Радиодонта
| Радиодонта Временной диапазон: | |
|---|---|
 | |
| Слева направо, сверху вниз: Amplectobelua symbrachiata , Anomalocaris canadensis , Aegirocassis benmoulai , Peytoia nathorsti , Lyrarapax unguispinus , Cambroraster falcatus и Hurdia victoria. | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | †Radiodonta Collins, 1996 |
| Families | |
| |
Radiodonta — вымерший отряд членистоногих стволовой группы , который пользовался успехом во всем мире в кембрийский период. Их можно назвать радиодонтами . [1] [2] [3] радиодонтаны , [4] [5] радиодонтиды , [6] аномалокариды , [7] или аномалокаридиды , [8] [9] [10] хотя два последних первоначально относятся к семейству Anomalocarididae , которое ранее включало все виды этого отряда, но теперь ограничивается лишь несколькими видами. [7] Радиодонты отличаются характерными лобными придатками, морфологически разнообразными и выполняющими самые разные функции. Радиодонты включали в себя самых ранних известных крупных хищников, но среди них также были просеиватели отложений и фильтраторы. [11] Некоторые из наиболее известных видов радиодонтов — кембрийские таксоны Anomalocaris canadensis , Hurdia victoria , Peytoia nathorsti , Titanokorys Ganessii , Cambroraster falcatus и Amplectobelua symbrachiata . выжившим представителям относятся подсемейство Aegirocassisinae из раннего ордовика Марокко К более поздним и раннего девона представитель Schinderhannes bartelsi из Германии .
Этимология
[ редактировать ]The name Radiodonta (Latin for radius "spoke of a wheel" and Greek for odoús "tooth") refers to the radial arrangement of tooth plates (oral cone) surrounding the mouth,[6] although these features are suggested to be absent in some radiodont species.[4][1]
Definition
[edit]The original diagnosis of order Radiodonta in 1996 is as follows:[6]
Radiodontids are bilaterally symmetrical, elongate arthropods with a nonmineralized cuticle typically most robust in the jaws and claws. The body is subdivided into two tagmata, much like the prosoma and opisthosoma of chelicerate arthropods. Typically, the front part shows no external segmentation, bears one pair of preoral claws, a pair of prominent eyes, and ventral jaws with radiating teeth. Some forms have additional rows of teeth and three or four postoral gnathobasic limb pairs. The trunk is metameric, typically with about 13 segments laterally developing imbricating lobes for swimming and gills for respiration, and may end in a prominent three-part tail. Some forms have gnathobasic trunk limbs.
In 2014, the clade Radiodonta was defined phylogenetically as a clade including any taxa closer to Anomalocaris canadensis than Paralithodes camtschaticus.[7] In 2019, it was redefined morphologically as animal bearing head carapace complex with central (H-) and lateral (P-) elements; outgrowths (endites) from frontal appendages bearing auxiliary spines; and reduced anterior flaps or bands of lamellae (setal blades) and strong tapering of body from anterior to posterior.[3]
Description
[edit]
Most radiodonts were significantly larger than the other Cambrian fauna, with typical body lengths of large taxa varying from 30 to 50 cm (12 to 20 in).[12] The largest described radiodont is the Early Ordovician species Aegirocassis benmoulai, which may have grown up to 2 m (6.6 ft) long.[10][2] A nearly complete specimen of a juvenile Lyrarapax unguispinus measured only 18 mm (0.71 in), making it among the smallest radiodont specimens known, though adults reached a length of 8.3 cm (3.3 in)[2][13] An isolated frontal appendage of a hurdiid with a length less than half that of the juvenile Lyrarapax is known, but it is not known whether this specimen pertains to an adult.[14] The largest known Cambrian radiodont was Amplectobelua, reaching lengths of up to 90 cm (35 in) based on an incomplete specimen.[15] Anomalocaris canadensis was also relatively large, estimated up to 34.2–37.8 cm (13.5–14.9 in) long,[2] and the Cambrian hurdiid Titanokorys approached around 50 cm (20 in) long.[16]
The body of a radiodont could be divided into two regions: head and trunk. The head is composed of only one body segment[17] known as the ocular somite, covered by sclerites (head carapace complex), bore arthropodized frontal appendages, ventral mouthparts (oral cone), and stalked compound eyes. The tapering trunk is composed of multiple body segments, each associated with pairs of flaps and gill-like structures (setal blades).[3]
Frontal appendage
[edit]

The anterior structures on the head are a pair of frontal appendages which have been referred to as 'claws', 'grasping appendages', 'feeding appendages', or 'great appendages' in previous studies (the last term is discouraged since the homology between frontal appendages and the original, morphologically distinct megacheiran great appendages is questionable.[17][18]). They are sclerotized (hardened) and arthropodized (segmented), bearing ventral endites (spines) on most of their podomeres (segmental units), and the endites may bear additional rows of auxiliary spines on their anterior and posterior margins.[19][3] The frontal appendage consists of two regions: the shaft ('peduncle',[2] 'base'[20] or 'promixal region'[2] in some studies) and the distal articulated region[19] (also referred to as 'claw'[20]). A triangular region covered by soft cuticle (arthrodial membrane) may occur on the ventral side between podomeres and provide flexibility.[21][11] Their purported pre-ocular and protocerebral origin suggest they are homologous to the primary antennae of Onychophora and the labrum of Euarthropoda (all arose from ocular somite),[17][9] while subsequent studies also suggest a deutocerebral origin and homologous with the chelicerae of Chelicerata and the antennae or 'great appendages' of other arthropods (all arose from post-ocular somite 1).[22] Since the morphology of the frontal appendages, especially those of the spines, always differs between species, it is one of the most important means of species identification.[19] In fact, many radiodonts are only known from a handful of fossilized frontal appendages.[21][19]
 Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species
Frontal appendages of Anomalocarididae, Amplectobeluidae, and possibly related species Frontal appendages of Tamisiocarididae
Frontal appendages of Tamisiocarididae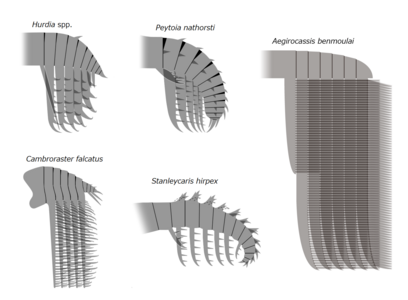 Frontal appendages of Hurdiidae
Frontal appendages of Hurdiidae



Oral cone
[edit]
The mouth is on the ventral side of the head, behind the attachment point of frontal appendages and is surrounded by a ring of tooth plates, forming the mouthpart known as oral cone ('jaws' in previous studies[6]). 3 or 4 tooth plates might be enlarged, giving the oral cone a triradial (e.g. Anomalocaris, Echidnacaris) or tetraradial (e.g. Hurdiidae, Lyrarapax) appearance.[23][13][24] The inner margin of tooth plates have spikes facing towards the mouth opening. Additional rows of internal tooth plates may occur in some hurdiid genera.[8][3] Detail reconstruction of some amplectobeluid oral cones are speculative, but they possibly did not present a typical radial arrangement.[4][1]
Head sclerites, eyes and trunk
[edit]
Three head sclerite (carapace) complex formed by a central H-element (anterior sclerite or head shield) and a pair of P-elements (lateral sclerites) cover the dorsal and laterovental surface of the animal's head.[3] The P-elements may connect to each other as well as the H-element by a narrow anterior extension (P-element neck or 'beak').[8][3] The head sclerites are small and ovoid in Anomalocarididae and Amplectobeluidae,[4][3] but often enlarged in Hurdiidae, corresponded to their distinct body shapes (streamlined in Anomalocarididae/Amplectobeluidae but often compact in Hurdiidae).[3] The head bore two stalked compound eyes, which may have had mobility,[25] and are located between the gaps formed by the posterior regions of the H-element and P-elements.[8][3] The compound eyes of Echidnacaris are exceptionally unstalked.[14] Some species possess an additional median eye behind the H-element.[22]


Contrary to the original diagnosis, the division of body segments (segmental boundaries) can be visible externally[10][5][3] and no known member of Radiodonta (except the putative radiodont Cucumericrus[10][26]) is known to have pediform trunk appendages (legs).[27] The trunk has numerous body segments (somites), tapering from anterior to posterior, with the anterior three or four segments significantly constricted into a neck region.[3]
 Variations of radiodont body flaps
Variations of radiodont body flaps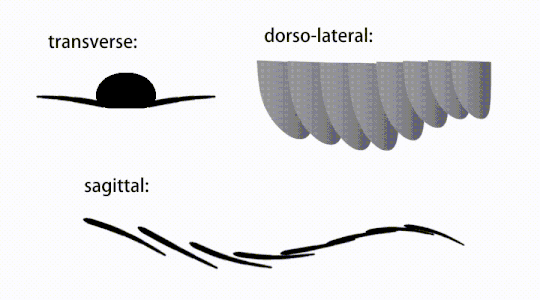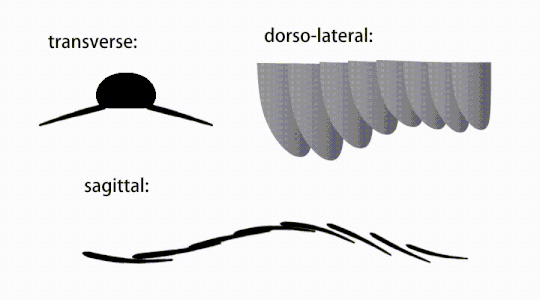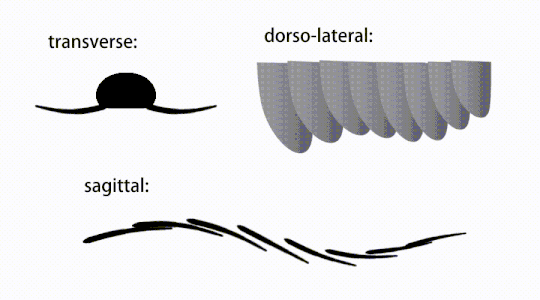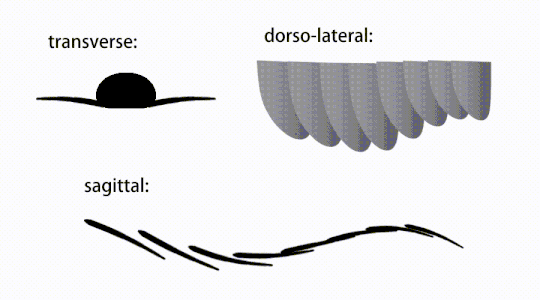 The movement of radiodont flapping appendages
The movement of radiodont flapping appendages Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps
Ventral view of a generalized GLS-bearing radiodont, showing gnathobase‐like structures (GLSs) associated with reduced anterior flaps



The trunk appendages were fin-like body flaps ('lateral flaps' or 'lobes' in some studies), usually one pair of ventral flaps per body segment, each slightly overlapping the one more anterior to it, but additional, non-overlapping sets of small dorsal flaps may occur in some Hurdiid species.[10] The flaps may have numerous vein-like structures (referred to as 'strengthening rays',[5] 'flap rays',[3] 'tranverse rods',[10] 'transverse lines'[28] or 'veins'[29]). The flaps on the neck region (referred to as 'reduced flaps',[4] 'neck flaps',[5] 'head flaps',[27] 'anterior flaps'[30] or 'differentiated flaps'[18]) are significantly reduced. In some species, jaw-like feeding appendages called gnathobase-like structures (GLSs) arose from each of the bases of their reduced neck flaps.[4][1] Numerous elongated blade-like extensions (referred to as lanceolate blades or lamellae[3]) arranged in a row, forming bands of gill-like structures known as setal blades, covered the dorsal surface of each body segment.[10] At least in Aegirocassis, each of the lanceolate blades are covered in wrinkles.[10] The ventral flaps may be homologous to the endopod of the biramous limbs of euarthropods and lobopodous limbs (lobopods) of gilled lobopodians, and the dorsal flaps and setal blades may be homologous to the exite and gill-bearing dorsal flaps of the former taxa.[31][10] The trunk may end either with a tail fan compose of 1 to 3 pairs of blades,[29][27][3] a pair of long furcae,[29][13][3] an elongated terminal structure,[27] or a featureless blunt tip.[10]
 Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian.
Stanleycaris, Hurdia, Aegirocassis, Peytoia and Cambroraster are all examples of hurdiid radiodonts. These were the most diverse and long lasting of the radiodont families, surviving from the Cambrian up until the Devonian. Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris.
Anomalocaris is a member of the anomalocarididae family, which at one point included all radiodonts, but now only includes a few genera such as Lenisicaris. Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts.
Amplectobelua and Lyrarapax are representatives of the amplectobeluidae which is a very inclusive family of mainly Chinese radiodonts. Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.
Echidnacaris and Tamisiocaris are examples of the family tamisiocarididae which were exclusively suspension feeding radiodonts from the Cambrian.




Internal structures
[edit]

Traces of muscles, digestive system and nervous system were described from some radiodont fossils. Pairs of well-developed muscles were connected to the ventral flaps located at the lateral cavities of each body segment.[27][9] Between the lateral muscles is a sophisticated digestive system, formed by a widening of the foregut and hindgut, both connected by a narrow midgut associated with six pairs of gut divercula (digestive glands).[27][5][32]
The brain of radiodonts was simpler than the three-segmented (compose of pro-, deuto- and tritocerebrum) brains of euarthropods, but further interpretations differ between studies. Based on Cong et al. 2014, the brain composed of only one brain segment originating from the ocular somite, the protocerebrum. The nerves of the frontal appendages and compound eyes arose from the anterior and lateral regions of the brain.[9][17] Based on Moysiuk & Caron 2022, the frontal appendage nerves arose from the ventral deutocerebrum, the second brain segment. The previous "frontal appendage nerves" actually represent median eye nerve.[22] In both interpretations, posterior to the brain was a pair of apparently unfused ventral nerve cords which ran through the animal's neck region.[9][22]
Paleoecology
[edit]Physiology
[edit]
Radiodonts were interpreted as nektonic or nektobenthic animals, with their morphology suggesting an active swimming lifestyle. The muscular, overlapping ventral flaps may have propelled the animal through the water, possibly by moving in a wave-like formation resembling modern rays and cuttlefish.[33][34] Pairs of dorsal flaps, which make up a tail fan in some species, may have helped steering and/or stabilizing the animal during locomotion.[10][35] In Anomalocaris, morphology of the tail fan even suggests it could rapidly change its swimming direction efficiently.[36] On the other hand, some hurdiids have features significantly specialized for a nektobenthic lifestyle, such as Cambroraster with its dome-like H-element similar to the carapace of a horseshoe crab.[3] Bands of setal blades with wrinkling lanceolate blades may have increased the surface area, suggesting they were gills, providing the animal's respiratory function.[27][10] Abundance of the remains of scleritzed structures such as disarticulated frontal appendages and head sclerite complexes, suggest that mass moulting events may have occurred among radiodonts,[10][3] a behavior which also has been reported in some other Cambrian arthropods such as trilobites.[37]
Diet
[edit]





Radiodonts had diverse feeding strategies, which could be categorized as raptorial predators, sediment sifters, or suspension, filter feeders.[2][38][11][39][40] For example, raptorial predators like Anomalocaris and Amplectobeluids might have been able to catch agile prey by using their raptorial frontal appendages; the latter even bore a robust endite for holding prey like a pincer.[26][21][4][11] With the smaller head carapace complex and large surface of arthrodial membranes, frontal appendages of these taxa had greater flexibility.[13] Stout frontal appendages of sediment sifters like Hurdia and Peytoia have serrated endites with mesial curvature, which could form a basket-like trap for raking through sediment and passing food items towards the well-developed oral cone.[3][11] Endites of frontal appendages from suspension/filter feeders like Tamisiocaris and Aegirocassis have flexible, densely-packed auxiliary spines, which could filter out organic components such as mesozooplankton and phytoplankton down to 0.5mm.[7][10] Frontal appendages of Caryosyntrips, which are unusual for radiodonts in having the direction of endite-bearing surfaces opposing one another and may have been able to manipulate and crush prey in a scissor-like slicing or grasping motion.[21][41]
Oral cones of radiodonts may have been used for suction and/or biting.[23][38][3] Together with the great variety of frontal appendages in different species of radiodonts, differentiation of oral cones between species suggests preferences of different diets as well.[38][11] For example, the triradial oral cone of Anomalocaris with irregular, tuberculated toothplates and a small opening may have been adapted to small and nektonic prey,[23][11] while the rigid tetraradial oral cones of Peytoia, Titanokorys, Hurdia, and one isolated oral cone attributed to Cambroraster with a larger opening and sometimes additional tooth plates may have been capable to consume larger food items relative to their body size and probably benthic or endobenthic prey.[23][38][3]
Classification
[edit]Taxonomic affinities
[edit]
| |||||||||||||||||||||||||||||||||||||||||||||
| Summarized phylogeny between Radiodonta and other Ecdysozoan taxa[42] |







 The radiodont/opabiniid-euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods
The radiodont/opabiniid-euarthropod intermediate Kylinxia, shares many of the characteristics found in both dinocaridids and euarthropods A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages
A life reconstruction of the basal deuteropod Erratus, which helped reveal the evolution of arthropod trunk appendages Mieridduryn is a dinocaridid panarthropod from the Middle Ordovician that shares traits with both opabiniids and radiodonts
Mieridduryn is a dinocaridid panarthropod from the Middle Ordovician that shares traits with both opabiniids and radiodonts




Most phylogenetic analyses suggest that radiodonts, alongside opabiniids (Opabinia and Utaurora[43]), are stem-group arthropods just basal to deuteropoda,[42] a clade including upper stem (e.g. fuxianhuiids and bivalved arthropods) and crown Euarthropoda (e.g. Artiopoda, Chelicerata and Mandibulata).[8][44][45][46][47][48][7][9][10][2][3][30][18][39][40][49][50][43] This interpretation is supported by numerous arthropod groundplan found on radiodonts and opabiniids, such as stalked compound eyes,[25] digestive glands,[32] trunk appendages forming by dorsal and ventral elements (precursor of arthropod biramous appendages).[10][50] Compared to opabiniids, which possess posterior mouth opening and fused frontalmost appendages (comparable to euarthropod posterior-facing labrum/hypostome complex),[17][43] radiodonts on the other hand featured euarthropod-like dorsal sclerite (H-element) and arthropodization (frontal appendages) on their head regions,[51][17][43] alongside cuticularized gut termini.[27] The fact that both radiodonts and opabiniids lack exoskeleton on their trunk region suggests that the origin of compound eyes and arthropodization (segmented appendages) precede arthrodization (full set of trunk exoskeleton) in the arthropod stem lineage.[42][52][53] The constricted neck region with feeding appendicular structures of some radiodont may also shed light on the origin of the sophisticated arthropod head, which was formed by the fusion of multiple anterior body segments.[4][17] Basal deuteropods that possess a mixture of radiodont/opabiniid characters like Kylinxia and Erratus, may represent intermediate forms between radiodonts, opabiniids and other euarthropods.[18][50]
 The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right)
The siberiid lobopodians Siberion (upper left), Megadictyon (bottom center) and Jianshanopodia (upper right)
 Megacheira or 'great appendage arthropods', a class of possible stem-chelicerate previously thought to be radiodont's close relative
Megacheira or 'great appendage arthropods', a class of possible stem-chelicerate previously thought to be radiodont's close relative



Taxa just basal to the radiodont, opabiniid and euarthropod branch are 'gilled lobopodians' like Pambdelurion and Kerygmachela, which occasionally united under the class Dinocaridida with opabibiids and radiodonts.[54][46] They have body flaps, digestive glands, large (presumely compound) eyes and specialized frontal appendages like the former taxa, but their frontal appendages are not arthropodized nor fused, eyes sessile, gill-like structures less prominent, and certainlly bore lobopod underneath each of their flaps.[55][10][56][43] Taxa even basal to 'gilled lobopodians' are siberiids like Megadictyon and Jianshanopodia,[42] a group of lobopodians that bore robust frontal appendages and digestive glands, but no body flaps. Such intermediate forms between lobopodian and radiodont/euarthropod suggest that the total-group Arthropoda arose from a paraphyletic lobopodian grade, alongside the other two extant panarthropod phyla Tardigrada and Onychophora.[57][42][17][58][52][53]
Previous studies may suggest radiodonts as a group other than stem-arthropods, such as a hitherto unknown phylum;[33] cycloneuralian worms undergone convergent with arthropods (based on the cycloneuralian-like radial mouthparts);[59][54] stem chelicerate euarthropods alongside megacheirans also known as great appendage arthropods (based on the similarity between radiodont frontal appendages, megacheiran great appendages and chelicerae);[60] or Schinderhannes bartelsi, which resolved as a hurdiid radiodont in recent analyses,[42][7][10][2][3][39][40] as a species more closely related to euarthropods than other radiodonts (based on some putative euarthropod-like features found on Schinderhannes).[35] However, neither each of them were supported by later investigations. The radial mouthparts are not cycloneuralian-exclusive and more likely present result of convergent evolution or ecdysozoan plesimorphy, since they also have been found in panarthropods such as tardigrade and some lobopodians;[61] radiodonts lacking definitive euarthropod features such as trunk tergites and multiple head appendages,[42] and the megacheiran great appendages were considered to be deutocerebral,[62][63] which could be non-homologous to the radiodont protocerebral frontal appendages;[9][17] putative euarthropod characters found on the single Schinderhannes fossil is questionable and may present other radiodont-like structures.[42]
Interrelationships
[edit]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogeny of Radiodonta after Moysiuk & Caron 2021[39] |








Traditionally, all radiodont species have been placed within one family, Anomalocarididae,[6] hence the previous common name 'anomalocaridid'[26][8] and it was still occasionally used to refer the whole order even after reclassification.[9][10] Since the reassignment done by Vinther et al. 2014, most of the radiodont species were reclassified within three new families: Amplectobeluidae, Tamisiocarididae[2][3] (formerly Cetiocaridae[7]), and Hurdiidae.[7][10][2][3] Including Anomalocarididae, the four recent radiodont families may form the clade Anomalocarida.[7]
The original description of the order Radiodonta included Anomalocaris, Laggania (later known as Peytoia), Hurdia, Proboscicaris, Amplectobelua, Cucumericrus, and Parapeytoia.[6] However, Proboscicaris is now regarded as a junior synonym of Hurdia, and Parapeytoia is considered to be a Megacheiran.[8][27][10] Due to the limited discovery, The position of Cucumericrus within Radiodonta is unclear, as it was either unselected by phylogenetic analysis[7][3][2][39][40] or resolved in a polytomy with Radiodonta and Euarthropoda.[10][13]
 One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont.
One of the poorly-known body parts (trunk appendage) of Cucumericrus decoratus; this species may not represent a true radiodont.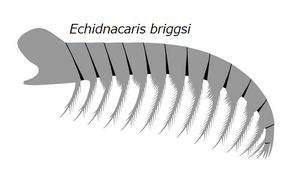 Frontal appendage of Echidnacaris briggsi, a tamisiocarid radiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.
Frontal appendage of Echidnacaris briggsi, a tamisiocarid radiodont that was once suggested to belong to the Anomalocaris genus until its description in 2023.


The first in-depth phylogenetic analysis of Radiodonta was conducted by Vinther et al. in 2014,[7] followed by a handful of subsequest studies with more or less modified results.[9][10][2][13][3][39][40][43] In most analysis, Caryosyntrips is the basal-most genus, but either resolved in a polytomy with other radiodonts and Euarthropoda (alongside Cucumericrus if included[10][13]) or outside of Radiodonta, casting doubt on its radiodont affinity.[64] With the exclusion of questionable Caryosyntrips and Cucumericrus, the monophyly of Radiodonta is widely supported,[7][9][10][2][13][3][39][40] with a few results suggest possible paraphyly (either the Anomalocarididae+Amplectobeluidae clade or Hurdiidae sister to Euarthropoda).[30][43] Putative synapomorphies of monophyletic Radiodonta including tripartite head sclerite complex and differentiated neck region.[3] The genus Anomalocaris in a broader sense always found to be polyphyletic, usually with "Anomalocaris" kunmingensis and "Anomalocaris" briggsi resolved as a member of Amplectobeluidae and Tamisiocarididae respectively.[7][9][10][2][3][39][40] Interrelationship of Amplectobeluidae is uncertain, as the amplectobeluid affinities of Lyrarapax and Ramskoeldia were occasionally questioned.[1][3][40] Monophyly of the speciose family Hurdiidae was recovered by most analysis and well-supported by several synapomorphies (e.g. distal articulated region of frontal appendage with proximal 5 podomeres bearing subequal endites[19][3]). Tamisiocarididae was often suggested to be sister group of Hurdiidae in 2010s,[7][10][2][3] but this position became questionable in subsequent studies.[22][24]
- Radiodonta
- ?Cucumericrus[26] (radiodont affinity questionable[65][64])
- ?Caryosyntrips[21] (radiodont affinity questionable[64])
- Anomalocarida
- Paranomalocaris[66] (placed within Anomalocarididae by some studies.[13][67])
- Laminacaris[68] (placed within Amplectobeluidae by some studies.[2])
- Houcaris (либо отнесенный к Anomalocarididae, [10] [13] [43] Амлектобелуиды [7] [2] или Tamisiocarididae. [69] [39] )
- Инновациикарис [70]
- Аномалокаридиды
- Anomalocaris (в более широком смысле некоторые виды могут быть отнесены к другим семействам. [7] [10] )
- Ленисикарис [20]
- Shucaris (отнесенный либо к Anomalocarididae, либо к Amplectobeluidae). [71] [64] )
- Амлектобелуиды
- Лирарапакс [9] (позиция подвергается сомнению в некоторых исследованиях. [1] )
- Эмблектобелуа [26]
- Рамскоэльдия [1] (позиция подвергается сомнению в некоторых исследованиях. [3] )
- Гуаншанкарис [67] [72]
- Shucaris (отнесенный либо к Anomalocarididae, либо к Amplectobeluidae). [71] [64] )
- Тамизиокаридиды
- Hurdiidae / Peytoiidae [64]
- Эгирокассисины [73]
- Пейтойя
- Шиндерханнес (позиция подвергается сомнению в некоторых исследованиях. [74] [75] [76] )
- Хурдия
- Стэнликарис [77]
- Пахвантия [2]
- Урсулинакарис [19]
- Камброрастер [3]
- ? Женгекарис (предполагаемый радиодонт хурдиид [65] )
- Вы убиты горем [78]
- Буккаспинея [79]
- Титанокорис [16]
| Разновидность | Исходное описание | Год назван | Семья | Возраст | Расположение | Лобный придаток | Головной склеритный комплекс |
|---|---|---|---|---|---|---|---|
| огурец Украшенный | Хоу, Бергстрем и Альберг | 1995 [26] | (не назначено) | Кембрийский этап 3 | Неизвестный | Неизвестный | |
| Кариосинтрипс зубчатый | Дэйли и Бадд | 2010 [21] | (не назначено) | Улиуань – Друмиан |  | Неизвестный | |
| Кариосинтрипс камурус | Пейтс и Дэйли | 2017 [41] | (не назначено) | Улиуань |  | Неполный [79] | |
| Кариосинтрипс твердый | Пейтс и Дэйли | 2017 [41] | (не назначено) | барабанщик |  | Неизвестный | |
| Параномалокарис многосегментарный | Ван, Хуан и Ху | 2013 [66] | Аномалокаридиды ? | Кембрийский этап 4 |  | Неизвестный | |
| Параномалокарис простой | Цзяо, Пейтс, Лерози-Обрил, Ортега-Эрнандес, Ян, Лан, Чжан | 2021 [67] | Аномалокаридиды ? | Кембрийский этап 4 |  | Неизвестный | |
| Ламинакарис химера | Го, Пейтс, Конг, Дейли, Эджкомб, Чен и Хоу | 2018 [68] | (спорный) | Кембрийский этап 3 |  | Неизвестный | |
| Инновациякарис маотианшаненсис | Цзэн, Чжао, Чжу | 2022 [70] | (не назначено) | Кембрийский этап 3 |  | P-элемент неизвестен [70] | |
| Инновации ? multispiniformis | Цзэн, Чжао, Чжу | 2022 [70] | (не назначено) | Кембрийский этап 3 | 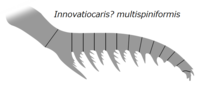 | Неизвестный | |
| Аномалокарис канадский | Уайтивс | 1892 [80] | Аномалокаридиды | Улиуань |  |  | |
| Lenisicaris pennsylvanica (ранее Anomalocaris pennsylvanica ) [20] | Сдерживать | 1929 | Аномалокаридиды | Кембрийский этап 3 |  | Неизвестный | |
| Ленисикарис тряпка | У, Ма, Линь, Сунь, Чжан и Фу | 2021 [20] | Аномалокаридиды | Кембрийский этап 3 |  | Неизвестный | |
| Аномалокарис далея | Патерсон, Гарсиа-Беллидоб и Эджкомб | 2023 | Аномалокаридиды | Кембрийский этап 4 |  | Неизвестный | |
| Houcaris magnabasis (ранее Anomalocaris magnabasis ) [69] | Пейтс, Дейли, Эджкомб, Конг и Либерман | 2019 | (спорный) | Кембрийский этап 4 |  | Неизвестный | |
| Houcaris saron (ранее Anomalocaris saron ) [69] | Хоу, Бергстрем и Альберг | 1995 | (спорный) | Кембрийский этап 3 |  | Неизвестный | |
| Эхиднакарис бриггси [24] | Не давай | 1995 | Тамизиокаридиды | Кембрийский этап 4 |  | Возможный H-элемент и уникальные латеральные склериты, связанные со сложными глазами [14] [24] | |
| Рамскоэльдия платьяканта | Конг, Эджкомб, Дейли, Го, Пейтс и Хоу | 2018 [1] | Амлектобелуиды | Кембрийский этап 3 |  | Неполный [1] | |
| Рамскоэльдиа похоже на | Конг, Эджкомб, Дейли, Го, Пейтс и Хоу | 2018 [1] | Амлектобелуиды | Кембрийский этап 3 |  | Неполный [1] | |
| Лирарапакс клауспинус | Конг, Ма, Хоу, Эджкомб и Штраусфилд | 2014 [9] | Амлектобелуиды | Кембрийский этап 3 |  | Горловина P-элемента неизвестна | |
| Лирарапакс трилобус | Конг, Дейли, Эджкомб, Хоу и Чен | 2016 [5] | Амлектобелуиды | Кембрийский этап 3 |  | P-элемент неизвестен | |
| Эмблектобелуа симбрациата | Хоу, Бергстрем и Альберг | 1995 [26] | Амлектобелуиды | Кембрийский этап 3 |  |  | |
| Эмблектобелуа стефененсис | Дэйли и Бадд | 2010 [21] | Амлектобелуиды | Улиуань |  | Неизвестный | |
| Гуаншанкарис кунмингенсис | Чжан и др. | 2023 [72] | Амлектобелуиды | Кембрийский этап 4 |  | Неизвестный | |
| Тамисиокарис северный | Дэйли и Пил | 2010 | Тамизиокаридиды | Кембрийский этап 3 |  | Неполный [7] | |
| Урсулинакарис grallae | Пейтс, Дейли и Баттерфилд | 2019 | Hurdiidae | Улиуань |  | Неизвестный | |
| Шиндерханнес Бартельси | Кюль, Бриггс и Раст | 2009 [35] | Hurdiidae | Эмсиан | Неполный [3] | Неполный [3] | |
| Стэнликарис гирпекс | Пейтс, Дейли и Ортега-Эрнандес | 2018 [77] | Hurdiidae | Улиуань |  | P-элемент неизвестен, возможно, отсутствует [22] | |
| Пейтоя натхорсти | Уолкотт | 1911 [81] | Hurdiidae | Улиуань – Друмиан |  | Неполный [3] | |
| Peytoia infercambriensis (ранее Cassubia infercambriensis ) [82] | Лендзион | 1975 | Hurdiidae | Кембрийский этап 3 |  | Неизвестный | |
| Эгирокассис бенмулай | Ван Рой, Дейли и Бриггс | 2015 [10] | Hurdiidae ( Aegirocassisinae ) | Тремадокийский |  |  | |
| Хурдия Виктория | Уолкотт | 1912 [83] | Hurdiidae | Улиуань – Друмиан |  |  | |
| Хурдия треугольная | Уолкотт | 1912 [83] | Hurdiidae | Улиуань | |  | |
| Камброрастером Скошено | Мойсюк и Карон | 2019 [3] | Hurdiidae | Улиуань |  |  | |
| Пахвантия пронзена копьем | Робисон и Ричардс | 1981 | Hurdiidae | барабанщик |  |  | |
| Кордатикарис полосатый | Сунь, Цзэн и Чжао | 2020 [78] | Hurdiidae | барабанщик | Неполный [78] |  | |
| Женгекарис Шанкоуэнсис | Ваннер, Чен, Хуан, Шарбонье и Ван | 2006 | Hurdiidae | Кембрийский этап 3 | Неизвестный |  | |
| я прикрыл рот | Пейтс, Лерози-Обрил, Дейли, Кир, Бонино и Ортега-Эрнандес | 2021 [79] | Hurdiidae | барабанщик |  | Неизвестный | |
| Титанокори гейнеси | Карон и Мойсюк | 2021 [40] | Hurdiidae | Улиуань |  |  | |
| Псевдоангустидонтус двойной остистый | Ван Рой и Тетли | 2006 | Hurdiidae ( Aegirocassisinae ) | Тремадокийский |  | Неизвестный | |
| Псевдоангустидонтус издигуа | Потин, Герио и Дейли | 2023 | Hurdiidae ( Aegirocassisinae ) | Тремадокийский |  | Неполный [73] | |
| Шукарис анкилосскелос | Ву и др. | 2024 | (спорный) | Кембрийский этап 2 – Кембрийский этап 3 |  | Неполный [71] |
История
[ редактировать ]
 Лобный придаток Anomalocaris canadensis
Лобный придаток Anomalocaris canadensis Оральный конус Peytoia nathorsti
Оральный конус Peytoia nathorsti H-элемент Hurdia victoria
H-элемент Hurdia victoria![Парные лобные придатки безымянного радиодонта-хурдиида[39]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg) Парные лобные придатки безымянного хурдиида . радиодонта- [39]
Парные лобные придатки безымянного хурдиида . радиодонта- [39]



![Парные лобные придатки безымянного радиодонта-хурдиида[39]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)
История радиодонтов сложна. Неполные экземпляры, относящиеся к разным частям тела одного и того же вида, исторически интерпретировались как принадлежащие к разным видам и даже разным типам. [6] [8] До того, как они были признаны группой, образцы радиодонтов были отнесены к пяти различным типам: Porifera , Cnidaria , Echinodermata , Annelida и Arthropoda . [6]
Первые известные образцы радиодонтов были собраны из пластов трилобитов горы Стивен Ричардом Г. МакКоннеллом из Геологической службы Канады в 1886 году. [6] или 1888 год. [80] Эти экземпляры были названы Anomalocaris canadensis в 1892 году палеонтологом GSC Джозефом Уайтавсом . [80] Уайтэйвс интерпретировал эти экземпляры, которые теперь известны как изолированные лобные придатки, как брюшко филлокарида ракообразного . [80] Дополнительные образцы радиодонтов были описаны в 1911 году Чарльзом Уолкоттом . [81] Изолированную ротовую шишку, которую он назвал Peytoia nathorsti , он интерпретировал как медузу, а плохо сохранившийся, но относительно полный экземпляр, который он назвал Laggania cambria , — как голотурию. [81] В 1912 году Уолкотт назвал Hurdia victoria и H. triangulata на основе изолированных H-элементов, которые он интерпретировал как панцири ракообразных. [83] Изолированные лобные придатки Peytoia и Hurdia были идентифицированы как придатки Sidneyia . , известные под общим названием «Придаток F» у Briggs 1979, в то время [81] Proboscicaris в P-элемент Hurdia был назван 1962 году и интерпретирован как панцирь двустворчатого членистоногого. [84]
Геологическая служба Канады инициировала пересмотр окаменелостей сланцев Бёрджесс в 1966 году под руководством Кембриджского университета палеонтолога Гарри Б. Уиттингтона . [6] Этот пересмотр в конечном итоге привел к открытию полного плана тела радиодонта. В 1978 году Саймон Конвей Моррис признал, что ротовой аппарат лаггании напоминал пейтою , но интерпретировал это как свидетельство того, что это было сложное ископаемое, состоящее из медузы пейтои и губки. [85] В 1979 году Дерек Бриггс признал, что окаменелости Anomalocaris были придатками, а не брюшком, но интерпретировал их как ходильные ноги рядом с «придатком F». [86] Лишь в 1985 году была признана истинная природа окаменелостей Anomalocaris , Laggania и Peytoia , и все они были отнесены к одному роду Anomalocaris . [33] Впоследствии было признано, что Anomalocaris представляет собой отдельную форму от двух других, что привело к разделению на два рода, последний из которых по-разному назывался Laggania и Peytoia, пока не было установлено, что оба представляют один и тот же вид, и Peytoia имеет приоритет. [23] Позже было признано, что некоторые окаменелости, отнесенные к этим таксонам, принадлежали другой форме, у которой был признан панцирь, состоящий из элементов Hurdia и Proboscicaris . Наконец, в 2009 году эти экземпляры были переописаны как Hurdia . [8] Даже после этих открытий частичные ошибочные идентификации (например, ротовой конус и лобные придатки пейтои) были отнесены к Anomalocaris. [6] и Хурдия , [8] соответственно) были выявлены и в последующих исследованиях. [23] [87]
Сам таксон Radiodonta был придуман в 1996 году Десмондом Коллинзом после того, как было установлено, что Anomalocaris и его родственники представляют собой особую линию, имеющую сходство с членистоногими, а не неизвестный до сих пор тип. [6] Коллинз также установил, что класс Dinocarida включает отряд Radiodonta, а также Opabiniidae , которые он признал отдельными из-за отсутствия у них характерной структуры орального конуса, свойственной радиодонтам. [6] Впервые Radiodonta получила филогенетическое определение в 2014 году. [7] Изначально считалось, что Radiodonta содержит одно семейство Anomalocarididae , но в 2014 году оно было разделено на четыре семейства: Amplectobeluidae , Anomalocarididae, Cetiocaridae и Hurdiidae . [7] Название Cetiocarididae не соответствовало Международному кодексу зоологической номенклатуры, поэтому в 2019 году было переименовано в Tamisiocarididae . [88]
До 2010-х годов радиодонты обычно считались крупными высшими хищниками, но открытие новых видов в течение этого десятилетия привело к значительному увеличению известного экологического и морфологического разнообразия группы. [7] [10] [2] [3] [89] [79] [39] [40]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к Конг, Пэй-Юнь; Эджкомб, Грегори Д.; Дейли, Эллисон С.; Го, Цзинь; Пейтс, Стивен; Хоу, Сянь-Гуан (2018). «Новые радиодонты с гнатобазоподобными структурами из кембрийской биоты Чэнцзян и значение для систематики Radiodonta» . Статьи по палеонтологии . 4 (4): 605–621. дои : 10.1002/spp2.1219 . ISSN 2056-2802 . S2CID 90258934 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Лерози-Обрил, Руди; Пейтс, Стивен (14 сентября 2018 г.). «Новый радиодонт, питающийся взвесью, предполагает эволюцию микропланктивных животных в кембрийском макронектоне» . Природные коммуникации . 9 (1): 3774. Бибкод : 2018NatCo...9.3774L . дои : 10.1038/s41467-018-06229-7 . ISSN 2041-1723 . ПМК 6138677 . ПМИД 30218075 . Данные дриад
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а Мойсюк Ю.; Карон, Ж.-Б. (14 августа 2019 г.). «Новый радиодонт-хурдиид из сланцев Бёрджесс свидетельствует об эксплуатации кембрийских инфаунистических источников пищи» . Труды Королевского общества B: Биологические науки . 286 (1908): 20191079. doi : 10.1098/rspb.2019.1079 . ПМК 6710600 . ПМИД 31362637 .
- ^ Jump up to: а б с д и ж г час Конг, Пэйюнь; Дейли, Эллисон С.; Эджкомб, Грегори Д.; Хоу, Сянгуан (30 августа 2017 г.). «Функциональная голова кембрийского радиодонта (стволовой группы Euarthropoda) Amplectobelua symbrachiata » . Эволюционная биология BMC . 17 (1): 208. дои : 10.1186/s12862-017-1049-1 . ISSN 1471-2148 . ПМК 5577670 . ПМИД 28854872 .
- ^ Jump up to: а б с д и ж Конг, Пэйюнь; Дейли, Эллисон С.; Эджкомб, Грегори Д.; Хоу, Сянгуан; Чен, Айлин (сентябрь 2016 г.). «Морфология радиодонтана Lyrarapax из биоты Чэнцзян раннего кембрия» . Журнал палеонтологии . 90 (4): 663–671. дои : 10.1017/jpa.2016.67 . ISSN 0022-3360 . S2CID 88742430 .
- ^ Jump up to: а б с д и ж г час я дж к л м Коллинз, Десмонд (1996). «Эволюция» Anomalocaris и его отнесение к классу членистоногих Dinocarida (nov.) и отряду Radiodonta (nov.)» . Журнал палеонтологии . 70 (2): 280–293. дои : 10.1017/S0022336000023362 . S2CID 131622496 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Винтер, Якоб; Штейн, Мартин; Лонгрич, Николас Р.; Харпер, Дэвид А.Т. (2014). «Аномалокарид, питающийся суспензией, из раннего кембрия» (PDF) . Природа . 507 (7493): 496–499. Бибкод : 2014Natur.507..496V . дои : 10.1038/nature13010 . ПМИД 24670770 . S2CID 205237459 .
- ^ Jump up to: а б с д и ж г час я дж Дейли, Эллисон С.; Бадд, Грэм Э.; Карон, Жан-Бернар; Эджкомб, Грегори Д.; Коллинз, Десмонд (2009). из сланца Берджесс «Аномалокаридид Hurdia и его значение для ранней эволюции членистоногих». Наука . 323 (5921): 1597–1600. Бибкод : 2009Sci...323.1597D . дои : 10.1126/science.1169514 . ПМИД 19299617 . S2CID 206517995 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Конг, Пэйюнь; Ма, Сяоя; Хоу, Сянгуан; Эджкомб, Грегори Д.; Штраусфельд, Николас Дж. (2014). «Структура мозга устраняет сегментарное сродство аномалокаридидных придатков». Природа . 513 (7519): 538–42. Бибкод : 2014Natur.513..538C . дои : 10.1038/nature13486 . ПМИД 25043032 . S2CID 4451239 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2015). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками». Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ПМИД 25762145 . S2CID 205242881 .
- ^ Jump up to: а б с д и ж г час Де Виво, Джачинто; Лаутеншлагер, Стефан; Винтер, Якоб (28 июля 2021 г.). «Трехмерное моделирование, неравенство и экология первых кембрийских хищников вершины» . Труды Королевского общества B: Биологические науки . 288 (1955): 20211176. doi : 10.1098/rspb.2021.1176 . ПМЦ 8292756 . ПМИД 34284622 .
- ^ Фостер, Дж. (2014). «Магический загадочный тур: биологическая психоделия сланцев Бёрджесс». Кембрийский океанский мир: древняя морская жизнь Северной Америки . Издательство Университета Индианы. стр. 195–252. ISBN 978-0253011824 .
- ^ Jump up to: а б с д и ж г час я дж Лю, Цзянни; Лерози-Обрил, Руди; Штайнер, Майкл; Данлоп, Джейсон А.; Шу, Деган; Патерсон, Джон Р. (01 ноября 2018 г.). «Происхождение хищного питания у молодых членистоногих, выявленное кембрийским радиодонтаном» . Национальный научный обзор . 5 (6): 863–869. дои : 10.1093/nsr/nwy057 . ISSN 2095-5138 .
- ^ Jump up to: а б с Пейтс, Стивен; Боттинг, Джозеф П.; Маккобб, Люси МЭ; Мьюир, Люси А. (2020). «Миниатюрная ордовикская гурдиида из Уэльса демонстрирует приспособляемость Radiodonta» . Королевское общество открытой науки . 7 (6): 200459. Бибкод : 2020RSOS....700459P . дои : 10.1098/rsos.200459 . ISSN 2054-5703 . ПМЦ 7353989 . ПМИД 32742697 .
- ^ Ву, Ю; Пейтс, Стивен; Поли, Дэниел; Чжан, Синлян; Фу, Дунцзин (03 ноября 2023 г.). «Быстрый рост крупного кембрийского хищника» . Национальный научный обзор . дои : 10.1093/nsr/nwad284 . ISSN 2095-5138 . ПМЦ 10833464 .
- ^ Jump up to: а б Карон, Ж.-Б.; Мойсюк Ю. (сентябрь 2021 г.). «Гигантский нектобентосный радиодонт из сланцев Берджесс и значение разнообразия панцирей хурдиид» . Королевское общество открытой науки . 8 (9): 210664. Бибкод : 2021RSOS....810664C . дои : 10.1098/rsos.210664 . ПМЦ 8424305 . ПМИД 34527273 .
- ^ Jump up to: а б с д и ж г час я Ортега-Эрнандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (01 мая 2017 г.). «Происхождение и эволюция головы панартропод - палеобиологическая перспектива и перспектива развития» . Строение и развитие членистоногих . Эволюция сегментации. 46 (3): 354–379. дои : 10.1016/j.asd.2016.10.011 . ISSN 1467-8039 . ПМИД 27989966 .
- ^ Jump up to: а б с д Цзэн, Хан; Чжао, Фанчэн; Ню, Кеченг; Чжу, Маоянь; Хуан, Диин (2020). «Раннекембрийский эуартропод с хищными придатками, похожими на радиодонтов» . Природа . 588 (7836): 101–105. Бибкод : 2020Natur.588..101Z . дои : 10.1038/s41586-020-2883-7 . ISSN 1476-4687 . ПМИД 33149303 . S2CID 226248177 .
- ^ Jump up to: а б с д и ж Пейтс, Стивен; Дейли, Эллисон С.; Баттерфилд, Николас Дж. (11 июня 2019 г.). «Первое сообщение о парных вентральных эндитах у хурдиидного радиодонта» . Зоологические письма . 5 (1): 18. дои : 10.1186/s40851-019-0132-4 . ISSN 2056-306X . ПМК 6560863 . ПМИД 31210962 .
- ^ Jump up to: а б с д и Ву, Ю; Ма, Цзясинь; Линь, Вэйлян; Солнце, АО; Чжан, Синлян; Фу, Дунцзин (01 мая 2021 г.). «Новые аномалокаридиды (Panarthropoda: Radiodonta) из нижнего кембрия Chengjiang Lagerstätte: биостратиграфические и палеобиогеографические последствия» . Палеогеография, Палеоклиматология, Палеоэкология . 569 : 110333. Бибкод : 2021PPP...569k0333W . дои : 10.1016/j.palaeo.2021.110333 . ISSN 0031-0182 . S2CID 233565727 .
- ^ Jump up to: а б с д и ж г час Дейли, Эллисон С.; Бадд, Грэм Э. (2010). «Новые придатки аномалокаридид из сланцев Бёрджесс, Канада» . Палеонтология . 53 (4): 721–738. Бибкод : 2010Palgy..53..721D . дои : 10.1111/j.1475-4983.2010.00955.x . ISSN 1475-4983 .
- ^ Jump up to: а б с д и ж г Мойсюк, Иосиф; Карон, Жан-Бернар (8 августа 2022 г.). «Трехглазый радиодонт с окаменелой нейроанатомией сообщает о происхождении головы и сегментации членистоногих» . Современная биология . 32 (15): 3302–3316.e2. дои : 10.1016/j.cub.2022.06.027 . ISSN 0960-9822 . ПМИД 35809569 . S2CID 250361698 .
- ^ Jump up to: а б с д и ж Дейли, Эллисон С.; Бергстрем, Ян (апрель 2012 г.). «Ротовая шишка Anomalocaris — это не классическая пейтоя » . Naturwissenschaften . 99 (6): 501–504. Бибкод : 2012NW.....99..501D . дои : 10.1007/s00114-012-0910-8 . ISSN 0028-1042 . ПМИД 22476406 . S2CID 2042726 .
- ^ Jump up to: а б с д Патерсон, Джон Р.; Гарсиа-Беллидо, Диего К.; Эджкомб, Грегори Д. (1 января 2023 г.). «Возвращение к раннекембрийским радиодонтам сланцев Эму Бэй: морфология и систематика» . Журнал систематической палеонтологии . 21 (1). дои : 10.1080/14772019.2023.2225066 . ISSN 1477-2019 . S2CID 259719252 .
- ^ Jump up to: а б Штраусфельд, Николас Дж.; Ма, Сяоя; Эджкомб, Грегори Д.; Форти, Ричард А.; Лэнд, Майкл Ф.; Лю, Ю; Конг, Пэйюнь; Хоу, Сянгуан (август 2015 г.). «Глаза членистоногих: летопись окаменелостей раннего кембрия и дивергентная эволюция зрительных систем» . Строение и развитие членистоногих . 45 (2): 152–172. дои : 10.1016/j.asd.2015.07.005 . ПМИД 26276096 .
- ^ Jump up to: а б с д и ж г Сиань-Гуан, Хоу; Бергстрем, Ян; Альберг, Пер (сентябрь 1995 г.). « Anomalocaris и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая». ГФФ . 117 (3): 163–183. дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Jump up to: а б с д и ж г час я Дейли, Эллисон С.; Эджкомб, Грегори Д. (2014). «Морфология Anomalocaris canadensis из сланцев Берджесс». Журнал палеонтологии . 88 (1): 68–91. дои : 10.1666/13-067 . S2CID 86683798 .
- ^ Дейли, Эллисон С.; Патерсон, Джон Р.; Эджкомб, Грегори Д.; Гарсиа-Беллидо, Диего К.; Джаго, Джеймс Б. (2013). «Новая анатомическая информация об Anomalocaris из кембрийских сланцев залива Эму в Южной Австралии и переоценка его предполагаемых хищнических привычек» . Палеонтология . 56 (5): 971–990. дои : 10.1111/пала.12029 . ISSN 1475-4983 . S2CID 128955512 .
- ^ Jump up to: а б с Чен, Цзюнь-юань; Рамшельд, Ларс; Чжоу, Гуй-цин (27 мая 1994 г.). «Доказательства монофилии и близости кембрийских гигантских хищников к членистоногим» . Наука . 264 (5163): 1304–1308. Бибкод : 1994Sci...264.1304C . дои : 10.1126/science.264.5163.1304 . ПМИД 17780848 . S2CID 1913482 .
- ^ Jump up to: а б с Ария, Седрик; Чжао, Фанчэн; Цзэн, Хан; Го, Цзинь; Чжу, Маоянь (08 января 2020 г.). «Окаменелости из Южного Китая дают новое определение строению тела предков эуартропод» . Эволюционная биология BMC . 20 (1): 4. дои : 10.1186/s12862-019-1560-7 . ISSN 1471-2148 . ПМК 6950928 . ПМИД 31914921 .
- ^ Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2013). Аномалокаридиды имели два набора боковых створок . 57-е ежегодное собрание Палеонтологической ассоциации. Цюрих, Швейцария.
- ^ Jump up to: а б Ваннье, Жан; Лю, Цзянни; Лерози-Обрил, Руди; Винтер, Якоб; Дэйли, Эллисон К. (2 мая 2014 г.). «Сложная пищеварительная система у ранних членистоногих» . Природные коммуникации . 5 (1): 3641. Бибкод : 2014NatCo...5.3641V . дои : 10.1038/ncomms4641 . ISSN 2041-1723 . ПМИД 24785191 .
- ^ Jump up to: а б с Уиттингтон, Гарри Блэкмор; Бриггс, Дерек Эрнест Гилмор (14 мая 1985 г.). «Самое крупное кембрийское животное, Anomalocaris , Берджесс-Шейл, Британская Колумбия» . Философские труды Лондонского королевского общества. Б. Биологические науки . 309 (1141): 569–609. Бибкод : 1985RSPTB.309..569W . дои : 10.1098/rstb.1985.0096 .
- ^ Усами, Ёсиюки (7 января 2006 г.). «Теоретическое исследование формы тела и схемы плавания Anomalocaris на основе гидродинамического моделирования». Журнал теоретической биологии . 238 (1): 11–17. Бибкод : 2006JThBi.238...11U . дои : 10.1016/j.jtbi.2005.05.008 . ISSN 0022-5193 . ПМИД 16002096 .
- ^ Jump up to: а б с Кюль, Габриэле; Бриггс, Дерек Э.Г.; Раст, Джес (6 февраля 2009 г.). «Членистоногие с большим придатком и радиальной пастью из сланца Хунсрюк нижнего девона, Германия». Наука . 323 (5915): 771–773. Бибкод : 2009Sci...323..771K . дои : 10.1126/science.1166586 . ISSN 0036-8075 . ПМИД 19197061 . S2CID 47555807 .
- ^ Шеппард, Калифорния; Соперник, Делавэр; Карон, Ж.-Б. (01.10.2018). «К гидродинамике хвостового плавника Anomalocaris» . Интегративная и сравнительная биология . 58 (4): 703–711. дои : 10.1093/icb/icy014 . HDL : 1974/22737 . ISSN 1540-7063 . ПМИД 29697774 .
- ^ Дейли, Эллисон; Драге, Харриет (01 сентября 2015 г.). «Ископаемая летопись шелушения и тенденции в поведении трилобитов при линьке» . Строение и развитие членистоногих . 45 (2): 71–96. дои : 10.1016/j.asd.2015.09.004 . ПМИД 26431634 .
- ^ Jump up to: а б с д Де Виво, Джачинто; Лаутеншлагер, Стефан; Винтер, Якоб (16 декабря 2016 г.). «Реконструкция ловкости пищевых придатков аномалокаридид проливает свет на экологию радиодонтана» .
- ^ Jump up to: а б с д и ж г час я дж к Мойсюк, Иосиф; Карон, Жан-Бернар (2021). «Исключительная многофункциональность в питающем аппарате среднекембрийского радиодонта» . Палеобиология . 47 (4): 704–724. Бибкод : 2021Pbio...47..704M . дои : 10.1017/pab.2021.19 . ISSN 0094-8373 . S2CID 236552819 .
- ^ Jump up to: а б с д и ж г час я дж Карон, Ж.-Б.; Мойсюк, Ю. (2021). «Гигантский нектобентосный радиодонт из сланцев Берджесс и значение разнообразия панцирей хурдиид» . Королевское общество открытой науки . 8 (9): 210664. Бибкод : 2021RSOS....810664C . дои : 10.1098/rsos.210664 . ПМЦ 8424305 . ПМИД 34527273 .
- ^ Jump up to: а б с Пейтс, С.; Дейли, AC (2017). « Кариосинтрипс : радиодонтан из кембрия Испании, США и Канады» . Статьи по палеонтологии . 3 (3): 461–470. дои : 10.1002/spp2.1084 . ISSN 2056-2802 . S2CID 135026011 .
- ^ Jump up to: а б с д и ж г час Ортега-Эрнандес, Хавьер (декабрь 2014 г.). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры Кембриджского философского общества . 91 (1): 255–273. дои : 10.1111/brv.12168 . ISSN 1469-185Х . ПМИД 25528950 . S2CID 7751936 .
- ^ Jump up to: а б с д и ж г час Пейтс, Стивен; Вулф, Джоанна М.; Лерози-Обрил, Руди; Дейли, Эллисон С.; Ортега-Эрнандес, Хавьер (09 февраля 2022 г.). «Новый опабиниид разнообразит самые странные чудеса стволовой группы эуартропод» . Труды Королевского общества B: Биологические науки . 289 (1968): 20212093. doi : 10.1098/rspb.2021.2093 . ПМЦ 8826304 . ПМИД 35135344 .
- ^ Эджкомб, Грегори Д. (01 марта 2010 г.). «Филогения членистоногих: обзор с точки зрения морфологии, молекулярных данных и летописи окаменелостей» . Строение и развитие членистоногих . Ископаемая летопись и филогения членистоногих. 39 (2): 74–87. дои : 10.1016/j.asd.2009.10.002 . ISSN 1467-8039 . ПМИД 19854297 .
- ^ Легг, Дэвид; Саттон, Марк; Эджкомб, Грегори; Карон, Жан-Бернар (10 октября 2012 г.). «Кембрийские двустворчатые членистоногие раскрывают происхождение артродизации» . Слушания. Биологические науки . 279 (1748): 4699–4704. дои : 10.1098/рспб.2012.1958 . ПМЦ 3497099 . ПМИД 23055069 .
- ^ Jump up to: а б Легг, Дэвид (2013). «Многочлениковые членистоногие из среднего кембрия Британской Колумбии (Канада)» . Журнал палеонтологии . 87 (3): 493–501. дои : 10.1666/12-112.1 . ISSN 0022-3360 . S2CID 86725173 .
- ^ Легг, Дэвид; Саттон, Марк; Эджкомб, Грегори (30 сентября 2013 г.). «Данные об окаменелостях членистоногих повышают соответствие морфологической и молекулярной филогении» . Природные коммуникации . 4 : 2485. Бибкод : 2013NatCo...4.2485L . дои : 10.1038/ncomms3485 . ПМИД 24077329 .
- ^ Легг, Дэвид А.; Ваннье, Жан (2013). «Сродство космополитических членистоногих Isoxys и его значение для происхождения членистоногих» . Летайя . 46 (4): 540–550. дои : 10.1111/лет.12032 . ISSN 1502-3931 .
- ^ О'Флинн, Роберт Дж.; Уильямс, Марк; Ю, Мэнсяо; Харви, Томас HP; Лю, Ю (2022). «Новый эуартропод с крупными лобными придатками из биоты Чэнцзян раннего кембрия» . Электронная палеонтология . 25 (1): 1–21. дои : 10.26879/1167 . ISSN 1094-8074 . S2CID 246779634 .
- ^ Jump up to: а б с Фу, Дунцзин; Легг, Дэвид А.; Дейли, Эллисон С.; Бадд, Грэм Э.; Ву, Ю; Чжан, Синлян (28 марта 2022 г.). «Эволюция двуветвистых придатков, выявленная кембрийским членистоногим с панцирем» . Философские труды Королевского общества B: Биологические науки . 377 ): 20210034.doi : ( 1847 10.1098/rstb.2021.0034 . ПМЦ 8819368 . ПМИД 35125000 . S2CID 246608509 .
- ^ Ортега-Эрнандес, Хавьер (15 июня 2015 г.). «Гомология головных склеритов у членистоногих сланцев Берджесс» . Современная биология . 25 (12): 1625–1631. дои : 10.1016/j.cub.2015.04.034 . ISSN 0960-9822 . ПМИД 25959966 .
- ^ Jump up to: а б Гирибет, Гонсало; Эджкомб, Грегори Д. (17 июня 2019 г.). «Филогения и эволюционная история членистоногих» . Современная биология . 29 (12): Р592–Р602. дои : 10.1016/j.cub.2019.04.057 . ISSN 0960-9822 . ПМИД 31211983 . S2CID 189926344 .
- ^ Jump up to: а б Эджкомб, Грегори Д. (2 ноября 2020 г.). «Происхождение членистоногих: объединение палеонтологических и молекулярных данных» . Ежегодный обзор экологии, эволюции и систематики . 51 (1): 1–25. doi : 10.1146/annurev-ecolsys-011720-124437 . ISSN 1543-592X . S2CID 225478171 .
- ^ Jump up to: а б Цзунцзе, Чжанхэ, Чжань ; Сяндун, Ван Сянгуан, Хоу; Ян Цзяю , Ин ;
- ^ Бадд, Грэм Э. (1998). «Морфология и филогенетическое значение Kerygmachela kierkegaardi Budd (формация Буэн, нижний кембрий, Северная Гренландия)» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 89 (4): 249–290. дои : 10.1017/S0263593300002418 . ISSN 1473-7116 . S2CID 85645934 .
- ^ Янг, Флетчер Дж.; Винтер, Якоб (2017). «Онихофороподобная миоанатомия кембрийского жаберного лобопода Pambdelurion Whittingtoni» . Палеонтология . 60 (1): 27–54. Бибкод : 2017Palgy..60...27Y . дои : 10.1111/пала.12269 . hdl : 1983/92180ef0-2205-4c65-9a70-90d59cfea2f4 . ISSN 1475-4983 . S2CID 55477207 .
- ^ Эджкомб, Грегори Д. (2010). «Филогения членистоногих: обзор с точки зрения морфологии, молекулярных данных и летописи окаменелостей» . Строение и развитие членистоногих . 39 (2–3): 74–87. дои : 10.1016/j.asd.2009.10.002 . ISSN 1467-8039 . ПМИД 19854297 .
- ^ Дейли, Эллисон С.; Антклифф, Джонатан Б.; Дрейдж, Харриет Б.; Пейтс, Стивен (22 мая 2018 г.). «Ранние окаменелости Euarthropoda и кембрийский взрыв» . Труды Национальной академии наук . 115 (21): 5323–5331. Бибкод : 2018PNAS..115.5323D . дои : 10.1073/pnas.1719962115 . ISSN 0027-8424 . ПМК 6003487 . ПМИД 29784780 .
- ^ Сиань-Гуан, Хоу; Бергстрем, Ян; Альберг, Пер (сентябрь 1995 г.). «Аномалокарис и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая» . ГФФ . 117 (3): 163–183. дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Хауг, Иоахим Т.; Валошек, Дитер; Маас, Андреас; Лю, Ю; Хауг, Кэролин (март 2012 г.). «Функциональная морфология, онтогенез и эволюция креветкообразных хищников-богомолов в кембрии: КРЕВЕТКООБОДНЫЕ КЕМБРИЙСКИЕ ХИЩНИКИ-богомолы» . Палеонтология . 55 (2): 369–399. дои : 10.1111/j.1475-4983.2011.01124.x . S2CID 82841481 .
- ^ Смит, Мартин Р.; Карон, Жан-Бернар (июнь 2015 г.). «Голова галлюцигении и глоточный каркас ранних экдизозоев» (PDF) . Природа . 523 (7558): 75–78. Бибкод : 2015Natur.523...75S . дои : 10.1038/nature14573 . ISSN 1476-4687 . ПМИД 26106857 . S2CID 205244325 .
- ^ Танака, Генго; Хоу, Сянгуан; Ма, Сяоя; Эджкомб, Грегори Д.; Штраусфельд, Николас Дж. (октябрь 2013 г.). «Хелицератный рисунок нервного поля у кембрийского членистоногого с большим придатком». Природа . 502 (7471): 364–367. Бибкод : 2013Natur.502..364T . дои : 10.1038/nature12520 . ISSN 1476-4687 . ПМИД 24132294 . S2CID 4456458 .
- ^ Ортега-Эрнандес, Хавьер; Лерози-Обрил, Руди; Пейтс, Стивен (18 декабря 2019 г.). «Склонность к сохранению нервной системы в кембрийских отложениях типа сланцев Бёрджесс» . Труды Королевского общества B: Биологические науки . 286 (1917): 20192370. doi : 10.1098/rspb.2019.2370 . ПМК 6939931 . ПМИД 31822253 .
- ^ Jump up to: а б с д и ж МакКолл, Кристиан Р.А. (13 декабря 2023 г.). «Большой пелагический лобопод из кембрийских сланцев Пиоче в Неваде» . Журнал палеонтологии : 1–16. дои : 10.1017/jpa.2023.63 . ISSN 0022-3360 .
- ^ Jump up to: а б Цзэн, Хан; Чжао, Фанчэн; Инь, Цзунцзюнь; Чжу, Маоянь (2 января 2018 г.). «Морфология разнообразных склеритов головы радиодонтана из раннего кембрия Чэнцзян Лагерштетте, юго-запад Китая» . Журнал систематической палеонтологии . 16 (1): 1–37. дои : 10.1080/14772019.2016.1263685 . ISSN 1477-2019 . S2CID 133549817 .
- ^ Jump up to: а б Юаньюань, Ван; Хуан, Диин; Ху, Шисюэ (01 ноября 2013 г.). «Новые лобные придатки аномалокардид из биоты Гуаньшань, восточная часть Юньнани» . Китайский научный бюллетень . 58 (32): 3937–3942. Бибкод : 2013ЧСБу..58.3937В . дои : 10.1007/s11434-013-5908-x . S2CID 83910779 .
- ^ Jump up to: а б с Цзяо, Дэ-гуан; Пейтс, Стивен; Лерози-Обрил, Руди; Ортега-Эрнандес, Хавьер; Ян, Цзе; Лан, Тиан; Чжан, Си-гуан (2021). «Эндемичные радиодонты кембрийской биоты Гуаньшаня 4-го этапа Южного Китая» . Acta Palaeontologica Polonica . 66 . дои : 10.4202/app.00870.2020 . ISSN 0567-7920 . S2CID 236683029 .
- ^ Jump up to: а б Го, Дж.; Пейтс, С.; Конг, П.; Дейли, AC; Эджкомб, Джорджия; Чен, Т.; Хоу, X. (2018). «Новый лобный придаток радиодонта (ствол Euarthropoda) с мозаикой признаков из кембрийской биоты (серия 2 этап 3) Чэнцзян» . Статьи по палеонтологии . 5 (1). ISSN 2056-2799 .
- ^ Jump up to: а б с Ву, Ю; Фу, Дунцзин; Ма, Цзясинь; Линь, Вэйлян; Солнце, АО; Чжан, Синлян (01.06.2021). « Houcaris gen. nov. из раннего кембрия (этап 3) Chengjiang Lagerstätte расширил палеогеографическое распространение тамизиокаридид (Panarthropoda: Radiodonta)» . ПалЗ . 95 (2): 209–221. дои : 10.1007/s12542-020-00545-4 . ISSN 1867-6812 . S2CID 235221043 .
- ^ Jump up to: а б с д Цзэн, Хан; Чжао, Фанчэн; Чжу, Маоянь (07 сентября 2022 г.). « Innovatiocaris , полный радиодонт из раннего кембрия Chengjiang Lagerstätte и его значение для филогении Radiodonta» . Журнал Геологического общества . 180 . дои : 10.1144/jgs2021-164 . ISSN 0016-7649 . S2CID 252147346 .
- ^ Jump up to: а б с Ву, Ю; Пейтс, Стивен; Лю, Конг; Чжан, Минцзин; Линь, Вэйлян; Ма, Цзясинь; Ву, Юхэн; Чай, Шу; Чжан, Синлян; Фу, Дунцзин (31 декабря 2024 г.). «Новый радиодонт из нижнего кембрия (серия 2, стадия 3) Чэнцзян Лагерштетте, Южный Китай, сообщает об эволюции структур питания у радиодонтов» . Журнал систематической палеонтологии . 22 (1). дои : 10.1080/14772019.2024.2364887 . ISSN 1477-2019 .
- ^ Jump up to: а б Чжан, Минцзин; Ву, Ю; Линь, Вэйлян; Ма, Цзясинь; Ву, Юхэн; Фу, Дунцзин (11 апреля 2023 г.). «Amplectobeluid Radiodont Guanshancaris gen. nov. из нижнего кембрия (этап 4) Гуаньшань Лагерштетте в Южном Китае: биостратиграфические и палеобиогеографические последствия» . Биология . 12 (4): 583. doi : 10.3390/biology12040583 . ISSN 2079-7737 . ПМЦ 10136193 . ПМИД 37106783 .
- ^ Jump up to: а б Потин, Гаэтан Ж.-М.; Герио, Пьер; Дейли, Эллисон К. (2023). «Лобные придатки радиодонтов из биоты Фезуата (Марокко) демонстрируют большое разнообразие и экологическую адаптацию к питанию взвесью в раннем ордовике» . Границы экологии и эволюции . 11 . дои : 10.3389/fevo.2023.1214109 . ISSN 2296-701X .
- ^ Ортега-Эрнандес, Хавьер (декабрь 2014 г.). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры Кембриджского философского общества . 91 (1): 255–273. дои : 10.1111/brv.12168 . ISSN 1469-185Х . ПМИД 25528950 . S2CID 7751936 .
- ^ Чжу, X.; Лерози-Обрил, Р.; Ортега-Эрнандес, Х. (2021). «Фуронгские (цзяншаньские) находки радиодонтов в Польше и Южном Китае и летопись окаменелостей Hurdiidae» . ПерДж . 9 . е11800. дои : 10.7717/peerj.11800 . ПМЦ 8312493 . ПМИД 34386302 .
- ^ Потин, Гаэтан Ж.-М.; Дейли, Эллисон К. (2023). «Значение Anomalocaris и других Radiodonta для понимания палеоэкологии и эволюции во время кембрийского взрыва» . Границы в науках о Земле . 11 . Бибкод : 2023FrEaS..1160285P . дои : 10.3389/feart.2023.1160285 . ISSN 2296-6463 .
- ^ Jump up to: а б Пейтс, Стивен; Дейли, Эллисон; Ортега-Эрнандес, Хавьер (3 марта 2018 г.). Ответ на комментарий к записи «Aysheaia prolata из формации Юта Уилер (Драмиан, кембрий) представляет собой лобный придаток радиодонтана Stanleycaris» с формальным описанием Stanleycaris .
- ^ Jump up to: а б с Сунь, Чжисинь; Цзэн, Хан; Чжао, Фанчэн (01 августа 2020 г.). «Новый радиодонт среднего кембрия из Северного Китая: последствия морфологического неравенства и пространственного распределения хурдиид» . Палеогеография, Палеоклиматология, Палеоэкология . 558 : 109947. Бибкод : 2020PPP...558j9947S . дои : 10.1016/j.palaeo.2020.109947 . ISSN 0031-0182 . S2CID 224868404 .
- ^ Jump up to: а б с д Пейтс, Стивен; Лерози-Обрил, Руди; Дейли, Эллисон С.; Кир, Карло; Бонино, Энрико; Ортега-Эрнандес, Хавьер (19 января 2021 г.). «Разнообразная фауна радиодонтов из формации Марджум в штате Юта, США (кембрий: драмиан)» . ПерДж . 9 : е10509. дои : 10.7717/peerj.10509 . ISSN 2167-8359 . ПМЦ 7821760 . ПМИД 33552709 .
- ^ Jump up to: а б с д Уайтэйвс, Дж. Ф. (1892). «Описание нового рода и вида филлокарид ракообразных из среднего кембрия горы Стивен, Британская Колумбия». Канадский рекорд науки . 5 (4).
- ^ Jump up to: а б с д Уолкотт, CD 1911a. Голотурии и медузы среднего кембрия . Кембрийская геология и палеонтология II. Разные коллекции Смитсоновского института, 57: 41–68.
- ^ Дейли, Эллисон; Легг, Дэвид (2 июля 2015 г.). «Морфологическая и таксономическая оценка древнейшего аномалокаридида из нижнего кембрия Польши» . Геологический журнал . -1 (5): 949–955. Бибкод : 2015ГеоМ..152..949Д . дои : 10.1017/S0016756815000412 . S2CID 130745134 .
- ^ Jump up to: а б с УОЛКОТТ, CD 1912. Среднекембрийские Branchiopoda, Malacostraca, Trilobita и Merostomata . Разные коллекции Смитсоновского института, 57: 145–228.
- ^ РОЛЬФ, WDI 1962. Два новых панциря членистоногих из сланцев Бёрджесс (средний кембрий) в Канаде. Музей сравнительной зоологии Бревиоры, 60: 1–9.
- ^ Конвей Моррис, С. (1978). «Лаггания камбрия Уолкотт: сложное ископаемое». Журнал палеонтологии . 52 (1): 126–131. JSTOR 1303799 .
- ^ Бриггс, DEG (1979). «Аномалокарис, крупнейшее известное кембрийское членистоногое». Палеонтология . 22 (3): 631–664.
- ^ Дейли, Эллисон; Бадд, Грэм; Карон, Жан-Бернар (01 октября 2013 г.). «Морфология и систематика аномалокарида Hurdia из среднего кембрия Британской Колумбии и Юты» . Журнал систематической палеонтологии . 11 . дои : 10.1080/14772019.2012.732723 . S2CID 86465719 .
- ^ Пейтс, Стивен; Дейли, Эллисон К. (2019). «Формация Кинзерс (Пенсильвания, США): наиболее разнообразный комплекс радиодонтов кембрийского этапа 4» . Геологический журнал . 156 (7): 1233–1246. Бибкод : 2019GeoM..156.1233P . дои : 10.1017/S0016756818000547 . S2CID 134299859 .
- ^ Патерсон, Джон Р.; Эджкомб, Грегори Д.; Гарсиа-Беллидо, Диего К. (2020). «Несопоставимые сложные глаза кембрийских радиодонтов свидетельствуют об их способе развития и разнообразной визуальной экологии» . Достижения науки . 6 (49). Бибкод : 2020SciA....6.6721P . дои : 10.1126/sciadv.abc6721 . hdl : 10141/622906 . ПМЦ 7821881 . ПМИД 33268353 .
