонского королевского общества. Серия Б, Биологические науки . 233 (606): 483–580. Бибкод : 1949РСТБ.233..483М . дои : 10.1098/rstb.1949.0003 . JSTOR 92441 .
- ^ «Бархатный червь» . Австралийский музей . 18 января 2022 г. . Проверено 17 февраля 2024 г.
- ^ «Что такое бархатный червь?» . Журнал BBC Science Focus . 5 июля 2022 г. Проверено 17 февраля 2024 г.
- ^ Оценка генома онихофорана: повторы, интроны и расширение семейства генов способствуют гигантизму генома у Epiperipatus Broadwayi
- ^ Холт, Джек (7 февраля 2010 г.). «Знакомство с онихофорами» . Протостомы > Ecdysozoa > Onychophora. Биол. 202: Систематическая биология. Селинсгроув, Пенсильвания: Университет Саскуэханны. Архивировано из оригинала 5 октября 2022 года . Проверено 1 ноября 2022 г.
Выделительная система: пареные метанефридии, открывающиеся собственными протоками на каждом внутреннем «сегменте». Передние нефридии модифицированы в слюнные железы; задние нефридии модифицированы в гонопоры.
- ^ Jump up to: а б с Майер, Георг (декабрь 2006 г.). «Строение и развитие онихофорных глаз: какой предковый орган зрения у членистоногих?». Строение и развитие членистоногих . 35 (4): 231–45. дои : 10.1016/j.asd.2006.06.003 . ПМИД 18089073 .
- ^ Эрикссон, Дж. (2003). Эволюция и развитие головы и нервной системы онихофора . Acta Universitatis Upsaliensis. ISBN 978-91-554-5613-9 . [ нужна страница ]
- ^ Оливейра, Иво де Сена; Франке, Франциска Анни; Геринг, Ларс; Шаффер, Стефан; Роуэлл, Дэвид М.; Век-Хейманн, Андреас; Монге-Нахера, Хулиан; Морера-Бренес, Берналь; Майер, Георг (17 декабря 2012 г.). «Неизведанное разнообразие признаков онихофоры (бархатных червей): сравнительное исследование трех видов перипатид» . ПЛОС ОДИН . 7 (12): е51220. Бибкод : 2012PLoSO...751220O . дои : 10.1371/journal.pone.0051220 . ISSN 1932-6203 . ПМЦ 3524137 . ПМИД 23284667 .
- ^ «Бархатный червь» . Австралийский музей .
- ^ Кемп, С. (1913). «Предварительная заметка о новом роде Onychophora с северо-восточной границы Индии» . Записи Индийского музея . 9 : 241–242.
- ^ Кемп, С. (1914). «Онихофора. Зоологические итоги Аборской экспедиции, 1911–1912 гг.» . Записи Индийского музея . 8 : 471–492. дои : 10.5962/bhl.part.1194 . S2CID 88237018 .
- ^ Оливейра, Иво де Сена; Франке, Франциска Анни; Геринг, Ларс; Шаффер, Стефан; Роуэлл, Дэвид М.; Век-Хейманн, Андреас; Монге-Нахера, Хулиан; Морера-Бренес, Берналь; Майер, Георг (17 декабря 2012 г.). «Неизведанное разнообразие признаков Onychophora (бархатных червей): сравнительное исследование трех видов перипатид» . ПЛОС ОДИН . 7 (12): е51220. Бибкод : 2012PLoSO...751220O . дои : 10.1371/journal.pone.0051220 . ПМЦ 3524137 . ПМИД 23284667 .
- ^ Пойнар, Джордж (сентябрь 1996 г.). «Ископаемые бархатные черви в балтийском и доминиканском янтаре: эволюция и биогеография онихофор». Наука . 273 (5280): 1370–1. Бибкод : 1996Sci...273.1370P . дои : 10.1126/science.273.5280.1370 . JSTOR 2891411 . S2CID 85373762 .
- ^ Эспинаса, Л.; Гарви, Р.; Эспинаса, Дж.; Фратто, Калифорния; Тейлор, С.; Тулкеридис, Т.; Аддисон, А. (21 января 2015 г.). «Пещерное жилище Онихофоры из лавовой трубки на Галапагосских островах» . Подземная биология . 15 : 1–10. дои : 10.3897/subtbiol.15.8468 .
- ^ «Бархатный червь» . Австралийский музей . Проверено 21 февраля 2024 г.
- ^ Взрыв в вечность и между: Космос . Джон Хасси. 31 июля 2014 г.
- ^ иногда называют «клеем», но в современной научной литературе называют слизью, например Баер, А.; Майер, Г. (октябрь 2012 г.). «Сравнительная анатомия слизистых желез онихофоры (бархатных червей)». Журнал морфологии . 273 (10): 1079–1088. дои : 10.1002/Jmor.20044 . ПМИД 22707384 . S2CID 701697 . , Баер, А.; Де Сена Оливейра, И.; Штайнхаген, М.; Бек-Сикингер, AG; Майер, Г. (2014). «Профилирование белков слизи: неинвазивный инструмент для идентификации видов Onychophora (бархатных червей)» . Журнал зоологической систематики и эволюционных исследований . 52 (4): 265–272. дои : 10.1111/jzs.12070 .
- ^ Баер, Александр; Майер, Георг (октябрь 2012 г.). «Сравнительная анатомия слизистых желез онихофоры (бархатных червей)». Журнал морфологии . 273 (10): 1079–88. дои : 10.1002/Jmor.20044 . ПМИД 22707384 . S2CID 701697 .
- ^ Jump up to: а б с Конча, Андрес; Мелладо, Паула; Морера-Бренес, Берналь; Сампайо-Коста, Криштиану; Махадеван, Л.; Монге-Нахера, Хулиан (март 2015 г.). «Колебания струи слизи бархатного червя вследствие пассивной гидродинамической неустойчивости» . Природные коммуникации . 6 : 6292. Бибкод : 2015NatCo...6.6292C . дои : 10.1038/ncomms7292 . ПМЦ 4382676 . ПМИД 25780995 .
- ^ Морера-Бренес, Берналь; Монге-Нахера, Хулиан (декабрь 2010 г.). «Новый гигантский вид плацентарных червей и механизм, с помощью которого онихофоры плетут свои сети (onychophora: Peripatidae)». Revista Biologia Tropical . 58 : 1127–1142. arXiv : 1511.00983 .
- ^ Jump up to: а б с д и ж г час Бенкендорф, Кирстен; Бердмор, Кейт; Гули, Эндрю А; Пакер, Николь Х; Тейт, Ноэль Н. (декабрь 1999 г.). «Характеристика секреции слизистых желез перипатуса Euperipatoides kanangrensis (Onychophora: Peripatopsidae)». Сравнительная биохимия и физиология Б . 124 (4): 457–65. дои : 10.1016/S0305-0491(99)00145-5 .
- ^ Холм, Эрик; Диппенаар-Шуман, Анси (2010). Путеводитель по Гого . Издательство ЛАПА. ISBN 978-0-7993-4689-3 . [ нужна страница ]
- ^ Хармер, Сидни Фредерик; Шипли, Артур Эверетт; и др. (1922). Перипаты, Многоножки, Насекомые . Кембриджская естественная история. Том. 5. Компания Макмиллан. [ нужна страница ]
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Рид, В.М. Сент-Джей; Хьюз, Р.Н. (22 мая 1987 г.). «Пищевое поведение и выбор добычи Macroperipatus torquatus (Onychophora)». Труды Лондонского королевского общества. Серия Б, Биологические науки . 230 (1261): 483–506. Бибкод : 1987РСПСБ.230..483Р . дои : 10.1098/rspb.1987.0030 . JSTOR 36199 . S2CID 83545214 .
- ^ Мозли, HN (1874 г.). «О строении и развитии Peripatus capensis» . Философские труды Лондонского королевского общества . 164 : 757–82. Бибкод : 1874RSPT..164..757M . дои : 10.1098/rstl.1874.0022 . JSTOR 109116 .
- ^ Зелинска, Тереза (2004). «Биологические аспекты передвижения» . В Пфайффере, Фридрих; Зелинска, Тереза (ред.). Ходьба: биологические и технологические аспекты . стр. 1–29. дои : 10.1007/978-3-7091-2772-8_1 . ISBN 978-3-211-22134-1 .
- ^ Jump up to: а б Мэнтон, С.М. (1950). «Эволюция механизмов передвижения членистоногих. Часть I. Передвижение перипатуса» . Журнал Лондонского Линнеевского общества, Зоология . 41 (282): 529–570. дои : 10.1111/j.1096-3642.1950.tb01699.x .
- ^ Jump up to: а б с д и ж г час я дж к л м Рейнхард, Джудит; Роуэлл, Дэвид М. (сентябрь 2005 г.). «Социальное поведение австралийского бархатного червя Euperipatoides rowelli (Onychophora: Peripatopsidae)». Журнал зоологии . 267 (1): 1–7. дои : 10.1017/S0952836905007090 .
- ^ Jump up to: а б Диас, Сидклей К.; Ло-Ман-Хунг, Нэнси Ф. (апрель 2009 г.). «Первая запись об онихофоране (Onychophora, Peripatidae), питающемся пауком-терафосидом (Araneae, Theraphosidae)» . Журнал арахнологии . 37 (1): 116–7. дои : 10.1636/ST08-20.1 . S2CID 86820446 .
- ^ Рид, В.М. Сент-Джей (июль 1988 г.). «Онихофора Тринидада, Тобаго и Малых Антильских островов». Зоологический журнал Линнеевского общества . 93 (3): 225–57. дои : 10.1111/j.1096-3642.1988.tb01362.x .
- ^ Смит, Дж., и Джонс, М. (2022). Материнская забота в Peripatus: доказательства до сих пор неизвестной репродуктивной стратегии онихофоров. Журнал зоологии, 301 (2), 125–134.
- ^ Уокер, Мюриэл Х.; Тейт, Ноэль Н. (декабрь 2004 г.). «Исследование эмбрионального развития и репродуктивного цикла яйцеживородящих австралийских Onychophora (Peripatopsidae)». Журнал зоологии . 264 (4): 333–54. дои : 10.1017/S0952836904005837 .
- ^ Монгенахера, Дж.; Барриентос, З.; Агилар, Ф. (1993). «Поведение Epiperipatus-biolleyi (Onychophora, Peripatidae) в лабораторных условиях». Обзор тропической биологии . 41 (3А): 689–696.
- ^ Эспинаса, Луис; Гарви, Радха; Эспинаса, Хорди; Фратто, Кристина; Тейлор, Стивен; Тулкеридис, Теофилос; Аддисон, Аарон (21 января 2015 г.). «Пещерное жилище Онихофора из лавовой трубки на Галапагосских островах. Пещерное жилище Онихофора из лавовой трубки на Галапагосских островах» . Подземная биология . 15 : 1–10. дои : 10.3897/subtbiol.15.8468 .
- ^ «Красный список видов, находящихся под угрозой исчезновения» МСОП . Красный список исчезающих видов МСОП .
- ^ Jump up to: а б Нью, Тим Р. (1995). «Онихофора в сохранении беспозвоночных: приоритеты, практика и перспективы». Зоологический журнал Линнеевского общества . 114 (1): 77–89. дои : 10.1006/zjls.1995.0017 .
- ^ Подсядловский Л.; Брабанд. А.; Майер, Г. (январь 2008 г.). «Полный митохондриальный геном Onychophoran Epiperipatus biolleyi раскрывает уникальный набор транспортных РНК и обеспечивает дополнительную поддержку гипотезы Ecdysozoa» . Молекулярная биология и эволюция . 25 (1): 42–51. дои : 10.1093/molbev/msm223 . ПМИД 17934206 .
- ^ Штраусфельд, Николас Дж.; Штраусфельд, Камилла Мок; Лозель, Руди; Роуэлл, Дэвид; Стоу, Салли (август 2006 г.). «Филогения членистоногих: организация мозга онихофоров предполагает архаичные отношения с линией ствола хелицератов» . Труды: Биологические науки . 273 (1596): 1857–1866. дои : 10.1098/rspb.2006.3536 . ПМЦ 1634797 . ПМИД 16822744 .
- ^ Чжан, Чжи-Цян (2011). Биоразнообразие животных: введение в классификацию более высокого уровня и таксономическое богатство (PDF) . Том. 3148. стр. 7–12. дои : 10.11646/zootaxa.3148.1.3 . ISBN 978-1-86977-849-1 . Архивировано (PDF) из оригинала 10 октября 2022 г.
{{cite book}}:|journal=игнорируется ( помогите ) - ^ Jump up to: а б с Оливейра, Иво де Сена; Рид, В. Морли Сент-Джей; Майер, Георг (2012). «Всемирный контрольный список онихофор (бархатных червей) с примечаниями по номенклатуре и статусу названий» . ZooKeys (211): 1–70. дои : 10.3897/zookeys.211.3463 . ПМЦ 3426840 . ПМИД 22930648 .
- ^ Монге-Нахера, Хулиан (1994). «Репродуктивные тенденции, тип среды обитания и особенности тела бархатных червей (Onychophora)» . Обзор тропической биологии . 42 (3): 611–622. ISSN 2215-2075 .
- ^ Седжвик, А. (1910). «Перипат папуенсис» . Природа . 83 (2117): 369–370. Бибкод : 1910Natur..83..369S . дои : 10.1038/083369b0 . ISSN 1476-4687 . S2CID 45711430 .
- ^ Jump up to: а б с д Гирибет, Гонсало; Эджкомб, Грегори Д. (3 марта 2020 г.). «30. Онихофора» . В Гирибете, Гонсало; Эджкомб, Грегори Д. (ред.). Древо жизни беспозвоночных . Издательство Принстонского университета. дои : 10.1515/9780691197067-032 . ISBN 978-0-691-19706-7 . S2CID 240645062 ; глава дои : 10.1515/9780691197067-032 ; книга дои : 10.1515/9780691197067 .
- ^ Jump up to: а б Майер, Георг (5 апреля 2007 г.). «Metaperipatus inae sp. nov. (Onychophora: Peripatopsidae) из Чили с новым типом яичников и кожным оплодотворением» . Зоотакса . 1440 (1): 21–37. дои : 10.11646/zootaxa.1440.1.2 . ISSN 1175-5334 – через ResearchGate.
- ^ Кемп, Стэнли (1914). «Онихофора» . Записи Индийского музея . 8 : 471–492. дои : 10.5962/bhl.part.1194 . S2CID 88237018 – через Библиотеку наследия биоразнообразия.
- ^ Jump up to: а б Смит, MR; Ортега Эрнандес, Дж. (2014). « Когти галлюцигении , похожие на онихофоры, и случай Tactopoda» . Природа . 514 (7522): 363–366. Бибкод : 2014Natur.514..363S . дои : 10.1038/nature13576 . ПМИД 25132546 . S2CID 205239797 .
- ^ Бадд, Грэм Э. (сентябрь – октябрь 2001 г.). «Почему членистоногие сегментированы?». Эволюция и развитие . 3 (5): 332–342. дои : 10.1046/j.1525-142X.2001.01041.x . ПМИД 11710765 . S2CID 37935884 .
- ^ Бергстрем, Ян; Хоу, Сиань Гуан (2001). «Кембрийские онихофоры или ксенусианцы». Зоологический вестник . 240 (3–4): 237–245. дои : 10.1078/0044-5231-00031 .
- ^ Ван Рой, Питер; Орр, Патрик Дж.; Боттинг, Джозеф П.; Мьюир, Люси А.; Винтер, Якоб; Лефевр, Бертран; эль Харири, Хадиджа; Бриггс, Дерек Э.Г. (май 2010 г.). «Ордовикская фауна типа сланцев Берджесс». Природа . 465 (7295): 215–218. Бибкод : 2010Natur.465..215V . дои : 10.1038/nature09038 . ПМИД 20463737 . S2CID 4313285 .
- ^ фон Биттер, Питер Х.; Пурнелл, Марк А.; Тетро, Денис К.; Стотт, Кристофер А. (2007). «Eramosa Lagerstätte — исключительно сохранившиеся мягкотелые биоты с мелководными ракушками и биотурбирующими организмами (силурийский период, Онтарио, Канада)». Геология . 35 (10): 879–882. Бибкод : 2007Гео....35..879В . дои : 10.1130/G23894A.1 .
- ^ Томпсон, Ида; Джонс, Дуглас С. (май 1980 г.). «Возможный онихофоран из пластов Мейзон-Крик в средней Пенсильвании в Северном Иллинойсе». Журнал палеонтологии . 54 (3): 588–596. JSTOR 1304204 .
- ^ Jump up to: а б Гримальди, Дэвид А.; Энгель, Майкл С.; Насимбене, Пол К. (март 2002 г.). «Ископаемый меловой янтарь из Мьянмы (Бирмы): его повторное открытие, биотическое разнообразие и палеонтологическое значение» . Новитаты Американского музея (3361): 1–71. doi : 10.1206/0003-0082(2002)361<0001:FCAFMB>2.0.CO;2 . hdl : 2246/2914 . S2CID 53645124 .
- ^ Бадд, Грэм Э. (2001). «Тихоходки как« членистоногие стволовой группы »: данные кембрийской фауны». Зоологический обзор . 240 (3–4): 265–279. дои : 10.1078/0044-5231-00034 .
- ^ Jump up to: а б Пойнар, Джордж (зима 2000 г.). «Ископаемые онихофоры из доминиканского и балтийского янтаря: Tertiapatus dominicanus ng, nsp. (Tertiapatidae n. fam.) и Succinipatopsis balticus ng, n.sp. (Succinipatopsidae n. fam.) с предлагаемой классификацией подтипа Onychophora» . Биология беспозвоночных . 119 (1): 104–109. дои : 10.1111/j.1744-7410.2000.tb00178.x . JSTOR 3227105 .
- ^ Бейкер, Кейтлин М; Бакман-Янг, Ребекка С; Коста, Криштиану С; Гирибет, Гонсало (9 декабря 2021 г.). Ся, Сюйхуа (ред.). «Филогеномный анализ бархатных червей (онихофоры) обнаруживает эволюционное излучение в неотропах» . Молекулярная биология и эволюция . 38 (12): 5391–5404. дои : 10.1093/molbev/msab251 . ISSN 1537-1719 . ПМЦ 8662635 . ПМИД 34427671 .
- ^ Jump up to: а б Монге-Нахера, Хулиан; Сянгуан, Хоу (декабрь 2002 г.). «Экспериментальная тафономия бархатных червей (Onychophora) и значение кембрийской модели «взрыва, неравенства и уничтожения»» . Обзор тропической биологии . 50 (3–4): 1133–1138. ПМИД 12947596 .
Внешние ссылки
[ редактировать ]- Фликр
- И.С. Оливейра; Л. Геринг; Г. Майер. «Сайт Онихофоры» . Архивировано из оригинала 14 июня 2017 года.
- Youtube , Склизкий и смертоносный бархатный червь , Смитсоновский канал
- . Британская энциклопедия (11-е изд.). 1911.
- Перипат обсуждается на программе RNZ Critter of the Week , 13 ноября 2015 г.