Аномалокарис
| Аномалокарис | |
|---|---|
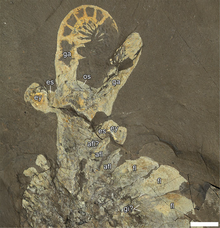 | |
| РОМИП 51212, практически полный экземпляр Anomalocaris canadensis . | |
 | |
| Восстановление жизни Anomalocaris canadensis . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Сорт: | † Динокаридид |
| Заказ: | † Радиодонта |
| Семья: | † Аномалокаридиды |
| Род: | † Аномалокарис Уайтэйвс , 1892 г. |
| Разновидность | |
(еще 8 безымянных видов [3] ) | |
Anomalocaris («в отличие от других креветок», или «аномальные креветки») — вымерший род радиодонтов , отряда рано расходящихся членистоногих группы стеблевой .
Он наиболее известен из типового вида A. canadensis, обнаруженного в формации Стивен (особенно в сланцах Бёрджесс ) в Британской Колумбии , Канада . Другой названный вид A. daleyae известен из несколько более древних сланцев залива Эму в Австралии . [2] Другие безымянные виды Anomalocaris известны из Китая и США . [3]
Как и у других радиодонтов, у Anomalocaris были плавательные створки, расположенные вдоль тела, большие сложные глаза и одна пара сегментированных лобных придатков , которые у Anomalocaris использовались для захвата добычи. По оценкам, длина достигает 34,2–37,8 см (1,12–1,24 фута), исключая лобные придатки и хвостовой веер. [4] Аномалокарис — одно из крупнейших животных кембрия и считается одним из самых ранних примеров высшего хищника . [5] [6] хотя другие были найдены в более старых кембрийских отложениях лагерштеттена .
С момента первоначального описания в конце 19 века, [7] лобные придатки были единственными известными окаменевшими частями, ошибочно принятыми за части тела других животных. [8] Его родство с радиодонтами было обнаружено в 1980-х годах, в частности, в журнальной статье 1985 года Гарри Б. Уиттингтона и Дерека Бриггса . [9] Туловище и рот реконструировались по другому роду радиодонтов до внесения исправлений в 1996 г. [8] и 2012. [10] Это типовой род Anomalocarididae , семейства, которое ранее включало всех радиодонтов, но в последнее время только Anomalocaris и несколько близкородственных таксонов. [3]
Открытие и идентификация
[ редактировать ]

С самого начала окаменелость Anomalocaris была ошибочно идентифицирована, после чего последовала серия ошибочных идентификаций и таксономических пересмотров. [9] [8] Как правильно описал Стивен Джей Гулд , популяризировавший кембрийский взрыв в своей книге 1989 года «Чудесная жизнь» :
[История Аномалокариса — это] рассказ о юморе, ошибках, борьбе, разочаровании и других ошибках, кульминацией которого является необычайное решение, объединившее кусочки трех «типов» в одно реконструированное существо, самое большое и свирепое существо кембрийского периода. организмы. [11]
Окаменелости аномалокариса были впервые собраны в 1886 году. [8] Ричард Г. МакКоннелл из Геологической службы Канады (GSC). Узнав о богатых окаменелостях в формации Стивен в Британской Колумбии, МакКоннелл поднялся на гору Стивен . 13 сентября 1886 года [12] [13] Он нашел множество трилобитов, а также два неизвестных экземпляра. [7] В августе 1891 года Анри-Марк Ами , помощник палеонтолога GSC, собрал множество трилобитов и брахиопод . окаменелостей [14] вместе с еще 48 неизвестными экземплярами. [15] Пятьдесят экземпляров были исследованы и описаны в 1892 году палеонтологом GSC Джозефом Фредериком Уайтэйвзом . [16] [17] Уайтэйвс интерпретировал их как брюшки -филлокарид ракообразных и дал полное научное название Anomalocaris canadensis . Он описывает ракообразных персонажей:
Сегменты тела или брюшка, которые у всех собранных экземпляров аномально уплощены с боков, немного выше или глубже длины, сверху шире, чем снизу, от каждого отходят пары вентральных придатков, почти равные по высоте или глубине самому сегменту. ... Родовое название Anomalocaris (от ανώμαλος , в отличие от — καρίς , креветка, т. е . в отличие от других других креветок) [видовое название, относящееся к Канаде] подсказывает необычная форма уроподов или вентральных придатков сегментов тела. и относительное положение каудального отдела позвоночника. [7]
В 1928 году датский палеонтолог Кай Хенриксен предположил, что Тузоя , членистоногое из сланцев Берджесс, известное только по панцирю, представляет собой недостающую переднюю половину Anomalocaris . [8] Художники Эли Шеверланж и Чарльз Р. Найт следовали этой интерпретации в своих изображениях Аномалокариса . [8]
Неизвестные в то время учёным части тела родственников Anomalocaris уже были описаны, но не признаны таковыми. Первый окаменелый рот такого вида животного обнаружил Чарльз Дулиттл Уолкотт , который принял его за медузу и отнес к роду Peytoia . Уолкотт также обнаружил лобный придаток, но не смог осознать сходство с открытием Уайтэйвса и вместо этого идентифицировал его как питающий придаток или хвост сосуществовавших Sidneyia . [18] В той же публикации, в которой он назвал Peytoia , Уолкотт назвал Laggania , таксон, который он интерпретировал как голотурию .
В 1966 году Геологическая служба Канады начала комплексный пересмотр летописи окаменелостей сланцев Бёрджесс под руководством Кембриджского университета палеонтолога Гарри Б. Уиттингтона . [8] В процессе этого пересмотра Уиттингтон и его ученики Саймон Конвей Моррис и Дерек Бриггс откроют истинную природу Anomalocaris и его родственников, но не без того, чтобы внести свой вклад в историю неправильных интерпретаций. [18] В 1978 году Конвей Моррис признал, что ротовой аппарат лаггании идентичен пейтое , но пришел к выводу, что лаггания представляет собой сложное ископаемое, состоящее из пейтои и губки Corralio undulata . [19] В 1979 году Бриггс признал, что окаменелости Anomalocaris представляли собой придатки, а не брюшки, и предположил, что это ходильные ноги гигантского членистоногого, а пищевой придаток, который Уолкотт определил для Сиднеи, был пищевым придатком аналогичного животного, называемого «придаток F». [16] Позже, очищая то, что, по его мнению, было неродственным экземпляром, Гарри Б. Уиттингтон удалил слой покрывающего камня и обнаружил однозначно связанный лобный придаток, идентичный Anomalocaris , и ротовой аппарат, похожий на Peytoia . [18] [20] Уиттингтон связал эти два вида, но исследователям потребовалось еще несколько лет, чтобы понять, что постоянно соседствующие пейтоя , лаггания и лобные придатки ( аномалокарис и «придаток F») на самом деле представляют собой единую группу огромных существ. [9] Эти два рода теперь объединены в отряд Radiodonta. [8] и широко известны как радиодонты или аномалокаридиды. Поскольку Пейтоя была названа первой, это общепринятое правильное название для всего животного. Однако первоначальный лобный придаток принадлежал более крупному виду, отличному от Peytoia и Laggania , и поэтому сохранил название Anomalocaris . [10]
В 2011 году сложные глаза Anomalocaris в были обнаружены во время палеонтологических раскопок в сланцах Emu Bay Shale на острове Кенгуру Австралии, что доказывает, что Anomalocaris действительно был членистоногим , как и предполагалось. Находка также показала, что глаза продвинутых членистоногих развились очень рано, до появления суставчатых ног или затвердевшего экзоскелета. [21] Позже этот экземпляр был идентифицирован как представитель нового вида Anomalocaris , A. daleyae . [2]
Многие виды ранее были отнесены к Anomalocaris , но последующие анализы поставили под сомнение это родовое отнесение. [22] [23] [24] и переклассифицировал их в разные роды. В 2021 году « А. » Сарон [25] и « А. » магнабасис [26] были переотнесены к новому роду Houcaris семейства Tamisiocarididae , [27] но последующий анализ показывает, что H. saron вместо этого является членом семейства Amplectobeluidae и что H ? magnabasis (обнаруженный как родственный таксон Amplectobeluidae) не образует монофилетическую кладу с другими видами Houcaris . [28] В том же году « A. » pennsylvanica была переведена в род Lenisicaris . [3] В 2022 году экземпляр ELRC 20001, который считался безымянным видом Anomalocaris или экземпляром целиком A. saron, получил новый род Innovatiocaris . [29] В 2023 году «А». kunmingensis был отнесен к новому роду Guanshancaris семейства Amplectobeluidae . [30] Множественный филогенетический анализ также показал, что «А». briggsi (tamisiocaridid) также не был видом Anomalocaris , [4] [31] [32] [33] и в 2023 году он был переведен в род Echidnacaris семейства Tamisiocarididae . [2]
Описание
[ редактировать ]
Для того времени, когда он жил, Аномалокарис был гигантским. Полный экземпляр A. canadensis , ROMIP 51211, имеет длину до 20,5 см (8,1 дюйма). [8] [34] (17,4 см (6,9 дюйма) в длину без учета лобных придатков и хвостового веера [4] ). Самый большой лобный придаток в вытянутом состоянии имеет длину до 18 см (7,1 дюйма). [35] и этот экземпляр A. canadensis достигал 34,2–37,8 см (1,12–1,24 фута) в длину тела, исключая лобные придатки и хвостовой веер. [4] [29] Предыдущая оценка длины тела до 1 м (3,3 фута) [16] маловероятно, исходя из соотношения частей тела [35] (длина тела всего лишь примерно в 2 раза превышает длину лобного придатка у A. canadensis [4] ) и размер крупнейшего лобного придатка. [35] A. daleyae (ранее A. cf. canadensis или A. aff. canadensis ) из сланцев залива Эму в Австралии крупнее, чем A. canadensis , с самым большим известным придатком длиной до 18,3 см (7,2 дюйма), что соответствует принадлежал человеку длиной 34,8–51,2 см (1,14–1,68 фута). [2] [4]
Аномалокарис продвигался по воде, взмахивая гибкими крыльями по бокам своего тела. [36] Каждая створка наклонена ниже еще одной, расположенной позади нее. [37] и это перекрытие позволило лепесткам на каждой стороне тела действовать как единый «плавник», максимизируя эффективность плавания. [36] Создание модели с дистанционным управлением показало, что этот режим плавания по своей природе стабилен. [38] подразумевая, что Anomalocaris не нуждался бы в сложном мозге, чтобы управлять балансом во время плавания. Тело было наиболее широким между третьей и пятой долями и сужалось к хвосту, с дополнительными 3 парами небольших створок в суженной области шеи. [8] [35] Около хвоста трудно различить доли, что затрудняет точный подсчет. [37] Для основных створок туловища у типового вида A. canadensis было 13 пар. [35]

 Подвижность туловищных придатков радиодонта (типа Anomalocaris ).
Подвижность туловищных придатков радиодонта (типа Anomalocaris ). головного склерита Структура Anomalocaris .
головного склерита Структура Anomalocaris .



У Anomalocaris был необычный дискообразный рот, известный как ротовой конус. Ротовой конус состоял из нескольких пластинок, расположенных трехрадиально. Три тарелки были довольно большими. Между каждой большой тарелкой можно было найти по три-четыре тарелки среднего размера, а между ними - несколько маленьких. Большая часть пластинок сморщена и имеет чешуйчатые бугорки возле ротового отверстия. [10] [39] Такой ротовой конус сильно отличается от конуса типичных хурдиид, радиодонтов таких как Peytoia и Hurdia , который гладкий и тетрарадиальный. [10] [32] Будучи общим признаком всех радиодонтов, Anomalocaris также имел три склерита на макушке и по бокам головы. [32] Верхний, известный как головной щит, спинной панцирь или Н-элемент, имел форму вытянутого вбок [40] овальной формы, с отчетливым венчиком по внешнему краю. [35] Остальные два боковых склерита, известные как Р-элементы, также были яйцевидными, но соединены перемычным выростом. [32] P-элементы ранее ошибочно интерпретировались как два огромных сложных глаза. [35] [32]

Перед ртом, на передней части головы, располагались два крупных лобных придатка. [8] Каждый лобный придаток Anomalocaris обычно имел 14 подомеров (сегментарные единицы, по крайней мере 1 для стержня и 13 для дистальной сочлененной области), причем каждый придаток был уплощен с боков (выше ширины). [35] Большинство подомеров имели на концах пару эндитов (вентральных шипов). [35] Сами эндиты были снабжены множеством вспомогательных шипов, которые ответвляются от переднего и заднего края эндитов. [25] [39] [41] [35] [1]
Хвост представлял собой большой веер, состоящий из трех [8] [35] пары больших боковых лопастей в форме плавников и одна конечная лопастная хвостовая часть. [35] Предыдущие исследования показали, что хвостовой веер использовался для перемещения корабля по кембрийским водам. [20] [36] в то время как дальнейшие гидродинамические исследования скорее показывают, что он был более приспособлен для обеспечения функции рулевого управления . [34] Жабры животного в виде длинных тонких волосообразных структур, известных как ланцетные лопасти, располагались рядами, образуя щетинковые лопасти. Щетинковые лопасти своим краем прикреплялись к верхней стороне животного, по две щетинок на каждый сегмент тела. Перегородка проходила посередине, разделяя жабры. [35]
На основе окаменелых глаз из сланцев залива Эму , принадлежащих к виду Anomalocaris daleyae. [2] Стебельчатые глаза Anomalocaris были в 30 раз мощнее, чем у трилобитов, которые, как долгое время считалось, имели самые развитые глаза среди всех современных видов. Поскольку у одного экземпляра в одном глазу было более 24 000 линз, разрешение глаз шириной 3 сантиметра (1,2 дюйма) могло бы конкурировать только с разрешением современной стрекозы , у которой в каждом глазу 28 000 линз. [21] Кроме того, оценка экдизозоа опсинов позволяет предположить, что Anomalocaris, возможно, обладали дихроматическим цветовым зрением . [42]
Палеобиология
[ редактировать ]Диета
[ редактировать ]
Интерпретация Anomalocaris как активного хищника широко распространена на протяжении всей истории исследований. [9] [8] [10] поскольку его хищные лобные придатки и железы средней кишки убедительно свидетельствуют о хищническом образе жизни. [43] [44] [5] Что касается A. canadensis , то его выдающиеся размеры среди фауны сланцев Берджесс также делают его одним из первых хищников . известных [5]
Однако давняя идея о том, что Anomalocaris питался животными с твердым телом, особенно его способность проникать в минерализованный экзоскелет трилобитов, была поставлена под сомнение, и многие недавние исследования считают более вероятным, что Anomalocaris охотился исключительно на добычу с мягким телом. [45] [10] [5] [6] У некоторых кембрийских трилобитов были обнаружены круглые или W-образные следы «укусов», которые были идентифицированы как имеющие ту же форму, что и ротовой аппарат Peytoia (ранее ошибочно идентифицированный как таковой у Anomalocaris). [46] [10] ). Более убедительные доказательства того, что Anomalocaris питались трилобитами, получены из копролита , который содержит части трилобитов и настолько велик, что радиодонты являются единственным известным организмом того периода, достаточно большим, чтобы произвести их. [46] Однако, поскольку у Anomalocaris отсутствует какая-либо минерализованная ткань, казалось маловероятным, что он сможет проникнуть в твердый кальцинированный экзоскелет трилобитов. [46] Скорее, копролиты могли быть произведены разными организмами, такими как трилобиты рода Redlichia . [39] Другая предполагаемая возможность заключалась в том, что Anomalocaris питался, хватая один конец добычи своим ротовым конусом, одновременно используя свои лобные придатки, чтобы быстро раскачивать другой конец животного вперед и назад. Это вызвало стресс, который использовал слабости кутикулы членистоногих , вызывая разрыв экзоскелета добычи и позволяя хищнику получить доступ к ее внутренностям. [46] Первоначально считалось, что такое поведение оказывало эволюционное давление на трилобитов, заставляя их сворачиваться, чтобы избежать сгибания до тех пор, пока они не сломаются. [46]

Отсутствие износа ротового аппарата радиодонтов позволяет предположить, что они не вступали в регулярный контакт с минерализованными панцирями трилобитов и, возможно, были лучше приспособлены для питания более мелкими, мягкотелыми организмами путем всасывания, так как у них произошла бы структурная поломка, если бы их использовали против доспехи трилобитов. [45] [39] Было высказано предположение, что A. canadensis был способен питаться организмами с твердым экзоскелетом из-за коротких и прочных шипов на его лобных придатках. [39] [26] Однако этот вывод основан исключительно на сравнении с хрупкими лобными придатками радиодонтов, питающихся взвесью (например, Echidnacaris и Houcaris spp.). [27] Типичное отсутствие повреждений эндитов на лобных придатках A. canadensis (повреждения имеются только у одного экземпляра) позволяет предположить, что они не использовались для захвата добычи с твердым панцирем. [6] В отличие от пейтои, ротовой конус которой более прямоугольный с короткими выступающими шипами, ротовой конус A. canadensis имеет меньшее и более неправильной формы отверстие, не допускающее сильных кусающих движений и указывающее на всасывание-питание для всасывания более мягких организмов. [10] Трехмерное моделирование различных лобных придатков радиодонтов также позволяет предположить, что A. canadensis более способна охотиться на более мелких (2–5 см в диаметре), активных животных с мягким телом (например, ветуколии ; свободно плавающие членистоногие, такие как изоксииды и гименокарины ; Нектокарис ). [5] [6]
Бикнелл и др. (2023) исследовали лобные придатки Anomalocaris , предполагая, что это был активный нектонный вершинный хищник. С вытянутыми лобными конечностями аномалокарис мог плавать с максимальной скоростью, подобно современным хищным водяным клопам . Его глаза подходят для охоты на добычу в хорошо освещенных водах. Аномалокарис охотился на различных свободноплавающих животных, поскольку существует большое разнообразие нектонных и пелагических мягкотелых животных. Вероятно, он не стал бы охотиться на донных животных, таких как трилобиты, учитывая возможность повреждения лобных придатков о субстрате при попытке схватить добычу со дна моря на скорости. Вместо этого другие животные, такие как другие радиодонты (например, Hurdia , Cambroraster , Titanokorys , Stanleycaris ) и парнокопытные (например, Sidneyia ), могли быть донными хищниками в сланцах Бёрджесс. [5] [6]
Палеоэкология
[ редактировать ]Особи Anomalocaris были обнаружены по всему миру, начиная с кембрийского этапа 3 и заканчивая гучжанским периодом . Помимо сланцев Бёрджесс и сланцев залива Эму, окаменелости были обнаружены в биоте Чэнцзян , формации Хунцзиншао , формации Баланг и формации Кайли в Китае, а также в формации Игар и формации Уикс в Соединенных Штатах. [3]
Anomalocaris canadensis обитала в сланцах Бёрджесс в относительно большом количестве. [1] В сланцах Берджесс Anomalocaris чаще встречается в более старых участках, особенно в пластах трилобитов горы Стивен . Однако в более молодых разрезах, таких как слой Phyllopod , Anomalocaris мог достигать гораздо больших размеров; примерно в два раза больше своих старших родственников-трилобитов. Эти редкие гигантские экземпляры ранее относили к отдельному виду Anomalocaris gigantea ; однако достоверность этого вида была поставлена под сомнение, [16] и в настоящее время является синонимом A. canadensis . [35]
Другие безымянные виды Anomalocaris живут в совершенно других условиях. [3] Например, Anomalocaris cf. canadensis (JS-1880) обитал в сланцах Маотяньшань , [3] мелкое тропическое море или даже дельта [47] на территории современного Китая. Anomalocaris daleyae (Emu Bay Shale) обитал в сопоставимых условиях; мелкие тропические воды кембрийской Австралии. [3] Сланец Маотяньшань и сланец залива Эму расположены очень близко друг к другу, их разделяет небольшой участок суши, вдали от сланцев Бёрджесс. [3] Эти два местонахождения также включали соответственно «Anomalocaris» kunmingensis и «Anomalocaris» briggsi , виды, которые ранее приписывались [48] [49] [39] [50] но таксономически маловероятно, что он принадлежит ни к Anomalocaris , ни даже к Anomalocarididae. [3] [51]
См. также
[ редактировать ]- 8564 Аномалокарис — астероид, названный в честь этого животного.
- Radiodonta — вымерший отряд членистоногих, состоящий из Anomalocaris и его родственников.
- Houcaris , Lenisicaris , Innovatiocaris , Guanshancaris , Echidnacaris , роды радиодонтов, содержащие виды, первоначально называвшиеся Anomalocaris .
- Эгирокассис , гигантский радиодонт-фильтратор из ордовикского Марокко .
- Кембрийский взрыв , крупное событие биоразнообразия, произошедшее в кембрийском периоде.
- Opabinia — род причудливых членистоногих стеблевой группы, отдаленно связанных с радиодонтами.
- Wiwaxia , род возможных моллюсков, имевший обильное количество углеродистых чешуек и живший рядом с Anomalocaris .
- Палеобиота сланцев Бёрджесс
Сноски
[ редактировать ]- ^ Перейти обратно: а б с Лерози-Обрил Р., Хегна Т.А., Бэбкок Л.Е., Бонино Э., Кир С. (19 мая 2014 г.). «Придатки членистоногих из формации Уикс Консерват-Лагерштетте: новые находки аномалокаридид в кембрии штата Юта, США» . Бюллетень геонаук : 269–282. дои : 10.3140/bull.geosci.1442 .
- ^ Перейти обратно: а б с д и ж Патерсон, Джон Р.; Гарсиа-Беллидоб, Диего К.; Эджкомб, Грегори Д. (10 июля 2023 г.). «Возвращение к раннекембрийским радиодонтам сланцев Эму Бэй: морфология и систематика» . Журнал систематической палеонтологии . 21 (1). Бибкод : 2023JSPal..2125066P . дои : 10.1080/14772019.2023.2225066 . S2CID 259719252 .
- ^ Перейти обратно: а б с д и ж г час я дж Ву Ю, Ма Дж, Линь В, Сунь А, Чжан Икс, Фу Д (2021). «Новые аномалокаридиды (Panarthropoda: Radiodonta) из нижнего кембрия Chengjiang Lagerstätte: биостратиграфические и палеобиогеографические значения». Палеогеография, Палеоклиматология, Палеоэкология . 569 : Артикул 110333. Бибкод : 2021PPP...56910333W . дои : 10.1016/j.palaeo.2021.110333 . S2CID 233565727 .
- ^ Перейти обратно: а б с д и ж Лерози-Обрил Р., Пейтс С. (сентябрь 2018 г.). «Новый радиодонт, питающийся взвесью, предполагает эволюцию микропланктивных животных в кембрийском макронектоне» . Природные коммуникации . 9 (1): 3774. Бибкод : 2018NatCo...9.3774L . дои : 10.1038/s41467-018-06229-7 . ПМК 6138677 . ПМИД 30218075 . Данные дриад
- ^ Перейти обратно: а б с д и ж Де Виво Дж., Лаутеншлагер С., Винтер Дж. (июль 2021 г.). «Трехмерное моделирование, неравенство и экология первых кембрийских хищников вершины» . Слушания. Биологические науки . 288 (1955): 20211176. doi : 10.1098/rspb.2021.1176 . ПМЦ 8292756 . ПМИД 34284622 .
- ^ Перейти обратно: а б с д и Бикнелл Р.Д., Шмидт М., Рахман И.А., Эджкомб Г.Д., Гутарра С., Дейли А.С. и др. (12 июля 2023 г.). «Хищные придатки кембрийского хищника Anomalocaris canadensis созданы для мягкой добычи и скорости» . Труды Королевского общества B: Биологические науки . 290 (2002). дои : 10.1098/rspb.2023.0638 . ISSN 0962-8452 . ПМЦ 10320336 . ПМИД 37403497 .
- ^ Перейти обратно: а б с Уайтэйвс Дж. Ф. (1892). «Описание нового рода и вида филлокарид ракообразных из среднего кембрия горы Стивен, Британская Колумбия» Canadian Record of Science . 5 (4): 205–208.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Коллинз Д. (1996). «Эволюция» Anomalocaris и его классификация в классе членистоногих Dinocarida (нов.) и отряде Radiodonta (нов.)». Журнал палеонтологии . 70 (2): 280–293. Бибкод : 1996JPal...70..280C . дои : 10.1017/S0022336000023362 . JSTOR 1306391 . S2CID 131622496 .
- ^ Перейти обратно: а б с д Уиттингтон Х.Б., Бриггс Д.Э. (1985). «Самое крупное кембрийское животное, Anomalocaris , Берджесс-Шейл, Британская Колумбия» . Философские труды Королевского общества Б. 309 (1141): 569–609. Бибкод : 1985RSPTB.309..569W . дои : 10.1098/rstb.1985.0096 .
- ^ Перейти обратно: а б с д и ж г час Дэйли AC, Бергстрем Дж (июнь 2012 г.). «Ротовая шишка Anomalocaris — это не классическая пейтоя » . Die Naturwissenschaften . 99 (6): 501–4. Бибкод : 2012NW.....99..501D . дои : 10.1007/s00114-012-0910-8 . ПМИД 22476406 . S2CID 2042726 .
- ^ Гулд С.Дж. (1989). Прекрасная жизнь: сланцы Бёрджесс и природа истории . Нью-Йорк: WW Нортон. п. 194. ИСБН 0-393-02705-8 . OCLC 18983518 .
- ^ «Сланец Бёрджесс: первые открытия» . burgess-shale.rom.on.ca . Королевский музей Онтарио . Проверено 25 марта 2023 г.
- ^ Моррис С.К. (1994-09-01), Мейсон Р. (ред.), «Как сланцы Берджесс пришли в Кембридж; и что произошло» , Cambridge Minds (1-е изд.), Cambridge University Press, стр. 126–141, doi : 10.1017/cbo9780511523007.011 , ISBN 978-0-521-45405-6 , получено 25 марта 2023 г.
- ^ Ферье В.Ф. (1893). Каталог стратиграфической коллекции канадских пород, подготовленной для Всемирной Колумбийской выставки, Чикаго, 1893 г. Оттава (Канада): Геологическая служба Канады/Государственное типографское бюро. п. 87. ИСБН 9781014253446 .
- ^ Вудворд Х (1902). «I. — Канадские Скалистые горы. Часть I: О коллекции окаменелостей среднего кембрия, полученной Эдвардом Уимпером, эсквайром, FRGS, с горы Стивен, Британская Колумбия» . Геологический журнал . 9 (12): 529–544. Бибкод : 1902GeoM....9..529W . дои : 10.1017/S001675680018149X . ISSN 0016-7568 . S2CID 131669387 .
- ^ Перейти обратно: а б с д Бриггс Д.Э. (1979). « Аномалокарис , крупнейшее известное кембрийское членистоногое» (PDF) . Палеонтология . 22 (3): 631–664. S2CID 134499952 .
- ^ Уайтэйвс Дж. Ф. (1892). «Описание нового рода и вида филлокарид ракообразных из среднего кембрия горы Стивен, Британская Колумбия». Канадский рекорд науки . 5 (4).
- ^ Перейти обратно: а б с Гулд С.Дж. (1989). Прекрасная жизнь: сланцы Бёрджесс и природа истории . Нью-Йорк: WW Нортон. стр. 194–206 . ISBN 978-0-393-02705-1 .
- ^ Конвей Моррис С. (1978). « Лаггания камбрия Уолкотт: сложное ископаемое». Журнал палеонтологии . 52 (1): 126–131. JSTOR 1303799 .
- ^ Перейти обратно: а б Конвей Моррис С. (1998). Тигель творения: сланцы Бёрджесс и появление животных . Оксфорд [Оксфордшир]: Издательство Оксфордского университета. стр. 56–9. ISBN 978-0-19-850256-2 .
- ^ Перейти обратно: а б Патерсон-младший, Гарсиа-Беллидо, округ Колумбия, Ли М.С., Брок Г.А., Джаго Дж.Б., Эджкомб, Г.Д. (декабрь 2011 г.). «Острое зрение у гигантского кембрийского хищника Anomalocaris и происхождение сложных глаз» . Природа . 480 (7376): 237–40. Бибкод : 2011Natur.480..237P . дои : 10.1038/nature10689 . ПМИД 22158247 . S2CID 2568029 .
- ^ Винтер Дж., Стейн М., Лонгрич Н.Р., Харпер Д.А. (март 2014 г.). «Аномалокарид, питающийся суспензией, из раннего кембрия» (PDF) . Природа . 507 (7493): 496–9. Бибкод : 2014Natur.507..496V . дои : 10.1038/nature13010 . ПМИД 24670770 . S2CID 205237459 .
- ^ Конг П., Ма Х., Хоу Х., Эджкомб Г.Д., Штраусфельд, штат Нью-Джерси (сентябрь 2014 г.). «Структура мозга устраняет сегментарное сродство аномалокаридидных придатков». Природа . 513 (7519): 538–42. Бибкод : 2014Natur.513..538C . дои : 10.1038/nature13486 . ПМИД 25043032 . S2CID 4451239 .
- ^ Ван Рой П., Дейли AC, Бриггс Д.Э. (июнь 2015 г.). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками». Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ПМИД 25762145 . S2CID 205242881 .
- ^ Перейти обратно: а б Сянь-Гуан Х., Бергстрем Дж., Альберг П. (1 сентября 1995 г.). «Anomalocaris и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая». ГФФ . 117 (3): 163–183. Бибкод : 1995GFF...117..163X . дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Перейти обратно: а б Пейтс С., Дейли А.С., Эджкомб Г.Д., Конг П., Либерман Б.С. (2019). «Систематика, сохранение и биогеография радиодонтов из южной части Большого Бассейна, США, во время верхнего Дайрана (кембрийская серия 2, этап 4)» . Статьи по палеонтологии . 7 : 235–262. дои : 10.1002/spp2.1277 . ISSN 2056-2799 . S2CID 204260554 .
- ^ Перейти обратно: а б Ву Ю, Фу Д, Ма Дж, Линь В, Сунь А, Чжан Икс (2021). « Houcaris gen. nov. из раннего кембрия (этап 3) Chengjiang Lagerstätte расширил палеогеографическое распространение тамизиокаридид (Panarthropoda: Radiodonta)». ПалЗ . 95 (2): 209–221. Бибкод : 2021PalZ...95..209W . дои : 10.1007/s12542-020-00545-4 . ISSN 1867-6812 . S2CID 235221043 .
- ^ МакКолл, Кристиан (13 декабря 2023 г.). «Большой пелагический лобопод из кембрийских сланцев Пиоче в Неваде» . Журнал палеонтологии . 97 (5): 1009–1024. Бибкод : 2023JPal...97.1009M . дои : 10.1017/jpa.2023.63 .
- ^ Перейти обратно: а б Цзэн Х, Чжао Ф, Чжу М (07 сентября 2022 г.). « Innovatiocaris , полный радиодонт из раннего кембрия Chengjiang Lagerstätte и его значение для филогении Radiodonta». Журнал Геологического общества . 180 . дои : 10.1144/jgs2021-164 . ISSN 0016-7649 . S2CID 252147346 .
- ^ Чжан М, Ву Ю, Линь В, Ма Дж, Ву Ю, Фу Д (апрель 2023 г.). «Amplectobeluid Radiodont Guanshancaris gen. nov. из нижнего кембрия (этап 4) Гуаньшань Лагерштетте в Южном Китае: биостратиграфические и палеобиогеографические последствия» . Биология . 12 (4): 583. doi : 10.3390/biology12040583 . ПМЦ 10136193 . ПМИД 37106783 .
- ^ Лю Дж., Лерози-Обрил Р., Штайнер М., Данлоп Дж.А., Шу Д., Патерсон-младший (01.11.2018). «Происхождение хищного питания у молодых членистоногих, выявленное кембрийским радиодонтаном» . Национальный научный обзор . 5 (6): 863–869. дои : 10.1093/nsr/nwy057 . ISSN 2095-5138 .
- ^ Перейти обратно: а б с д и Мойсюк Дж., Кэрон Дж.Б. (август 2019 г.). «Новый радиодонт-хурдиид из сланцев Бёрджесс свидетельствует об эксплуатации кембрийских инфаунистических источников пищи» . Слушания. Биологические науки . 286 (1908): 20191079. doi : 10.1098/rspb.2019.1079 . ПМК 6710600 . ПМИД 31362637 .
- ^ Мойсюк Дж., Кэрон Дж.Б. (2021). «Исключительная многофункциональность в питающем аппарате среднекембрийского радиодонта» . Палеобиология . 47 (4): 704–724. Бибкод : 2021Pbio...47..704M . дои : 10.1017/pab.2021.19 . ISSN 0094-8373 . S2CID 236552819 .
- ^ Перейти обратно: а б Шеппард К.А., Ривал Д.Э., Кэрон Дж.Б. (октябрь 2018 г.). «К гидродинамике хвостового плавника Anomalocaris». Интегративная и сравнительная биология . 58 (4): 703–711. дои : 10.1093/icb/icy014 . HDL : 1974/22737 . ПМИД 29697774 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н Дейли AC, Эджкомб, Джорджия (январь 2014 г.). «Морфология Anomalocaris canadensis из сланцев Бёрджесс» . Журнал палеонтологии . 88 (1): 68–91. Бибкод : 2014JPal...88...68D . дои : 10.1666/13-067 . S2CID 86683798 .
- ^ Перейти обратно: а б с Усами Ю. (январь 2006 г.). «Теоретическое исследование формы тела и схемы плавания Anomalocaris на основе гидродинамического моделирования». Журнал теоретической биологии . 238 (1): 11–7. Бибкод : 2006JThBi.238...11U . дои : 10.1016/j.jtbi.2005.05.008 . ПМИД 16002096 .
- ^ Перейти обратно: а б Уиттингтон Х.Б., Бриггс Д.Э. (1985). «Самое крупное кембрийское животное, Anomalocaris , сланцы Бёрджесс, Британская Колумбия» . Философские труды Королевского общества B (полный текст бесплатно). 309 (1141): 569–609. Бибкод : 1985RSPTB.309..569W . дои : 10.1098/rstb.1985.0096 .
- ^ Бриггс Д.Э. (май 1994 г.). «Гигантские хищники кембрия Китая». Наука . 264 (5163): 1283–4. Бибкод : 1994Sci...264.1283B . дои : 10.1126/science.264.5163.1283 . ПМИД 17780843 .
- ^ Перейти обратно: а б с д и ж Дейли AC, Патерсон-младший, Эджкомб GD, Гарсиа-Беллидо DC, Джаго Дж.Б. (2013). Донохью П. (ред.). «Новая анатомическая информация об Anomalocaris из кембрийских сланцев залива Эму в Южной Австралии и переоценка его предполагаемых хищнических привычек» . Палеонтология . 56 (5): 971–990. Бибкод : 2013Palgy..56..971D . дои : 10.1111/пала.12029 .
- ^ Цзэн Х, Чжао Ф, Инь Цз, Чжу М (2 января 2018 г.). «Морфология разнообразных склеритов головы радиодонтана из раннего кембрия Чэнцзян Лагерштетте, юго-запад Китая» . Журнал систематической палеонтологии . 16 (1): 1–37. Бибкод : 2018JSPal..16....1Z . дои : 10.1080/14772019.2016.1263685 . ISSN 1477-2019 . S2CID 133549817 .
- ^ Лю Цюй (01 сентября 2013 г.). «Первое открытие придатков аномалокаридид из формации Баланг (кембрийская серия 2) в Хунани, Китай». Алчеринга: Австралазийский журнал палеонтологии . 37 (3): 338–343. Бибкод : 2013Алч...37..338Л . дои : 10.1080/03115518.2013.753767 . ISSN 0311-5518 . S2CID 129212098 .
- ^ Флеминг Дж. Ф., Кристенсен Р. М., Соренсен М. В., Парк Т. С., Аракава К., Блакстер М. и др. (декабрь 2018 г.). «Молекулярная палеонтология освещает эволюцию зрения экдизозоев» . Слушания. Биологические науки . 285 (1892): 20182180. doi : 10.1098/rspb.2018.2180 . ПМК 6283943 . ПМИД 30518575 .
- ^ Ваннье Дж., Лю Дж., Лерози-Обрил Р., Винтер Дж., Дэйли AC (май 2014 г.). «Сложная пищеварительная система у ранних членистоногих» . Природные коммуникации . 5 (1): 3641. Бибкод : 2014NatCo...5.3641V . дои : 10.1038/ncomms4641 . ПМИД 24785191 .
- ^ Де Виво Дж., Лаутеншлагер С., Винтер Дж. (16 декабря 2016 г.). Реконструкция ловкости пищевых придатков аномалокаридид проливает свет на экологию радиодонтана (Отчет).
- ^ Перейти обратно: а б Хагадорн JW (август 2009 г.). «Откусив от Аномалокариса » (PDF) . Смит М.Р., О'Брайен Л.Дж., Кэрон Дж.Б. (ред.). Аннотация Том . Международная конференция по кембрийскому взрыву (Уолкотт, 2009 г.) . Торонто, Онтарио, Канада: Консорциум Burgess Shale (опубликовано 31 июля 2009 г.). ISBN 978-0-9812885-1-2 .
- ^ Перейти обратно: а б с д и Недин С (1999). « Хищничество Anomalocaris на неминерализованных и минерализованных трилобитах». Геология . 27 (11): 987–990. Бибкод : 1999Geo....27..987N . doi : 10.1130/0091-7613(1999)027<0987:APONAM>2.3.CO;2 .
- ^ Салех Ф., Ци С., Буатойс Л.А., Мангано М.Г., Пас М., Воше Р. и др. (март 2022 г.). «Биота Чэнцзян обитала в дельтовой среде» . Природные коммуникации . 13 (1): 1569. Бибкод : 2022NatCo..13.1569S . дои : 10.1038/s41467-022-29246-z . ПМЦ 8943010 . ПМИД 35322027 .
- ^ Недин, Кристофер (1995). Сланец залива Эму, окаменелость нижнего кембрия Лагерштатт, остров Кенгуру, Южная Австралия .
- ^ Ван Ю, Хуан Д, Ху С (01 ноября 2013 г.). «Новые лобные придатки аномалокардид из биоты Гуаньшань, восточная часть Юньнани» . Китайский научный бюллетень . 58 (32): 3937–3942. Бибкод : 2013ЧСБу..58.3937В . дои : 10.1007/s11434-013-5908-x . ISSN 1861-9541 .
- ^ Джинс Дж. «Картирование мировых месторождений типа сланцев Бёрджесс» . www.virtualmuseum.ca . Проверено 16 сентября 2019 г.
- ^ Цзяо Д.Г., Пейтс С., Лерози-Обрил Р., Ортега-Эрнандес Дж., Ян Дж., Лан Т., Чжан С.Г. (2021). «Эндемичные радиодонты кембрийской биоты Гуаньшаня 4-го этапа Южного Китая» . Acta Palaeontologica Polonica . 66 . дои : 10.4202/app.00870.2020 . ISSN 0567-7920 .
Внешние ссылки
[ редактировать ]- « Аномалокарис канадский » . Галерея окаменелостей сланцев Бёрджесс . Виртуальный музей Канады. 2011. Архивировано из оригинала 12 ноября 2020 г. Проверено 21 января 2023 г.
- Anomalocaris с анимацией плавания Домашняя страница
- Берджесс-Шейл: Anomalocaris canadensis (проточленистоногие) , Смитсоновский институт.

- Кембрийские членистоногие
- Окаменелости сланцев Бёрджесс
- Окаменелости сланцев Маотяньшань
- Радиодонта
- Ископаемые таксоны, описанные в 1892 г.
- Таксоны, названные Джозефом Фредериком Уайтавсом
- Кембрийское вымирание родов
- Уилер Шейл
- Emu Bay Shale
- Палеозойская жизнь Северо-Западных территорий.
- Высшие хищники
- Окаменелости Канады