Бразилодон
| Бразилодон Временной диапазон: поздний триас. ~ | |
|---|---|
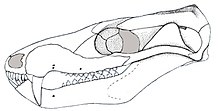 | |
| Реконструированный череп, вид сбоку. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | Терапсида |
| Клэйд : | Цинодонтия |
| Клэйд : | Млекопитающие |
| Семья: | † Бразилодонтиды Бонапарт и др. , 2005 г. |
| Род: | † Бразилодон Бонапарт и др. , 2003 г. |
| Разновидность: | † Б. четырехугольная |
| Биномиальное имя | |
| † Бразилодон четырехугольный Бонапарт и др. , 2003 г. | |
| Синонимы | |
| |
Brasilodon («зуб из Бразилии») — вымерший род небольших млекопитающих цинодонтов , которые жили на территории современной Бразилии в норийский период позднего триаса , около 225,42 миллиона лет назад. Хотя полных скелетов обнаружено не было, длина бразилодона оценивается в 12 сантиметров (4,7 дюйма). Его зубной ряд показывает, что он, скорее всего, был насекомоядным . Род монотипический , содержит только вид B. Quadrangularis . Бразилодон принадлежит к семейству Brasilodontidae , представители которого были одними из ближайших родственников млекопитающих, единственных ныне живущих цинодонтов. Два других рода бразилодонтид, и Minicynodon , теперь считаются младшими синонимами Brasilodon Brasilitherium .
Открытие и присвоение имени
[ редактировать ]
Первые три экземпляра, относящиеся к Brasilodon Quadrangularis, были найдены на стоянке Линья-Сан-Луис, карьере недалеко от города Фашинал-ду-Сотурно в штате Риу-Гранди-ду-Сул . Породы, где был найден Бразилиодон , относятся к верхней части толщи Канделария суперпоследовательности Санта-Мария, соответствующей традиционной формации Катуррита , датируемой ранним норийским возрастом позднего триаса . [1] [2] [3] Голотип послеклыковые (UFRGS-PV-0611-T) представляет собой хорошо сохранившийся череп, на котором сохранились как левый, так и правый верхние зубы , но отсутствует нижняя челюсть. Упомянутый экземпляр UFRGS-PV-0716-T состоит из левой передней части черепа с сохранением 6 постклыков. Экземпляр UFRGS-PV-0628-T состоит из части черепа, включая нижнюю челюсть, с сохраненной большей частью верхнего и нижнего зубного ряда. [1] множество других экземпляров Brasilodon С тех пор было обнаружено как в Факсинале-ду-Сотурно, так и в Канделарии , содержащих как краниальный, так и посткраниальный материал. [1] [2]
Род Brasilodon был назван в статье 2003 года Хосе Ф. Бонапарта , Агустина Г. Мартинелли, Сезара Л. Шульца и Рожерио Руберта. Родовое название Brasilodon происходит от названия страны Бразилии, где он был найден, и от греческого слова odon , что означает «зуб». Предполагаемое значение - «зуб из Бразилии». Видовой эпитет Quadrangularis относится к прямоугольной форме верхних послеклыковых зубов . [1]
вид Brasilitherium riograndensis В той же статье 2003 года по шести экземплярам был назван . Экземпляры, отнесенные к Brasilitherium, были обнаружены в тех же местах, что и Brasilitherium , и отличались от этого таксона в основном меньшими размерами, другими пропорциями черепа и наличием бугорка d в нижних постклыках Brasilitherium , но не у Brasilodon. . [1] [4] В статье 2005 года семейство Brasilodontidae было выделено для двух родов. [4] В 2010 году третий вид бразилодонтид, Minicynodon maieri , был назван Бонапартом и др. , основанный на единственном хорошо сохранившемся черепе из Факсинал-ду-Сотурно. Этот вид отличался от Brasilodon прочным прикреплением костей крыши черепа , а от Brasilitherium - отсутствием бугорка b в нижних постклыках. Миницинодон также интерпретировался как обладающий двойным челюстным суставом, в отличие от двух других видов. [5] Более поздние исследования поставили под сомнение достоверность Brasilitherium и Minicynodon , предположив вместо этого, что они, наряду с Brasilodon , просто представляют собой индивидуальные вариации внутри одного вида. В этом случае Brasilodon является действительным таксоном, тогда как Brasilitherium и Minicynodon являются недействительными младшими синонимами . [2] [6]
Описание
[ редактировать ]Бразилодон был небольшим производным цинодонтом, предполагаемая общая длина которого составляла около 12 сантиметров (4,7 дюйма). [7] Он демонстрировал многие черты млекопитающих, в том числе хорошо развитое вторичное небо , симметричное развитие зубов и более развитую анатомию ушей, чем у более ранних цинодонтов. [4]
Череп
[ редактировать ]

Длина черепа разных экземпляров бразилодона сильно различалась: от 20 до 55 миллиметров (от 0,79 до 2,17 дюйма), главным образом из-за различий в возрасте. [7] Череп имел низкую, вытянутую форму, если смотреть сбоку. Верхняя часть черепной коробки была довольно широкой, с коротким и низким теменным гребнем . [1] У бразилодона отсутствовали префронтальные и посторбитальные кости, которые были обнаружены у более базальных пробайногнатов, таких как прозостродон . не было заглазничной перемычки За глазницей . Скуловая дуга (скула) была довольно низкой и тонкой. [2] [4]
Нижняя челюсть состояла в основном из зубной кости , несущей зубы. Кончик нижней челюсти был загнут вверх. Симфиз прикрепления , соединение между двумя половинками зубной кости, не сросся, с шероховатой поверхностью в месте . связок Меккелева борозда располагалась вблизи нижнего края зубной кости. Постдентарные кости, набор костей нижней челюсти, расположенный позади зубной кости, были сильно редуцированы по сравнению с состоянием более примитивных цинодонтов. Как и у большинства цинодонтов, не относящихся к млекопитающим, челюстной сустав бразилодона включал квадратную кость (кость черепа, гомологичную наковальне современных млекопитающих) и суставную кость (одну из постзубных костей, гомологичную молоточку ). [4] Также мог иметь место контакт между зубной и чешуйчатой костью , при этом суставной отросток зубной кости имел утолщенный конец, что, по-видимому, является предшественником полностью развитого мыщелка зубной кости у более производных млекопитающих . Квадратная кость имела хорошо развитый стременный отросток, подобный таковому у базальных млекопитающих Morganucodon . [4]
Передняя часть неба состояла из хорошо развитого вторичного неба , образованного верхнечелюстными костями и небными костями , которое простиралось примерно до последнего постклыка. [6] На вторичном небе была бороздка, в которую могли входить нижние постклыки, когда рот был закрыт. Позади вторичного неба располагалось первичное небо , образованное сошником и крыловидными костями . Между крыловидными костями существовала пара промежутков, называемых межкрыловидными впадинами . Хорошо развитые межкрыловидные пустоты известны у базальных цинодонтов, таких как Procynosuchus , но у более продвинутых групп они обычно уменьшены или отсутствуют, поэтому их присутствие у Brasilodon , вероятно, является производным заболеванием. Крыловидные отростки имели поперечный контакт с базиптеригоидным отростком базисфеноидальной кости . [4] Базикраний был широким , если смотреть снизу. Проотическая . и опистотическая кости были слиты в каменистую кость , имевшую четко мысок выраженный [2] Имелись отдельные отверстия (отверстия) для верхнечелюстного и нижнечелюстного нервов. Рядом с crista interfenestralis находилось большое углубление, называемое «стременной выемкой», в котором было несколько отверстий. Затылочные мыщелки располагались дальше ламбдовидного гребня . [4]
Краниальная эндокаст , внутренняя поверхность черепной коробки, является важным источником информации о мозге доисторических животных, поскольку сам мозг очень редко сохраняется в окаменелостях. Исследование 2013 года, проведенное Родригесом и соавт. описал эндокаст экземпляра UFRGS-PV-1043-T, который первоначально был отнесен к роду Brasilitherium . Эндокаст имел длину 17,67 миллиметров (0,696 дюйма), что составляет 46,5% от общей длины этого черепа. Максимальная ширина эндокаста составляла 7,17 миллиметра (0,282 дюйма). Форма эндокаста указывает на то, что у животного были хорошо развиты обонятельные луковицы — части мозга, связанные с обонянием. С другой стороны, полушария головного мозга были меньше, чем у базальных млекопитающих, таких как Morganucodon . Неясно, ли бразилодон обладал неокортексом — частью мозга, обнаруженной у современных млекопитающих. Было интерпретировано, что коэффициент энцефализации (относительный размер мозга), вероятно, выше, чем у более базальных цинодонтов, таких как traversodontid Massetognathus , но ниже, чем у базальных млекопитающих и современных млекопитающих. Как и другие цинодонты, не относящиеся к млекопитающим, бразилодон, вероятно, имел лиссэнцефалическую (гладкую) поверхность мозга. [8]
Зубной ряд
[ редактировать ]
Как и большинство цинодонтов, бразилодон обладал гетеродонтным зубным рядом, разделенным на резцы , клыки и постклыки. Экземпляры, первоначально отнесенные к Brasilodon, не сохранили ни одного верхнего резца, но некоторые экземпляры, отнесенные к Brasilitherium, имели 4 пары резцов в верхней челюсти. [9] Было 3 пары нижних резцов; первый нижний резец был вытянутым (направленным вперед). [2] У взрослых особей клыки были большими и сплюснутыми из стороны в сторону, но не имели зазубрин . [4] У мелких особей, отнесенных к Brasilitherium , было две пары функциональных клыков, которые лишь немногим превышали размеры резцов. [9]
В обеих челюстях насчитывалось до 8 пар постклыков. [4] Постклыки имели прямоугольную форму, если смотреть в сторону темени . Форма верхних постклыков была незначительной, тогда как нижние имели больше вариаций. [1] Верхние постклыки были значительно меньше нижних, что наблюдалось у многих других пробаиногнатов. [10] Пост-клыки были «триконодонтами». [2] с тремя основными выступами, расположенными примерно по прямой линии. Был большой центральный бугор, называемый бугорком А на верхних зубах и бугором А на нижних, бугорок меньшего размера перед центральным бугорком (бугор B/b) и еще один небольшой бугор позади центрального бугра (бугор С/b). в). [1] В нижних постклыках некоторых экземпляров позади бугорка с имелся дополнительный бугорок (бугорок d); отсутствие или наличие этого выступа использовалось, чтобы отличить бразилодона от бразилитериума . [1] но вполне вероятно, что отсутствие острия d у некоторых экземпляров вызвано стираемостью зубов у пожилых особей. [6] Постклыки также имели различное количество дополнительных бугорков, иногда образующих поясную извилину . У более старых особей между клыками и постклыками был большой зазор ( диастема ), который образовался из-за того, что первый постклык был выброшен, но не заменен. В средних нижних задних клыках существовала система «язычок-паз» для блокировки между соседними зубами, с бугорком d, соединяющим бугор b и дополнительный бугор е, но эта система, по-видимому, не функционировала из-за расстояния между зубами. Корни постклыков не были полностью разделены, как у современных млекопитающих, но имели заметное сужение посередине, что придавало им восьмеричное поперечное сечение. [11] корней Питательные каналы тоже были разделены. [2] Постклыковые корни были прикреплены к челюсти кольцом окостеневшей периодонтальной связки . [11]
Позвонки и ребра
[ редактировать ]
Позвонки мало бразилодона . изучены В одном экземпляре (UFRGS-PV-1043-T) сохранилось 4 изолированных пресакральных позвонка, все из которых плохо сохранились. Центры позвонков были платицелозными , с плоским передним концом и вогнутым задним концом. Такая форма также наблюдается у некоторых тритилодонтид и базальных млекопитающих, но не у более базальных цинодонтов, у которых обычно были амфицельные позвонки, вогнутые с обеих сторон. Нервный канал был довольно большим, а боковые стенки тонкими. У этого экземпляра нервные дуги срослись с центрами, что указывает на то, что он взрослый. На одном из позвонков сохраняется небольшой цилиндрический диапофиз (поперечный отросток). Зигапофизы . (суставные отростки, четыре отростка каждого позвонка, соединяющие между собой соседние позвонки) сохранились плохо, но постзигапофизы, по-видимому, были почти горизонтальными и располагались довольно высоко на позвонках Форма основания нервного отростка (заостренное расширение на верхней поверхности позвонка) указывает на то, что он был удлинен в переднезаднем направлении (в направлении вперед-назад). [2]
В экземпляре УФРГС-ПВ-1043-Т также сохранились 4 изолированных фрагмента ребер . Ребра уплощены в передне-заднем направлении. Многие непробайногнатные цинодонты, включая большинство циногнатов , имели широкие выступы, известные как реберные пластины на ребрах ; эти пластинки отсутствуют у большинства пробаиногнатий, включая Brasilodon . [2]
Конечности и пояса конечностей
[ редактировать ]У экземпляра УФРГС-ПВ-1043-Т сохранилась нижняя часть правой лопатки (лопатка), а два других связанных с ней костных фрагмента, вероятно, представляют собой среднюю часть дорсального (верхнего) края и передний (передний) край. , соответственно. Лопатка представляла собой тонкую кость с суженным основанием, как и у других эпицинодонтов . Верхний край был несколько выпуклым, а заднедорсальный (верхний спинной) угол заострен. Передний и задний края имели выступающие вбок фланцы, как это наблюдается у других эпицинодонтов, не имеющих млекопитающих. имелся хорошо развитый акромиальный На конце переднего края отросток. Акромион был направлен вперед, как и у многих других цинодонтов. имелась глубокая подостная ямка Со стороны лопатки , а надостная ямка, по-видимому, отсутствовала. Гленоидная плечевой фасетка лопатки (область, где она соединялась с костью ) была вогнутой и имела несколько яйцевидную форму. Гленоид, по-видимому, был направлен вниз, в отличие от гленоидов, обращенных вбок у некоторых рано расходящихся цинодонтов. Коракоидная и прокоракоидная кости не сохранились, но на лопатке имелись суставные поверхности для этих костей. [2]

В образце UFRGS-PV-1043-T полностью сохранилась левая плечевая кость (кость плеча) длиной 15,6 мм (0,61 дюйма). Плечевая кость была тонкой. Она была несколько искривлена, с углом наклона 15 градусов между противоположными концами кости. Искривленная плечевая кость встречается у большинства цинодонтов за пределами Терии (клады, включающей современных сумчатых и плацентарных ), в том числе у современных однопроходных . Однако скручивание плечевой кости у бразилодона было относительно низким по сравнению с таковым у многих других нетерийских цинодонтов. На проксимальной стороне (стороне, соединяющейся с плечом) головка плечевой кости имела полусферическую форму, типичную для млекопитающих, но в отличие от более базальных цинодонтов, у которых головка плечевой кости в целом была более яйцевидной. В отличие от большинства териев головка плечевой кости обращена заднелатерально (назад и в сторону). шел гребень от головки плечевой кости до эктепимыщелкового гребня На другом конце кости . В отличие от более базальных цинодонтов, бразилодон имел отчетливую больший бугорок , хотя и был совсем маленьким. Малый бугорок имел толстую и луковицеобразную форму. Он был крупнее большого бугорка, как у базальных млекопитающих и однопроходных, но в отличие от многобугорковых , спалакотериид и териан. Дельтопекторальный гребень сливался с большим бугорком и занимал 48% длины плечевой кости. Гребень был довольно большим и крепким, как у других цинодонтов, не относящихся к млекопитающим, однопроходных и роющих (роющих) териев, таких как кроты . На другой стороне кости был еще один гребень, известный как гребень Терес . Этот гребень был обнаружен у большинства мезозойских цинодонтов, но отсутствовал у Винцелестов и териан. Тересный гребень был примерно такой же длины, как дельтопекторальный гребень, и простирался от меньшего бугорка до большого овоидного выступа около середины кости. Этот выступ, возможно, служил якорем для большой круглой мышцы . Подобная структура существовала у многих других мезозойских цинодонтов, но не у мультитуберкулят, спалакотериид или териан. На передней поверхности плечевой кости имелась широкая двуглавая борозда , окаймленная дельтопекторальным и круглым гребнями, аналогичная таковой у многих других мезозойских цинодонтов. Двуглавая борозда, вероятно, служила местом прикрепления клювовидно-плечевой мышцы . Дистальная сторона плечевой кости (сторона, соединяющаяся с лучевой и локтевой костью ) предплечья имела ширину 43% длины плечевой кости. Он был уже, чем у многих других цинодонтов, не являющихся млекопитающими, а также у однопроходных, но шире, чем у базальных млекопитающих Morganucodon и териан Didelphis . Энтепимыщелок был довольно крепким и имел выступ , направленный к проксимальному концу кости. Эктепимыщелок был более узким и менее выступающим , чем энтепимыщелок. Энтепикондилярное отверстие представляло собой большое и удлиненное отверстие. не было И наоборот, эктепикондилярного отверстия ; это отверстие было обнаружено у большинства цинодонтов более базально, чем у бразилодона , но отсутствовало у тритилодонтид и большинства млекопитающих. Локтевой . и радиальный мыщелки (части, соединяющиеся с локтевой и лучевой костью соответственно) имели луковицеобразную форму, как это наблюдается у других цинодонтов, не являющихся млекопитающими Мыщелки были разделены узкой бороздкой и не имели Трохлеарная (шкивообразная) форма, наблюдаемая у многих млекопитающих кроновой группы, таких как терианы или однопроходные стеблевые Kryoryctes . [12] Локтевой мыщелок был виден как на передней, так и на задней стороне плечевой кости. Его передняя сторона была несколько сжата в поперечном направлении. Лучевой мыщелок был больше локтевого, но не был виден на задней стороне кости. Ямка локтевого отростка (впадина на задней стороне плечевой кости) была довольно мелкой, в отличие от более глубокой ямки, наблюдаемой у мультитуберкулезных, винцелестов и териан. [2]

Длина левого радиуса UFRGS-PV-1043-T составляет 14 миллиметров (0,55 дюйма). Лучевая кость была тонкой, с круглым поперечным сечением вблизи проксимального конца (ближе к локтю). Головка лучевой кости (там, где она сочленялась с плечевой костью) имела примерно круглую форму, с чашеобразным углублением посередине, окруженным выпуклым краем. Голова была несколько наклонена в переднемедиальную (переднюю правую) сторону. На задне-медиальной (задней правой) стороне головы имелась небольшая суставная фасетка в месте сочленения с лучевой вырезкой локтевой кости. мышцы не было Лучевой бугристости для прикрепления двуглавой . Дистальный конец кости (ближе к запястью) имел форму приплюснутого прямоугольника. Дистальный конец был несколько изогнут задне-медиально, что позволяло кости пересекать переднюю часть локтевой кости, как у тритилодонтид, морганукодона и опоссумов. Дистальная суставная поверхность (в месте соединения с запястьем) также имела прямоугольную форму. [2]
Правая локтевая кость UFRGS-PV-1043-T имеет длину 16,5 миллиметров (0,65 дюйма). Как и у большинства цинодонтов, локтевая кость была узкой медиолатерально (из стороны в сторону) и имела сигмовидную (s-образную) форму. По бокам кости имелись две бороздки, идущие от одного конца кости к другому, окруженные с передней и задней стороны утолщенными краями. Считается, что бороздки служили точками прикрепления мышц-разгибателей и сгибателей. На проксимальном конце кости имелся хорошо развитый и окостеневший локтевой отросток. За исключением Trucidocynodon и tritylodontids, ни один цинодонт, более базальный, чем Brasilodon, не имел окостеневшего локтевого отростка, и считается, что он был хрящевым у этих таксонов . Олекранон бразилодона составлял чуть менее 20% длины локтевой кости и был направлен вперед относительно остальной части кости. Олекранон был несколько толще из стороны в сторону, чем остальная часть кости. Полулунная вырезка , в месте сочленения локтевой кости с локтевым мыщелком плечевой кости, была большой и полукруглой. локтевой отросток был довольно небольшим, как у большинства базальных цинодонтов и докодонтной млекопитающей формы Haldanodon , но в отличие от многих других млекопитающих, у которых локтевой отросток образует заметный гребень. [2]

Таз , (тазовая кость) UFRGS-PV-1043-T сохраняет полную левую вертлужную впадину и лобковую кость а также неполную левую подвздошную и седалищную кости . Вертлужная впадина – это место, где бедренная кость задней конечности сочленяется с тазом. Вертлужная впадина бразилодона представляла собой глубокую сферическую впадину, обращенную вбок. Три кости, составляющие вертлужную впадину (лобковая, подвздошная и седалищная), были слиты вместе, швов не было видно; это особенность, которую бразилодон разделял с млекопитающими, в то время как кости были более отчетливыми у некоторых более базальных цинодонтов, таких как траверсодонтиды . Кости образовывали три отдельных гребня, известных как надвертлужные опоры вокруг вертлужной впадины. Контрфорсы были разделены промежутками в 2–3 миллиметра (0,079–0,118 дюйма), в отличие от современных териан, у которых вокруг вертлужной впадины имеется полностью окостеневший край. промежутки, вероятно, были заполнены волокнистым хрящом У живого животного . Лопатка подвздошной кости сохранилась плохо, но форма основания указывает на то, что постацебулярная (направленная назад) часть лопатки либо редуцирована, либо отсутствовала. Бразилодон , как и другие прозостродонты . Лобковый бугр располагался ниже вертлужной впадины, как и у большинства пробайногнатов, включая современных териев. У базальных эпицинодонтов, однопроходных и спалакотериид Akidolestes лобковая бугристость вместо этого располагалась перед вертлужной впадиной. Лобковая и седалищная кость образовывали большое овоидное запирательное отверстие . [2]

Правое бедро (бедренная кость) UFRGS-PV-1043-T имеет длину 15,9 мм (0,63 дюйма). Тело бедренной кости было в основном прямым, но с заметным изгибом вперед вблизи тазобедренного сустава, как у других цинодонтов, не относящихся к млекопитающим; у современных млекопитающих этот изгиб развит менее развито. Проксимальная часть стержня (ближе к бедру) имела преимущественно квадратную форму, но дистально (к колену) становилась более сжатой спереди назад (к колену), одновременно становясь шире из стороны в сторону. Проксимальный и дистальный концы кости имели одинаковую ширину. На проксимальном конце кости головка бедренной кости (сочленяющаяся с вертлужной впадиной бедра) имела полусферическую форму. Голова была отделена от остальной части кости короткой бедренной шейкой , как у тритилодонтид и млекопитающих; у более примитивных цинодонтов не было шейки бедра. Голова была наклонена под углом около 60 градусов медиально (вправо) по отношению к продольной оси бедренной кости, что напоминало состояние у мультитуберкулезных и спалакотериид. В отличие от млекопитающих, здесь не было ямка головки на головке бедренной кости. Чуть дистальнее головки бедренной кости находился хорошо развитый выступ, известный как большой вертел на левой стороне кости . Отчетливый большой вертел также обнаружен у тритилодонтид и млекопитающих, но у большего количества базальных цинодонтов большой вертел сливался с головкой бедренной кости. На правой стороне кости имелся короткий гребень, называемый малым вертелом . Малый вертел направлен вправо и виден на передней стороне кости, как и у многих других пробайногнатов. У более базальных цинодонтов он обычно был более направлен назад и не был виден на передней стороне. Малый вертел не простирался так далеко к проксимальному концу кости, как у базальных млекопитающих. не было Третьего вертела . На задней стороне бедренной кости, между большим и малым вертелами, имелось глубокое и узкое углубление, называемое межвертельной ямкой . На дистальном крае этой ямки имелся межвертельный гребень. который соединял два вертела. Этот гребень также встречается у большинства териан, но отсутствовал у большинства мезозойских групп цинодонтов. Имелась относительно неглубокая треугольная впадина, расположенная дистальнее межвертельного гребня; вероятно, это служило точкой прикрепления приводящих мышц бедра. На задней стороне дистального конца бедренной кости находились два мыщелка, которые должны были соединяться с большеберцовой костью , — медиальный и латеральный мыщелки. Медиальный мыщелок сдавлен из стороны в сторону. Подколенная ямка имела треугольную форму и располагалась близко к мыщелкам. [2]

UFRGS-PV-1043-T сохраняет почти полную левую большеберцовую кость (голень). Это была тонкая кость, длина сохранившихся частей составляла 16,1 миллиметра (0,63 дюйма). Голень в основном прямой, но с небольшим медиальным (вправо) изгибом в проксимальной части (в сторону колена); у более базальных цинодонтов обычно был более сильный медиальный изгиб большеберцовой кости. Стержень становился все более уплощенным к дистальному концу (по направлению к лодыжке). В проксимальной части имелись две суставные фасетки — медиальная и латеральная , сочленяющиеся с мыщелками бедренной кости. Латеральная фасетка была шире медиальной. Дистальный конец большеберцовой кости не сохранился. [2]
UFRGS-PV-1043-T сохраняет две левые кости предплюсны : пяточную и таранную . Пяточная кость (пяточная кость) бразилодона имела низкую и широкую форму, в отличие от узкой пяточной кости, встречающейся у мультитуберкулезных, « симметродонтных » жангеотерий и териев. Он имел выпуклую верхнюю поверхность и вогнутую нижнюю поверхность. имелся хорошо развитый бугорок На заднем конце кости ; эта особенность обнаружена у большинства эуцинодонтов, но отсутствует у более базальных таксонов, таких как Thrinaxodon . имелся малоберцовый выступ На латеральной (левой) стороне пяточной кости . На медиальной (правой) стороне находился выступ, известный как sustentaculum tali , который соединялся с нижней стороной таранной кости (лодыжки). У базальных цинодонтов, таких как Thrinaxodon , не было обнаружено sustentaculum tali, что позволяет предположить, что он был хрящевым, если вообще существовал в этих таксонах. Левая таранная кость была меньше пяточной кости. Он имел полусферическую дорсомедиальную (вверху справа) поверхность и плоскую латероплантарную (внизу слева) поверхность. Передний конец таранной кости имел небольшую головку. Шея, отделявшая голову от остальной части кости, была короче, чем у современных териан. [2]
UFRGS-PV-1043-T также сохраняет две неполные плюсневые кости , состоящие из проксимального и дистального конца, которые должны были соединяться с предплюснами и фалангами соответственно . На проксимальном конце имелось круглое углубление, а на дистальном — два симметричных мыщелка. Возможный фрагмент метаподии (плюсневой или пястной кости ) также известен из экземпляра UFRGS-PV-0765-T, который имел широкую проксимальную часть и более узкое стержне. У обоих этих экземпляров также сохранились некоторые фаланги (кости пальцев), а у UFRGS-PV-0765-T сохранилась почти полная средняя фаланга. Эта фаланга была довольно короткой, с вогнутым проксимальным концом и двумя небольшими мыщелками на дистальном конце. [2]
Классификация
[ редактировать ]Род Brasilodon принадлежит к Brasilodontidae, семейству продвинутых пробайногнатных цинодонтов. Наряду с Brasilodon два современных рода ( Brasilitherium и Minicynodon к семейству отнесены ), оба из которых, вероятно, являются синонимами Brasilodon . [2] [6] Protheriodon , род из среднего триаса ( ладинского периода ) в Бразилии, и Panchetocynodon , малоизвестный цинодонт из раннего триаса ( индского периода ) в Индии, также были помещены в Brasilodontidae в статье Хосе Бонапарта 2013 года. [13] Однако совсем недавно было обнаружено, что протериодон является пробайногнатом, рано расходящимся и не связанным с бразилодонтидами. [14] в то время как Panchetocynodon в остальном рассматривается как таксон incertae sedis , отношения которого с другими таксонами неясны из-за его неполноты. [9] [15]
Особенности черепа и зубных рядов бразилодона указывают на то, что это был производный цинодонт, тесно связанный с млекопитающими, единственными сохранившимися цинодонтами. Филогенетический анализ обычно обнаруживает его как члена клады Mammaliamorpha, расположенной ближе к млекопитающим, чем тритилодонтиды, но за пределами Mammaliaformes, более эксклюзивной клады, содержащей млекопитающих и их ближайших родственников, таких как морганукодонты , докодонты и харамииданы . адаптирована Приведенная ниже кладограмма на основе анализа Уоллеса и др., проведенного в 2019 году. : [16]


| Пробайногнатия |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]
Передвижение
[ редактировать ]Особенности постчерепа указывают на то, что бразилодон был обобщенным животным, способным к различным способам передвижения , включая рытье и лазание. Большая бугристость возле дельтопекторального гребня плечевой кости означает, что большая круглая мышца (важная мышца-ретрактор передней конечности) была довольно большой, как это видно у современных ископаемых грызунов . Однако роющая адаптация бразилодона была менее выражена, чем у современных ископаемых млекопитающих, а также у многих других цинодонтов, не относящихся к млекопитающим. Доказательства способности к сканированию (лазанию) включают полусферические головки плечевых и бедренных костей, которые обеспечивали широкий диапазон вращения плечевых и тазобедренных суставов, хорошо развитый эктепикондилярный гребень, полусферическую головку и почти круглую головку лучевой кости, которая имела бы придавал высокую подвижность локтю и небольшому локтевому отростку. Некоторые особенности плечевой кости, в том числе перекрученный стержень, указывают на то, что у бразилодона были раскидистые или полураскидистые передние конечности. С другой стороны, особенности таза и задних конечностей, такие как плохо развитая поствертлужная часть лопатки подвздошной кости, указывают на более прямое положение. парасагиттальное ) положение задних конечностей. [2]
Замена зуба
[ редактировать ]Согласно одной теории, подробно изложенной в статье Мартинелли и др. была 2010 г. У бразилодона замена зубов полифиодонтом , при этом зубы заменялись более одного раза на протяжении всей жизни животного, хотя неясно, сколько последовательных постклыковых замен было у бразилодона . [11] Постклыковая замена прекратилась задолго до смерти человека, что привело к чрезвычайно сильному износу зубов у пожилых людей. В некоторых случаях зубы человека со временем изнашивались до менее половины высоты оставшейся коронки. [4] Размер постклыковой диастемы указывает на то, что передние постклыки могли быть утрачены быстрее, чем у примитивного цинодонта Thrinaxodon . [11] [10]
По данным Мартинелли и др. (2010) постклыки заменялись поочередно. Замена постклыков должна была происходить сзади вперед, как у некоторых других пробаиногнатий, не относящихся к млекопитающим, и, таким образом, отличалась от замены у ранних млекопитающих, у которых постклыки заменялись спереди назад. Возможно, это было связано с неограниченным ростом черепа ранних пробайногнатов. [11] Вариации замены зубов, возможно, также можно объяснить диетой: у травоядных / всеядных траверсодонтидов были расширенные постклыки и последовательная замена зубов. [17] а плотоядные и насекомоядные пробайногнаты, не относящиеся к млекопитающим, такие как Brasilodon, имеют вместо этого поочередную замену зубов. [11]
Исследование 2022 года, проведенное Cabreira et al. предполагает, что бразилодон на самом деле был дифиодонтом , сменившим зубы только один раз, как и большинство современных млекопитающих. Согласно этой теории, постклыки у бразилодона состояли из набора молочных зубов , которые замещались, и набора постоянных зубов, разделенных на премоляры и коренные зубы . Исследование показывает, что молочные постклыки у Brasilodon прорезывались последовательно в направлении зад-на-перед, тогда как коренные зубы добавлялись в противоположном направлении, спереди назад. [7]
Палеоэкология
[ редактировать ]
Экземпляры бразилодона обнаружены в двух местонахождениях. Голотип был найден в местности Линья-Сан-Луис, в муниципалитете Фашинал-ду-Сотурно. Другие экземпляры были найдены в местности Сесмариа-ду-Пинхал в муниципалитете Канделария; оба эти места находятся в бразильской части бассейна Параны . [18] Породы, в которых был обнаружен Brasilodon, относятся к верхней части толщи Канделария, которая соответствует биостратиграфической единице, известной как Риограндия зона комплекса . Риограндия AZ датируется ранним норианским возрастом позднего триаса. [3] около 225,42 миллиона лет назад. [7] Цинодонты — один из наиболее распространенных и таксономически разнообразных элементов этой зоны объединения. [5] и представлены многими экземплярами хорошей сохранности. [19] Помимо бразилодона , цинодонты представлены трителедонтидами Irajatherium и Riograndia , базальной млекопитающей Botucaraitherium и индетерминантными траверсодонтидами . Другие животные включают дицинодонта Jachaleria , проколофонида Soturnia , лепидозавроморф Cargninia , Clevosaurus и Lanceirosphenodon , а также avemetatarsalians Faxinalipterus , Guaibasaurus , Macrocollum , Maehary , [3] Сакизавр и Унайзавр . неопределенные останки темноспондилов , фитозавров и других групп. Также были обнаружены [18]
Место, где были найдены эти окаменелости, представляет собой речную систему, характеризующуюся большим количеством мелкого песчаника, образующего песчаные пласты, образовавшиеся в результате осаждения осадков в бассейне во время пиковых потоков. [1] [19]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д и ж г час я дж Бонапарт, Дж. Ф.; Мартинелли, АГ; Шульц, CL; Руберт, Р. (2003). «Сестринская группа млекопитающих: мелкие цинодонты из позднего триаса южной Бразилии» . Revista Brasileira de Paleontologia . 5 :5–27.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с т в Гиньяр, ML; Мартинелли, АГ; Соарес, МБ (2019). «Посткраниальная анатомия Brasilodon Quadrangularis и приобретение черт млекопитающих среди цинодонтов, не являющихся млекопитающими» . ПЛОС ОДИН . 14 (5): e0216672. дои : 10.1371/journal.pone.0216672 . ПМК 6510408 .
- ^ Перейти обратно: а б с Келлнер, AWA; Хольгадо, Б.; Грилло, О.; Претто, ФА; Кербер, Л.; Пиньейро, Флорида; Соарес, МБ; Шульц, CL; Лопес, RT; Араужо, О.; Мюллер, RT (2022). «Повторная оценка Faxinalipterus minimus , предполагаемого триасового птерозавра из южной Бразилии, с описанием нового таксона» . ПерДж . 10 : е13276. дои : 10.7717/peerj.13276 . ПМЦ 9074864 .
- ^ Перейти обратно: а б с д и ж г час я дж к Бонапарт, Дж. Ф.; Мартинелли, АГ; Шульц, CL (2005). «Новые сведения о бразилодоне и бразилитериуме (Cynodontia, Probainognathia) из позднего триаса южной Бразилии» . Revista Brasileira de Paleontologia . 8 (1): 25–46.
- ^ Перейти обратно: а б Бонапарт Дж.; Шульц, К.; Соареш, М.; Мартинелли, А. (2010). «Местная фауна Факсинал-ду-Сотурно, поздний триас Риу-Гранди-ду-Сул, Бразилия» . Бразильский журнал палеонтологии . 13 (3): 233–246. дои : 10.4072/rbp.2010.3.07 .
- ^ Перейти обратно: а б с д Лю, Дж.; Олсен, П. (2010). «Филогенетические отношения Eucynodontia (Amniota: Synapsida)» . Журнал эволюции млекопитающих . 17 (3): 151–176. дои : 10.1007/s10914-010-9136-8 .
- ^ Перейти обратно: а б с д Кабрейра, Сан-Франциско; Шульц, CL; да Силва, ЛР; Лора, LHP; Пакульски, К.; до Рего, RCB; Соарес, МБ; Смит, ММ; Рихтер, М. (2022). «Замена дифиодонтных зубов бразилодона - евцинодонта позднего триаса, бросающего вызов времени происхождения млекопитающих». Журнал анатомии : joa.13756. дои : 10.1111/joa.13756 .
- ^ Родригес, разыгрывающий; Руф, И.; Шульц, CL (2014). «Изучение цифровой краниальной эндокасты цинодонта, не являющегося млекопитающим, Brasilitherium riograndensis (поздний триас, Бразилия) и ее значимости для эволюции мозга млекопитающих». Палеонтологическая газета . 88 (3): 329–352. дои : 10.1007/s12542-013-0200-6 .
- ^ Перейти обратно: а б с Мартинелли, AG (2017). «Вклад в знания о пробайногнатских цинодонах (Therapsida, Cynodontia, Probainognathia) из триаса Южной Америки и их влияние на происхождение Mammaliaformes» (на португальском языке).
- ^ Перейти обратно: а б Соарес, МБ; Мартинелли, АГ; де Оливейра, ТВ (2014). «Новый прозостродонтовый цинодонт (Therapsida) из позднетриасовой зоны комплекса Риограндия (суперпоследовательность Санта-Мария) на юге Бразилии» . Анналы Бразильской академии наук . 86 (4): 1673–1691. дои : 10.1590/0001-3765201420140455 . hdl : 10183/119079 .
- ^ Перейти обратно: а б с д и ж Мартинелли, АГ; Бонапарт, Дж. Ф. (2010). «Постсобачья замена у Brasilodon и Brasilitherium (Cynodontia, Probainognathia) и ее значение в эволюции цинодонтов» . Палеонтология и динозавры со времен Латинской Америки : 179–186.
- ^ Придмор, Пенсильвания; Рич, TH; Викерс-Рич, П.; Гамбарян, ПП (2005). «Тахиглоссидная плечевая кость из раннего мела Юго-Восточной Австралии». Журнал эволюции млекопитающих . 12 (3–4): 359–378. дои : 10.1007/s10914-005-6959-9 .
- ^ Бонапарт, Дж. Ф. (2013). «Эволюция Brasilodontidae (Cynodontia-Eucynodontia)». Историческая биология . 25 (5–6): 643–653. дои : 10.1080/08912963.2012.731154 .
- ^ Мартинелли, АГ; Элтинк, Э.; Да-Роса, А. КАК; Лангер, MC (2017). «Новый цинодонт из формации Санта-Мария на юге Бразилии улучшает разнообразие пробайногнатий позднего триаса». Статьи по палеонтологии . 3 (3): 401–423. дои : 10.1002/spp2.1081 .
- ^ Дас, ДП; Гупта, А. (2012). «Новая находка цинодонтов из формации Панчет нижнего триаса, долина Дамодар». Журнал Геологического общества Индии . 79 (2): 175–180. дои : 10.1007/s12594-012-0022-2 .
- ^ Уоллес, РВС; Мартинес, Р.; Роу, Т. (2019). «Первая запись базального млекопитающего из формации Ишигуаласто раннего позднего триаса в Аргентине» . ПЛОС ОДИН . 14 (8): e0218791. дои : 10.1371/journal.pone.0218791 . ПМК 6685608 .
- ^ Мартинелли, АГ; Соарес, МБ; Шванке, К. (2016). «Два новых цинодонта (Therapsida) из среднего-раннего позднего триаса Бразилии и комментарии к южноамериканским пробайногнатам» . ПЛОС ОДИН . 11 (10): e0162945. дои : 10.1371/journal.pone.0162945 . ПМК 5051967 .
- ^ Перейти обратно: а б Мартинелли, АГ; Эскобар, Дж.А.; Францискини, Х.; Кербер, Л.; Мюллер, RT; Руберт, Р.; Шульц, CL; Да-Роза, А.. А.С. (2021). «Новая находка сталекериид-дицинодонта (Therapsida, Dicynodontia) из позднего триаса на юге Бразилии и биостратиграфические замечания о зоне объединения Риограндия». Историческая биология . 33 (11): 3101–3110. дои : 10.1080/08912963.2020.1850715 .
- ^ Перейти обратно: а б де Оливейра, ТВ; Мартинелли, АГ; Соареш, МБ (2011). «Новая информация об Irajatherium hernandezi Martinelli, Bonaparte, Schultz & Rubert 2005 (Eucynodontia, Tritheledontidae) из верхнего триаса (формация Катуррита, бассейн Парана) в Бразилии». Палеонтологический журнал . 85 (1): 67–82. дои : 10.1007/s12542-010-0078-5 .





