Радиодонта
| Радиодонта Временной диапазон:
| |
|---|---|

| |
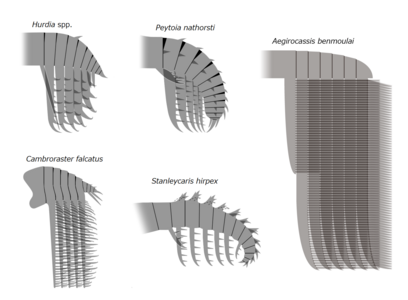
| Слева направо, сверху вниз: Amplectobelua symbrachiata , Anomalocaris canadensis , Aegirocassis benmoulai , Peytoia nathorsti , Lyrarapax unguispinus , Cambroraster falcatus и Hurdia victoria. | |
| Научная классификация | |
| Королевство: | |
| Тип: | |
| Сорт: | |
| Заказ: | † Радиодонта
Коллинз , 1996 год. |
| Семьи | |
| |
Radiodonta — вымерший отряд членистоногих стволовой группы , который пользовался успехом во всем мире в кембрийский период. Их можно назвать радиодонтами . [ 1 ] [ 2 ] [ 3 ] радиодонтаны , [ 4 ] [ 5 ] радиодонтиды , [ 6 ] аномалокариды , [ 7 ] или аномалокаридиды , [ 8 ] [ 9 ] [ 10 ] хотя два последних первоначально относятся к семейству Anomalocarididae , которое ранее включало все виды этого отряда, но теперь ограничивается лишь несколькими видами. [ 7 ] Радиодонты отличаются характерными лобными придатками, морфологически разнообразными и выполняющими самые разные функции. Радиодонты включали в себя самых ранних известных крупных хищников, но среди них также были просеиватели отложений и фильтраторы. [ 11 ] Некоторые из наиболее известных видов радиодонтов — кембрийские таксоны Anomalocaris canadensis , Hurdia victoria , Peytoia nathorsti , Titanokorys Ganessii , Cambroraster falcatus и Amplectobelua symbrachiata . выжившим представителям относятся подсемейство Aegirocassisinae из раннего ордовика Марокко К последним и раннего девона представитель Schinderhannes bartelsi из Германии.
Этимология
[ редактировать ]Название Radiodonta (от латинского radius «спица колеса» и от греческого odoús «зуб») относится к радиальному расположению зубных пластинок (ротового конуса), окружающих рот. [ 6 ] хотя предполагается, что эти особенности отсутствуют у некоторых видов радиодонтов. [ 4 ] [ 1 ]
Определение
[ редактировать ]Первоначальный диагноз отряда Radiodonta в 1996 г. следующий: [ 6 ]
Радиодонтиды — двусторонне-симметричные, удлиненные членистоногие с неминерализованной кутикулой, обычно наиболее прочной в челюстях и когтях. Тело подразделяется на две тагматы , во многом напоминающие и опистосому хелицеровых членистоногих просому . Обычно передняя часть не имеет внешней сегментации, несет одну пару предоральных когтей, пару выдающихся глаз и вентральные челюсти с радиально расходящимися зубами. Некоторые формы имеют дополнительные ряды зубов и три-четыре посторальные пары гнатобазальных конечностей. Туловище метамерное , обычно имеет около 13 сегментов, по бокам развиваются черепитчатые лопасти для плавания и жабры для дыхания, и может заканчиваться выступающим трехчастным хвостом. Некоторые формы имеют гнатобазные туловищные конечности.
В 2014 году клада Radiodonta была определена филогенетически как клада, включающая любые таксоны, более близкие к Anomalocaris canadensis, чем к Paralithodes camtschaticus . [ 7 ] В 2019 году он был переопределен морфологически как панцирный комплекс головы животного с центральными (H-) и боковыми (P-) элементами; выросты (эндиты) лобных придатков, несущие вспомогательные шипы; уменьшенные передние створки или полосы пластинок (щеточные лопасти) и сильное сужение тела спереди назад. [ 3 ]
Описание
[ редактировать ]
Большинство радиодонтов были значительно крупнее представителей другой кембрийской фауны, при этом типичная длина тела крупных таксонов варьировалась от 30 до 50 см (от 12 до 20 дюймов). [ 12 ] Самый крупный из описанных радиодонтов — раннеордовикский вид Aegirocassis benmoulai , который мог достигать 2 м (6,6 футов) в длину. [ 10 ] [ 2 ] Почти полный экземпляр молодого Lyrarapax unguispinus имел размеры всего 18 мм (0,71 дюйма), что делает его одним из самых маленьких известных экземпляров радиодонтов, хотя взрослые особи достигали длины 8,3 см (3,3 дюйма). [ 2 ] [ 13 ] Известен изолированный лобный придаток хурдииды длиной менее половины длины молоди Lyrarapax , однако неизвестно, принадлежит ли этот экземпляр взрослой особи. [ 14 ] Самым крупным известным кембрийским радиодонтом был Amplectobelua , достигавший длины до 90 см (35 дюймов), судя по неполному экземпляру. [ 15 ] Anomalocaris canadensis также был относительно крупным, его длина оценивалась до 34,2–37,8 см (13,5–14,9 дюйма). [ 2 ] а кембрийская хурдия Titanokorys достигала длины около 50 см (20 дюймов). [ 16 ]
Тело радиодонта можно разделить на две части: голову и туловище. Голова состоит только из одного сегмента тела. [ 17 ] известный как глазной сомит, покрытый склеритами (комплекс головного панциря), нес членистоногие лобные придатки, вентральный ротовой аппарат (ротовой конус) и на ножках сложные глаза . Сужающийся туловище состоит из множества сегментов тела, каждый из которых связан с парой створок и жаберными структурами (щетиночными лопастями). [ 3 ]
Лобный придаток
[ редактировать ]

Передние структуры головы представляют собой пару лобных придатков, которые в предыдущих исследованиях назывались «когтями», «хватательными придатками», «питающими придатками» или «большими придатками» (последний термин не рекомендуется, поскольку существует гомология между лобные придатки и исходные, морфологически отличные от мегахейрана больших придатков вызывают сомнения. [ 17 ] [ 18 ] ). Они склеротизированы (затвердевшие) и членистоногие (сегментированы), несут вентральные эндиты (шипы) на большинстве подомеров (сегментарных единиц), а эндиты могут нести дополнительные ряды вспомогательных шипов на переднем и заднем краях. [ 19 ] [ 3 ] Лобный придаток состоит из двух отделов: стержня («ножка», [ 2 ] 'база' [ 20 ] или «промиксальная область» [ 2 ] в некоторых исследованиях) и дистальная суставная область [ 19 ] (также называемый «коготь» [ 20 ] ). Треугольная область, покрытая мягкой кутикулой (артродиальной мембраной), может располагаться на вентральной стороне между подомерами и обеспечивать гибкость. [ 21 ] [ 11 ] Их предполагаемое преокулярное и протоцеребральное происхождение предполагает, что они гомологичны первичным усикам ( все они возникли Onychophora и верхней губе Euarthropoda из глазного сомита ). [ 17 ] [ 9 ] в то время как последующие исследования также предполагают дейтоцеребральное происхождение и гомологичны хелицерам Chelicerata и » других членистоногих (все они возникли усикам или « большим придаткам из постглазного сомита 1). [ 22 ] Поскольку морфология лобных придатков, особенно шипов, всегда различается у разных видов, она является одним из важнейших способов видовой идентификации. [ 19 ] Фактически, многие радиодонты известны только по горстке окаменевших лобных придатков. [ 21 ] [ 19 ]
-
 Лобные придатки Anomalocarididae , Amplectobeluidae и, возможно, родственных видов.
Лобные придатки Anomalocarididae , Amplectobeluidae и, возможно, родственных видов. -
 Лобные придатки Tamisiocarididae
Лобные придатки Tamisiocarididae -
 Лобные придатки Hurdiidae
Лобные придатки Hurdiidae



Оральный конус
[ редактировать ]
Рот находится на вентральной стороне головы, позади точки прикрепления лобных придатков, и окружен кольцом зубных пластинок, образуя ротовой аппарат, известный как ротовой конус («челюсти» в предыдущих исследованиях). [ 6 ] ). 3 или 4 зубные пластинки могут быть увеличены, придавая ротовому конусу трехлучевой (например, Anomalocaris , Echidnacaris ) или тетрарадиальный (например, Hurdiidae , Lyrarapax ) вид. [ 23 ] [ 13 ] [ 24 ] Внутренний край зубных пластинок имеет шипы, обращенные в сторону ротового отверстия. Дополнительные ряды внутренних зубных пластинок могут встречаться у некоторых родов хурдиид . [ 8 ] [ 3 ] Детальная реконструкция некоторых амплектобелюидных оральных конусов является спекулятивной, но, возможно, они не имели типичного радиального расположения. [ 4 ] [ 1 ]
Склериты головы, глаз и туловища
[ редактировать ]
Три головных склерита ( панциря ) комплекса, образованные центральным Н-элементом (передним склеритом или головным щитом) и парой Р-элементов (латеральными склеритами), покрывают дорсальную и латеро-вентальную поверхность головы животного. [ 3 ] P-элементы могут соединяться друг с другом, а также с H-элементом узким передним продолжением (шеей или «клювом» P-элемента). [ 8 ] [ 3 ] Головные склериты мелкие и яйцевидные у Anomalocarididae и Amplectobeluidae . [ 4 ] [ 3 ] но часто увеличены у Hurdiidae , что соответствует их отличительной форме тела (обтекаемое у Anomalocarididae/Amplectobeluidae, но часто компактное у Hurdiidae). [ 3 ] На голове были два на ножках сложных глаза , которые, возможно, имели подвижность. [ 25 ] и располагаются между щелями, образованными задними областями Н-элемента и Р-элементов. [ 8 ] [ 3 ] Сложные глаза Echidnacaris исключительно без стеблей. [ 14 ] Некоторые виды обладают дополнительным срединным глазом позади H-элемента. [ 22 ]


В отличие от первоначального диагноза разделение сегментов тела (границы сегментов) может быть видно внешне. [ 10 ] [ 5 ] [ 3 ] и ни один известный представитель Radiodonta (за исключением предполагаемого радиодонта Cucumericrus [ 10 ] [ 26 ] ), как известно, имеет ножкообразные придатки туловища (ноги). [ 27 ] Туловище имеет многочисленные сегменты тела ( сомиты ), сужающиеся от передней части к задней, при этом передние три или четыре сегмента значительно сужены в область шеи. [ 3 ]
-
 Варианты лоскутов тела радиодонта
Варианты лоскутов тела радиодонта -
 Движение хлопающих придатков радиодонта.
Движение хлопающих придатков радиодонта. -
 Вентральный вид генерализованного радиодонта с GLS, показывающий гнатобазоподобные структуры (GLS), связанные с уменьшенными передними лоскутами.
Вентральный вид генерализованного радиодонта с GLS, показывающий гнатобазоподобные структуры (GLS), связанные с уменьшенными передними лоскутами.



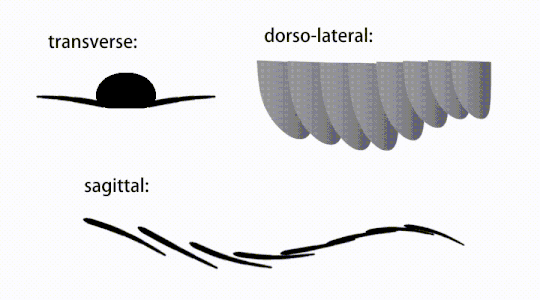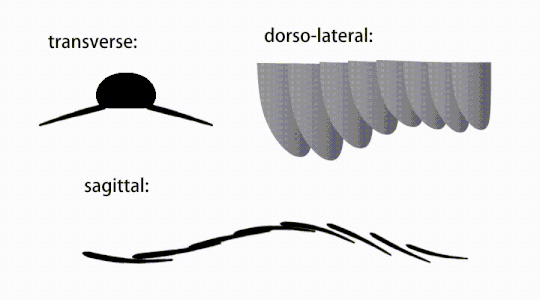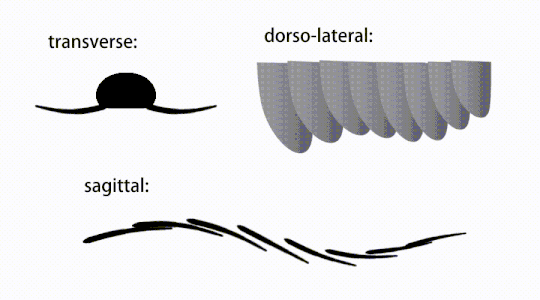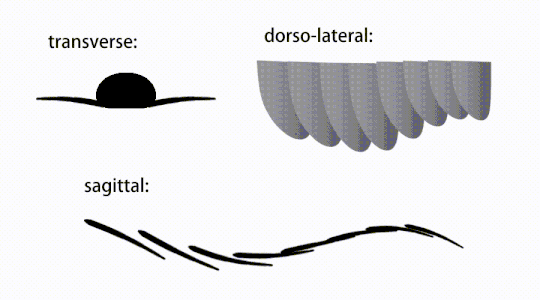
Придатки туловища представляли собой плавниковые створки тела («боковые створки» или «лопасти» в некоторых исследованиях), обычно одна пара брюшных створков на каждый сегмент тела, каждая из которых слегка перекрывала одну, расположенную впереди него, но дополнительные, неперекрывающиеся наборы. небольшие спинные лоскуты могут встречаться у некоторых видов хурдий . [ 10 ] Лоскуты могут иметь многочисленные веноподобные структуры (называемые «укрепляющими лучами»). [ 5 ] 'хлопковые лучи', [ 3 ] «поперечные стержни», [ 10 ] «поперечные линии» [ 28 ] или «вены» [ 29 ] ). Лоскуты в области шеи (называемые «уменьшенными лоскутами») [ 4 ] «шейные лоскуты», [ 5 ] 'хлопки на голове', [ 27 ] «передние лоскуты» [ 30 ] или «дифференцированные закрылки» [ 18 ] ) значительно сокращаются. У некоторых видов челюстеобразные пищевые придатки, называемые гнатобазоподобными структурами (GLS), возникают из каждого основания уменьшенных шейных клапанов. [ 4 ] [ 1 ] Многочисленные удлиненные лопастные расширения (называемые ланцетными лопастями или пластинками). [ 3 ] ), расположенные в ряд, образующие полосы жаберных структур, известных как щетинковые лопасти, покрывали дорсальную поверхность каждого сегмента тела. [ 10 ] По крайней мере, у Aegirocassis каждая ланцетная пластинка покрыта морщинками. [ 10 ] Вентральные створки могут быть гомологичны эндоподам двуветвистых конечностей эуартропод и лобоподам (лобоподам) жаберных лобоподий , а дорсальные створки и щетинковые пластинки могут быть гомологичны выходным и жаберным спинным створкам первых таксонов. [ 31 ] [ 10 ] Ствол может заканчиваться хвостовым опахалом, состоящим из 1-3 пар лопастей. [ 29 ] [ 27 ] [ 3 ] пара длинных фурк, [ 29 ] [ 13 ] [ 3 ] удлиненная терминальная структура, [ 27 ] или безликий тупой кончик. [ 10 ]
-
 Stanleycaris , Hurdia , Aegirocassis , Peytoia и Cambroraster — все это примеры радиодонтов- хурдиид . Это были самые разнообразные и долговечные из семейств радиодонтов, сохранившиеся от кембрия до девона.
Stanleycaris , Hurdia , Aegirocassis , Peytoia и Cambroraster — все это примеры радиодонтов- хурдиид . Это были самые разнообразные и долговечные из семейств радиодонтов, сохранившиеся от кембрия до девона. -
 Anomalocaris — член семейства anomalocarididae , которое когда-то включало всех радиодонтов, но теперь включает лишь несколько родов, таких как Lenisicaris .
Anomalocaris — член семейства anomalocarididae , которое когда-то включало всех радиодонтов, но теперь включает лишь несколько родов, таких как Lenisicaris . -
 Amplectobelua и Lyrarapax являются представителями amplectobeluidae , очень обширного семейства, состоящего преимущественно из китайских радиодонтов.
Amplectobelua и Lyrarapax являются представителями amplectobeluidae , очень обширного семейства, состоящего преимущественно из китайских радиодонтов. -
 Echidnacaris и Tamisiocaris являются примерами семейства Tamisiocarididae , которое питалось исключительно суспензией радиодонтов из кембрия.
Echidnacaris и Tamisiocaris являются примерами семейства Tamisiocarididae , которое питалось исключительно суспензией радиодонтов из кембрия.




Внутренние структуры
[ редактировать ]

Следы мышц , пищеварительной и нервной системы были описаны на некоторых окаменелостях радиодонтов. Пары хорошо развитых мышц соединялись с брюшными створками, расположенными в боковых полостях каждого сегмента тела. [ 27 ] [ 9 ] Между боковыми мышцами находится сложная пищеварительная система, образованная расширением передней и задней кишки, соединенных узкой средней кишкой, связанной с шестью парами диверкулов кишечника (пищеварительных желез). [ 27 ] [ 5 ] [ 32 ]
Мозг радиодонтов был проще, чем трехсегментный (состоящий из про-, дейто- и тритоцеребрума) мозг эуартропод , но дальнейшие интерпретации различаются в разных исследованиях. На основе Cong et al. 2014, мозг состоит только из одного сегмента мозга, происходящего из глазного сомита, протоцеребрума. Нервы лобных придатков и сложных глаз отходят от передних и латеральных отделов мозга. [ 9 ] [ 17 ] По данным Мойсюка и Кэрона, 2022, нервы лобных придатков возникли из вентрального дейтоцеребрума, второго сегмента мозга. Предыдущие «нервы лобного придатка» на самом деле представляют собой срединный глазной нерв. [ 22 ] В обеих интерпретациях позади мозга находилась пара явно несросшихся вентральных нервных шнуров , которые проходили через область шеи животного. [ 9 ] [ 22 ]
Палеоэкология
[ редактировать ]Физиология
[ редактировать ]
Радиодонты были интерпретированы как нектонные или нектобентосные животные, морфология которых предполагает активный плавательный образ жизни. Мускулистые, перекрывающиеся брюшные створки, возможно, продвигали животное по воде, возможно, двигаясь волнообразным образом, напоминающим современных скатов и каракатиц . [ 33 ] [ 34 ] Пары спинных крыльев, которые у некоторых видов образуют хвостовой веер, возможно, помогали управлять и/или стабилизировать животное во время передвижения. [ 10 ] [ 35 ] У Anomalocaris морфология веера хвоста даже позволяет предположить, что он может быстро и эффективно менять направление плавания. [ 36 ] значительно специализированными для нектобентосного образа жизни, например Cambroraster с его куполообразным Н-элементом, похожим на панцирь мечехвоста С другой стороны, некоторые хурдииды обладают особенностями , . [ 3 ] Полосы щетинковых пластинок со складчатыми ланцетными пластинками, возможно, увеличивали площадь поверхности, что позволяет предположить, что это были жабры функцию животного , обеспечивающие дыхательную . [ 27 ] [ 10 ] Обилие остатков склеритизованных структур, таких как разрозненные лобные придатки и комплексы склеритов головы, позволяет предположить, что линьки . среди радиодонтов могли происходить массовые события [ 10 ] [ 3 ] поведение, о котором также сообщалось у некоторых других кембрийских членистоногих, таких как трилобиты . [ 37 ]
Диета
[ редактировать ]





У радиодонтов были разнообразные стратегии питания, которых можно было разделить на хищных хищников , просеивателей отложений или суспензионных фильтраторов . [ 2 ] [ 38 ] [ 11 ] [ 39 ] [ 40 ] Например, хищные хищники, такие как Anomalocaris и Amplectobeluids, могли ловить ловкую добычу, используя свои хищные лобные придатки; последний даже имел прочный эндит, позволяющий удерживать добычу, как клешни . [ 26 ] [ 21 ] [ 4 ] [ 11 ] При меньшем головном панцирном комплексе и большой поверхности артродиальных мембран лобные придатки этих таксонов обладали большей гибкостью. [ 13 ] Толстые передние придатки просеивающих отложений, таких как Hurdia и Peytoia , имеют зазубренные эндиты с медиальной кривизной, которые могут образовывать корзинчатую ловушку для сгребания осадка и прохождения пищевых продуктов к хорошо развитому ротовому конусу. [ 3 ] [ 11 ] Эндиты лобных придатков взвеси/фильтраторов, таких как Tamisiocaris и Aegirocassis, имеют гибкие, плотно упакованные вспомогательные шипы, которые могут отфильтровывать органические компоненты, такие как мезозоопланктон и фитопланктон, размером до 0,5 мм. [ 7 ] [ 10 ] Фронтальные придатки Caryosyntrips , которые необычны для радиодонтов тем, что имеют направление несущих эндит поверхностей, противоположных друг другу, и, возможно, были способны манипулировать и раздавливать добычу ножницеобразными разрезающими или хватательными движениями. [ 21 ] [ 41 ]
Ротовые конусы радиодонтов могли использоваться для присасывания и/или кусания. [ 23 ] [ 38 ] [ 3 ] Наряду с большим разнообразием лобных придатков у разных видов радиодонтов, дифференциация ротовых шишек между видами позволяет предположить и предпочтения разных диет. [ 38 ] [ 11 ] Например, трехлучевой ротовой конус Anomalocaris с неровными бугорчатыми зубными пластинками и небольшим отверстием, возможно, был приспособлен к мелкой и нектонной добыче. [ 23 ] [ 11 ] в то время как жесткие тетрарадиальные оральные шишки Peytoia , Titanokorys , Hurdia и один изолированный ротовой конус, приписываемый Cambroraster, с большим отверстием и иногда дополнительными зубными пластинками, возможно, были способны потреблять более крупные пищевые продукты по сравнению с размером их тела и, вероятно, донную или эндобентосную добычу. . [ 23 ] [ 38 ] [ 3 ]
Классификация
[ редактировать ]Таксономическое сходство
[ редактировать ]
| |||||||||||||||||||||||||||||||||||||||||||||
| Обобщенная филогения Radiodonta и других таксонов Ecdysozoa. [ 42 ] |






-

-
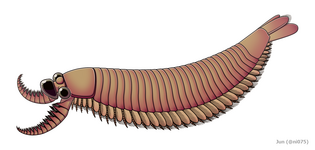 Радиодонт/ опабиниид - эуартропод , промежуточная килинксия , разделяет многие характеристики, обнаруженные как у динокаридид, так и у эуартропод.
Радиодонт/ опабиниид - эуартропод , промежуточная килинксия , разделяет многие характеристики, обнаруженные как у динокаридид, так и у эуартропод. -
 Реконструкция жизни базального дейтеропода Erratus , которая помогла выявить эволюцию придатков туловища членистоногих.
Реконструкция жизни базального дейтеропода Erratus , которая помогла выявить эволюцию придатков туловища членистоногих. -
 Мириддурин - динокаридидный панартропод из среднего ордовика, имеющий общие черты как с опабиниидами, так и с радиодонтами.
Мириддурин - динокаридидный панартропод из среднего ордовика, имеющий общие черты как с опабиниидами, так и с радиодонтами.




Большинство филогенетических анализов показывают, что радиодонты, наряду с опабиниидами ( Opabinia и Utaurora [ 43 ] ), являются стеблевой группы членистоногими , являющимися базальными по отношению к дейтероподам , [ 42 ] клада, включающая верхний стебель (например, fuxianhuiids и двустворчатые членистоногие) и корончатые Euarthropoda (например, Artiopoda , Chelicerata и Mandibulata ). [ 8 ] [ 44 ] [ 45 ] [ 46 ] [ 47 ] [ 48 ] [ 7 ] [ 9 ] [ 10 ] [ 2 ] [ 3 ] [ 30 ] [ 18 ] [ 39 ] [ 40 ] [ 49 ] [ 50 ] [ 43 ] Эта интерпретация подтверждается многочисленными планами строения членистоногих, обнаруженными на радиодонтах и опабиниидах, таких как на стебельках сложные глаза , [ 25 ] пищеварительные железы, [ 32 ] туловищные придатки, образующиеся дорсальными и вентральными элементами (предшественники двуветвистых придатков членистоногих). [ 10 ] [ 50 ] По сравнению с опабиниидами, имеющими заднее ротовое отверстие и почти сросшиеся передние придатки (сравнимо с обращенным назад комплексом губа /гипостома у эуартропод), [ 17 ] [ 43 ] радиодонты, с другой стороны, имели дорсальный склерит, напоминающий эуартропод (H-элемент), и членистоногие (лобные придатки) в областях головы, [ 51 ] [ 17 ] [ 43 ] рядом с кутикуляризованными концами кишки. [ 27 ] Тот факт, что как у радиодонтов, так и у опабиниид отсутствует экзоскелет в области туловища, позволяет предположить, что возникновение сложных глаз и членистоногих (сегментированных придатков) предшествует артродизации (полный набор экзоскелета туловища) в стволовой линии членистоногих. [ 42 ] [ 52 ] [ 53 ] Суженная область шеи с питающими аппендикулярными структурами некоторых радиодонтов также может пролить свет на происхождение сложной головы членистоногого, образовавшейся в результате слияния множества передних сегментов тела. [ 4 ] [ 17 ] Базальные дейтроподы, обладающие смесью признаков радиодонтов и опабиниид, таких как Kylinxia и Erratus , могут представлять собой промежуточные формы между радиодонтами, опабиниидами и другими эуартроподами. [ 18 ] [ 50 ]
-
 Сибирские лобоподии Siberion (вверху слева), Megadictyon (внизу в центре) и Jianshanopodia (вверху справа)
Сибирские лобоподии Siberion (вверху слева), Megadictyon (внизу в центре) и Jianshanopodia (вверху справа) -
-
 Мегахейра , или «большие членистоногие с придатками», класс возможных стволовых хелицератов, ранее считавшихся близким родственником радиодонта.
Мегахейра , или «большие членистоногие с придатками», класс возможных стволовых хелицератов, ранее считавшихся близким родственником радиодонта.



Таксоны, базальные к ветви радиодонтов, опабиниид и эуартропод, представляют собой « жаберные лобоподии », такие как Pambdelurion и Kerygmachela , которые иногда объединялись в класс Dinocaridida с опабибиидами и радиодонтами. [ 54 ] [ 46 ] У них есть створки тела, пищеварительные железы, большие (предположительно сложные) глаза и специализированные лобные придатки, как у предыдущих таксонов, но их лобные придатки не членистоногие и не слиты, глаза сидячие, жаберные структуры менее заметны и, конечно же, под каждым из них имеются лобоподы. их закрылки. [ 55 ] [ 10 ] [ 56 ] [ 43 ] Таксонами, даже базальными по отношению к «жаберным лобоподиям», являются сиберииды, такие как Megadictyon и Jianshanopodia . [ 42 ] группа лобоподов , у которых были крепкие лобные придатки и пищеварительные железы, но не было створок тела. Такие промежуточные формы между лобоподиями и радиодонтами/эуартроподами предполагают, что вся группа Arthropoda возникла из парафилетического класса лобоподий, наряду с двумя другими существующими панартропод типами Tardigrada и Onychophora . [ 57 ] [ 42 ] [ 17 ] [ 58 ] [ 52 ] [ 53 ]
Предыдущие исследования могут предполагать, что радиодонты относятся к группе, отличной от стволовых членистоногих, например, к неизвестному до сих пор типу ; [ 33 ] циклоневралии n черви, подвергшиеся конвергенции с членистоногими (на основе циклоневралианоподобного радиального ротового аппарата); [ 59 ] [ 54 ] стеблевые хелицеровые эуартроподы наряду с мегахейранами , также известными как большие придаточные членистоногие (на основании сходства между радиодонтными лобными придатками, большими придатками мегахейран и хелицерами ); [ 60 ] или Schinderhannes bartelsi , оказался хурдиидным радиодонтом, который в недавних анализах [ 42 ] [ 7 ] [ 10 ] [ 2 ] [ 3 ] [ 39 ] [ 40 ] как вид, более тесно связанный с эуартроподами, чем с другими радиодонтами (на основании некоторых предполагаемых черт, напоминающих эуартропод, обнаруженных на Schinderhannes ). [ 35 ] Однако ни одно из них не было подтверждено последующими расследованиями. Радиальные ротовые части не являются исключительными для циклонейралий и, скорее всего, представляют собой результат конвергентной эволюции или экдизозоа таких плезиморфии , поскольку они также были обнаружены у панартропод, как тихоходки и некоторые лобоподии ; [ 61 ] радиодонты, у которых отсутствуют характерные особенности эуартропод, такие как тергиты туловища и множественные придатки головы, [ 42 ] а большие придатки мегахейрана считались дейтоцеребральными, [ 62 ] [ 63 ] которые могли быть негомологичны радиодонтным протоцеребральным лобным придаткам; [ 9 ] [ 17 ] предполагаемые признаки эуартропод, обнаруженные на единственной окаменелости Шиндерханнеса , сомнительны и могут представлять собой другие структуры, подобные радиодонтам. [ 42 ]
Взаимосвязи
[ редактировать ]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Филогения Radiodonta по Мойсюку и Карону, 2021 г. [ 39 ] |








Традиционно все виды радиодонтов относили к одному семейству Anomalocarididae . [ 6 ] отсюда и предыдущее общее название «аномалокаридид». [ 26 ] [ 8 ] и иногда он все еще использовался для обозначения всего ордена даже после реклассификации. [ 9 ] [ 10 ] После переназначения Vinther et al. В 2014 году большинство видов радиодонтов были переклассифицированы в три новых семейства: Amplectobeluidae , Tamisiocarididae. [ 2 ] [ 3 ] (ранее Cetiocaridae [ 7 ] ) и Hurdiidae . [ 7 ] [ 10 ] [ 2 ] [ 3 ] Четыре современных семейства радиодонтов, включая Anomalocarididae, могут образовывать кладу Anomalocarida. [ 7 ]
Первоначальное описание отряда Radiodonta включало Anomalocaris , Laggania (позже известную как Peytoia ), Hurdia , Proboscicaris , Amplectobelua , Cucumericrus и Parapeytoia . [ 6 ] Однако теперь Proboscicaris рассматривается как младший синоним Hurdia , а Parapeytoia считается мегахейраном . [ 8 ] [ 27 ] [ 10 ] Из-за ограниченного открытия положение Cucumericrus внутри Radiodonta неясно, поскольку он не был выбран с помощью филогенетического анализа. [ 7 ] [ 3 ] [ 2 ] [ 39 ] [ 40 ] или разрешается политомией с Radiodonta и Euarthropoda . [ 10 ] [ 13 ]
-
 Одна из малоизвестных частей тела (придаток туловища) Cucumericrus Decoratus ; этот вид может не представлять собой настоящего радиодонта.
Одна из малоизвестных частей тела (придаток туловища) Cucumericrus Decoratus ; этот вид может не представлять собой настоящего радиодонта. -
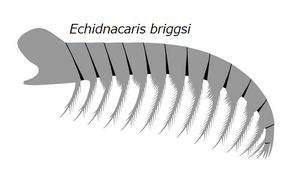 Лобный придаток Echidnacaris briggsi , радиодонта тамизиокарида , который когда-то предполагался принадлежащим к роду Anomalocaris до его описания в 2023 году.
Лобный придаток Echidnacaris briggsi , радиодонта тамизиокарида , который когда-то предполагался принадлежащим к роду Anomalocaris до его описания в 2023 году.


Первый углубленный филогенетический анализ Radiodonta был проведен Vinther et al . в 2014 году, [ 7 ] за которым последовало несколько дополнительных исследований с более или менее измененными результатами. [ 9 ] [ 10 ] [ 2 ] [ 13 ] [ 3 ] [ 39 ] [ 40 ] [ 43 ] В большинстве анализов Caryosyntrips является самым базальным родом, но он либо разрешается в политомии с другими радиодонтами и Euarthropoda (наряду с Cucumericrus , если он включен). [ 10 ] [ 13 ] ) или за пределами Radiodonta, что ставит под сомнение его родство с радиодонтом. [ 64 ] За исключением сомнительных Caryosyntrips и Cucumericrus , монофилия Radiodonta. широко поддерживается [ 7 ] [ 9 ] [ 10 ] [ 2 ] [ 13 ] [ 3 ] [ 39 ] [ 40 ] с некоторыми результатами можно предположить возможную парафилию (либо клада Anomalocarididae + Amplectobeluidae, либо сестра Hurdiidae Euarthropoda). [ 30 ] [ 43 ] Предполагаемые синапоморфии монофилетических Radiodonta, включая трехчастный комплекс склеритов головы и дифференцированную область шеи. [ 3 ] Род Anomalocaris в более широком смысле всегда считается полифилетическим , обычно с «Anomalocaris» kunmingensis и «Anomalocaris» briggsi, отнесенными к Amplectobeluidae и Tamisiocarididae соответственно. [ 7 ] [ 9 ] [ 10 ] [ 2 ] [ 3 ] [ 39 ] [ 40 ] Взаимосвязь Amplectobeluidae неясна, поскольку сходство Lyrarapax и Ramskoeldia с амплектобелуидами иногда подвергалось сомнению. [ 1 ] [ 3 ] [ 40 ] Монофилия особого семейства Hurdiidae была обнаружена большинством анализов и хорошо подтверждена несколькими синапоморфиями (например, дистальная сочлененная область лобного придатка с проксимальными 5 подомерами, несущими почти одинаковые эндиты). [ 19 ] [ 3 ] ). В 2010-х годах Tamisiocarididae часто считали сестринской группой Hurdiidae. [ 7 ] [ 10 ] [ 2 ] [ 3 ] но эта позиция стала сомнительной в последующих исследованиях. [ 22 ] [ 24 ]
- Радиодонта
- ? Огурец [ 26 ] (родство радиодонтов сомнительно [ 65 ] [ 64 ] )
- ? Кариосинтрипсы [ 21 ] (родство радиодонтов сомнительно [ 64 ] )
- Аномалокарида
- Параномалокарис [ 66 ] (по некоторым исследованиям отнесен к Anomalocarididae. [ 13 ] [ 67 ] )
- Ламинакарис [ 68 ] (по некоторым исследованиям отнесен к Amplectobeluidae. [ 2 ] )
- Houcaris (либо отнесенный к Anomalocarididae, [ 10 ] [ 13 ] [ 43 ] Амлектобелуиды [ 7 ] [ 2 ] или Tamisiocarididae. [ 69 ] [ 39 ] )
- Вы инновационный [ 70 ]
- Аномалокаридиды
- Anomalocaris (в более широком смысле некоторые виды могут быть отнесены к другим семействам. [ 7 ] [ 10 ] )
- Вы будете смягчены [ 20 ]
- Shucaris (отнесенный либо к Anomalocarididae, либо к Amplectobeluidae). [ 71 ] [ 64 ] )
- Амлектобелуиды
- Лирарапакс [ 9 ] (позиция подвергается сомнению в некоторых исследованиях. [ 1 ] )
- Эмлектобелуа [ 26 ]
- Рамскоэльдия [ 1 ] (позиция подвергается сомнению в некоторых исследованиях. [ 3 ] )
- Гуаншанкарис [ 67 ] [ 72 ]
- Shucaris (отнесенный либо к Anomalocarididae, либо к Amplectobeluidae). [ 71 ] [ 64 ] )
- Тамизиокаридиды
- Hurdiidae / Peytoiidae [ 64 ]
- Эгирокассисины [ 73 ]
- Пейтойя
- Шиндерханнес (позиция подвергается сомнению в некоторых исследованиях. [ 74 ] [ 75 ] [ 76 ] )
- Хурдия
- Стэнликарис [ 77 ]
- Пахвантия [ 2 ]
- Урсулинакарис [ 19 ]
- Камброрастер [ 3 ]
- ? Женгекарис (предполагаемый радиодонт хурдиид [ 65 ] )
- Вы убиты горем [ 78 ]
- Буккаспинея [ 79 ]
- Титанокори [ 16 ]
| Разновидность | Исходное описание | Год назван | Семья | Возраст | Расположение | Лобный придаток | Головной склеритный комплекс |
|---|---|---|---|---|---|---|---|
| огурец Украшенный | Хоу, Бергстрем и Альберг | 1995 [ 26 ] | (не назначено) | Кембрийский этап 3 | Неизвестный | Неизвестный | |
| Кариосинтрипс зубчатый | Дэйли и Бадд | 2010 [ 21 ] | (не назначено) | Улиуань – Друмиан | 
|
Неизвестный | |
| Кариосинтрипс камурус | Пейтс и Дэйли | 2017 [ 41 ] | (не назначено) | Улиуань | 
|
Неполный [ 79 ] | |
| Кариосинтрипс твердый | Пейтс и Дэйли | 2017 [ 41 ] | (не назначено) | барабанщик | 
|
Неизвестный | |
| Параномалокарис многосегментарный | Ван, Хуан и Ху | 2013 [ 66 ] | Аномалокаридиды ? | Кембрийский этап 4 | 
|
Неизвестный | |
| Параномалокарис простой | Цзяо, Пейтс, Лерози-Обрил, Ортега-Эрнандес, Ян, Лан, Чжан | 2021 [ 67 ] | Аномалокаридиды ? | Кембрийский этап 4 | 
|
Неизвестный | |
| Ламинакарис химера | Го, Пейтс, Конг, Дейли, Эджкомб, Чен и Хоу | 2018 [ 68 ] | (спорный) | Кембрийский этап 3 | 
|
Неизвестный | |
| Инновациякарис маотианшаненсис | Цзэн, Чжао, Чжу | 2022 [ 70 ] | (не назначено) | Кембрийский этап 3 | 
|
P-элемент неизвестен [ 70 ] | |
| Инновации ? multispiniformis | Цзэн, Чжао, Чжу | 2022 [ 70 ] | (не назначено) | Кембрийский этап 3 | 
|
Неизвестный | |
| Аномалокарис канадский | Уайтивс | 1892 [ 80 ] | Аномалокаридиды | Улиуань | 
|

| |
| Lenisicaris pennsylvanica (ранее Anomalocaris pennsylvanica ) [ 20 ] | Сдерживать | 1929 | Аномалокаридиды | Кембрийский этап 3 | 
|
Неизвестный | |
| Ленисикарис тряпка | У, Ма, Линь, Сунь, Чжан и Фу | 2021 [ 20 ] | Аномалокаридиды | Кембрийский этап 3 | 
|
Неизвестный | |
| Аномалокарис далея | Патерсон, Гарсиа-Беллидоб и Эджкомб | 2023 | Аномалокаридиды | Кембрийский этап 4 | 
|
Неизвестный | |
| Houcaris magnabasis (ранее Anomalocaris magnabasis ) [ 69 ] | Пейтс, Дейли, Эджкомб, Конг и Либерман | 2019 | (спорный) | Кембрийский этап 4 | 
|
Неизвестный | |
| Houcaris saron (ранее Anomalocaris saron ) [ 69 ] | Хоу, Бергстрем и Альберг | 1995 | (спорный) | Кембрийский этап 3 | 
|
Неизвестный | |
| Эхиднакарис Бриггсия [ 24 ] | Не давай | 1995 | Тамизиокаридиды | Кембрийский этап 4 | 
|
Возможный H-элемент и уникальные латеральные склериты, связанные со сложными глазами [ 14 ] [ 24 ] | |
| Рамскоэльдия платьяканта | Конг, Эджкомб, Дейли, Го, Пейтс и Хоу | 2018 [ 1 ] | Амлектобелуиды | Кембрийский этап 3 | 
|
Неполный [ 1 ] | |
| Рамскоэльдиа похоже на | Конг, Эджкомб, Дейли, Го, Пейтс и Хоу | 2018 [ 1 ] | Амлектобелуиды | Кембрийский этап 3 | 
|
Неполный [ 1 ] | |
| Лирарапакс клауспинус | Конг, Ма, Хоу, Эджкомб и Штраусфилд | 2014 [ 9 ] | Амлектобелуиды | Кембрийский этап 3 | 
|
Горловина P-элемента неизвестна | |
| Лирарапакс трилобус | Конг, Дейли, Эджкомб, Хоу и Чен | 2016 [ 5 ] | Амлектобелуиды | Кембрийский этап 3 | 
|
P-элемент неизвестен | |
| Эмблектобелуа симбрациата | Хоу, Бергстрем и Альберг | 1995 [ 26 ] | Амлектобелуиды | Кембрийский этап 3 | 
|

| |
| Эмблектобелуа стефененсис | Дэйли и Бадд | 2010 [ 21 ] | Амлектобелуиды | Улиуань | 
|
Неизвестный | |
| Гуаншанкарис кунмингенсис | Чжан и др. | 2023 [ 72 ] | Амлектобелуиды | Кембрийский этап 4 | 
|
Неизвестный | |
| Тамисиокарис северный | Дэйли и Пил | 2010 | Тамизиокаридиды | Кембрийский этап 3 | 
|
Неполный [ 7 ] | |
| Урсулинакарис grallae | Пейтс, Дейли и Баттерфилд | 2019 | Hurdiidae | Улиуань | 
|
Неизвестный | |
| Шиндерханнес Бартельси | Кюль, Бриггс и Раст | 2009 [ 35 ] | Hurdiidae | Эмсиан | Неполный [ 3 ] | Неполный [ 3 ] | |
| Стэнликарис гирпекс | Пейтс, Дейли и Ортега-Эрнандес | 2018 [ 77 ] | Hurdiidae | Улиуань | 
|
P-элемент неизвестен, возможно, отсутствует [ 22 ] | |
| Пейтоя натхорсти | Уолкотт | 1911 [ 81 ] | Hurdiidae | Улиуань – Друмиан | 
|
Неполный [ 3 ] | |
| Peytoia infercambriensis (ранее Cassubia infercambriensis ) [ 82 ] | Лендзион | 1975 | Hurdiidae | Кембрийский этап 3 | 
|
Неизвестный | |
| Эгирокассис бенмулай | Ван Рой, Дейли и Бриггс | 2015 [ 10 ] | Hurdiidae ( Aegirocassisinae ) | Тремадокийский | 
|

| |
| Хурдия Виктория | Уолкотт | 1912 [ 83 ] | Hurdiidae | Улиуань – Друмиан | 
|

| |
| Хурдия треугольная | Уолкотт | 1912 [ 83 ] | Hurdiidae | Улиуань |
|

| |
| Камброрастером Скошено | Мойсюк и Карон | 2019 [ 3 ] | Hurdiidae | Улиуань | 
|

| |
| Пахвантия пронзена копьем | Робисон и Ричардс | 1981 | Hurdiidae | барабанщик | 
|

| |
| Кордатикарис полосатый | Сунь, Цзэн и Чжао | 2020 [ 78 ] | Hurdiidae | барабанщик | Неполный [ 78 ] | 
| |
| Женгекарис Шанкоуэнсис | Ваннер, Чен, Хуан, Шарбонье и Ван | 2006 | Hurdiidae | Кембрийский этап 3 | Неизвестный |  | |
| я прикрыл рот | Пейтс, Лерози-Обрил, Дейли, Кир, Бонино и Ортега-Эрнандес | 2021 [ 79 ] | Hurdiidae | барабанщик | 
|
Неизвестный | |
| Титанокори гейнеси | Карон и Мойсюк | 2021 [ 40 ] | Hurdiidae | Улиуань | 
|

| |
| Псевдоангустидонтус двойной остистый | Ван Рой и Тетли | 2006 | Hurdiidae ( Aegirocassisinae ) | Тремадокийский | 
|
Неизвестный | |
| Псевдоангустидонтус издигуа | Потин, Герио и Дейли | 2023 | Hurdiidae ( Aegirocassisinae ) | Тремадокийский | 
|
Неполный [ 73 ] | |
| Шукарис анкилосскелос | Ву и др. | 2024 | (спорный) | Кембрийский этап 2 – Кембрийский этап 3 |  |
Неполный [ 71 ] |
История
[ редактировать ]
-
 Лобный придаток Anomalocaris canadensis
Лобный придаток Anomalocaris canadensis -
 Оральный конус Peytoia nathorsti
Оральный конус Peytoia nathorsti -
 H-элемент Hurdia victoria
H-элемент Hurdia victoria -
![Парные лобные придатки безымянного радиодонта-хурдиида[39]](//upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg) Парные лобные придатки безымянного хурдиида . радиодонта- [ 39 ]
Парные лобные придатки безымянного хурдиида . радиодонта- [ 39 ]



![Парные лобные придатки безымянного радиодонта-хурдиида[39]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)
История радиодонтов сложна. Неполные экземпляры, относящиеся к разным частям тела одного и того же вида, исторически интерпретировались как принадлежащие к разным видам и даже разным типам. [ 6 ] [ 8 ] До того, как они были признаны группой, образцы радиодонтов были отнесены к пяти различным типам: Porifera , Cnidaria , Echinodermata , Annelida и Arthropoda . [ 6 ]
Первые известные образцы радиодонтов были собраны из пластов трилобитов горы Стивен Ричардом Г. МакКоннеллом из Геологической службы Канады в 1886 году. [ 6 ] или 1888 год. [ 80 ] Эти экземпляры были названы Anomalocaris canadensis в 1892 году палеонтологом GSC Джозефом Уайтавсом . [ 80 ] Уайтэйвс интерпретировал эти экземпляры, которые теперь известны как изолированные лобные придатки, как брюшко филлокарида ракообразного . [ 80 ] Дополнительные образцы радиодонтов были описаны в 1911 году Чарльзом Уолкоттом . [ 81 ] Изолированную ротовую шишку, которую он назвал Peytoia nathorsti , он интерпретировал как медузу, а плохо сохранившийся, но относительно полный экземпляр, который он назвал Laggania cambria , — как голотурию. [ 81 ] В 1912 году Уолкотт назвал Hurdia victoria и H. triangulata на основе изолированных H-элементов, которые он интерпретировал как панцири ракообразных. [ 83 ] Изолированные лобные придатки Peytoia и Hurdia были идентифицированы как придатки Sidneyia . , известные под общим названием «Придаток F» у Briggs 1979, в то время [ 81 ] Proboscicaris в P-элемент Hurdia был назван 1962 году и интерпретирован как панцирь двустворчатого членистоногого. [ 84 ]
Геологическая служба Канады инициировала пересмотр окаменелостей сланцев Бёрджесс в 1966 году под руководством Кембриджского университета палеонтолога Гарри Б. Уиттингтона . [ 6 ] Этот пересмотр в конечном итоге приведет к открытию полного плана тела радиодонта. В 1978 году Саймон Конвей Моррис признал, что ротовой аппарат лаггании напоминал пейтою , но интерпретировал это как свидетельство того, что это было сложное ископаемое, состоящее из медузы пейтоя и губки. [ 85 ] В 1979 году Дерек Бриггс признал, что окаменелости Anomalocaris были придатками, а не брюшком, но интерпретировал их как ходильные ноги рядом с «придатком F». [ 86 ] Лишь в 1985 году была признана истинная природа окаменелостей Anomalocaris , Laggania и Peytoia , и все они были отнесены к одному роду Anomalocaris . [ 33 ] Впоследствии было признано, что Anomalocaris представляет собой отдельную форму от двух других, что привело к разделению на два рода, последний из которых по-разному назывался Laggania и Peytoia, пока не было установлено, что оба представляют один и тот же вид, и Peytoia имеет приоритет. [ 23 ] Позже было признано, что некоторые окаменелости, отнесенные к этим таксонам, принадлежали другой форме, у которой был признан панцирь, состоящий из элементов Hurdia и Proboscicaris . Наконец, в 2009 году эти экземпляры были переописаны как Hurdia . [ 8 ] Даже после этих открытий частичные ошибочные идентификации (например, ротовой конус и лобные придатки пейтои) были отнесены к Anomalocaris. [ 6 ] и Хурдия , [ 8 ] соответственно) были выявлены и в последующих исследованиях. [ 23 ] [ 87 ]
Сам таксон Radiodonta был придуман в 1996 году Десмондом Коллинзом после того, как было установлено, что Anomalocaris и его родственники представляют собой особую линию, имеющую сходство с членистоногими, а не неизвестный до сих пор тип. [ 6 ] Коллинз также установил, что класс Dinocarida включает отряд Radiodonta, а также Opabiniidae , которые он признал отдельными из-за отсутствия у них характерной структуры орального конуса, свойственной радиодонтам. [ 6 ] Впервые Radiodonta получила филогенетическое определение в 2014 году. [ 7 ] Изначально считалось, что Radiodonta содержит одно семейство Anomalocarididae , но в 2014 году оно было разделено на четыре семейства: Amplectobeluidae , Anomalocarididae, Cetiocaridae и Hurdiidae . [ 7 ] Название Cetiocarididae не соответствовало Международному кодексу зоологической номенклатуры, поэтому в 2019 году было переименовано в Tamisiocarididae . [ 88 ]
До 2010-х годов радиодонты обычно считались одинаково крупными высшими хищниками, но открытие новых видов в течение этого десятилетия привело к значительному увеличению известного экологического и морфологического разнообразия группы. [ 7 ] [ 10 ] [ 2 ] [ 3 ] [ 89 ] [ 79 ] [ 39 ] [ 40 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г час я дж к Конг, Пэй-Юнь; Эджкомб, Грегори Д.; Дейли, Эллисон С.; Го, Цзинь; Пейтс, Стивен; Хоу, Сянь-Гуан (2018). «Новые радиодонты с гнатобазоподобными структурами из кембрийской биоты Чэнцзян и значение для систематики Radiodonta» . Статьи по палеонтологии . 4 (4): 605–621. дои : 10.1002/spp2.1219 . ISSN 2056-2802 . S2CID 90258934 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Лерози-Обрил, Руди; Пейтс, Стивен (14 сентября 2018 г.). «Новый радиодонт, питающийся взвесью, предполагает эволюцию микропланктивных животных в кембрийском макронектоне» . Природные коммуникации . 9 (1): 3774. Бибкод : 2018NatCo...9.3774L . дои : 10.1038/s41467-018-06229-7 . ISSN 2041-1723 . ПМК 6138677 . ПМИД 30218075 . Данные дриад
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из в ах есть также и аль являюсь а Мойсюк Ю.; Карон, Ж.-Б. (14 августа 2019 г.). «Новый радиодонт-хурдиид из сланцев Берджесс свидетельствует об эксплуатации кембрийских инфаунистических источников пищи» . Труды Королевского общества B: Биологические науки . 286 (1908): 20191079. doi : 10.1098/rspb.2019.1079 . ПМК 6710600 . ПМИД 31362637 .
- ^ Jump up to: а б с д и ж г час Конг, Пэйюнь; Дейли, Эллисон С.; Эджкомб, Грегори Д.; Хоу, Сянгуан (30 августа 2017 г.). «Функциональная голова кембрийского радиодонта (стволовой группы Euarthropoda) Amplectobelua symbrachiata » . Эволюционная биология BMC . 17 (1): 208. дои : 10.1186/s12862-017-1049-1 . ISSN 1471-2148 . ПМК 5577670 . ПМИД 28854872 .
- ^ Jump up to: а б с д и ж Конг, Пэйюнь; Дейли, Эллисон С.; Эджкомб, Грегори Д.; Хоу, Сянгуан; Чен, Айлин (сентябрь 2016 г.). «Морфология радиодонтана Lyrarapax из биоты Чэнцзян раннего кембрия» . Журнал палеонтологии . 90 (4): 663–671. дои : 10.1017/jpa.2016.67 . ISSN 0022-3360 . S2CID 88742430 .
- ^ Jump up to: а б с д и ж г час я дж к л м Коллинз, Десмонд (1996). «Эволюция» Anomalocaris и его отнесение к классу членистоногих Dinocarida (nov.) и отряду Radiodonta (nov.)» . Журнал палеонтологии . 70 (2): 280–293. дои : 10.1017/S0022336000023362 . S2CID 131622496 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т Винтер, Якоб; Штейн, Мартин; Лонгрич, Николас Р.; Харпер, Дэвид А.Т. (2014). «Аномалокарид, питающийся суспензией, из раннего кембрия» (PDF) . Природа . 507 (7493): 496–499. Бибкод : 2014Natur.507..496V . дои : 10.1038/nature13010 . ПМИД 24670770 . S2CID 205237459 .
- ^ Jump up to: а б с д и ж г час я дж Дейли, Эллисон С.; Бадд, Грэм Э.; Карон, Жан-Бернар; Эджкомб, Грегори Д.; Коллинз, Десмонд (2009). из сланца Берджесс «Аномалокаридид Hurdia и его значение для ранней эволюции членистоногих». Наука . 323 (5921): 1597–1600. Бибкод : 2009Sci...323.1597D . дои : 10.1126/science.1169514 . ПМИД 19299617 . S2CID 206517995 .
- ^ Jump up to: а б с д и ж г час я дж к л м н Конг, Пэйюнь; Ма, Сяоя; Хоу, Сянгуан; Эджкомб, Грегори Д.; Штраусфельд, Николас Дж. (2014). «Структура мозга устраняет сегментарное сродство аномалокаридидных придатков». Природа . 513 (7519): 538–42. Бибкод : 2014Natur.513..538C . дои : 10.1038/nature13486 . ПМИД 25043032 . S2CID 4451239 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб и объявление но из Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2015). «Гомология конечностей туловища аномалокаридид, выявленная гигантским фильтратором с парными створками». Природа . 522 (7554): 77–80. Бибкод : 2015Natur.522...77V . дои : 10.1038/nature14256 . ПМИД 25762145 . S2CID 205242881 .
- ^ Jump up to: а б с д и ж г час Де Виво, Джачинто; Лаутеншлагер, Стефан; Винтер, Якоб (28 июля 2021 г.). «Трехмерное моделирование, неравенство и экология первых кембрийских хищников вершины» . Труды Королевского общества B: Биологические науки . 288 (1955): 20211176. doi : 10.1098/rspb.2021.1176 . ПМЦ 8292756 . ПМИД 34284622 .
- ^ Фостер, Дж. (2014). «Магический загадочный тур: Биологическая психоделия сланцев Бёрджесс». Кембрийский океанический мир: древняя морская жизнь Северной Америки . Издательство Университета Индианы. стр. 195–252. ISBN 978-0253011824 .
- ^ Jump up to: а б с д и ж г час я дж Лю, Цзянни; Лерози-Обрил, Руди; Штайнер, Майкл; Данлоп, Джейсон А.; Шу, Деган; Патерсон, Джон Р. (1 ноября 2018 г.). «Происхождение хищного питания у молодых членистоногих, выявленное кембрийским радиодонтаном» . Национальный научный обзор . 5 (6): 863–869. дои : 10.1093/nsr/nwy057 . ISSN 2095-5138 .
- ^ Jump up to: а б с Пейтс, Стивен; Боттинг, Джозеф П.; Маккобб, Люси МЭ; Мьюир, Люси А. (2020). «Миниатюрная ордовикская хурдия из Уэльса демонстрирует приспособляемость Radiodonta» . Королевское общество открытой науки . 7 (6): 200459. Бибкод : 2020RSOS....700459P . дои : 10.1098/rsos.200459 . ISSN 2054-5703 . ПМЦ 7353989 . ПМИД 32742697 .
- ^ Ву, Ю; Пейтс, Стивен; Поли, Дэниел; Чжан, Синлян; Фу, Дунцзин (3 ноября 2023 г.). «Быстрый рост крупного кембрийского хищника» . Национальный научный обзор . дои : 10.1093/nsr/nwad284 . ISSN 2095-5138 . ПМЦ 10833464 .
- ^ Jump up to: а б Карон, Ж.-Б.; Мойсюк Ю. (сентябрь 2021 г.). «Гигантский нектобентосный радиодонт из сланцев Берджесс и значение разнообразия панцирей хурдиид» . Королевское общество открытой науки . 8 (9): 210664. Бибкод : 2021RSOS....810664C . дои : 10.1098/rsos.210664 . ПМЦ 8424305 . ПМИД 34527273 .
- ^ Jump up to: а б с д и ж г час я Ортега-Эрнандес, Хавьер; Янссен, Ральф; Бадд, Грэм Э. (1 мая 2017 г.). «Происхождение и эволюция головы панартропод - палеобиологическая перспектива и перспектива развития» . Строение и развитие членистоногих . Эволюция сегментации. 46 (3): 354–379. дои : 10.1016/j.asd.2016.10.011 . ISSN 1467-8039 . ПМИД 27989966 .
- ^ Jump up to: а б с д Цзэн, Хань; Чжао, Кеченг; Хуан, Диин (2020). членистоногое животное с хищными придатками, . кембрийский похожими » радиодонтов на « Ранний .588..101Z .doi : 10.1038 s41586-020-2883-7 ISSN 1476-4687 . PMID 33149303 /
- ^ Jump up to: а б с д и ж Пейтс, Стивен; Дейли, Эллисон С.; Баттерфилд, Николас Дж. (11 июня 2019 г.). «Первое сообщение о парных вентральных эндитах у хардидного радиодонта» . Зоологические письма . 5 (1): 18. дои : 10.1186/s40851-019-0132-4 . ISSN 2056-306X . ПМК 6560863 . ПМИД 31210962 .
- ^ Jump up to: а б с д и Ву, Ю; Ма, Цзясинь; Линь, Вэйлян; Солнце, АО; Чжан, Синлян; Фу, Дунцзин (1 мая 2021 г.). «Новые аномалокаридиды (Panarthropoda: Radiodonta) из нижнего кембрия Chengjiang Lagerstätte: биостратиграфические и палеобиогеографические последствия» . Палеогеография, Палеоклиматология, Палеоэкология . 569 : 110333. Бибкод : 2021PPP...569k0333W . дои : 10.1016/j.palaeo.2021.110333 . ISSN 0031-0182 . S2CID 233565727 .
- ^ Jump up to: а б с д и ж г час Дейли, Эллисон С.; Бадд, Грэм Э. (2010). «Новые придатки аномалокаридид из сланцев Бёрджесс, Канада» . Палеонтология . 53 (4): 721–738. Бибкод : 2010Palgy..53..721D . дои : 10.1111/j.1475-4983.2010.00955.x . ISSN 1475-4983 .
- ^ Jump up to: а б с д и ж г Мойсюк, Иосиф; Карон, Жан-Бернар (8 августа 2022 г.). «Трехглазый радиодонт с окаменелой нейроанатомией сообщает о происхождении головы и сегментации членистоногих» . Современная биология . 32 (15): 3302–3316.e2. дои : 10.1016/j.cub.2022.06.027 . ISSN 0960-9822 . ПМИД 35809569 . S2CID 250361698 .
- ^ Jump up to: а б с д и ж Дейли, Эллисон С.; Бергстрем, Ян (апрель 2012 г.). «Ротовая шишка Anomalocaris — это не классическая пейтоя » . Naturwissenschaften . 99 (6): 501–504. Бибкод : 2012NW.....99..501D . дои : 10.1007/s00114-012-0910-8 . ISSN 0028-1042 . ПМИД 22476406 . S2CID 2042726 .
- ^ Jump up to: а б с д Патерсон, Джон Р.; Гарсиа-Беллидо, Диего К.; Эджкомб, Грегори Д. (1 января 2023 г.). «Возвращение к радиодонтам ранних кембрийских сланцев Эму-Бей: морфология и систематика» . Журнал систематической палеонтологии . 21 (1). дои : 10.1080/14772019.2023.2225066 . ISSN 1477-2019 . S2CID 259719252 .
- ^ Jump up to: а б Штраусфельд, Николас Дж.; Ма, Сяоя; Эджкомб, Грегори Д.; Форти, Ричард А.; Лэнд, Майкл Ф.; Лю, Ю; Конг, Пэйюнь; Хоу, Сянгуан (август 2015 г.). «Глаза членистоногих: летопись окаменелостей раннего кембрия и дивергентная эволюция зрительных систем» . Строение и развитие членистоногих . 45 (2): 152–172. дои : 10.1016/j.asd.2015.07.005 . ПМИД 26276096 .
- ^ Jump up to: а б с д и ж г Сиань-Гуан, Хоу; Бергстрем, Ян; Альберг, Пер (сентябрь 1995 г.). « Anomalocaris и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая». ГФФ . 117 (3): 163–183. дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Jump up to: а б с д и ж г час я Дейли, Эллисон С.; Эджкомб, Грегори Д. (2014). «Морфология Anomalocaris canadensis из сланцев Берджесс». Журнал палеонтологии . 88 (1): 68–91. дои : 10.1666/13-067 . S2CID 86683798 .
- ^ Дейли, Эллисон С.; Патерсон, Джон Р.; Эджкомб, Грегори Д.; Гарсиа-Беллидо, Диего К.; Джаго, Джеймс Б. (2013). «Новая анатомическая информация об Anomalocaris из кембрийских сланцев залива Эму в Южной Австралии и переоценка его предполагаемых хищнических привычек» . Палеонтология . 56 (5): 971–990. дои : 10.1111/пала.12029 . ISSN 1475-4983 . S2CID 128955512 .
- ^ Jump up to: а б с Чен, Цзюнь-юань; Рамшельд, Ларс; Чжоу, Гуй-цин (27 мая 1994 г.). «Доказательства монофилии и близости кембрийских гигантских хищников к членистоногим» . Наука . 264 (5163): 1304–1308. Бибкод : 1994Sci...264.1304C . дои : 10.1126/science.264.5163.1304 . ПМИД 17780848 . S2CID 1913482 .
- ^ Jump up to: а б с Ария, Седрик; Чжао, Фанчэн; Цзэн, Хан; Го, Цзинь; Чжу, Маоянь (8 января 2020 г.). «Окаменелости из Южного Китая дают новое определение строению тела предков эуартропод» . Эволюционная биология BMC . 20 (1): 4. дои : 10.1186/s12862-019-1560-7 . ISSN 1471-2148 . ПМК 6950928 . ПМИД 31914921 .
- ^ Ван Рой, Питер; Дейли, Эллисон С.; Бриггс, Дерек Э.Г. (2013). Аномалокаридиды имели два набора боковых створок . 57-е ежегодное собрание Палеонтологической ассоциации. Цюрих, Швейцария.
- ^ Jump up to: а б Ваннье, Жан; Лю, Цзянни; Лерози-Обрил, Руди; Винтер, Якоб; Дейли, Эллисон К. (2 мая 2014 г.). «Сложная пищеварительная система у ранних членистоногих» . Природные коммуникации . 5 (1): 3641. Бибкод : 2014NatCo...5.3641V . дои : 10.1038/ncomms4641 . ISSN 2041-1723 . ПМИД 24785191 .
- ^ Jump up to: а б с Уиттингтон, Гарри Блэкмор; Бриггс, Дерек Эрнест Гилмор (14 мая 1985 г.). «Самое крупное кембрийское животное, Anomalocaris , Берджесс-Шейл, Британская Колумбия» . Философские труды Лондонского королевского общества. Б. Биологические науки . 309 (1141): 569–609. Бибкод : 1985RSPTB.309..569W . дои : 10.1098/rstb.1985.0096 .
- ^ Усами, Ёсиюки (7 января 2006 г.). «Теоретическое исследование формы тела и схемы плавания Anomalocaris на основе гидродинамического моделирования». Журнал теоретической биологии . 238 (1): 11–17. Бибкод : 2006JThBi.238...11U . дои : 10.1016/j.jtbi.2005.05.008 . ISSN 0022-5193 . ПМИД 16002096 .
- ^ Jump up to: а б с Кюль, Габриэле; Бриггс, Дерек Э.Г.; Раст, Джес (6 февраля 2009 г.). «Членистоногие с большим придатком и радиальной пастью из сланца Хунсрюк нижнего девона, Германия». Наука . 323 (5915): 771–773. Бибкод : 2009Sci...323..771K . дои : 10.1126/science.1166586 . ISSN 0036-8075 . ПМИД 19197061 . S2CID 47555807 .
- ^ Шеппард, Калифорния; Соперник, Делавэр; Карон, Ж.-Б. (1 октября 2018 г.). «К гидродинамике хвостового плавника Anomalocaris» . Интегративная и сравнительная биология . 58 (4): 703–711. дои : 10.1093/icb/icy014 . HDL : 1974/22737 . ISSN 1540-7063 . ПМИД 29697774 .
- ^ Дейли, Эллисон; Драге, Харриет (1 сентября 2015 г.). «Ископаемая летопись шелушения и тенденции линьки трилобитов» . Строение и развитие членистоногих . 45 (2): 71–96. дои : 10.1016/j.asd.2015.09.004 . ПМИД 26431634 .
- ^ Jump up to: а б с д Де Виво, Джачинто; Лаутеншлагер, Стефан; Винтер, Якоб (16 декабря 2016 г.). «Реконструкция ловкости пищевых придатков аномалокаридид проливает свет на экологию радиодонтана» .
- ^ Jump up to: а б с д и ж г час я дж к Мойсюк, Иосиф; Карон, Жан-Бернар (2021). «Исключительная многофункциональность в питающем аппарате среднекембрийского радиодонта» . Палеобиология . 47 (4): 704–724. Бибкод : 2021Pbio...47..704M . дои : 10.1017/pab.2021.19 . ISSN 0094-8373 . S2CID 236552819 .
- ^ Jump up to: а б с д и ж г час я дж Карон, Ж.-Б.; Мойсюк, Ю. (2021). «Гигантский нектобентосный радиодонт из сланцев Берджесс и значение разнообразия панцирей хурдиид» . Королевское общество открытой науки . 8 (9): 210664. Бибкод : 2021RSOS....810664C . дои : 10.1098/rsos.210664 . ПМЦ 8424305 . ПМИД 34527273 .
- ^ Jump up to: а б с Пейтс, С.; Дейли, AC (2017). « Кариосинтрипс : радиодонтан из кембрия Испании, США и Канады» . Статьи по палеонтологии . 3 (3): 461–470. дои : 10.1002/spp2.1084 . ISSN 2056-2802 . S2CID 135026011 .
- ^ Jump up to: а б с д и ж г час Ортега-Эрнандес, Хавьер (декабрь 2014 г.). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры Кембриджского философского общества . 91 (1): 255–273. дои : 10.1111/brv.12168 . ISSN 1469-185Х . ПМИД 25528950 . S2CID 7751936 .
- ^ Jump up to: а б с д и ж г час Пейтс, Стивен; Вулф, Джоанна М.; Лерози-Обрил, Руди; Дейли, Эллисон С.; Ортега-Эрнандес, Хавьер (9 февраля 2022 г.). «Новый опабиниид разнообразит самые странные чудеса стволовой группы эуартропод» . Труды Королевского общества B: Биологические науки . 289 (1968): 20212093. doi : 10.1098/rspb.2021.2093 . ПМЦ 8826304 . ПМИД 35135344 .
- ^ Эджкомб, Грегори Д. (1 марта 2010 г.). «Филогения членистоногих: обзор с точки зрения морфологии, молекулярных данных и летописи окаменелостей» . Строение и развитие членистоногих . Ископаемая летопись и филогения членистоногих. 39 (2): 74–87. дои : 10.1016/j.asd.2009.10.002 . ISSN 1467-8039 . ПМИД 19854297 .
- ^ Легг, Дэвид; Саттон, Марк; Эджкомб, Грегори; Карон, Жан-Бернар (10 октября 2012 г.). «Кембрийские двустворчатые членистоногие раскрывают происхождение артродизации» . Слушания. Биологические науки . 279 (1748): 4699–4704. дои : 10.1098/рспб.2012.1958 . ПМЦ 3497099 . ПМИД 23055069 .
- ^ Jump up to: а б Легг, Дэвид (2013). «Многочлениковые членистоногие из среднего кембрия Британской Колумбии (Канада)» . Журнал палеонтологии . 87 (3): 493–501. дои : 10.1666/12-112.1 . ISSN 0022-3360 . S2CID 86725173 .
- ^ Легг, Дэвид; Саттон, Марк; Эджкомб, Грегори (30 сентября 2013 г.). «Данные об окаменелостях членистоногих повышают соответствие морфологической и молекулярной филогении» . Природные коммуникации . 4 : 2485. Бибкод : 2013NatCo...4.2485L . дои : 10.1038/ncomms3485 . ПМИД 24077329 .
- ^ Легг, Дэвид А.; Ваннье, Жан (2013). «Сродство космополитических членистоногих Isoxys и его значение для происхождения членистоногих» . Летайя . 46 (4): 540–550. дои : 10.1111/лет.12032 . ISSN 1502-3931 .
- ^ О'Флинн, Роберт Дж.; Уильямс, Марк; Ю, Мэнсяо; Харви, Томас HP; Лю, Ю (2022). «Новый эуартропод с крупными лобными придатками из биоты Чэнцзян раннего кембрия» . Электронная палеонтология . 25 (1): 1–21. дои : 10.26879/1167 . ISSN 1094-8074 . S2CID 246779634 .
- ^ Jump up to: а б с Фу, Дунцзин; Легг, Дэвид А.; Дейли, Эллисон С.; Бадд, Грэм Э.; Ву, Ю; Чжан, Синлян (28 марта 2022 г.). «Эволюция двуветвистых придатков, выявленная кембрийским членистоногим с панцирем» . Философские труды Королевского общества B: Биологические науки . 377 ): 20210034.doi : ( 1847 10.1098/rstb.2021.0034 . ПМЦ 8819368 . ПМИД 35125000 . S2CID 246608509 .
- ^ Ортега-Эрнандес, Хавьер (15 июня 2015 г.). «Гомология головных склеритов у членистоногих сланцев Берджесс» . Современная биология . 25 (12): 1625–1631. дои : 10.1016/j.cub.2015.04.034 . ISSN 0960-9822 . ПМИД 25959966 .
- ^ Jump up to: а б Гирибет, Гонсало; Эджкомб, Грегори Д. (17 июня 2019 г.). «Филогения и эволюционная история членистоногих» . Современная биология . 29 (12): Р592–Р602. дои : 10.1016/j.cub.2019.04.057 . ISSN 0960-9822 . ПМИД 31211983 . S2CID 189926344 .
- ^ Jump up to: а б Эджкомб, Грегори Д. (2 ноября 2020 г.). «Происхождение членистоногих: объединение палеонтологических и молекулярных данных» . Ежегодный обзор экологии, эволюции и систематики . 51 (1): 1–25. doi : 10.1146/annurev-ecolsys-011720-124437 . ISSN 1543-592X . S2CID 225478171 .
- ^ Jump up to: а б Чжанхэ, Чжань ; Сянгуан, Хоу; Ян Цзяю , Ин Цзунцзе , 10.1.1.693.5869
- ^ Бадд, Грэм Э. (1998). «Морфология и филогенетическое значение Kerygmachela kierkegaardi Budd (формация Буэн, нижний кембрий, Северная Гренландия)» . Труды Королевского общества Эдинбурга по наукам о Земле и окружающей среде . 89 (4): 249–290. дои : 10.1017/S0263593300002418 . ISSN 1473-7116 . S2CID 85645934 .
- ^ Янг, Флетчер Дж.; Винтер, Якоб (2017). «Онихофороподобная миоанатомия кембрийского жаберного лобопода Pambdelurion Whittingtoni» . Палеонтология . 60 (1): 27–54. Бибкод : 2017Palgy..60...27Y . дои : 10.1111/пала.12269 . hdl : 1983/92180ef0-2205-4c65-9a70-90d59cfea2f4 . ISSN 1475-4983 . S2CID 55477207 .
- ^ Эджкомб, Грегори Д. (2010). «Филогения членистоногих: обзор с точки зрения морфологии, молекулярных данных и летописи окаменелостей» . Строение и развитие членистоногих . 39 (2–3): 74–87. дои : 10.1016/j.asd.2009.10.002 . ISSN 1467-8039 . ПМИД 19854297 .
- ^ Дейли, Эллисон С.; Антклифф, Джонатан Б.; Дрейдж, Харриет Б.; Пейтс, Стивен (22 мая 2018 г.). «Ранние окаменелости Euarthropoda и кембрийский взрыв» . Труды Национальной академии наук . 115 (21): 5323–5331. Бибкод : 2018PNAS..115.5323D . дои : 10.1073/pnas.1719962115 . ISSN 0027-8424 . ПМК 6003487 . ПМИД 29784780 .
- ^ Сиань-Гуан, Хоу; Бергстрем, Ян; Альберг, Пер (сентябрь 1995 г.). «Аномалокарис и другие крупные животные фауны Чэнцзян нижнего кембрия на юго-западе Китая» . ГФФ . 117 (3): 163–183. дои : 10.1080/11035899509546213 . ISSN 1103-5897 .
- ^ Хауг, Иоахим Т.; Валошек, Дитер; Маас, Андреас; Лю, Ю; Хауг, Кэролин (март 2012 г.). «Функциональная морфология, онтогенез и эволюция креветкообразных хищников-богомолов в кембрии: КРЕВЕТООБОДНЫЕ БОГОМОЛА КЕМБРИЙСКИЕ ХИЩНИКИ» . Палеонтология . 55 (2): 369–399. дои : 10.1111/j.1475-4983.2011.01124.x . S2CID 82841481 .
- ^ Смит, Мартин Р.; Карон, Жан-Бернар (июнь 2015 г.). «Голова галлюцигении и глоточный каркас ранних экдизозойных» (PDF) . Природа . 523 (7558): 75–78. Бибкод : 2015Natur.523...75S . дои : 10.1038/nature14573 . ISSN 1476-4687 . ПМИД 26106857 . S2CID 205244325 .
- ^ Танака, Генго; Хоу, Сянгуан; Ма, Сяоя; Эджкомб, Грегори Д.; Штраусфельд, Николас Дж. (октябрь 2013 г.). «Хелицератный рисунок нервного поля у кембрийского членистоногого с большим придатком». Природа . 502 (7471): 364–367. Бибкод : 2013Natur.502..364T . дои : 10.1038/nature12520 . ISSN 1476-4687 . ПМИД 24132294 . S2CID 4456458 .
- ^ Ортега-Эрнандес, Хавьер; Лерози-Обрил, Руди; Пейтс, Стивен (18 декабря 2019 г.). «Склонность к сохранению нервной системы в кембрийских отложениях типа сланцев Бёрджесс» . Труды Королевского общества B: Биологические науки . 286 (1917): 20192370. doi : 10.1098/rspb.2019.2370 . ПМК 6939931 . ПМИД 31822253 .
- ^ Jump up to: а б с д и ж МакКолл, Кристиан Р.А. (13 декабря 2023 г.). «Большой пелагический лобопод из кембрийских сланцев Пиоче в Неваде» . Журнал палеонтологии : 1–16. дои : 10.1017/jpa.2023.63 . ISSN 0022-3360 .
- ^ Jump up to: а б Цзэн, Хан; Чжао, Фанчэн; Инь, Цзунцзюнь; Чжу, Маоянь (2 января 2018 г.). «Морфология разнообразных склеритов головы радиодонтана из раннего кембрия Чэнцзян Лагерштетте, юго-запад Китая» . Журнал систематической палеонтологии . 16 (1): 1–37. дои : 10.1080/14772019.2016.1263685 . ISSN 1477-2019 . S2CID 133549817 .
- ^ Jump up to: а б Юаньюань, Ван; Хуан, Диин; Ху, Шисюэ (1 ноября 2013 г.). «Новые лобные придатки аномалокардид из биоты Гуаньшань, восточная часть Юньнани» . Китайский научный бюллетень . 58 (32): 3937–3942. Бибкод : 2013ЧСБу..58.3937В . дои : 10.1007/s11434-013-5908-x . S2CID 83910779 .
- ^ Jump up to: а б с Цзяо, Дэ-гуан; Пейтс, Стивен; Лерози-Обрил, Руди; Ортега-Эрнандес, Хавьер; Ян, Цзе; Лан, Тиан; Чжан, Си-гуан (2021). «Эндемичные радиодонты кембрийской биоты Гуаньшаня 4-го этапа Южного Китая» . Acta Palaeontologica Polonica . 66 . дои : 10.4202/app.00870.2020 . ISSN 0567-7920 . S2CID 236683029 .
- ^ Jump up to: а б Го, Дж.; Пейтс, С.; Конг, П.; Дейли, AC; Эджкомб, Джорджия; Чен, Т.; Хоу, X. (2018). «Новый лобный придаток радиодонта (ствол Euarthropoda) с мозаикой признаков кембрийской биоты (серия 2 этап 3) Чэнцзян» . Статьи по палеонтологии . 5 (1). ISSN 2056-2799 .
- ^ Jump up to: а б с Ву, Ю; Фу, Дунцзин; Ма, Цзясинь; Линь, Вэйлян; Солнце, АО; Чжан, Синлян (1 июня 2021 г.). « Houcaris gen. nov. из раннего кембрия (этап 3) Chengjiang Lagerstätte расширил палеогеографическое распространение тамизиокаридид (Panarthropoda: Radiodonta)» . ПалЗ . 95 (2): 209–221. дои : 10.1007/s12542-020-00545-4 . ISSN 1867-6812 . S2CID 235221043 .
- ^ Jump up to: а б с д Цзэн, Хан; Чжао, Фанчэн; Чжу, Маоянь (7 сентября 2022 г.). « Innovatiocaris , полный радиодонт из раннего кембрия Chengjiang Lagerstätte и его значение для филогении Radiodonta» . Журнал Геологического общества . 180 . дои : 10.1144/jgs2021-164 . ISSN 0016-7649 . S2CID 252147346 .
- ^ Jump up to: а б с Ву, Ю; Пейтс, Стивен; Лю, Конг; Чжан, Минцзин; Линь, Вэйлян; Ма, Цзясинь; Ву, Юхэн; Чай, Шу; Чжан, Синлян; Фу, Дунцзин (31 декабря 2024 г.). «Новый радиодонт из нижнего кембрия (серия 2, стадия 3) Чэнцзян Лагерштетте, Южный Китай, сообщает об эволюции структур питания у радиодонтов» . Журнал систематической палеонтологии . 22 (1). дои : 10.1080/14772019.2024.2364887 . ISSN 1477-2019 .
- ^ Jump up to: а б Чжан, Минцзин; Ву, Ю; Линь, Вэйлян; Ма, Цзясинь; Ву, Юхэн; Фу, Дунцзин (11 апреля 2023 г.). «Amplectobeluid Radiodont Guanshancaris gen. nov. из нижнего кембрия (этап 4) Гуаньшань Лагерштетте в Южном Китае: биостратиграфические и палеобиогеографические последствия» . Биология . 12 (4): 583. doi : 10.3390/biology12040583 . ISSN 2079-7737 . ПМЦ 10136193 . ПМИД 37106783 .
- ^ Jump up to: а б Потин, Гаэтан Ж.-М.; Герио, Пьер; Дейли, Эллисон К. (2023). «Лобные придатки радиодонтов из биоты Фезуата (Марокко) демонстрируют большое разнообразие и экологическую адаптацию к питанию взвесью в раннем ордовике» . Границы экологии и эволюции . 11 . дои : 10.3389/fevo.2023.1214109 . ISSN 2296-701X .
- ^ Ортега-Эрнандес, Хавьер (декабрь 2014 г.). «Осмысление «нижней» и «верхней» стеблевой группы Euarthropoda с комментариями о строгом использовании названия Arthropoda von Siebold, 1848». Биологические обзоры Кембриджского философского общества . 91 (1): 255–273. дои : 10.1111/brv.12168 . ISSN 1469-185Х . ПМИД 25528950 . S2CID 7751936 .
- ^ Чжу, X.; Лерози-Эйприл, Р.; Ортега-Эрнандес, Дж. (2021). «Фуронгские (цзяншаньские) находки радиодонтов в Польше и Южном Китае и летопись окаменелостей Hurdiidae» . ПерДж . 9 . е11800. дои : 10.7717/peerj.11800 . ПМЦ 8312493 . ПМИД 34386302 .
- ^ Потин, Гаэтан Ж.-М.; Дейли, Эллисон К. (2023). «Значение Anomalocaris и других Radiodonta для понимания палеоэкологии и эволюции во время кембрийского взрыва» . Границы в науках о Земле . 11 . Бибкод : 2023FrEaS..1160285P . дои : 10.3389/feart.2023.1160285 . ISSN 2296-6463 .
- ^ Jump up to: а б Пейтс, Стивен; Дейли, Эллисон; Ортега-Эрнандес, Хавьер (3 марта 2018 г.). Ответ на комментарий к записи «Aysheaia prolata из формации Юта Уилер (Драмиан, кембрий) представляет собой лобный придаток радиодонтана Stanleycaris» с формальным описанием Stanleycaris .
- ^ Jump up to: а б с Сунь, Чжисинь; Цзэн, Хан; Чжао, Фанчэн (1 августа 2020 г.). «Новый радиодонт среднего кембрия из Северного Китая: последствия морфологического неравенства и пространственного распределения хурдиид» . Палеогеография, Палеоклиматология, Палеоэкология . 558 : 109947. Бибкод : 2020PPP...558j9947S . дои : 10.1016/j.palaeo.2020.109947 . ISSN 0031-0182 . S2CID 224868404 .
- ^ Jump up to: а б с д Пейтс, Стивен; Лерози-Эйприл, Руди; Дейли, Эллисон С.; Кир, Чарльз; Бонино, Генри; Ортега-Эрнандес, Хавьер (19 января 2021 г.). «Разнообразная фауна радиодонтов из формации Марджум в штате Юта, США (кембрий: драмиан) » ПерДж . 9 : е10509. дои : 10.7717/peerj.10509 . ISSN 2167-8359 . ПМЦ 7821760 . ПМИД 33552709 .
- ^ Jump up to: а б с д Уайтэйвс, Дж. Ф. (1892). «Описание нового рода и вида филлокарид ракообразных из среднего кембрия горы Стивен, Британская Колумбия». Канадский рекорд науки . 5 (4).
- ^ Jump up to: а б с д Уолкотт, CD 1911a. Голотурии и медузы среднего кембрия . Кембрийская геология и палеонтология II. Разные коллекции Смитсоновского института, 57: 41–68.
- ^ Дейли, Эллисон; Легг, Дэвид (2 июля 2015 г.). «Морфологическая и таксономическая оценка древнейшего аномалокаридида из нижнего кембрия Польши» . Геологический журнал . -1 (5): 949–955. Бибкод : 2015ГеоМ..152..949Д . дои : 10.1017/S0016756815000412 . S2CID 130745134 .
- ^ Jump up to: а б с УОЛКОТТ, CD 1912. Среднекембрийские Branchiopoda, Malacostraca, Trilobita и Merostomata . Разные коллекции Смитсоновского института, 57: 145–228.
- ^ РОЛЬФ, WDI 1962. Два новых панциря членистоногих из сланцев Бёрджесс (средний кембрий) в Канаде. Музей сравнительной зоологии Бревиоры, 60: 1–9.
- ^ Конвей Моррис, С. (1978). «Лаггания камбрия Уолкотт: сложное ископаемое». Журнал палеонтологии . 52 (1): 126–131. JSTOR 1303799 .
- ^ Бриггс, DEG (1979). «Аномалокарис, крупнейшее известное кембрийское членистоногое». Палеонтология . 22 (3): 631–664.
- ^ Дейли, Эллисон; Бадд, Грэм; Карон, Жан-Бернар (1 октября 2013 г.). «Морфология и систематика аномалокарида Hurdia из среднего кембрия Британской Колумбии и Юты» . Журнал систематической палеонтологии . 11 . дои : 10.1080/14772019.2012.732723 . S2CID 86465719 .
- ^ Пейтс, Стивен; Дейли, Эллисон К. (2019). «Формация Кинзерс (Пенсильвания, США): наиболее разнообразный комплекс радиодонтов кембрийского этапа 4» . Геологический журнал . 156 (7): 1233–1246. Бибкод : 2019GeoM..156.1233P . дои : 10.1017/S0016756818000547 . S2CID 134299859 .
- ^ Патерсон, Джон Р.; Эджкомб, Грегори Д.; Гарсиа-Беллидо, Диего К. (2020). «Несопоставимые сложные глаза кембрийских радиодонтов свидетельствуют об их способе развития и разнообразной визуальной экологии» . Достижения науки . 6 (49). Бибкод : 2020SciA....6.6721P . дои : 10.1126/sciadv.abc6721 . hdl : 10141/622906 . ПМЦ 7821881 . ПМИД 33268353 .
