Лобоподии
| Лобоподии Временной диапазон: Потомки таксонов Onychophora , Tardigrada и Euarthropoda доживают до недавних, возможных эдиакарских ихноокаменелостей. [2] | |
|---|---|
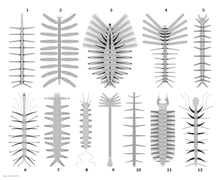 | |
| Реконструкция различных лобоподий. 1: Microdictyon sinicum , 2: Diania cactiformis , 3: monstruosus , 4: Luolishania longicruris , 5: Onychodictyonferox Facivermis , 6: Hallucigenia sparsa , 7: Aysheaia pedunculata , 8: Antennacanthopodia gracilis , 9: , Collinsovermis 10: Paucipodia inermis , 11: Jianshanopodia Decora , 12: Hallucigenia fortis. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Subkingdom: | Эуметазоа |
| Клэйд : | ПараХоксозоа |
| Клэйд : | билатерия |
| Клэйд : | Нефрозоа |
| (без рейтинга): | протостомия |
| Супертип: | Экдисозоа |
| (без рейтинга): | панартроподы |
| Тип: | † «Лобоподия» Снодграсс 1938 год |
| Группы включены | |
| |
| Кладистически включенные, но традиционно исключенные таксоны | |
Кронная группа Euarthropoda | |
| Синонимы | |
| |
Лобоподии являются членами неформальной группы Lobopodia. [3] (от греческого , что означает «тупые ступни»), или формально установленный тип Lobopoda Cavalier-Smith (1998). [4] Это панартроподы с короткими ногами, называемые лобоподами . [5] термин, который также может использоваться как общее название этой группы. [6] [7] Хотя определения лобоподий могут различаться в разных источниках, [8] обычно это относится к группе мягкотелых морских червеобразных ископаемых панартропод, таких как Aysheaia и Hallucigenia . Однако другие роды, такие как Kerygmachela и Pambdelurion (имеющие сходные с другими группами черты), часто называют «жаберными лобоподиями». [9] [5] [10]
Самые старые почти полные ископаемые лобоподии относятся к нижнему кембрию ; некоторые также известны из ордовика , силура и каменноугольного периода лагерштеттенов . [11] [12] [13] Некоторые из них имеют закаленные когти, пластины или шипы, которые обычно сохраняются в виде углеродистых или минерализованных микроокаменелостей в кембрийских слоях. [14] [15] Эта группировка считается парафилетической , поскольку считается, что три ныне живущие группы панартропод ( Arthropoda , Tardigrada и Onychophora ) произошли от предков лобоподов.
Определения
[ редактировать ]
Концепция лобоподиан варьируется от автора к автору. [8] Его наиболее общий смысл относится к набору преимущественно кембрийских червеобразных таксонов панартропод, содержащих лобоподы, например, Aysheaia , Hallucigenia и Xenusion , которые традиционно объединялись как «Xenusians» или «Xenusiids» (класс Xenusia). [16] [17] [18] Некоторые роды динокаридид , такие как Opabinia , Pambdelurion и Kerygmachela , также можно рассматривать как лобоподий. [19] [10] иногда их называют более конкретно «жаберными лобоподиями» или «жаберными лобоподиями». [20] Это традиционное, неформальное использование слова «Lobopodia» рассматривает его как эволюционную ступень , включающую только вымерших панартропод у основания кроны панартропод. Коронные панартроподы включают три существующих типа панартропод — Onychophora (бархатные черви), Tardigrada (водяные медведи) и Arthropoda (членистоногие), а также их самого недавнего общего предка и всех его потомков. Таким образом, в этом смысле лобоподии состоят из различных базальных панартропод. [5] [21] [22] [8] [23] [24] Это соответствует букве «А» на изображении слева.
Альтернативное, более широкое определение Lobopodia также включает Onychophora и Tardigrada. [16] [19] [4] два современных типа панартропод, которые все еще имеют лобоподовые конечности. Это определение, соответствующее «С», является морфологическим и зависит от внешнего сходства придатков («лобоподов»). Таким образом, он является парафилетическим , за исключением Euarthropods, которые являются потомками некоторых Lobopodians, на основании их сильно различающейся морфологии конечностей. [5] «Лобоподия» также использовалась для обозначения предполагаемой сестринской клады Arthropoda, состоящей из дошедших до нас Onychophora и Tardigrada, а также их самого недавнего общего предка и всех его потомков. Это определение делает Lobopodia монофилетическим таксоном, если оно действительно действительно (то есть, если тихоходки и онихофоры ближе друг к другу, чем любой из них к членистоногим), но исключает все таксоны линии Euarthropod, традиционно считающиеся лобоподами. Однако его достоверность сомнительна, поскольку существует ряд гипотез относительно внутренней филогении Panarthropoda. [25] Самое широкое определение рассматривает Lobopodia как монофилетический супертип, эквивалентный объему Panarthropoda по . Согласно этому определению, обозначенному буквой «D» на изображении, лобоподии больше не рассматриваются как эволюционная ступень, а как клада, содержащая не только ранние, внешне «лобоподические» формы, но и всех их потомков, включая современных панартроподов. [26] [6]
Лобоподии исторически иногда включали Pentastomida , [27] группа паразитических панартропод, которые традиционно считались уникальным типом . [28] [29] и анатомические исследования показали, но последующие филогеномные что это высокоспециализированный таксон ракообразных . [30] [31] [32] [33] [34]
Представитель таксонов
[ редактировать ]

Наиболее известные роды включают Aysheaia , которая была обнаружена в канадских сланцах Берджесс , и Hallucigenia , известные как в сланцах Чэньцзян Маотяньшань, так и в сланцах Берджесс. Aysheaia pedunculata имеет морфологию, очевидно, базовую для лобоподий. [21] - например, значительно кольчатая кутикула, терминальное ротовое отверстие, специализированные почти лобные придатки и короткие лобоподы с терминальными когтями. Hallucigenia sparsa известна своей сложной историей интерпретации — первоначально она была реконструирована с длинными, похожими на ходули ногами и загадочными мясистыми спинными выступами, и долгое время считалась ярким примером того, как природа экспериментировала с самым разнообразным и причудливым телом. конструкции во время кембрия. [35] Однако дальнейшие открытия показали, что эта реконструкция поместила животное вверх тормашками: интерпретация «ходулей» как спинных шипов прояснила, что мясистые «спинные» выступы на самом деле были удлиненными лобоподами. Более поздняя реконструкция даже поменяла местами передний и задний концы животного: выяснилось, что выпуклый отпечаток, ранее считавшийся головой, на самом деле был содержимым кишечника, выбрасываемым из заднего прохода. [14] [22]
Микродиктион — еще один харизматичный и особый род лобоподий, напоминающий Hallucigenia , но вместо шипов он нес пары сетчатых пластинок, которые часто встречаются разрозненными и известны как пример небольших ракушечных окаменелостей (SSF). Xenusion имеет самую старую летопись окаменелостей среди описанных лобоподий, которая может восходить к кембрийскому этапу 2 . [16] [10] Луолишания — яркий пример лобоподий с несколькими парами специализированных придатков. [36] Жаберные лобоподии Kerygmachela и Pambdelurion проливают свет на взаимоотношения между лобоподами и членистоногими , поскольку они имеют как сходство, так и характеристики лобоподий, связанные со стеблевой группой членистоногих. [20] [26]
Морфология
[ редактировать ]
 Максимальный размер трех видов Hallucigenia (сверху: H. fortis , H. hongmeia и H. sparsa ) в масштабе.
Максимальный размер трех видов Hallucigenia (сверху: H. fortis , H. hongmeia и H. sparsa ) в масштабе. Окаменелости Xenusion , лобопода, который мог вырасти до 20 сантиметров.
Окаменелости Xenusion , лобопода, который мог вырасти до 20 сантиметров.


Большинство лобоподий имели длину всего несколько сантиметров, тогда как некоторые роды вырастали до более чем 20 сантиметров. [7] Их тела кольчатые , хотя наличие кольчатых колец может различаться в зависимости от положения или таксона, и иногда их трудно различить из-за их близкого расположения и низкого рельефа на ископаемых материалах. [37] Тело и придатки округлые в поперечном сечении. [37]
Голова
[ редактировать ]

Из-за обычно плохой сохранности подробные реконструкции области головы доступны лишь для нескольких видов лобоподий. [38] [22] Голова лобоподия более или менее выпуклая. [5] и иногда обладает парой предглазных, предположительно протоцеребральных [23] придатки - например, первичные усики [39] [36] [23] [40] или хорошо развитые лобные придатки, [41] [19] [42] [7] [5] которые индивидуализированы из лобоподов туловища [23] [43] (за исключением Antennacanthopodia , у которых имеются две пары головных придатков вместо одной [39] ). Ротовой аппарат может состоять из рядов зубов. [37] [22] [42] [7] [44] или конический хоботок. [38] [5] [45] Глаза могут быть представлены одним глазком или многочисленными. [46] пары простых глазков, [5] как было показано в Луолишании [36] (= Миралуолишания [46] [47] ), Оватовермис , [45] Ониходикцион , [38] Галлюцигения , [22] Фацивермис , [47] и, в меньшей степени, Айшеайя . [38] Однако у жаберных лобоподий, таких как Kerygmachela , глаза представляют собой относительно сложные светоотражающие пятна. [48] это могло иметь сложный характер. [49]
Туловище и лобоподы
[ редактировать ]

Туловище удлиненное и состоит из многочисленных сегментов тела ( сомитов ), каждый из которых несет пару ног, называемых лобоподами. [5] или лобоподические конечности. [19] Границы сегментов не так внешне значимы, как у членистоногих, хотя у некоторых видов они обозначены гетерономными кольцами (т. е. чередованием плотности колец, соответствующим положению границ сегментов). [50] [36] [21] Сегменты туловища могут нести другие внешние, соответствующие сегментам структуры, такие как узлы (например, Hadranax , [6] Керигмачела [19] ), сосочки (например, Onychodictyon [38] ), шиповидные/пластинчатые склериты (например, панцирные лобоподии [5] ) или боковые створки (например, жаберные лобоподии [19] [44] ). Туловище может заканчиваться парой лобоподов (например, Aysheaia , Hallucigenia sparsa ). [22] или хвостообразное расширение (например, Paucipodia , Siberion , Jianshanopodia ). [50] [37] [7] [27]
Лобоподы гибкие, имеют слабоконическую форму, сужаются от тела к кончикам, которые могут [21] [5] или не может [39] [51] [10] медвежьи когти. Когти, если они есть, представляют собой закаленные структуры, по форме напоминающие крючок или слегка изогнутый шип. [37] [52] [36] [21] [5] Лобоподы с клешнями обычно имеют две клешни, но известны и одиночные клешни (например, задние лобоподы луолишаниид [36] [45] [40] ), а также более двух (например, три у Tritonychus , [53] семь в Айшеайе [41] ) в зависимости от его сегментной или таксономической принадлежности. [21] У некоторых родов лобоподы несут дополнительные структуры, такие как шипы (например, Diania [51] ), мясистые выросты (например, Onychodictyon [38] ) или бугорки (например, Jianshanopodia [7] ). не обнаружено Признаков членистоногих (развитие затвердевшего экзоскелета и сегментарного деления на придатках панартропод) у известных представителей лобоподий, даже у тех, кто принадлежит к стеблевой группе членистоногих (например, жаберные лобоподии и сибирииды), и есть подозрение на членистоногие на конечности Диании [54] считается неверным толкованием. [51] [10]
Дифференциация (тагмоз) между сомитами туловища почти не происходит, за исключением галлюцигенид и луолишаниид, у которых многочисленные пары передних лобопод значительно тонкие (галлюцигениды) или щетинистые (луолишанииды) в отличие от их задних аналогов. [5] [22] [45] [24] [40]
Внутренние структуры
[ редактировать ]
Кишка лобоподий часто прямая, недифференцированная, [55] и иногда сохраняются в летописи окаменелостей в трех измерениях. У некоторых экземпляров кишечник заполнен осадком. [37] Кишка состоит из центральной трубки, занимающей всю длину туловища лобоподии. [7] который не сильно меняется по ширине - по крайней мере, не систематически. Однако у некоторых групп, особенно у жаберных лобоподий и сибириид, кишечник окружен парами серийно повторяющихся почковидных кишечных дивертикулов (пищеварительных желез). [7] [42] [55] У некоторых экземпляров части кишки лобоподий могут сохраняться в трех измерениях. Это не может быть результатом фосфатизации, которая обычно отвечает за трехмерное сохранение кишечника. [56] потому что содержание фосфатов в кишках менее 1%; в составе присутствуют кварц и мусковит. [37] Кишечник представителя Paucipodia варьирует по ширине, наибольшая ширина приходится на центр тела. Его положение в полости тела фиксировано слабо, поэтому возможна гибкость.

О нервной анатомии лобоподий известно немного из-за скудных и по большей части неоднозначных ископаемых свидетельств. Возможные следы нервной системы обнаружены у Paucipodia , Megadictyon и Antennacanthopodia . [42] [37] [39] Первое и пока единственное подтвержденное свидетельство существования нервных структур лобоподий получено от жаберных лобоподий Kerygmachela в работе Park et al. 2018 — представлен мозг, состоящий только из протоцеребрума (самого переднего церебрального ганглия панартропод ) , который напрямую связан с глазными нервами и лобными придатками, что позволяет предположить протоцеребральное происхождение головы лобоподов, а также всех панартропод . [49]
У некоторых современных экдизозоев, таких как приапулиды и онихофоры , имеется слой самых внешних круговых мышц и слой самых внутренних продольных мышц. У онихофоров имеется также третий, промежуточный, слой переплетенных косых мышц. Мускулатура жаберного лобоподия Pambdelurion имеет аналогичную анатомию. [57] но у лобоподийных Tritonychus наблюдается противоположная картина: самые наружные мышцы являются продольными, а самый внутренний слой состоит из круговых мышц. [53]
Категории
[ редактировать ]В зависимости от внешней морфологии лобопдианы могут относиться к разным категориям - например, к общим червеобразным таксонам, таким как «ксенузиид» или «ксенузиан»; ксенузииды со склеритом в виде «бронированных лобоподий»; и таксоны как с крепкими лобными придатками, так и с боковыми створками, как «жаберные лобоподии». Некоторые из них первоначально были определены в таксономическом смысле (например, класс Xenusia), но ни один из них не был признан монофилетическим в дальнейших исследованиях. [21] [8] [23]
Бронированные лобоподии
[ редактировать ]

Бронированные лобоподии относились к ксенузиидам лобоподиям, которые несли повторяющиеся склериты, такие как шипы или пластинки на туловище (например, Hallucigenia , Microdictyon , Luolishania ) или лобоподам (например, Diania ). Напротив, лобоподий без склеритов можно назвать «небронированными лобоподиями». [39] [10] Функцию склеритов интерпретировали как защитную броню и/или точки прикрепления мышц. [50] [58] [5] В ряде случаев сохранились лишь разрозненные склериты животного, которые представляли собой компонент мелких ракушковых окаменелостей (SSF). [50] [14] предполагалось, что панцирные лобоподии связаны с онихофорами и могут даже представлять собой кладу . В некоторых предыдущих исследованиях [58] но их филогенетическое положение в более поздних исследованиях противоречиво. ( см. текст )
Жаберные лобоподии
[ редактировать ]
Динокаридиды , имеющие сходство с лобоподиями (из-за общих черт, таких как кольчатое кольцо и лобоподии), называются «жаберными лобоподиями». [8] [44] [57] или «жаберные лобоподы». [20] Эти формы имеют пару створок на каждом сегменте туловища, но в остальном не имеют признаков членистоногих, в отличие от более продвинутых динокаридид, таких как Radiodonta , у которых есть крепкие и склеротизированные лобные придатки. Жаберные лобоподии охватывают как минимум четыре рода: Pambdelurion , Kerygmachela , Utahnax и Mobulavermis . [10] [59] Опабиния также может подпадать под эту категорию в более широком смысле. [8] [60] хотя наличие лобоподов в этом роде окончательно не доказано. [61] Omnidens , род, известный только по ротовому аппарату, похожему на памбделурион , также может быть жаберным лобоподием. [44] Створки тела могли функционировать как плавательные придатки, так и жабры. [20] и, возможно, гомологичны дорсальным лоскутам радиодонтов и выходам Euarthropoda . [19] [60] Некоторые до сих пор оспаривают, были ли эти роды настоящими лобоподиями. [62] Однако они широко признаны членистоногими стеблевой группы, базальными по отношению к радиодонтам. [8] [21] [22] [23]
Сиберион и подобные таксоны
[ редактировать ]
Сиберион , Мегадиктион и Цзяньшаноподии можно сгруппировать как сиберииды (отряд Siberiida ), [27] цзяньшаноподианцы [21] или «гигантские лобоподии» [63] по некоторым литературам. Обычно они крупные — длина тела колеблется от 7 [27] и 22 сантиметра [42] (от 2¼ до 8⅔ дюймов) - лобоподии ксенузиид с расширенным туловищем, лобоподами с толстым туловищем без признаков когтей и, в первую очередь, с парой крепких лобных придатков. [8] За исключением, пожалуй, Сибириона , [27] [21] у них также есть пищеварительные железы, как у жаберных лобоподий и базальных эуартропод. [7] [42] [8] [55] Их анатомия представляет собой переходные формы между типичными ксенузиидами и жаберными лобоподиями. [27] в конечном итоге поместив их под самое базальное положение стеблевой группы членистоногих. [7] [42] [8] [23]
Палеоэкология
[ редактировать ]

Лобоподии, возможно, занимали широкий спектр экологических ниш . [5] Хотя у большинства из них были недифференцированные придатки и прямой кишечник, что позволяет предположить простой образ жизни, питающийся отложениями, [5] сложные пищеварительные железы и большие размеры жаберных лобоподий и сибериид позволили бы им потреблять более крупные продукты питания, [5] [55] а их крепкие лобные придатки могут даже указывать на хищнический образ жизни. [7] [55] С другой стороны, луолишанииды, такие как Luolishania и Ovatiovermis, имеют сложные перистые лобоподы, которые, предположительно, образовывали «корзины» для подвешивания или фильтрования . [36] [45] Лобоподы с изогнутыми конечными когтями, возможно, дали некоторым лобоподам способность лазить по веществам. [5]
известно немного О физиологии лобоподий . Есть данные, позволяющие предположить, что лобоподии линяют так же, как и другие таксоны экдизозойных , но очертания и орнаментация затвердевшего склерита не менялись в течение онтогенеза . [50] [15] Жаберные структуры на створках тела жаберных лобоподий и разветвленные отростки на лобоподиях Цзяньшаноподий могут обеспечивать дыхательную функцию ( жабры ). [20] [7] Памбделурион может контролировать движение своих лобоподов подобно онихофорам . [57]
Распределение
[ редактировать ]В кембрийском периоде лобоподы демонстрировали значительную степень биоразнообразия . Из каждого ордовикского и силурийского периодов известно по одному виду. [12] [64] еще несколько известны из каменноугольного периода (Мазон-Крик) — это свидетельствует о нехватке исключительных лагерстаттенов в посткембрийских отложениях.
Филогения
[ редактировать ]
| |||||||||||||||||||||||||||||||||||||||
| Нейтрализованная филогения между лобоподиями и другими Ecdysozoa . таксонами [21] [8] [22] [60] [23] [45] [24] Существующие таксоны панартропод выделены жирным шрифтом . Отношения между общей группой современных типов панартропод не решены. |







Общая филогенетическая интерпретация лобоподий кардинально изменилась с момента их открытия и первого описания. [5] Переназначения основаны не только на новых ископаемых данных, но также на новой эмбриологической , нейроанатомической и геномной (например, экспрессии генов , филогомике ) информации, наблюдаемой у существующих таксонов панартропод . [5] [23] [65]
На основании их явно онихофороподобной морфологии (например, кольчатая кутикула, лобоподический придаток с когтями) первоначально считалось, что лобоподии представляют собой группу палеозойских онихофоров. [66] [17] [18] [67] [5] Эта интерпретация была оспорена после открытия лобоподий с характеристиками членистоногих и тихоходок . [68] предполагая, что сходство между лобоподиями и онихофорами представляет собой более глубокие панартропод наследственные черты ( плезиоморфии ), а не эксклюзивные для онихофоранов характеристики ( синапоморфии ). [24] Например, британский палеонтолог Грэм Бадд рассматривает Lobopodia как базальную ступень, от которой произошли типы Onychophora и Arthropoda, при этом Aysheaia сравнима с предковым планом, а такие формы, как Kerygmachela и Pambdelurion, переход, который через динокаридиды представляют собой привели к строению тела членистоногих. [58] Орнаментация поверхности Айшеайи, если она гомологична склеритам палеосколецид , может представлять собой более глубокое звено, связывающее ее с внешними циклоневральными группами. [58] Многие дальнейшие исследования последовали и расширили эту идею, в целом согласившись с тем, что все три типа панартропод имеют лобоподий в своих стволовых линиях. [5] [21] [22] [23] [45] [24] Таким образом, лобоподии являются парафилетическими и включают в себя последнего общего предка членистоногих, онихофоров и тихоходок. [5]
Членистоногие стеблевой группы
[ редактировать ]По сравнению с другими стволовыми группами панартропод, предположения о лобоподических членах стволовой группы членистоногих относительно последовательны - сиберииды, такие как Megadictyon и Jianshanopodia, занимают самую базальную позицию, жаберные лобоподии Pambdelurion и Kerygmachela разветвляются следующим и, наконец, приводят к кладе, состоящей из Opabinia . Radiodonta и Euarthropoda (членистоногие кроновой группы). [21] [8] [22] [60] [23] [45] [24] Их положение внутри стеблевой группы членистоногих определяется многочисленными планами строения членистоногих и промежуточными формами (например, членистоногими пищеварительными железами, радиодонтоподобными лобными придатками и дорсо-вентральными аппендикулярными структурами, связанными с двуветвистыми придатками членистоногих). [8] [23] Лобоподическое происхождение членистоногих также подтверждено геномными исследованиями существующих таксонов - экспрессия генов подтверждает гомологию между придатками членистоногих и онихофорными лобоподами, что позволяет предположить, что современные менее сегментированные членистоногие придатки произошли от кольчатых лобоподовых конечностей. [43] С другой стороны, первичные усики и лобные придатки лобоподий и динокаридид могут быть гомологичны комплексу верхней губы /гипостомы эуартропод, что подтверждается их протоцеребральным происхождением. [8] [23] [49] и особенности развития верхней губы современных членистоногих. [43] [23]
 Радиодонты стеблевой группы представляют собой членистоногих с жаберными лобоподиеподобными лоскутами тела, членистоногими лобными придатками и на стебельках сложными глазами .
Радиодонты стеблевой группы представляют собой членистоногих с жаберными лобоподиеподобными лоскутами тела, членистоногими лобными придатками и на стебельках сложными глазами . Реставрация Памбделуриона, «жаберного лобопода», родственного членистоногим, имеющего как пары лобопод, так и боковые створки.
Реставрация Памбделуриона, «жаберного лобопода», родственного членистоногим, имеющего как пары лобопод, так и боковые створки.


Первоначально считалось, что Diania , род панцирных лобоподов с толстыми и колючими ногами, относится к группе стволовых членистоногих на основании его явно членистоногих (членистоногих) придатков туловища. [54] Однако такая интерпретация сомнительна, поскольку данные оригинального описания не согласуются с предполагаемыми филогенетическими связями. [69] [70] Дальнейшее повторное обследование даже показало, что подозрение на членистоногие на ногах Диании было неверно истолковано — хотя позвоночник мог затвердеть, оставшаяся кутикула ног Диании была мягкой (не затвердевшей и не склеритизованной), без каких-либо признаков поворотного сустава и артродиальной мембраны, позволяют предположить, что ноги представляют собой лобоподы только с широко расположенными кольцами. [51] [10] Таким образом, повторное исследование в конечном итоге отвергло доказательства членистоногих (склеротизации, сегментации и сочленения) придатков, а также фундаментальные взаимоотношения между Дианией и членистоногими. [51] [10]
Онихофораны стволовой группы
[ редактировать ]
Хотя Antennacanthopodia широко считается онихофораном стволовой группы, [21] [22] [53] [45] [24] [47] положение других родов ксенузиид, которые ранее считались родственными онихофоранам, является спорным - в дальнейших исследованиях большинство из них были предложены как онихофораны стволовой группы. [21] [22] [53] [47] или базальные панартроподы, [45] [24] [40] с несколькими видами ( Aysheaia [45] [24] [40] или Ониходикцион Ферокс [21] [22] ) иногда предполагают, что это тихоходки стеблевой группы. Исследование, проведенное в 2014 году, показало, что Hallucigenia представляют собой онихофораны стволовой группы, основанные на их когтях, которые перекрываются внутренние структуры, напоминающие структуры современных онихофоранов. [21] Эта интерпретация была подвергнута сомнению в более поздних исследованиях, поскольку структуры могут представлять собой плезиоморфию панартропод . [24] [40]
Тихоходки стеблевой группы
[ редактировать ]
Таксоны лобоподий стеблевой группы тихоходок неясны. [5] Айшеайя [45] [24] [40] или Ониходикцион Ферокс [21] [22] Было предложено стать возможным членом на основании большого количества когтей (у Aysheaia ) и/или конечных лобопод с когтями, обращенными вперед (у обоих таксонов). [21] Хотя это и не получило широкого признания, есть даже предположения, что Tardigrada сама по себе представляет собой самого базального панартропода или ветвь между стволовой группой членистоногих. [68] Однако в статье 2023 года луолишанииды оказались ближайшими родственниками тихоходок по различным морфологическим характеристикам. [72]
Панартроподы стеблевой группы
[ редактировать ]Неясно, какие лобоподии представляют собой членов стеблевой группы панартропод, которые разветвились незадолго до появления последнего общего предка современных типов панартропод. Айшеайя , возможно, занимала это положение на основании своей очевидно базовой морфологии; [58] [21] [22] [47] в то время как другие исследования скорее предполагают луолишаниид и галлюцигенид, [45] [24] [40] два таксона лобоподий, которые также были отнесены к онихофорам стволовой группы. [5] [21] [22] [53] [47]
Описанные роды
[ редактировать ]



По состоянию на 2018 год описано более 20 родов лобоподий. [10] описываемые как лобоподии Mureropodia apae и Aysheaia prolata, считаются разрозненными лобными придатками радиодонтов Ископаемые материалы , Caryosyntrips и Stanleycaris соответственно. [73] [74] [75] является предположили, что синонимом Luolishania . Miraluolishania Некоторые исследования [36] [76] [47] Позже выяснилось, что загадочный Facivermis представляет собой узкоспециализированный род лобоподий луолишаниид. [27] [45] [47]
- Ацинокрикус [5]
- Антеннакантоподии [39]
- Айшеайя [41]
- Карботубул [13]
- Кардиодиктион [77]
- Коллинз [78]
- Коллинсовермис [40]
- События [54] [51]
- Энтотирей [79]
- Фацивермис [47]
- Фузуконхарий [80]
- Хадранакс [81]
- Галлюцигения [17] [18] [52] [22]
- Цзяньшаноподия [7]
- Керигмачела [20] [19] [49]
- Ленисамбулатрикс [10]
- Пещера Шанья [18] (= Миралуолишания ) [36] [47]
- Мегадиктион [82] [42]
- Микродиктион [50]
- Мобулавермис [83]
- Омнидены ? [84]
- Ониходикцион [77] [38]
- Орстенотубулус [85]
- Оватовермис [45]
- Памбделурион [26] [57] [44]
- Парвибеллус ? [83]
- Пауциподии [86] [37]
- Квадратапора [80]
- Сибирь [27]
- Танахита [24]
- Тритоних [53]
- Ютанакс [87]
- Ксенюзия [16]
- Юти ? [88]
Ссылки
[ редактировать ]- ^ Хауг, Дж. Т.; Майер, Г.; Хауг, К.; Бриггс, DEG (2012). «Неонихофоровый лобопод каменноугольного периода демонстрирует долгосрочное выживание кембрийского морфотипа» . Современная биология . 22 (18): 1673–1675. дои : 10.1016/j.cub.2012.06.066 . ПМИД 22885062 .
- ^ Чен, Чжэ; Чен, Сян; Чжоу, Чуаньмин; Юань, Сюньлай; Сяо, Шухай (6 июня 2018 г.). «Позднеэдиакарские следы, оставленные двусторонними животными с парными придатками» . Достижения науки . 4 (6). дои : 10.1126/sciadv.aao6691 . hdl : 10919/84444 .
- ^ Снодграсс, RE (1938). «Эволюция кольчатых червей, онихофор и членистоногих» . Разные коллекции Смитсоновского института . 97 (6): 1–159.
- ^ Jump up to: а б Кавальер-Смит, Т. (1998). «Пересмотренная система жизни шести королевств». Биологические обзоры . 73 (3): 203–266. doi : 10.1017/S0006323198005167 (неактивен 26 апреля 2024 г.). ПМИД 9809012 .
{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка ) - ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа Ортега-Эрнандес, Хавьер (05 октября 2015 г.). «Лобоподии» . Современная биология . 25 (19): Р873–Р875. дои : 10.1016/j.cub.2015.07.028 . ISSN 0960-9822 . ПМИД 26439350 . Ошибка цитирования: именованная ссылка «:0» была определена несколько раз с разным содержимым (см. страницу справки ).
- ^ Jump up to: а б с Бадд, Грэм; Пил, Джон (1 декабря 1998 г.). «Новый лобопод Xenusiid из раннекембрийской фауны Sirius Passet Северной Гренландии» . Палеонтология . 41 : 1201–1213.
- ^ Jump up to: а б с д и ж г час я дж к л м Цзянни Лю; Деган Шу; Цзянь Хан; Чжифей Чжан и Синлян Чжан (2006). «Большой лобопод ксенузиид со сложными придатками из нижнего кембрия Chengjiang Lagerstätte» (PDF) . Акта Палеонтол. Пол . 51 (2): 215–222. Архивировано (PDF) из оригинала 10 октября 2022 г. Проверено 9 февраля 2011 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот Ортега-Эрнандес, Хавьер (декабрь 2014 г.). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры Кембриджского философского общества . 91 (1): 255–273. дои : 10.1111/brv.12168 . ISSN 1469-185Х . ПМИД 25528950 . S2CID 7751936 .
- ^ Лю, Цзянни; Данлоп, Джейсон А. (15 марта 2014 г.). «Кембрийские лобоподы: обзор недавнего прогресса в нашем понимании их морфологии и эволюции» . Палеогеография, Палеоклиматология, Палеоэкология . 398 : 4–15. Бибкод : 2014PPP...398....4L . дои : 10.1016/j.palaeo.2013.06.008 . ISSN 0031-0182 .
- ^ Jump up to: а б с д и ж г час я дж к Оу, Цян; Майер, Георг (01 декабря 2018 г.). «Кембрийский небронированный лобопод, † Lenisambulatrix humboldti gen. et sp. nov., по сравнению с новым материалом † Diania cactiformis » . Научные отчеты . 8 (1): 13667. Бибкод : 2018NatSR...813667O . дои : 10.1038/s41598-018-31499-y . ПМК 6147921 . ПМИД 30237414 .
- ^ ван Рой, П.; Орр, ПиДжей; Боттинг, JP; Мьюир, Луизиана; Винтер, Дж.; Лефевр, Б.; Харири, Кентукки; Бриггс, DEG (2010). «Ордовикская фауна типа сланцев Берджесс». Природа . 465 (7295): 215–218. Бибкод : 2010Natur.465..215V . дои : 10.1038/nature09038 . ПМИД 20463737 . S2CID 4313285 .
- ^ Jump up to: а б фон Биттер, PH; Пурнелл, Массачусетс; Тетро, Дания; Стотт, Калифорния (2007). «Eramosa Lagerstätte — исключительно сохранившаяся мягкотелая биота с мелководными ракушками и биотурбирующими организмами (силурийский период, Онтарио, Канада)». Геология . 35 (10): 879. Бибкод : 2007Гео....35..879В . дои : 10.1130/G23894A.1 . S2CID 11561169 .
- ^ Jump up to: а б Хауг, Дж. Т.; Майер, Г.; Хауг, К.; Бриггс, DEG (2012). «Неонихофоровый лобопод каменноугольного периода демонстрирует долгосрочное выживание кембрийского морфотипа» . Современная биология . 22 (18): 1673–1675. дои : 10.1016/j.cub.2012.06.066 . ПМИД 22885062 .
- ^ Jump up to: а б с Карон, Ж.-Б.; Смит, MR; Харви, THP (2013). «За сланцами Бёрджесс: кембрийские микроокаменелости отслеживают взлет и падение галлюцигенидных лобоподий» . Труды Королевского общества B: Биологические науки . 280 (1767): 20131613. doi : 10.1098/rspb.2013.1613 . ПМЦ 3735267 . ПМИД 23902914 .
- ^ Jump up to: а б Топпер, ТП; Сковстед, CB; Пил, Дж. С.; Харпер, DAT (2013). «Линька у лобоподийного ониходиктиона нижнего кембрия Гренландии» . Летайя . 46 (4): 490–495. дои : 10.1111/лет.12026 .
- ^ Jump up to: а б с д Дзик, Ежи; Крумбигель, Гюнтер (1989). «Самый старый онихофоран Xenusion: связь, соединяющая типы?». Летайя . 22 (2): 169–181. дои : 10.1111/j.1502-3931.1989.tb01679.x . ISSN 1502-3931 .
- ^ Jump up to: а б с Рамшельд, Л.; Сянгуан, Хоу (1991). «Новые сходства загадочных многоклеточных животных с раннекембрийскими животными и онихофорами». Природа . 351 (6323): 225–228. Бибкод : 1991Natur.351..225R . дои : 10.1038/351225a0 . ISSN 1476-4687 . S2CID 4309565 .
- ^ Jump up to: а б с д ХОУ, СЯНГУАН; БЕРГСТРЁМ, ЯН (1 мая 1995 г.). "Кембрийские лобоподы - предки современных онихофоров?" . Зоологический журнал Линнеевского общества . 114 (1): 3–19. дои : 10.1111/j.1096-3642.1995.tb00110.x . ISSN 0024-4082 .
- ^ Jump up to: а б с д и ж г час Бадд, Грэм Э. (1998). «Морфология и филогенетическое значение Kerygmachela kierkegaardi Budd (формация Буэн, нижний кембрий, Северная Гренландия)». Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 89 (4): 249–290. дои : 10.1017/S0263593300002418 . ISSN 1473-7116 . S2CID 85645934 .
- ^ Jump up to: а б с д и ж Бадд, Грэм (август 1993 г.). «Кембрийский жаберный лобопод из Гренландии». Природа . 364 (6439): 709–711. Бибкод : 1993Natur.364..709B . дои : 10.1038/364709a0 . ISSN 1476-4687 . S2CID 4341971 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в Смит, Мартин Р.; Ортега-Эрнандес, Хавьер (2014). «Когти галлюцигении, похожие на онихофоры, и случай Tactopoda» (PDF) . Природа . 514 (7522): 363–366. Бибкод : 2014Natur.514..363S . дои : 10.1038/nature13576 . ПМИД 25132546 . S2CID 205239797 . Архивировано (PDF) из оригинала 10 октября 2022 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р Смит, Мартин Р.; Карон, Жан-Бернар (июль 2015 г.). «Голова галлюцигении и глоточный каркас ранних экдизозойных» . Природа . 523 (7558): 75–78. Бибкод : 2015Natur.523...75S . дои : 10.1038/nature14573 . ISSN 0028-0836 . ПМИД 26106857 . S2CID 205244325 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Ортега-Эрнандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (01 мая 2017 г.). «Происхождение и эволюция головы панартропод - палеобиологическая перспектива и перспектива развития» . Строение и развитие членистоногих . Эволюция сегментации. 46 (3): 354–379. дои : 10.1016/j.asd.2016.10.011 . ISSN 1467-8039 . ПМИД 27989966 .
- ^ Jump up to: а б с д и ж г час я дж к л м Сиветер, Дерек Дж.; Бриггс, Дерек Э.Г.; Сиветер, Дэвид Дж.; Саттон, Марк Д.; Легг, Дэвид (2018). «Трехмерно сохранившийся лобопод из Херефордширского (силурийского) Лагерштетте, Великобритания» . Королевское общество открытой науки . 5 (8): 172101. дои : 10.1098/rsos.172101 . ПМК 6124121 . ПМИД 30224988 .
- ^ Смит, Фрэнк В.; Гольдштейн, Боб (01 мая 2017 г.). «Сегментация тихоходок и разнообразие сегментных моделей панартропод». Строение и развитие членистоногих . Эволюция сегментации. 46 (3): 328–340. дои : 10.1016/j.asd.2016.10.005 . ISSN 1467-8039 . ПМИД 27725256 .
- ^ Jump up to: а б с Форти, Ричард А.; Томас, Ричард Х. (31 декабря 1997 г.). Отношения членистоногих . Springer Science & Business Media. ISBN 978-0-412-75420-3 .
- ^ Jump up to: а б с д и ж г час Дзик, Ежи (1 июля 2011 г.). «Переход от ксенузиан к аномалокаридидам внутри лобоподий» (PDF) . Боллеттино делла Сосьета Палеонтологического Итальянского общества . 50 : 65–74. Архивировано (PDF) из оригинала 10 октября 2022 года.
- ^ Пентастомида - Sociedad Entomológica Aragonesa
- ^ Трактат по зоологии - анатомия, таксономия, биология. Ракообразные, Том 5
- ^ Райли, Дж.; Баная, А.А.; Джеймс, Дж.Л. (1 августа 1978 г.). «Филогенетические отношения пентастомид: аргументы в пользу их включения в состав ракообразных» . Международный журнал паразитологии . 8 (4): 245–254. дои : 10.1016/0020-7519(78)90087-5 . ISSN 0020-7519 .
- ^ Лавров Денис В.; Браун, Уэсли М.; Бур, Джеффри Л. (7 марта 2004 г.). «Филогенетическое положение взаимоотношений пентастомид и (пан) ракообразных» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 271 (1538): 537–544. дои : 10.1098/rspb.2003.2631 . ПМК 1691615 . ПМИД 15129965 .
- ^ Режье, Джером К.; Шульц, Джеффри В.; Цвик, Андреас; Хасси, апрель; Болл, Бернард; Ветцер, Регина; Мартин, Джоэл В.; Каннингем, Клиффорд В. (2010). «Отношения членистоногих, выявленные с помощью филогеномного анализа последовательностей, кодирующих ядерные белки» . Природа . 463 (7284): 1079–1083. Бибкод : 2010Natur.463.1079R . дои : 10.1038/nature08742 . ISSN 1476-4687 . ПМИД 20147900 . S2CID 4427443 .
- ^ Лосано-Фернандес, Хесус; Джакомелли, Маттиа; Флеминг, Джеймс Ф; Чен, Альберт; Винтер, Якоб; Томсен, Филип Фрэнсис; Гленнер, Хенрик; Палеро, Ферран; Легг, Дэвид А; Илифф, Томас М; Пизани, Давиде (01 августа 2019 г.). «Эволюция панкрустообразных, освещенная богатыми таксонами наборами данных геномного масштаба с расширенной выборкой реминожек» . Геномная биология и эволюция . 11 (8): 2055–2070. дои : 10.1093/gbe/evz097 . ISSN 1759-6653 . ПМК 6684935 . ПМИД 31270537 .
- ^ Рабет, Николя (2021), «Ракообразные», Справочник по морским модельным организмам в экспериментальной биологии , CRC Press, стр. 271–287, doi : 10.1201/9781003217503-15 , ISBN 978-1-003-21750-3 , S2CID 239958264
- ^ Гулд, С.Дж. (1989). Прекрасная жизнь: сланцы Бёрджесс и природа истории . WW Нортон и компания. Бибкод : 1989wlbs.book.....G . [ нужна страница ]
- ^ Jump up to: а б с д и ж г час я Ма, Сяоя; Хоу, Сянгуан; Бергстрём, январь (2009). «Морфология Luolishania longicruris (нижний кембрий, Чэнцзян Лагерштетте, юго-запад Китая) и филогенетические отношения внутри лобоподий» . Строение и развитие членистоногих . 38 (4): 271–291. дои : 10.1016/j.asd.2009.03.001 . ISSN 1467-8039 . ПМИД 19293001 .
- ^ Jump up to: а б с д и ж г час я Хоу, Сянь-Гуан; Ма, Сяо-Я; Чжао, Цзе; Бергстрём, январь (2004). «Лобоподия Paucipodia inermis из фауны Чэнцзян нижнего кембрия, Юньнань, Китай» . Летайя . 37 (3): 235–244. дои : 10.1080/00241160410006555 .
- ^ Jump up to: а б с д и ж г Оу, Цян; Шу, Деган; Майер, Георг (11 декабря 2012 г.). «Кембрийские лобоподии и современные онихофоры дают новое представление о ранней цефализации панартропод» . Природные коммуникации . 3 (1): 1261. Бибкод : 2012NatCo...3.1261O . дои : 10.1038/ncomms2272 . ISSN 2041-1723 . ПМЦ 3535342 . ПМИД 23232391 .
- ^ Jump up to: а б с д и ж Оу, Цян; Лю, Цзянни; ШУ, ДЕГАН; Хан, Цзянь; Чжан, Чжифэй; Ван, Сяоцяо; Лэй, Цяньпин (01 мая 2011 г.). «Редкий онихофороподобный лобопод из нижнего кембрия Чэнцзян Лагерштетте, Юго-Западный Китай, и его филогенетическое значение» . Журнал палеонтологии . 85 (3): 587–594. Бибкод : 2011JPal...85..587O . дои : 10.1666/09-147R2.1 . JSTOR 23020193 . S2CID 53056128 .
- ^ Jump up to: а б с д и ж г час я Карон, Жан-Бернар; Ария, Седрик (2020). «Монстр Коллинзов, шипистый лобопод, питающийся взвесью, из кембрийских сланцев Бёрджесс в Британской Колумбии» . Палеонтология . 63 (6): 979–994. дои : 10.1111/пала.12499 . ISSN 1475-4983 . S2CID 225593728 .
- ^ Jump up to: а б с Уиттингтон, Гарри Блэкмор (16 ноября 1978 г.). «Животное-лобопод Aysheaia Pedunculata Walcott, средний кембрий, сланцы Бёрджесс, Британская Колумбия» . Философские труды Лондонского королевского общества. Б. Биологические науки . 284 (1000): 165–197. Бибкод : 1978RSPTB.284..165W . дои : 10.1098/rstb.1978.0061 .
- ^ Jump up to: а б с д и ж г час Лю, Дж.; Шу, Д.; Хан, Дж.; Чжан, З.; Чжан, X. (2007). «Морфоанатомия лобопода Magadictyon cf. Haikouensis из раннего кембрия Чэнцзян Лагерштетте, Южный Китай». Акта Зоология . 88 (4): 279–288. дои : 10.1111/j.1463-6395.2007.00281.x .
- ^ Jump up to: а б с Йокуш, Элизабет Л. (1 сентября 2017 г.). «Развитие и эволюционные перспективы происхождения и разнообразия придатков членистоногих» . Интегративная и сравнительная биология . 57 (3): 533–545. дои : 10.1093/icb/icx063 . ISSN 1540-7063 . ПМИД 28957524 .
- ^ Jump up to: а б с д и Винтер, Якоб; Поррас, Луис; Янг, Флетчер; Бадд, Грэм; Эджкомб, Грегори (01 сентября 2016 г.). «Ротовой аппарат кембрийского жаберного лобопода Pambdelurion Whittingtoni » . Палеонтология . 59 (6): 841–849. Бибкод : 2016Palgy..59..841V . дои : 10.1111/пала.12256 . hdl : 1983/16da11f1-5231-4d6c-9968-69ddc5633a8a . S2CID 88758267 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п Карон, Жан-Бернар; Ария, Седрик (31 января 2017 г.). «Кембрийские лобоподии, питающиеся взвесью, и ранняя радиация панартропод» . Эволюционная биология BMC . 17 (1): 29. Бибкод : 2017BMCEE..17...29C . дои : 10.1186/s12862-016-0858-y . ISSN 1471-2148 . ПМЦ 5282736 . ПМИД 28137244 .
- ^ Jump up to: а б Шенеманн, Бриджит; Лю, Цзянь-Ни; Шу, Де-Ган; Хан, Цзянь; Чжан, Чжи-Фей (2009). «Миниатюрная оптимизированная зрительная система в нижнем кембрии». Летайя . 42 (3): 265–273. дои : 10.1111/j.1502-3931.2008.00138.x . ISSN 1502-3931 .
- ^ Jump up to: а б с д и ж г час я дж Ховард, Ричард Дж.; Хоу, Сянгуан; Эджкомб, Грегори Д.; Салге, Тобиас; Ши, Сяомэй; Ма, Сяоя (27 февраля 2020 г.). «Живущий в трубке лобопод раннего кембрия» . Современная биология . 30 (8): 1529–1536.e2. дои : 10.1016/j.cub.2020.01.075 . ISSN 0960-9822 . ПМИД 32109391 .
- ^ Флеминг, Джеймс Ф.; Кристенсен, Райнхардт Мёбьерг; Соренсен, Мартин Винтер; Пак Тэ-Юн С.; Аракава, Казухару; Блакстер, Марк; Ребекки, Лорена; Гуидетти, Роберто; Уильямс, Том А.; Робертс, Николас В.; Винтер, Якоб (05 декабря 2018 г.). «Молекулярная палеонтология освещает эволюцию зрения экдизозоев» . Труды Королевского общества B: Биологические науки . 285 (1892): 20182180. doi : 10.1098/rspb.2018.2180 . ISSN 0962-8452 . ПМК 6283943 . ПМИД 30518575 .
- ^ Jump up to: а б с д Пак Тэ-Юн С.; Ким, Джи-Хун; Ууу, Джусун; Парк, Чанкунь; Ли, Вон Янг; Смит, М. Пол; Харпер, Дэвид А.Т.; Янг, Флетчер; Нильсен, Арне Т. (9 марта 2018 г.). «Мозг и глаза Керигмачелы свидетельствуют о протоцеребральном происхождении головы панартропода» . Природные коммуникации . 9 (1): 1019. Бибкод : 2018NatCo...9.1019P . дои : 10.1038/s41467-018-03464-w . ISSN 2041-1723 . ПМЦ 5844904 . ПМИД 29523785 .
- ^ Jump up to: а б с д и ж Чен, Дж. Я., Чжоу, GQ, Рамшельд, Л. (1995a). Кембрийский лобопод Microdictyon sinicum . Бюллетень Национального музея естествознания 5, 1–93 (Тайчжун, Тайвань).
- ^ Jump up to: а б с д и ж Ма, Сяоя; Эджкомб, Грегори; Легг, Дэвид; Хоу, Сянгуан (8 мая 2013 г.). «Морфология и филогенетическое положение кембрийского лобопода Diania cactiformis » . Журнал систематической палеонтологии . 12 (4): 445–457. дои : 10.1080/14772019.2013.770418 . S2CID 220463025 .
- ^ Jump up to: а б Штайнер, М.; Ху, SX; Лю, Дж.; Кеупп, Х. (2 февраля 2012 г.). «Новый вид галлюцигений из кембрийской формации Улунцин 4-го этапа Юньнани (Южный Китай) и строение склеритов лобоподий» . Бюллетень геонаук : 107–124. дои : 10.3140/bull.geosci.1280 . ISSN 1802-8225 .
- ^ Jump up to: а б с д и ж Чжан, Си-Гуан; Смит, Мартин Р.; Ян, Цзе; Хоу, Джин-Бо (2016). «Онихофораноподобная мускулатура у фосфатизированного кембрийского лобопода» . Письма по биологии . 12 (9): 20160492. doi : 10.1098/rsbl.2016.0492 . ISSN 1744-9561 . ПМК 5046927 . ПМИД 27677816 .
- ^ Jump up to: а б с Лю, Цзянни; Штайнер, Майкл; Данлоп, Джейсон А.; Кеупп, Хельмут; Шу, Деган; Оу, Цян; Хан, Цзянь; Чжан, Чжифэй; Чжан, Синлян (февраль 2011 г.). «Бронированный кембрийский лобопод из Китая с членистоногими придатками». Природа . 470 (7335): 526–530. Бибкод : 2011Natur.470..526L . дои : 10.1038/nature09704 . ISSN 1476-4687 . ПМИД 21350485 . S2CID 4324509 .
- ^ Jump up to: а б с д и Ваннье, Жан; Лю, Цзянни; Лерози-Обрил, Руди; Винтер, Якоб; Дэйли, Эллисон К. (2 мая 2014 г.). «Сложная пищеварительная система у ранних членистоногих» . Природные коммуникации . 5 (1): 3641. Бибкод : 2014NatCo...5.3641V . дои : 10.1038/ncomms4641 . ISSN 2041-1723 . ПМИД 24785191 .
- ^ Баттерфилд, Нью-Джерси (2002). «Кишки Leanchoilia и интерпретация трехмерных структур в окаменелостях типа сланцев Берджесс». Палеобиология . 28 : 155–171. doi : 10.1666/0094-8373(2002)028<0155:LGATIO>2.0.CO;2 . ISSN 0094-8373 . S2CID 85606166 .
- ^ Jump up to: а б с д Янг, Флетчер Дж.; Винтер, Якоб (2017). «Онихофороподобная миоанатомия кембрийского жаберного лобопода Pambdelurion Whittingtoni » (PDF) . Палеонтология . 60 (1): 27–54. Бибкод : 2017Palgy..60...27Y . дои : 10.1111/пала.12269 . hdl : 1983/92180ef0-2205-4c65-9a70-90d59cfea2f4 . ISSN 1475-4983 . S2CID 55477207 . Архивировано (PDF) из оригинала 10 октября 2022 г.
- ^ Jump up to: а б с д и Бадд, GE (2001). «Почему членистоногие сегментированы?». Эволюция и развитие . 3 (5): 332–42. дои : 10.1046/j.1525-142X.2001.01041.x . ПМИД 11710765 . S2CID 37935884 .
- ^ МакКолл, CRA (2023). «Большой пелагический лобопод из кембрийских сланцев Пиоче в Неваде». Журнал палеонтологии . 97 (5): 1009–1024. дои : 10.1017/jpa.2023.63 .
- ^ Jump up to: а б с д Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2015). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками». Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ISSN 1476-4687 . ПМИД 25762145 . S2CID 205242881 .
- ^ Бадд, Грэм Э.; Дейли, Эллисон К. (январь 2012 г.). «Доли и лобоподы Opabinia regalis из среднекембрийских сланцев Бёрджесс: доли Opabinia» . Летайя . 45 (1): 83–95. дои : 10.1111/j.1502-3931.2011.00264.x .
- ^ Чен, Ж.-й; Рамскольд, Л.; Чжоу, Г.-к (1994). «Доказательства монофилии и близости кембрийских гигантских хищников к членистоногим» . Наука . 264 (5163): 1304–1308. Бибкод : 1994Sci...264.1304C . дои : 10.1126/science.264.5163.1304 . ISSN 0036-8075 . ПМИД 17780848 . S2CID 1913482 .
- ^ Эджкомб, Грегори Д. (2 ноября 2020 г.). «Происхождение членистоногих: объединение палеонтологических и молекулярных данных» . Ежегодный обзор экологии, эволюции и систематики . 51 (1): 1–25. doi : 10.1146/annurev-ecolsys-011720-124437 . ISSN 1543-592X . S2CID 225478171 .
- ^ Уиттл, Р.Дж.; Габботт, SE; Олдридж, Р.Дж.; Терон, Дж. (2009). «Ордовикский лобопод из сланца Сум-Шале-Лагерштетте, Южная Африка» . Палеонтология . 52 (3): 561–567. Бибкод : 2009Palgy..52..561W . дои : 10.1111/j.1475-4983.2009.00860.x . S2CID 129609503 .
- ^ Эджкомб, Грегори Д. (1 июня 2009 г.). «Палеонтологические и молекулярные данные, связывающие членистоногих, онихофоров и других экдизозоа» . Эволюция: образование и информационно-пропагандистская деятельность . 2 (2): 178–190. дои : 10.1007/s12052-009-0118-3 . ISSN 1936-6434 .
- ^ Робисон, РА (1985). «Сродство Aysheaia (Onychophora) с описанием нового кембрийского вида». Журнал палеонтологии . 59 (1): 226–235. ISSN 0022-3360 . JSTOR 1304837 .
- ^ Бергстрем, Дж.; Хоу, Сянь Гуан (1 декабря 2001 г.). «Кембрийские онихофоры или ксенусианцы» . Зоологический вестник . 240 (3–4): 237–245. дои : 10.1078/0044-5231-00031 .
- ^ Jump up to: а б Бадд, Грэм Э. (1 января 2001 г.). «Тихоходки как« членистоногие стволовой группы »: данные кембрийской фауны». Zoologischer Anzeiger - Журнал сравнительной зоологии . 240 (3): 265–279. дои : 10.1078/0044-5231-00034 . ISSN 0044-5231 .
- ^ Маунс, Росс CP; Уиллс, Мэтью А. (август 2011 г.). «Филогенетическое положение Диании под вопросом» . Природа . 476 (7359): E1, обсуждение E3–4. Бибкод : 2011Natur.476E...1M . дои : 10.1038/nature10266 . ISSN 1476-4687 . ПМИД 21833044 . S2CID 4417903 .
- ^ Легг, Дэвид; Ма, Сяоя; Вулф, Джоанна; Ортега-Эрнандес, Хавьер; Эджкомб, Грегори; Саттон, Марк (11 августа 2011 г.). «Повторный анализ филогении лобоподий» . Природа . 476 (7359): E2–3, обсуждение E3. Бибкод : 2011Natur.476Q...1L . дои : 10.1038/nature10267 . ПМИД 21833046 . S2CID 4310063 .
- ^ Маас, Андреас; Валошек, Дитер (1 января 2001 г.). «Кембрийские производные стволовой линии ранних членистоногих, пентастомид, тихоходок и лобоподов: перспектива Орстена» . Zoologischer Anzeiger - Журнал сравнительной зоологии . 240 (3): 451–459. дои : 10.1078/0044-5231-00053 . ISSN 0044-5231 .
- ^ Ким, Джи-Хун; Смит, Фрэнк В.; Ким, Сангхи; Ро, Хён Су; Чжан, Синлян; Лю, Цзянни; Пак, Тэ Юн С. (2023). «Кембрийские лобоподии проливают свет на происхождение строения тела тихоходок» . Труды Национальной академии наук . 120 (28): e2211251120. дои : 10.1073/pnas.2211251120 . ПМЦ 10334802 . ПМИД 37399417 .
- ^ Пейтс, Стивен; Дейли, Эллисон К. (2017). «Кариосинтрипс: радиодонтан из кембрия Испании, США и Канады» . Статьи по палеонтологии . 3 (3): 461–470. дои : 10.1002/spp2.1084 . ISSN 2056-2802 . S2CID 135026011 .
- ^ «Aysheaia prolata из формации Юта Уиллер (Друмский период, кембрий) представляет собой лобный придаток радиодонтана Stanleycaris — Acta Palaeontologica Polonica» . www.app.pan.pl. Проверено 8 января 2020 г.
- ^ «Ответ на комментарий на тему «Aysheaia prolata из формации Юта Уиллер (Друмский период, кембрий) представляет собой лобный придаток радиодонтана Stanleycaris» с формальным описанием Stanleycaris — Acta Palaeontologica Polonica» . www.app.pan.pl. Проверено 8 января 2020 г.
- ^ Лю, Цзянни; Шу, ДеГан; Хан, Цзянь; Чжан, Чжифей (1 мая 2008 г.). «Сравнительное изучение кембрийских лобопод Miraluolishania и Luolishania» . Китайский научный бюллетень . 53 (1): 87–93. Бибкод : 2008ЧСБу..53...87Л . дои : 10.1007/s11434-007-0428-1 . S2CID 128619311 .
- ^ Jump up to: а б Сянгуан, Хоу; Рамшельд, Ларс; Бергстрем, Ян (1991). «Состав и сохранение фауны Чэнцзян – мягкотелой биоты нижнего кембрия» . Зоологика Скрипта . 20 (4): 395–411. дои : 10.1111/j.1463-6409.1991.tb00303.x . ISSN 1463-6409 . S2CID 85077111 .
- ^ Ян, Цзе; Ортега-Эрнандес, Хавьер; Гербер, Сильвен; Баттерфилд, Николас Дж.; Хоу, Джин-бо; Лан, Тиан; Чжан, Си-гуан (14 июля 2015 г.). «Сверхбронированный лобопод из кембрия Китая и раннее неравенство в эволюции Onychophora» . Труды Национальной академии наук . 112 (28): 8678–8683. Бибкод : 2015PNAS..112.8678Y . дои : 10.1073/pnas.1505596112 . ISSN 0027-8424 . ПМК 4507230 . ПМИД 26124122 .
- ^ Ария, К.; Карон, Ж.-Б. (2024). «Глубокое происхождение стратегий артикуляции у панартропод: данные от нового лобопода луолишаниид (Panarthropoda) из пластов тюльпанов, сланцы Берджесс». Журнал систематической палеонтологии . 22 (1). 2356090. Бибкод : 2024JSPal..2256090A . дои : 10.1080/14772019.2024.2356090 .
- ^ Jump up to: а б Чжан, Си-Гуан; Олдридж, Ричард Дж. (2007). «Развитие и диверсификация туловищных пластинок лобоподий нижнего кембрия» . Палеонтология . 50 (2): 401–415. Бибкод : 2007Palgy..50..401Z . дои : 10.1111/j.1475-4983.2006.00634.x . ISSN 1475-4983 . S2CID 85293118 .
- ^ БАДД, GE, ПИЛ, Дж. С. (1998). Новый лобопод ксенузиид из раннекембрийской фауны Sirius Passet Северной Гренландии . Палеонтология , 41, 6, 1201–1213.
- ^ Луо, HL, Ху, SX и Чен, LZ Раннекембрийская фауна Чэнцзян из региона Куньмин, Китай . 129 (Юньнаньская научно-техническая пресса, 1999).
- ^ Jump up to: а б МакКолл, Кристиан (13 декабря 2023 г.). «Большой пелагический лобопод из кембрийских сланцев Пиоче в Неваде» . Журнал палеонтологии . 97 (5): 1009–1024. дои : 10.1017/jpa.2023.63 . Проверено 13 декабря 2023 г.
- ^ Хоу, Сянгуан; Бергстрем, Ян; Цзе, Ян (2006). «Отличие аномалокаридид от членистоногих и приапулид». Геологический журнал . 41 (3–4): 259–269. Бибкод : 2006GeolJ..41..259X . дои : 10.1002/gj.1050 . S2CID 83582128 .
- ^ Маас, Андреас; Майер, Георг; Кристенсен, Рейнхардт М.; Валошек, Дитер (1 декабря 2007 г.). «Кембрийский микролобопод и эволюция передвижения и размножения членистоногих» . Китайский научный бюллетень . 52 (24): 3385–3392. Бибкод : 2007ЧСБу..52.3385М . дои : 10.1007/s11434-007-0515-3 . ISSN 1861-9541 . S2CID 83993887 .
- ^ Цзюнь-юань, Чэнь; Гуй-цин, Чжоу; Рамшельд, Ларс (1994). «Новое раннекембрийское животное, похожее на онихофора, Paucipodia gen. nov., из фауны Чэнцзян, Китай» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 85 (4): 275–282. дои : 10.1017/S0263593300002042 . ISSN 1473-7116 . S2CID 131555757 .
- ^ Лерози-Обрил, Р.; Ортега-Эрнандес, Х. (2022). «Новый лобопод из среднего кембрия штата Юта: эволюционировали ли плавательные закрылки тела конвергентно у членистоногих стеблевой группы?». Статьи по палеонтологии . 8 (3): е1450. дои : 10.1002/spp2.1450 . S2CID 250076505 .
- ^ Смит, Мартин Р.; Лонг, Эмма Дж.; Дхунгана, Алавья; Добсон, Кэтрин Дж.; Ян, Цзе; Чжан, Сигуан (2024). «Системы органов личинки кембрийского членистоногого». Природа . 624 : 123–128. дои : 10.1038/s41586-024-07756-8 .