Махародонтины
| Махародонтины Временной диапазон: | |
|---|---|
 | |
| Homotherium venezuelensis , Machairodus aphanistus , Metailurus sp. и смилодон фаталис | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Плотоядный |
| Подотряд: | Фелиформия |
| Семья: | кошачьих |
| Подсемейство: | † Махародонтины Гилл , 1872 г. |
| Подгруппы | |
Machairodontinae — вымершее подсемейство хищных млекопитающих семейства Felidae (настоящие кошки). Они были обнаружены в Азии , Африке , Северной Америке , Южной Америке и Европе от среднего миоцена до раннего голоцена .
В состав Machairodontinae входят многие вымершие хищники, широко известные как « саблезубые кошки », включая знаменитый род Smilodon , а также другие кошки, у которых лишь незначительно увеличились размер и длина верхнечелюстных клыков . Название означает «кинжал-зуб», от греческого μάχαιρα ( махайра ), меч. Иногда саблезубыми кошками называют и других хищных млекопитающих с удлиненными зубами, хотя они к кошачьим не относятся. Помимо махайродонтов, ) возникли и другие саблезубые хищники у нимравид , барбурофелид , махаэроидинов , гиенодонтов и даже у двух групп метатерий ( тилакосмилид -спарасодонтов и дельтатероидов . [1]
Этимология
[ редактировать ]Махайрос, от древнегреческого: махайра, букв. «махайра» означает меч, аодонтос (odóntos), что означает «зуб».
Эволюция
[ редактировать ]Семья кошачьих
[ редактировать ]
Machairodontinae возникли в среднем миоцене Европы. [2] Ранние кошачьи Pseudaelurus Quaddentatus продемонстрировали тенденцию к удлинению верхних клыков и, как полагают, лежат в основе эволюции махайродонтов. [3] Самый ранний известный род махайродонтов — среднемиоценовый Miomachairodus из Африки и Турции. [1] До позднего миоцена махайродонтины в нескольких местах сосуществовали вместе с барбурофелидами , архаичными крупными хищниками, которые также имели длинные саблезубы. [1]
Традиционно выделялись три разных трибы махайродонтинов: Smilodontini с типичными зубастыми формами, такими как Megantereon и Smilodon , Machairodontini или Homotherini с ятаганозубыми кошками, такими как Machairodus или Homotherium , и Metailurini , содержащие такие роды, как Dinofelis. и Метайлурус . Однако некоторые недавно перегруппировали Metailurini в другое подсемейство кошачьих, Felinae, вместе со всеми современными кошками. [1] Последним махайродонтином является популяция Smilodon , исчезнувшая в позднем плейстоцене - раннем голоцене в Америке. [4]
Судя по митохондриальной ДНК, последовательностям извлеченным из окаменелостей , линии Homotherium и Smilodon , по оценкам, разошлись около 18 млн лет назад. [5]
Название «саблезубые тигры» вводит в заблуждение. Махародонты не принадлежали к тому же подсемейству, что и тигры, нет никаких свидетельств того, что они имели тигриный рисунок шерсти, и не все представители этой широкой группы животных жили и охотились так же, как современные тигры . Анализ ДНК, опубликованный в 2005 году, подтвердил и уточнил кладистический анализ , показав, что Machairodontinae рано отделились от предков современных кошек и не имеют тесного родства ни с одним из ныне живущих видов кошачьих. [1]
Саблезубые также во многих местах соседствовали с коническими зубами. В Африке и Евразии саблезубые кошки конкурировали с несколькими пантеринами и гепардами вплоть до раннего или среднего плейстоцена. Homotherium сохранялся в Северной Европе даже до позднего плейстоцена. В Америке они сосуществовали с пумой , американским львом , американским гепардом и ягуаром вплоть до позднего плейстоцена. Саблезубые и конусозубые кошки конкурировали друг с другом за пищевые ресурсы, пока не вымерли последние из первых. Все современные кошачьи имеют верхние клыки более или менее конической формы.
Классификация
[ редактировать ]| Племя | Изображение | Род | Разновидность |
|---|---|---|---|
| Неуверенное сиденье | † Чадаилурус [6] [7] Бонис и др., 2018 г. |
| |
| † Гомотерини |  | † Амфимахайрод [8] Крецой, 1929 год. |
|
 | † Гомотерий Фабрини, 1890 г. |
| |
| † Локотунджайлурус Верделин, 2003 г. |
| ||
 | † Нимравидес [8] Сент-Китс, 1958 год. |
| |
 | † Xenosmilus Martin и др., 2000 г. |
| |
| † Махайродонтини | † Hemimachairodus Koenigswald, 1974 г. |
| |
 | † Махайродус Кауп, 1833 г. |
| |
| † Лунчуансмилюс Цзянцзо и др., 2022 г. |
| ||
| † Адейлосмилус Цзянцзо, Верделин, Сан и др., 2022 г. |
| ||
| † Таову Цзянцзо, Верделин, Сан и др., 2022 г. |
| ||
| † Миомахайрод Шмидт-Киттлер 1976 г. |
| ||
| † Метаилурины | † Адельфаилурус Хиббард, 1934 г. |
| |
 | † Динофелис Зданский, 1924 г. |
| |
 | † Метайлурус Зданский, 1924 г. |
| |
| † Стенаилурус |
| ||
| † Фортуниктис Понс-Мойя, 1987 г. |
| ||
| † Йоши [9] Спасов и Гераадс, 2014 г. |
| ||
| † Смилодонтини |  | † Мегантереон Круазе и Жобер, 1828 г. |
|
| † Парамахайрод Пилигрим, 1913 г. |
| ||
| † Промегантереон [8] Крецой, 1938 год. |
| ||
 | † Ризосмилодон Уоллес и Халберт, 2013 г. |
| |
 | † Смилодон Лунд, 1842 г. |
|
Филогения
[ редактировать ]Филогенетические отношения Machairodontinae показаны на следующей кладограмме: [10] [11] [12] [13] [14]
| † Махародонтины |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционная история и происхождение фенотипа
[ редактировать ]До недавнего открытия в 1990-х годах кладбища окаменелостей позднего миоцена, известного как Batallones-1 , экземпляры предков Smilodontini и Homotheriini были редкими и фрагментарными, поэтому эволюционная история фенотипа саблезубых - фенотипа, затрагивающего краниомандибулярные, шейные и передние конечности - анатомия была в значительной степени неизвестна. [17] [18] До раскопок Баталлонес-1 преобладающей гипотезой было то, что высокоразвитый саблезубый фенотип быстро возник в результате плейотропной эволюции. [19] Batollnes-1 обнаружил новые экземпляры Promegantereon ogygia , предка Smilodontini, и Machairodus aphanistus , предка Homotheriini, проливая свет на историю эволюции. [17] [18] (Хотя предок Smilodontini первоначально был отнесен к роду Paramachairodus , позже он был изменен на род Promegantereon ). [20] размером с леопарда P. ogygia (живущий 9,0 млн лет назад) населял Испанию (и, возможно, дополнительную территорию), а его наиболее изученные потомки, представители рода Smilodon размером с тигра , жили до 10 000 лет назад в Америке. [21] размером со льва M. aphanistus (живущий 15,0 млн лет назад) кочевал по Евразии, как и его наиболее изученные потомки, представители рода Homotherium размером со льва (живущий 3,0–5,0 млн лет назад).
Текущая гипотеза эволюции саблезубого фенотипа, ставшая возможной благодаря Batollnes-1, заключается в том, что этот фенотип возник постепенно с течением времени посредством мозаичной эволюции . [17] [18] Хотя точная причина неясна, текущие результаты подтверждают гипотезу о том, что необходимость быстрого убийства добычи была основным фактором, определяющим развитие фенотипа в течение эволюционного времени. Как указывает большое количество случаев сломанных зубов, биотическая среда саблезубых кошек была отмечена острой конкуренцией. [22] [23]
Сломанные зубы указывают на частоту контакта зубов с костью. Увеличение контакта зубов с костями предполагает либо увеличение потребления туш, либо быстрое поедание добычи, либо усиление агрессии по поводу добычи – все три из которых указывают на уменьшение доступности добычи, усиливая конкуренцию между хищниками. Такая конкурентная среда благоприятствует более быстрому убийству добычи, потому что, если добыча отбирается до ее потребления (например, в результате конкуренции), энергетические затраты на поимку этой добычи не возмещаются, и, если это происходит достаточно часто в течение жизни хищник, это приведет к смерти от истощения или голода. Самые ранние приспособления, улучшающие скорость убийства добычи, присутствуют в черепе и нижней челюсти P. ogygia и M. aphanistus . [17] [18] и в шейных позвонках [18] и передняя конечность [24] ogygia P. . Они предоставляют дополнительные морфологические доказательства важности скорости в эволюции фенотипа саблезубых. [24]
Скелет
[ редактировать ]Череп
[ редактировать ]Наиболее изученным отделом группы махайродонтов является череп, а именно зубы. Благодаря большому спектру родов, хорошей представленности окаменелостей, сравнимым современным родственникам, разнообразию внутри группы и хорошему пониманию обитаемых экосистем подсемейство махайродонтов представляет собой один из лучших способов исследования для анализа гиперплотоядных животных, специализации и взаимоотношения хищника и жертвы. [25]

Махародонты делятся на два типа: кортикозубые и ятаганозубые. Кортикозубые кошки имели удлиненные, узкие верхние клыки и, как правило, коренастое тело. Ятаганзубые кошки имели более широкие и короткие верхние клыки и типично гибкое тело с более длинными ногами. Кошки с более длинными зубами часто имели костный выступ , простирающийся от нижней челюсти . Однако один род, Xenosmilus , известный только по двум довольно полным окаменелостям, сломал эту форму; обладая как толстыми, тяжелыми конечностями, характерными для кошек с кинжалами, так и толстыми клыками, как у кошек с ятаганскими зубами.
У плотоядных животных было уменьшено количество зубов, поскольку они специализировались на поедании мяса, а не на измельчении растений или насекомых. У кошек меньше всего зубов среди всех хищников , а у махайродонтов их число еще больше сокращается. сохраняются шесть резцов , два клыка и шесть премоляров У большинства махайродонтов в каждой челюсти два коренных зуба , а в верхней челюсти — только . Некоторые роды, такие как Smilodon , несут только восемь премоляров и на один меньше на нижней челюсти, оставляя только четыре больших премоляра на нижней челюсти вместе с двумя низкорослыми клыками и шестью толстыми резцами. Клыки плавно изогнуты назад, имеются зубцы, но они незначительны и стираются с возрастом, в результате чего у большинства махайродонтов среднего возраста (около четырех или пяти лет) зубцы отсутствуют. [ нужна ссылка ] Подобные подсказки в костях помогают палеонтологам оценить возраст особи для изучения популяции давно вымершего животного.
Более длинные клыки требуют большего зияния. Лев с углом раскрытия 95° не мог носить клыки длиной девять дюймов, потому что у него не было бы зазора между нижними и верхними клыками больше дюйма или около того, чего было бы недостаточно для убийства. Махародонтам, как и другим группам животных, которые приобрели подобные зубы в результате конвергентной эволюции , нужен был способ изменить свои черепа, чтобы разместить клыки несколькими способами.


Основными ингибиторами большого зева у млекопитающих являются височная и жевательная мышцы задней части челюсти. Эти мышцы способны быть мощными и претерпевать значительную степень модификаций для изменения силы укуса, но они не очень эластичны из-за своей толщины, расположения и силы. Чтобы открыть пасть шире, этим видам нужно было уменьшить мышцы и изменить их форму. Первым шагом в этом было уменьшение венечного отростка . Жевательные, и особенно височные, мышцы прикрепляются к этой выступающей полоске кости, поэтому сокращение этого отростка означало сокращение мышц. Меньшая масса каждой мышцы обеспечивала большую эластичность и меньшую устойчивость к широкому зеву. Изменение формы височной мышцы в этом отношении привело к увеличению расстояния между началом и местом прикрепления, так что мышца стала длиннее и компактнее, что, как правило, является более подходящим форматом для этого типа растяжки. Это сокращение привело к ослаблению клева.
Черепа махайродонтов позволяют предположить еще одно изменение формы височной мышцы. Основным препятствием для открытия челюстей является то, что височная мышца порвется, если ее растянуть до критической степени вокруг суставного отростка при открытии рта. У современных кошачьих затылочная кость отходит назад, но височные мышцы, прикрепляющиеся к этой поверхности, напрягаются при широком раскрытии челюсти, поскольку мышца обхватывает суставной отросток. Чтобы уменьшить растяжение височной мышцы вокруг неподвижного отростка, у махайродонтов развился череп с более вертикальной затылочной костью. угол У домашней кошки зева составляет 80°, а у льва — 91°. У смилодона зияние составляет 128°, а угол между ветвью нижней челюсти и затылочной костью - 100°. Этот угол является основным ограничивающим фактором зияния, и уменьшение угла затылочной кости относительно неба рта, как это видно у Smilodon , позволило зиянию еще больше увеличиться. Если бы затылочная кость не была натянута к небу и ближе к перпендикулярно , зазор теоретически будет меньше, примерно 113°. [ нужна ссылка ]
Черепа многих саблезубых хищников, в том числе махайродонтов, высокие сверху вниз и короткие спереди назад. Скуловые дуги сжаты, часть черепа с чертами лица, например глазами, выше, а морда короче. [26] Эти изменения помогают компенсировать увеличенный зев. [27] У махайродонтов также были уменьшены нижние клыки, сохраняя расстояние между клыками верхней и нижней челюстей. [ нужна ссылка ]





Посткраниальный скелет
[ редактировать ]Кортикозубые махайродонты, в том числе смилодон , мегантерон и парамахайродус , отличаются выносливостью и силой, при этом самый примитивный ( парамахайродус ) меньше и гибче, чем более продвинутый смилодон ; промежуточный Мегантереон находится между ними. Они не были выносливыми бегунами с короткими лапками и плюснами и тяжелым телом. По сравнению с современными львами , их грудные клетки имели бочкообразную форму с узкими передними концами и расширенными задними концами. Их лопатки были очень хорошо развиты, особенно у Smilodon , что позволяло обеспечить большую площадь поверхности прикрепления массивных плечевых и трехглавых мышц. Шейные позвонки очень крепкие, а крепления мышц были мощными и крепкими. Поясничный отдел позвоночника укорочен. Хвосты, от самых примитивных до самых продвинутых, становились все короче и короче, в результате чего у Смилодона появился хвост, похожий на рысь . Если рассматривать только посткраниальные останки, то они по строению больше похожи на современные. медведям, чем современным кошкам. [28]
Ятаганзубые махайродонты ( Machairodontini , Homotherini и Metailurini ) представляют собой гораздо более разнообразную группу. Клыки этой более крупной группы значительно короче и, как правило, толще, но все же намного длиннее, чем у любой современной кошки. Из-за разнообразия родов трудно проиллюстрировать конкретный тип. Махародонтины были первыми среди махайродонтинов и кошачьих в целом, которые достигли размеров, близких к львиным, и уже на ранних этапах своей эволюции в миоцене демонстрировали впечатляющие верхние клыки, но помимо них сохраняли относительно кошачью морфологию, которая была больше похожа на современную. пантерины , чем более производные махайродонтины более поздних периодов. Махайродус, похоже, был отличным прыгуном. Гомотерины в целом были более специализированными, и уже самые ранние таксоны, такие как Lokotunjailurus, были удивительно длинноногими и худощавыми, хотя и такими же большими, как современный лев, и эта тенденция еще больше усилилась в плиоцен-плейстоценовом роде Homotherium, который когда-то считался стопохождение , но было доказано, что оно является пальцевидным . [13] Сыворотка Homotherium , наиболее известный вид из плейстоцена Северной Америки, имела наклонную спину, что, возможно, делало ее превосходной в беге на большие расстояния, подобно ныне живущей пятнистой гиене . У него также была хорошо развитая зрительная кора, большая носовая полость , которая позволяла лучше усваивать кислород, и меньшие, лишь частично втягивающиеся когти, которые могли функционировать как шипы для лучшего сцепления с землей. очень активный образ жизни и беглость . [29] Однако Xenosmilus , гомотерин среднего плейстоцена из Флориды и близкий родственник Homotherium , нарушил эту тенденцию, поскольку у него были как ятагановидные зубы, так и громоздкое и сильное строение, более типичное для кортикозубов. [30]
Третье племя ятаган-зубых, Метайлурини, имело общее сходство с современными кошками, но было весьма разнообразным с точки зрения морфологии: виды варьировались от маленького гепарда до маленького льва. Некоторые из них имели сравнительно короткие, почти конические верхние клыки, похожие на современные кошки, тогда как некоторые виды имели ярко выраженные махайродонтные черты. Однако, в отличие от гомотерин и смилодонтинов, даже у наиболее производных металуринов сохранились длинные хвосты, длинные задние ноги и длинный позвоночник. [31] В среднем у ятаганозубых кошек было больше зубов, чем у среднего махайродонта с кортикальными зубами, с шестью премолярами на нижней челюсти . При рассмотрении только посткраниальных остатков однозубых махайродонтов многие их формы были сравнительно схожи с современными пантеринами (роды Panthera и Neofelis ). [28]
Производная анатомия и диета
[ редактировать ]
Сила укуса
[ редактировать ]Челюсти махайродонтов, особенно более продвинутых видов с более длинными клыками, таких как смилодон и мегантереон , необычайно слабы. Цифровые реконструкции черепов львов и смилодона показывают, что последний плохо справился бы со стрессом, связанным с удержанием борющейся добычи. [32] Основная проблема заключалась в нагрузках, которым подвергалась нижняя челюсть: сильная сила угрожала сломать челюсть, поскольку давление оказывалось на ее самые слабые места.
Сила укуса смилодона была бы в три раза слабее льва, если бы он использовал только челюстные мышцы. Однако мышцы шеи, которые соединялись с задней частью черепа, были сильнее и давили на голову, заставляя череп опускаться вниз. Когда челюсть была чрезмерно вытянута, мышцы челюсти не могли сокращаться, но мышцы шеи прижимали голову вниз, заставляя клыки прижиматься ко всему, что им сопротивлялось. Когда рот был закрыт достаточно сильно, мышцы челюсти могли немного поднять нижнюю челюсть. [32]

Диета
[ редактировать ]Иногда кости окаменелого хищника сохраняются достаточно хорошо, чтобы сохранить узнаваемые белки, принадлежащие тому виду, который он потреблял при жизни. [33] [34] Анализ стабильных изотопов этих белков показал, что Smilodon охотился в основном на бизонов и лошадей, а иногда и на наземных ленивцев и мамонтов, в то время как Homotherium часто охотился на молодых мамонтов и других травоядных, таких как вилорогая антилопа и снежный баран, когда мамонты были недоступны. [30] Исследования Xenosmilus, опубликованные в 2022 году, посвященные стиранию зубов смилодона и следам укусов на костях пекари Platygonus , проведенные Xenosmilus, позволяют предположить, что махайродонты были способны эффективно сдирать и обезжиривать мясные туши при кормлении. Они также демонстрируют степень потребления костей наравне с современными львами, которые сами могут и регулярно поедают более мелкие кости во время еды. [35]
Лицо
[ редактировать ]Американский палеонтолог Джордж Миллер изложил набор особенностей, ранее не мыслившихся у мягких тканей махайродонтов, в частности смилодона . [25]
Первым изменением, которое он предложил во внешности махайродонтов, были более низкие уши, или, скорее, иллюзия нижних ушей из-за более высокого сагиттального гребня . Однако расположение ушей у современных кошачьих всегда одинаково, даже у особей, имеющих гребни, сравнимые по размерам с гребнями саблезубых кошек. [36] Расположение ушных раковин , или внешних ушей, а также цвет шерсти зависят от человека, проводящего реконструкцию. Большие или маленькие, заостренные или округлые, высокие или низкие, окаменелости не отражают этих характеристик, оставляя их открытыми для интерпретации.
Миллер также предложил нос, похожий на мопса. За исключением мопсов и подобных им собак, ни у одного современного хищника нет курносого носа, поскольку это неестественно созданная черта, возникшая в результате селекционного разведения. Относительно низкое распространение курносого носа привело к тому, что его обычно игнорировали. [37] Обоснование Миллера основано на втягивании носовых костей смилодона . Критика теории Миллера сравнивает носовые кости львов и тигров. Львы, по сравнению с тиграми, также имеют сильно втянутые носовые кости, но у льва ринарий , или наружный нос, втянут не больше, чем у тигра. Таким образом, курносый нос смилодона, предложенный Миллером, мало что подтверждает в физическом строении сопоставимых животных. [ нужна ссылка ] По данным Антона, Гарсиа-Переа и Тернера (1998), ноздри современных кошачьих всегда занимают одинаковое положение, независимо от длины носовых костей, которая у Smilodon находится в пределах диапазона, наблюдаемого у современных видов. [36]
Третья предложенная идея – удлинение губ на 50%. Хотя другие его гипотезы были в значительной степени отвергнуты, последняя широко используется в современных изображениях. Миллер утверждает, что более длинные губы обеспечивают большую эластичность, необходимую для укуса добычи с более широким раскрытием. Хотя этот аргумент оспаривается в научном сообществе, [ нужна ссылка ] тем не менее, он по-прежнему поддерживается художниками. Научная критика отмечает, что губы современных кошек, особенно крупных видов, обладают невероятной эластичностью, а губа обычной длины хорошо растягивается, несмотря на большую степень раскрытия. [38] и что у современных хищников линия губ всегда находится впереди жевательной мышцы, которая у Smilodon располагалась сразу за хищными. [36] Тем не менее, реконструкции Smilodon , Machairodus и других видов показаны с длинными губами, часто напоминающими челюсти больших собак.
Исследования Homotherium и Smilodon показывают, что ятаганозубые махайродонты, такие как сам Homotherium, обладали верхними губами и тканью десен, которые могли эффективно скрывать и защищать их верхние клыки; эта черта была у них общей с современными видами кошек, в то время как у смилодона были клыки, которые оставались частично обнаженными и выступали за губы и подбородок, даже когда пасть была закрыта из-за их большой длины. [39]
Вокализации
[ редактировать ]Сравнение подъязычных костей смилодона и львов показывает, что первые и, возможно, другие махайродонты потенциально могли рычать, как их современные родственники. [40] [41]
Социальное поведение
[ редактировать ]Смилодон
[ редактировать ]Исследование 2009 года сравнило соотношение социальных и одиночных хищников в заповедниках Южной Африки и Танзании с соотношением окаменелостей в калифорнийских смоляных ямах Ла Бреа , хорошо известном пласте окаменелостей плейстоцена. [42] и как они реагировали на записанные звуки умирающей добычи, чтобы сделать вывод, был ли Смилодон социальным или нет. Когда-то смоляные ямы Ла Бреа представляли собой глубокую смолу, в которой оказывались ловушки животных. Когда они умирали, их крики привлекали хищников, которые, в свою очередь, тоже попадали в ловушку. Он считается лучшим пластом окаменелостей плейстоцена в Северной Америке по количеству животных, пойманных и сохранившихся в смоле, и может быть аналогичен ситуации, созданной в исследовании. Предполагалось, что одиночные хищники не будут приближаться к источникам таких звуков из-за опасности конфронтации с другими хищниками. Социальным хищникам, таким как львы, нечего бояться других хищников, и они с готовностью присоединяются к этим призывам. Исследование пришло к выводу, что эта последняя ситуация наиболее точно соответствует соотношению животных, обнаруженных в смоляных ямах Ла-Бреа, и, следовательно, смилодон, скорее всего, был социальным. [42]
Гомотерий
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( Март 2017 г. ) |

В пещере Фризенхан, штат Техас, были обнаружены останки почти 400 молодых мамонтов вместе со скелетами Homotherium . Homotherium Предполагается, что группы специализировались на охоте на молодых мамонтов и перетаскивали добычу в укромные пещеры, чтобы питаться внутри, а не на открытом воздухе. Они также сохраняли превосходное ночное зрение, и ночная охота в арктических регионах, вероятно, была их основным методом охоты. [43]
Современный лев способен в больших количествах убивать ослабленных взрослых и здоровых полувзрослых слонов, поэтому гомотерий аналогичного размера , вероятно, мог бы совершить тот же подвиг с молодыми мамонтами. [44] Это подтверждается изотопным анализом. Но идея о том, что кошка, даже очень крупная и, возможно, социальная, способна совместно «перетащить» 180-килограммового (400 фунтов) детеныша мамонта на любое реальное расстояние в пещеру, не повредив при этом его зубы, вызвала резкую критику. [ кем? ] Его наклонная спина и мощная поясничная часть позвонков наводили на мысль о медвежьем телосложении, поэтому он мог тянуть тяжести, но ломать клыки - судьба, от которой с некоторой частотой страдали Махайродус и Смилодон , - не наблюдается у Homotherium . Более того, на костях этих молодых мамонтов имеются характерные следы резцов Homotherium , что указывает на то, что они могли эффективно перерабатывать большую часть мяса на туше, что указывает на то, что именно они, а не падальщики, тащили туши в пещеры. [31] Исследование костей также показывает, что туши этих мамонтов были расчленены кошками перед тем, как их утащили, что указывает на то, что Homotherium расчленял свою добычу, чтобы перевезти ее в безопасное место и не дать падальщикам забрать с трудом добытую еду. Имеющиеся данные также показывают, что кошки могли эффективно отделять плоть от костей таким образом, что оставляли заметные следы. [45]
Палеопатология
[ редактировать ]
Machairodus — еще один род, в котором мало окаменелостей, что позволяет предположить его социальную природу, но клыки у этих видов ломаются чаще, чем у других, и после этого проявляются признаки обширного заживления. Самец Amphimachairodus giganteus из Китая, хранящийся в Институте палеонтологических исследований Бабиарца, представляет собой пожилую особь со сломанным клыком, изношенным в результате использования после перерыва. Однако человек умер от тяжелой инфекции носа — травмы, которую у социального хищника было бы больше шансов вылечить, поэтому череп можно интерпретировать по-разному. [46] Взрослым клыкам молодых махайродов требовалось исключительно много времени, чтобы прорезаться и начать использоваться, поэтому до тех пор он полностью зависел от заботы своих родителей. [ нужна ссылка ]
Еще одним примером палеопатологии, подтверждающим социальную гипотезу, является большое количество окаменелостей смилодонов из смоляных ям Ла Бреа, имеющих охотничьи травмы. Помимо травм, полученных в результате перенапряжения во время охоты, более серьезные травмы явно предполагают социальный характер. Животные могли быть искалечены еще долгое время после заживления травмы, у них были опухшие лодыжки, выраженная хромота и ограничение подвижности, которое сохранялось в течение многих лет. [40] В одном из таких случаев речь идет о несовершеннолетнем, страдающем от разбитого таза, который зажил. Особь с трудом могла бы использовать поврежденную конечность и медленно хромала, отдавая предпочтение трем остальным, будучи совершенно неспособной охотиться самостоятельно. [47] Если бы одинокому хищнику удалось пережить такую тяжелую травму, это был бы очень редкий случай. Гораздо более вероятно, что такое животное не могло бы двигаться с одного места на земле в течение нескольких месяцев и могло выжить, только если бы ему приносили еду или тащилось к добыче, устроенной родственниками. [ нужна ссылка ]
Опровержение социальной гипотезы
[ редактировать ]Вопрос о социальности до сих пор остается дискуссионным. Сильная поддержка традиционной концепции одиночного смилодона обнаруживается в его мозге. У большинства социальных хищников, включая людей, серых волков и львов, мозг немного больше, чем у их родственников-одиночек. У смилодона был относительно небольшой мозг, что предполагало меньшую способность к сложному совместному поведению, например, к охоте в группах. [48]
Однако сломанные кости, похоже, все еще поддерживают социальность. [ по мнению кого? ] Лучшим объяснением исцеления животных от серьезных ран в одиночку является то, что кошки создают метаболические резервы, которые можно использовать в случае необходимости. Гепарда часто рассматривают как плохой пример, поскольку это специализированный вид с более хрупким телосложением, чем другие кошки. Было замечено, что более крупные и крепкие виды кошек, такие как львы и леопарды, восстанавливаются после серьезных травм, таких как сломанные челюсти и порванные мышцы. [ нужна ссылка ]
Функциональность сабель
[ редактировать ]Колющие удары
[ редактировать ]Было высказано предположение, что махайродонты использовали свои саблевидные зубы во время охоты, схватывая животное, открывая ему пасть и поворачивая голову вниз с достаточной силой, чтобы проколоть кожу и плоть животного. Когда-то было высказано предположение, что зубы сабли использовались так же, как нож . [25] Первоначально клыки казались инструментами огромной силы и разрушительной способности, используемыми для дробления позвонков или для вскрытия панцирных животных, таких как глиптодонты . [ нужна ссылка ]
Однако зубы состоят из эмали без опоры , и их можно было бы легко сломать о твердый материал, например кость. Также утверждалось, что нижняя челюсть и неспособность очень широко открыть рот были бы препятствием для эффективного нанесения удара ножом. [25] По этим причинам эта концепция была отвергнута научным сообществом. [ нужна ссылка ]
Сексуальная характеристика
[ редактировать ]Возможно, этот раздел содержит оригинальные исследования . ( Ноябрь 2013 г. ) |

Длинные клыки также могли быть продуктом полового отбора , подобно львиной гриве, и использовались для ухаживания, сексуальной демонстрации и социального статуса. Их клыки уже давно признаны относительно хрупкими, а мышцы челюстей недостаточно сильными, поэтому любая хищническая функция сомнительна. [49]
Однако когда черта используется для усиления сексуального влечения, обычно только один пол, обычно мужчины, проявляет эту особенность. У всех видов махайродонтов как самцы, так и самки имеют эти клыки и, за небольшим исключением, как у Machairodus , имеют одинаковую форму. [ нужна ссылка ] Обычно также существует разница в размерах между полами, но махайродонты мужского и женского пола, по-видимому, были одинакового размера. Кроме того, такой уровень полового отбора кажется чрезмерным, учитывая, что у человека будут серьезные нарушения в еде и общих функциях. [25]
Уборка мусора
[ редактировать ]Одно из предположений состоит в том, что большинство махайродонтов были падальщиками . Это делает клыки по большей части нефункциональными и часто сочетается с гипотезой полового отбора. Многие современные хищники в большей или меньшей степени питаются мусором. Сильное обоняние и хороший слух могли бы помочь найти трупы или украсть добычу других хищников, таких как ужасные волки или короткомордые медведи , и при этом не было бы необходимости бегать, как это видно по коренастому телосложению большинства махайродонтов. [25]

Многие современные кошки демонстрируют эту смесь черт. Львы — способные охотники, но будут воровать, когда им представится такая возможность. Тигры и пумы закапывают добычу и возвращаются позже, чтобы продолжить трапезу, даже через несколько дней. Все кошки предпочитают убивать больных или раненых, и существует тонкая грань между животным, настолько больным, что оно не может двигаться, и мертвым животным. Обилие скелетов смилодонов в смоляных ямах Ла Бреа в Калифорнии также подтверждает эту гипотезу. Животные, попавшие в ямы, либо умирали, либо были мертвы — от такой еды настоящий гиперхищник, такой как современный гепард , отказался бы. [ нужна ссылка ] Эта гипотеза является самой старой, но до сих пор считается жизнеспособной. [ нужна ссылка ]
Противодействие этой концепции наблюдается во многих частях тела кошки. Зубы чисто плотоядные, [25] неспособны измельчать растительный материал, как это делают всеядные зубы собак и медведей. позволяет Форма хищников эффективно резать плоть, а не хрустеть кости, как у современных пятнистых гиен. Поскольку оба пола имеют эти клыки и присутствуют дополнительные модификации черепа, махайродонты, вероятно, в некоторой степени были оппортунистами. [ нужна ссылка ]
Гипотезы о кусании шеи
[ редактировать ]Этот раздел нуждается в дополнительных цитатах для проверки . ( январь 2014 г. ) |
Более распространенный и широко распространенный взгляд на охоту на махайродонтов - это укус с перерезанием горла. Современные кошки используют горловой зажим — укус, расположенный в верхней части горла, чтобы задушить добычу, сжимая трахею. [25] Их клыки служат для прокалывания кожи и в основном обеспечивают лучший захват и не наносят существенного вреда добыче. С другой стороны, махайродонты нанесли бы ущерб, если бы использовали ту же технику, что и их современные родственники. [50]
Основным недостатком этих методов является то, что большое количество пролитой крови могут учуять другие находящиеся поблизости хищники, такие как другие махайродонты или ужасные волки. Хищники часто образуют конкурентные отношения, в которых доминирование может переходить от одного вида к другому, как это видно на примере современного льва и пятнистой гиены в Африке. В таких ситуациях склоки – не редкость. Баланс сил и доминирования между этими высшими хищниками остается загадкой из-за социального фактора. Численная сила может сыграть значительную роль в этой борьбе. Например, считается, что ужасные волки путешествовали небольшими стаями, и, хотя они подчинялись индивидуально, их численности могло быть достаточно, чтобы заставить махайродонта отказаться от добычи.
Тем не менее, кошка могла бы найти добычу, убитую ужасными волками. Два одиночных махайродонта быстро выработали бы иерархию с первым. [ нужны разъяснения ] индивидуальная доминанта. Из-за этой неопределенности большая часть ниши махайродонтов до сих пор неизвестна. Все несколько вариантов этой гипотезы требуют подавленного и неподвижного животного.
Генерал «кусай и отступай»
[ редактировать ]Первая гипотеза, связанная с чувствительной шеей, заключается в том, что кошка просто удерживала животное, а затем укусила шею, без особого указания места, чтобы вызвать серьезное повреждение крови, а затем отступила, позволив животному истечь кровью. Условия включают в себя не кусать заднюю часть шеи, где контакт с позвонками может сломать зубы, но глубокий укус в любом месте шеи может оказаться смертельным. [51]
Этот общий укус будет использоваться везде, где его можно достичь, и для него потребуется меньше хищников. По сравнению с гипотезой стрижки живота, один Мегантереон мог убить большого оленя и, возможно, лошадь с небольшой опасностью сломать клыки. Это связано с тем, что укус может быть нанесен, в то время как хищник по большей части держит свое тело позади добычи, избегая подбрасывания ног, но при этом продолжая давить своим весом тела, чтобы удерживать ее на месте. Это был бы быстрый укус, подходящий для стиля выслеживания и охоты из засады, подразумеваемого тяжелыми и сильными телами большинства махайродонтов. Одинокий махайродонт также мог ранить таким образом крупное животное-жертву, затем отпустить его и следовать за ним, пока оно не упадет от шока.
Общая гипотеза «укуси и отступи» подверглась критике. [ кем? ] из-за своей кровопролитности и потому, что борющаяся добыча могла привлечь любых хищников и падальщиков в этом районе. Идея о том, что одно животное может ранить, отпустить и преследовать добычу, получила более сильное противодействие. Кошки редко уходят от добычи, пока не наедятся, иначе она рискует быть украдена другими хищниками.
В частности, Xenosmilus мог использовать этот метод, поскольку все зубы во рту были зазубренными и выровнены таким образом, что образовывали единую режущую поверхность. [52]

«Откусить и сжать»
[ редактировать ]Когда животное ранено укусом махайродонта (игнорируя расположение кровеносных сосудов, которые в этой гипотезе незначительны), клыки должны были быть вставлены за дыхательное горло, а премоляры охватывали бы дыхательное горло . В этом варианте говорится, что махайродонт сжимал трахею после укуса, одновременно удушая и ранив жертву. Прокол крупных кровеносных сосудов в горле и вызвать массивное кровотечение ускорит смерть животного.
Современные кошки и, предположительно, базальные роды всех кошек, такие как Pseudaelurus и Proailurus , используют зажим на горле как обычный метод уничтожения добычи. Удушение заглушило бы звук испуганной добычи - метод, используемый современными гепардами и леопардами. Рана от клыков и недостаток воздуха могли убить животное-жертву.
Этот метод может полностью свести на нет эффект раны, нанесенной клыками. Если оставить клыки в ране, это затруднит приток крови к телу и может продлить жизнь животному, даже если добыча не может издавать звуки. В этом методе убийства нет существенного преимущества у более длинных клыков по сравнению с предковыми кошками с их короткими клыками конической формы. Во всяком случае, опасность сломать зубы, застрявшие в горле паникующего животного, даже если оно хорошо сдерживается, перевешивает возможные преимущества, поэтому этот метод часто рассматривается как маловероятный. [ кем? ]

Осторожный «режущий прикус»
[ редактировать ]Еще один вариант [13] предполагает, что продвинутые махайродонты были узкоспециализированными, достаточными для того, чтобы получить определенную геометрию, позволяющую проколоть четыре основных кровеносных сосуда в горле животного-жертвы за один укус. Эта гипотеза будет включать в себя осторожный укус с целью проколоть кровеносные сосуды, аналогичный, но более точный, чем гипотеза укуса и сжатия, после чего махайродонт отступит и позволит животному очень быстро истечь кровью.
Хотя этот метод и кровавый, из всех гипотез этот метод потребует наименьшего времени, чтобы убить животное. Из-за различий в анатомии видов, на которых, возможно, охотятся махайродонты, геометрия, необходимая, например, для убийства лошади, может не подойти для бизона. Это потребовало бы, чтобы род или даже конкретный вид были узкоспециализированы для одного типа животных-жертв. Это могло бы дать объяснение их исчезновению, поскольку перемещение или исчезновение этого вида-жертвы привело бы к гибели его специализированного хищника.
Кажется, высокая специализация [ кому? ] крайняя и ненужная версия перерезания горла, основанная на принципе «укуси и отступи», но предположение о том, что виды махайродонтов стали более специализированными для охоты на один вид добычи, обычно считается приемлемым, пока это заблуждение [ кем? ] что махайродонты охотились «только» на этот вид. Однако это не решит проблему беспорядка и громких звуков, которые, вероятно, связаны с таким видом поклевки. Вероятно, понадобилось бы более одного человека, чтобы обеспечить полное подавление животного.
«Стрижка живота»
[ редактировать ]В 1985 году американский палеонтолог Уильям Акерстен предложил стригущий укус. [53] [54] Этот метод убийства похож на стиль убийства, наблюдаемый сегодня у гиен и собак. Группа махайродонтов захватила и полностью подчинила добычу, удерживая ее неподвижно, пока один из группы вгрызался в брюшную полость , отдергивался и разрывал тело.
Чтобы этот метод работал, необходимо соблюдать определенную последовательность движений. Во-первых, животное должно быть полностью подчинено, а хищные махайродонты должны быть социальными, чтобы несколько особей могли удержать животное-жертву. Особь, готовящаяся нанести смертельный укус, открывала рот максимально широко и нижней челюстью нажимала на кожу живота. Создавая углубление там, где нижние клыки и резцы прижимаются к коже, на коже над нижними зубами создается небольшая складка, когда нижняя челюсть поднимается вверх. Затем верхние клыки прижимаются к коже, а мышцы шеи используются для опускания головы, поэтому вместо того, чтобы тянуть челюсть «вверх», череп прижимается «вниз». Когда клыки прокалывают кожу, они опускаются до тех пор, пока угол раскрытия рта не составит примерно 45°, при этом нижняя челюсть подтягивается вверх, а череп все еще остается вдавленным. Небольшие выступы на передней части нижней челюсти большинства махайродонтов использовались для вдавливания черепа. Когда пасть животного закрыта, оно удерживает толстый лоскут кожи между челюстями, за клыками, и животное использует мышцы нижней части спины и передних конечностей, чтобы оттянуться назад, отрывая лоскут от тела. Когда эта большая рана открыта, кишки остаются незакрытыми, а артерии и вены разрываются. Истекающее кровью животное умрет в течение нескольких минут, а шок от повторяющихся укусов, вырывающих из тела внутренности, может ускорить этот процесс. [55]
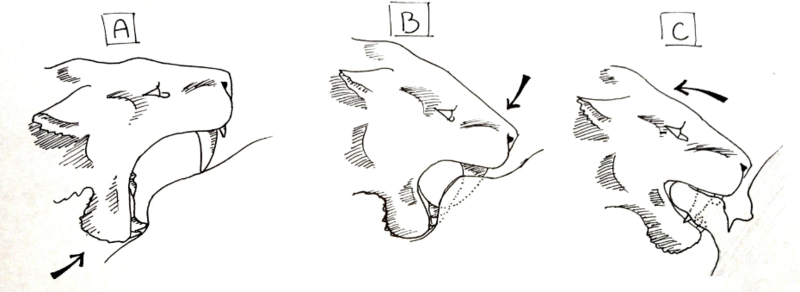
Этот метод позволяет социальным махайродонтам наносить добыче большие раны. За этим последует массивная кровопотеря, и, хотя эта социальная группа будет кровопролитной, она сможет дать отпор практически любому животному, привлеченному в этот район. Укус не обязательно должен быть конкретным, его можно повторить, чтобы ускорить смерть животного, и это уже наблюдается в методах убийства нескольких современных видов, таких как пятнистая гиена. Клыки реже ломаются из-за более мягкого характера живота по сравнению с горлом, а подергивания в животе не так усиливаются, как в шее. Гипотеза о разрыве живота в целом считалась весьма правдоподобной. [ кем? ] В смоляных ямах Ла Бреа случаи сломанных клыков у смилодонов редки, и этому менее рискованный метод мог способствовать этому. [56]

Однако резкий прикус мог быть проблематичным для махайродонтов по нескольким причинам. Большинство копытных очень чувствительны в области живота и задних конечностей, и большинству хищников гораздо легче поймать и усмирить животное, похожее на домашнюю корову, манипулируя головой и передними конечностями. Опустив животное на землю и поместив его между парами ног, махайродонт подвергся бы большому риску получить удар ногой. Сила такого удара легко могла бы сломать зубы, нижнюю челюсть или ногу, а также покалечить или убить кошку.
Общительность могла бы решить эту проблему, если бы один человек нанес смертельный укус, в то время как другие удерживали животное на месте. Более того, диаметр брюшка крупного копытного животного, такого как бизон, мог быть слишком большим, а кожа слишком натянутой, чтобы махайродонт мог вообще схватить лоскут кожи, не говоря уже о том, чтобы оторвать его от тела. Третья проблема, связанная со стригущим укусом, заключается в том, что для успеха клыкам придется проделать большую дыру в животе животного, но вместо этого они могут просто содрать кожу и оставить две длинные порезы. Эта рана может быть болезненной и кровоточить, но животное, скорее всего, не истечет кровью и все равно сможет убежать и выжить, вместо того, чтобы истечь кровью.
В 2004 году в эксперименте использовалась пара механических алюминиевых челюстей, отлитых на основе компьютерной томографии Smilodon Fatalis из смоляных ям Ла Бреа, для имитации нескольких техник укуса, возможно, используемых Smilodon , включая режущий укус, на свежей туше домашней коровы. [57] Было обнаружено, что живот коровы был слишком большим в диаметре, чтобы клыки могли проколоть кожу, но вместо этого они были отклонены от тела, а нижняя челюсть блокировала им доступ. Однако модель вытянула челюсть вверх, как кусают современные кошки, а махайродонты, скорее всего, этого не сделали, а вместо этого прижали череп вниз с помощью мышц шеи. Этот недостаток процедуры может свести на нет результаты и оставить гипотезу о стрижке живота нетронутой.
Примечания и ссылки
[ редактировать ]- ^ Jump up to: а б с д и Ларс В. ван ден Хук Остенде, Майкл Морло и Дорис Нагель (июль 2006 г.). «Окаменелости объяснили 52 Величественных убийцы: саблезубых кошек». Геология сегодня . 22 (4): 150–157. дои : 10.1111/j.1365-2451.2006.00572.x . S2CID 128960196 .
- ^ Верделин, Ларс; Ямагучи, Нобуюки; Джонсон, МЫ (январь 2010 г.). «Филогения и эволюция кошек (Felidae)». В Макдональде, DW; Ловеридж, Эй Джей (ред.). Биология и охрана диких кошачьих . Издательство Оксфордского университета. стр. 59–82.
- ^ Джорди Аугусти: Мамонты, саблезубые и гоминиды, 65 миллионов лет эволюции млекопитающих в Европе , Columbia University Press, 2002. ISBN 0-231-11640-3
- ^ Манцуэтти, А.; Переа, Д.; Джонс, В.; Убилла, М.; Риндеркнехт, А. (2020). «Чрезвычайно большой череп саблезубой кошки из Уругвая (поздний плейстоцен – ранний голоцен, формация Долорес): размер тела и палеобиологические значения». Алчеринга: Австралазийский журнал палеонтологии . 44 (2): 332–339. Бибкод : 2020Алч...44..332М . дои : 10.1080/03115518.2019.1701080 . S2CID 216505747 .
- ^ Пайманс, JLA; Барнетт, Р.; Гилберт, MTP; Сепеда-Мендоса, ML; Реймер, JWF; де Вос, Дж.; Зазула, Г.; Нагель, Д.; Барышников Г.Ф.; Леонард, Дж.А.; Роланд, Н.; Вестбери, Миннесота; Барлоу, А.; Хофрейтер, М. (19 октября 2017 г.). «Эволюционная история саблезубых кошек на основе древней митогеномики» . Современная биология . 27 (21): 3330–3336.e5. дои : 10.1016/j.cub.2017.09.033 . ПМИД 29056454 .
- ^ Бонис, Луи де; Пенье †, Стефан; Маккей, Хасан Тайссо; Ликиус, Андосса; Виньо, Патрик; Брюне, Мишель (15 февраля 2018 г.). «Новые саблезубые кошачьи (Carnivora, Mammalia) на местонахождении гоминидов Торос Меналла (поздний миоцен, Чад)» (PDF) . Геодиверситас . 40 (1): 69. doi : 10.5252/geodiversitas2018v40a3 . S2CID 134769588 . Проверено 30 ноября 2022 г.
- ^ «Палеопрофиль: кот Чад» . Архивировано из оригинала 06 апреля 2018 г. Проверено 5 апреля 2018 г.
- ^ Jump up to: а б с д и ж г Антон, Маурисио (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. ISBN 9780253010421 .
- ^ Спасов, Николай; Гераадс, Денис (2014). «Новый кошачий из позднего миоцена Балкан и состав рода Metailurus Zdansky, 1924 (Carnivora, Felidae)». Журнал эволюции млекопитающих . 22 : 45–56. дои : 10.1007/s10914-014-9266-5 . S2CID 14261386 .
- ^ «Палеобиологическая база данных» . Архивировано из оригинала 25 марта 2012 г. Проверено 16 июня 2011 г.
- ^ Тернер, Алан (1990). «Эволюция гильдии крупных наземных хищников во время плио-плейстоцена в Африке». Геобиос . 23 (3): 349–368. Бибкод : 1990Geobi..23..349T . дои : 10.1016/0016-6995(90)80006-2 .
- ^ Мартин, LD; Бабиарз, JP; Неаполь, Вирджиния; Херст, Дж. (2000). «Три способа быть саблезубым котом». Naturwissenschaften . 87 (1): 41–44. Бибкод : 2000NW.....87...41M . дои : 10.1007/s001140050007 . ПМИД 10663132 . S2CID 1216481 .
- ^ Jump up to: а б с Тернер, Алан (1997). Большие кошки и их ископаемые родственники . Нью-Йорк: Издательство Колумбийского университета. п. 60. ИСБН 978-0-231-10228-5 .
- ^ Уоллес, Южная Каролина; Халберт, RC (2013). Ларсон, Грегер (ред.). «Новый махайродонт из фауны пальметто (ранний плиоцен) Флориды с комментариями о происхождении смилодонтини (млекопитающих, хищников, кошачьих)» . ПЛОС ОДИН . 8 (3): e56173. Бибкод : 2013PLoSO...856173W . дои : 10.1371/journal.pone.0056173 . ПМЦ 3596359 . ПМИД 23516394 .
- ^ Спасов, Николай; Гераадс, Денис (2015). «Новый кошачий из позднего миоцена Балкан и состав рода Metailurus Zdansky, 1924 (Carnivora, Felidae)». Журнал эволюции млекопитающих . 22 : 45–56. дои : 10.1007/s10914-014-9266-5 . S2CID 14261386 .
- ^ «Rhizosmilodon fiteae :: Музей естественной истории Флориды» . Архивировано из оригинала 4 октября 2016 г. Проверено 4 октября 2016 г.
- ^ Jump up to: а б с д Антон, М.; MJ Салеса; Дж. Моралес; А. Тернер (2004). «Первые известные полные черепа ятаганообразной кошки Machairodus aphanistus (Felidae, Carnivora) из испанской стоянки Баталлонес-1 позднего миоцена». Журнал палеонтологии позвоночных . 24 (4): 957–969. doi : 10.1671/0272-4634(2004)024[0957:FKCSOT]2.0.CO;2 . S2CID 85985476 .
- ^ Jump up to: а б с д и Салеса, MJ; М. Антон; А. Тернер; Дж. Моралес (2005). «Аспекты функциональной морфологии черепного и шейного скелета саблезубой кошки Paramachairodus ogygia (Kaup, 1832) (Felidae, Machairodontinae) из позднего миоцена Испании: значение для происхождения смертельного укуса махайродонта» . Зоологический журнал Линнеевского общества . 144 (3): 363–377. дои : 10.1111/j.1096-3642.2005.00174.x .
- ^ Доусон, MR; РК Стаки; Л. Кришталка; СиСи Блэк (1986). «Machaeroides simpsoni, новый вид, старейший из известных саблезубых кредонтов (Mammalia) из затерянной хижины эоцена». Вклад в геологию, Университет Вайоминга, специальный доклад . 3 : 177–182.
- ^ Салеса, MJ; М. Антон; А. Тернер; Л. Алькала; П. Монтойя; Дж. Моралес (2010). «Систематический пересмотр позднемиоценовых саблезубых кошачьих Paramachaedrodus в Испании». Палеонтология . 53 (6): 1369–1391. Бибкод : 2010Palgy..53.1369S . дои : 10.1111/j.1475-4983.2010.01013.x . S2CID 83268098 .
- ^ Тернер, А. (1997). Большие кошки и их ископаемые родственники: иллюстрированный путеводитель по их эволюции и естественной истории . Нью-Йорк: Издательство Колумбийского университета.
- ^ Ван Валкенбург, Б.; Ф. Гертель (1993). «Тяжелые времена в Ла-Бреа – поломка зубов у крупных хищников позднего плейстоцена». Наука . 261 (5120): 456–459. Бибкод : 1993Sci...261..456V . дои : 10.1126/science.261.5120.456 . ПМИД 17770024 . S2CID 39657617 .
- ^ Ван Валкенбург, Б. (2009). «Издержки хищничества: перелом зубов у плейстоценовых и современных хищников» . Биологический журнал Линнеевского общества . 96 : 68–81. дои : 10.1111/j.1095-8312.2008.01108.x .
- ^ Jump up to: а б Салеса, MJ; М. Антон; А. Тернер; Дж. Моралес (2010). «Функциональная анатомия передней конечности Promegantereon ogygia (Felidae, Machairodontinae, Smilodontini) из позднего миоцена Испании и истоки модели саблезубых кошачьих» . Журнал анатомии . 216 (3): 381–396. дои : 10.1111/j.1469-7580.2009.01178.x . ПМЦ 2829396 . ПМИД 20039979 .
- ^ Jump up to: а б с д и ж г час я Тернер, Алан (1997). Большие кошки и их ископаемые родственники . Издательство Колумбийского университета.
- ^ Кристиансен, Пер (2008). «Эволюция формы черепа и нижней челюсти у кошек (Carnivora: Felidae)» . ПЛОС ОДИН . 3 (7): e2807. Бибкод : 2008PLoSO...3.2807C . дои : 10.1371/journal.pone.0002807 . ПМЦ 2475670 . ПМИД 18665225 .
- ^ Ладен, Грег. «Блог Грега Ладена: О проекте» . Архивировано из оригинала 13 июня 2011 года.
- ^ Jump up to: а б Горе, Стивен; Лоури, Антон (4 мая 2006 г.). «Как построить суперхищника млекопитающих». Зоология . 111 (3): 196–203. дои : 10.1016/j.zool.2007.07.008 . ПМИД 18313908 .
- ^ Антон, Маурисио; Галобарт, Ангел; Тернер, Алан (1 мая 2005 г.). «Сосуществование ятаганозубых кошек, львов и гомининов в европейском плейстоцене. Значение посткраниальной анатомии Homotherium latidens (Оуэн) для сравнительной палеоэкологии» . Четвертичные научные обзоры . 24 (10): 1287–1301. Бибкод : 2005QSRv...24.1287A . doi : 10.1016/j.quascirev.2004.09.008 . ISSN 0277-3791 .
- ^ Jump up to: а б Антон, Морис (2013). Саблезубый . п. 136.
- ^ Jump up to: а б Антон, Маурисио (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. стр. 227–228. ISBN 9780253010421 .
- ^ Jump up to: а б Брайнер, Жанна (октябрь 2007 г.). «Саблезубый кот укусил слабака» . Живая наука . Архивировано из оригинала 12 декабря 2011 г. Проверено 5 июля 2011 г.
- ^ «Стабильные изотопы в археологии» . Архивировано из оригинала 27 декабря 2013 г. Проверено 10 сентября 2011 г.
- ^ Р. Х. Тыкот. «Стабильные изотопы и диета: вы то, что вы едите» (PDF) . Архивировано (PDF) из оригинала 2 апреля 2012 г. Проверено 10 сентября 2011 г.
- ^ Домингес-Родриго, М., Эгеланд, К.П., Кобо-Санчес, Л. и др. Поведение потребления туш саблезубых особей и динамика плейстоценовых крупных гильдий хищников. Sci Rep 12, 6045 (2022 г.). https://doi.org/10.1038/s41598-022-09480-7
- ^ Jump up to: а б с Антон, М.; Гарсиа-Переа, Р.; Тернер, А. (1998). «Реконструированный облик лица саблезубого кошачьего смилодона» . Зоологический журнал Линнеевского общества . 124 (4): 369–386. дои : 10.1111/j.1096-3642.1998.tb00582.x .
- ^ Антон, Маурисио (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. стр. 168–173. ISBN 9780253010421 .
- ^ Абдулла, Сара (28 января 1999 г.). «Улыбка смилодона» . Природа . дои : 10.1038/news990128-5 . Архивировано из оригинала 5 ноября 2012 г. Проверено 5 июля 2011 г.
- ^ Антон, Маурисио; Силисео, Хема; Пастор Хуан Ф.; Салеса, Мануэль Дж. (2022). «Скрытое оружие: пересмотренная реконструкция анатомии лица и жизненного облика саблезубой кошки Homotherium latidens (Felidae, Machairodontinae)» . Четвертичные научные обзоры . 284 : 107471. Бибкод : 2022QSRv..28407471A . doi : 10.1016/j.quascirev.2022.107471 . hdl : 10261/270770 . S2CID 248168629 .
- ^ Jump up to: а б Местел, Рози (1 апреля 1993 г.). «Саблезубые сказки» . Откройте для себя журнал . Архивировано из оригинала 25 декабря 2011 г. Проверено 15 июля 2011 г.
- ^ «Скульптура саблезубого кота» . Архивировано из оригинала 9 января 2012 г. Проверено 10 сентября 2011 г.
- ^ Jump up to: а б Карбоне, К.; Мэддокс, Т.; Фанстон, П.Дж.; Миллс, MGL; Гретер, Г.Ф; Ван Валкенбург, Б. (23 февраля 2009 г.). «Параллели между воспроизведением и просачиванием смолы плейстоцена позволяют предположить социальность вымершего саблезубого кота Смилодона» . Письма по биологии . 5 (1): 81–85. дои : 10.1098/rsbl.2008.0526 . ПМЦ 2657756 . ПМИД 18957359 .
- ^ Меткалф, Джессика З. «Климат позднего плейстоцена и палеоэкология хоботных» . Архивировано из оригинала 19 марта 2012 г. Проверено 10 сентября 2011 г.
- ^ Антон, Маурисио (2013). Саблезубый . Блумингтон, Индиана: Издательство Университета Индианы. п. 29. ISBN 9780253010421 .
- ^ «Питание саблезубых кошек» . 08 марта 2008 г. Архивировано из оригинала 14 марта 2017 г. Проверено 14 марта 2017 г.
- ^ «Саблезубый кот, китайский Machairodus giganteus череп » . Архивировано из оригинала 26 апреля 2011 г. Проверено 3 июля 2011 г.
- ^ Шермис, Стюарт (1983). «Исцеление массивного перелома таза у смилодона из Ранко Ла Бреа, Калифорния» (PDF) . ПалеоБиос . 1 (3): 12–126. Архивировано из оригинала (PDF) 28 марта 2012 г.
- ^ МакКолл, Шерман; Неаполь, Вирджиния; Мартин, Ларри (2003). «Оценка поведения вымерших животных: был ли Смилодон социальным?» . Мозговое поведение. Эвол . 61 (3): 159–64. дои : 10.1159/000069752 . ПМИД 12697957 . S2CID 2756104 .
- ^ Суитек, Брайан. «Пересмотренный репост: Какие у тебя большие зубы» . Научные блоги. Архивировано из оригинала 21 февраля 2014 г. Проверено 5 октября 2012 г.
- ^ Андерссон, К.; Норман, Д.; Верделин, Л. (2011). Соарес, Дафна (ред.). «Саблезубые хищники и убийство крупной добычи» . ПЛОС ОДИН . 6 (10): e24971. Бибкод : 2011PLoSO...624971A . дои : 10.1371/journal.pone.0024971 . ПМК 3198467 . ПМИД 22039403 .
- ^ МакГенри, ЧР; Роу, С.; Клаузен, PD; Морено, К.; Каннингем, Э. (2007). «Супермоделирование саблезубого кота, хищническое поведение Smilodon Fatalis, выявленное с помощью компьютерного 3D-моделирования с высоким разрешением» . Труды Национальной академии наук . 104 (41): 16010–16015. Бибкод : 2007PNAS..10416010M . дои : 10.1073/pnas.0706086104 . ПМК 2042153 . ПМИД 17911253 .
- ^ «Ксеносмилюс ходсонае» . Музей Флориды. 3 апреля 2017 г.
- ^ Пейдж, Джейк. Слышат ли кошки ногами?: Откуда берутся кошки, что мы о них знаем . [ нужна полная цитата ]
- ^ «Саблезубые коты» . Архивировано из оригинала 6 октября 2011 г. Проверено 10 сентября 2011 г.
- ^ «Доисторические хищники: Саблезубый зуб. Часть 4» . Ютуб . Архивировано из оригинала 22 мая 2016 г. Проверено 26 ноября 2016 г.
- ^ Кваммен, Дэвид (2003). Божье чудовище: хищник-людоед в джунглях истории и разума . WW Нортон и компания. ISBN 9780393051407 .
- ^ Гордер, ПФ (2004). «Имитация следов укусов». Вычисления в науке и технике . 6 (3): 4–6. Бибкод : 2004CSE.....6c...4G . дои : 10.1109/MCISE.2004.1289301 . S2CID 515458 .
- Отчет об исследовании группы Барнетта в журнале Current Biology , 9 августа 2005 г .: Росс Барнетт и др.: «Эволюция вымерших саблезубых и американской гепардоподобной кошки» в журнале Current Biology , Vol. 15, R589-R590, 9 августа 2005 г.
Дальнейшее чтение
[ редактировать ]- Ван Валкенбург, Б. (2007). «Дежавю: эволюция морфологии питания хищных животных» . Интегративная и сравнительная биология . 47 (1): 147–163. дои : 10.1093/icb/icm016 . ПМИД 21672827 . . Полный анализ конвергентной эволюции гиперплотоядных животных
Внешние ссылки
[ редактировать ]Диаграммы
- Эволюция кошачьей формы саблезубого черепа в блоге Нимравида.
- Схемы саблезубого черепа
- Диаграммы Марисио Антона. Все графитовые рисунки принадлежат Антону, а также нескольким другим художникам. Предпоследний рисунок изображает универсальность общего прикуса шеи и включает сравнение Machairodus и Panthera leo . головы и лица
губы Миллера
- Иллюстрации губ Миллера в фазах агрессии и неагрессии. (Обязательно нажмите в любом месте страницы сразу после входа — это слайд-шоу, и переход к следующему изображению осуществляется через 5 секунд.)
- художником-биологом Марисио Антоном Изображение Махайродуса без губ Миллера, сделанное . (это зарубежный сайт — изображение расположено на две трети вниз с надписью « Изображение: Маурисио Антон» )
- Дальнейшая иллюстрация (Антон) отсутствия губ Миллера у рода Homotherium.
| Гомотерини |  | |
|---|---|---|
| Метаюрины | ||
| Смилодонтини | ||
| Неопределенное место: | ||
| Базы данных органов управления : Национальные |
|---|