Скажи волк
| Скажи волк Временной диапазон: | |
|---|---|
 | |
| Установленный скелет, Музей естественной истории Штернберга. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Плотоядный |
| Семья: | Псовые |
| Подсемейство: | Клыки |
| Племя: | Клыки |
| Подтриба: | Собачий |
| Род: | † Эноцион Мерриам , 1918 год. [2] |
| Разновидность: | † А. жесткий |
| Биномиальное имя | |
| † Эноцион дирус | |
| Подвиды [3] | |
| Синонимы | |
Ужасный волк ( Aenocyon dirus) [10] / iː ˈ n ɒ s aɪ . ɒ n ˈ d aɪ r ə s / ) — вымершая собака . Ужасный волк жил в Америке (возможно, единственная находка также известна из Восточной Азии ) в эпохи позднего плейстоцена и раннего голоцена (125 000–9 500 лет назад). Вид был назван в 1858 году, через четыре года после того, как был найден первый экземпляр. два подвида Выделяют : Aenocyon dirus guildayi и Aenocyon dirus dirus . Самая большая коллекция его окаменелостей была получена в смоляных ямах Ранчо Ла Бреа в Лос-Анджелесе .
Останки ужасного волка были обнаружены в самых разных средах обитания, включая равнины, луга и некоторые лесные горные районы Северной Америки, а также засушливые саванны Южной Америки. Высота участков варьируется от уровня моря до 2255 метров (7400 футов). Окаменелости ужасных волков редко находили к северу от 42° северной широты ; над этой широтой было только пять неподтвержденных сообщений. Считается, что это ограничение ареала связано с ограничениями температуры, добычи или среды обитания, вызванными близостью к Лаврентиды и Кордильер существовавшим в то время ледниковым щитам .
Ужасный волк был примерно такого же размера, как самые крупные современные серые волки ( Canis lupus ): юконский волк и северо-западный волк . А. д. guildayi весил в среднем 60 кг (132 фунта), а A. d. Дирус составлял в среднем 68 кг (150 фунтов). Его череп и зубные ряды соответствовали таковым у C. lupus , но зубы были крупнее и обладали большей режущей способностью, а сила укуса клыка была сильнее, чем у любого известного Canis вида . Считается, что эти характеристики являются адаптацией для охоты на мегатравоядных животных позднего плейстоцена , а в Северной Америке его добычей, как известно, были западные лошади , наземные ленивцы , мастодонты , древние бизоны и верблюды . Его исчезновение произошло во время четвертичного вымирания вместе с его основными видами добычи. Его зависимость от мегатравоядных животных была предложена как причина его вымирания, а также изменение климата и конкуренция с другими видами или комбинация этих факторов. Согласно датированным останкам, ужасные волки жили всего 9500 лет назад.
Таксономия
[ редактировать ]С 1850-х годов в Соединенных Штатах находили ископаемые останки вымерших крупных волков, и не сразу было ясно, принадлежат ли они к одному виду. Первый экземпляр того, что позже стало ассоциироваться с Aenocyon dirus, был найден в середине 1854 года в русле реки Огайо недалеко от Эвансвилля, штат Индиана . Окаменевшая челюстная кость с щечными зубами была получена геологом Джозефом Грэнвиллом Норвудом у коллекционера из Эвансвилля Фрэнсиса А. Линка. Палеонтолог Джозеф Лейди определил, что этот экземпляр представляет собой вымерший вид волка, и назвал его Canis primaevus . [4] Письма Норвуда Лейди хранятся вместе с типовым экземпляром (первым представителем вида, имеющим письменное описание) в Академии естественных наук Филадельфии . В 1857 году, исследуя долину реки Ниобрара в Небраске, Лейди нашел позвонки вымершего вида Canis , о которых он сообщил в следующем году под названием C. dirus . [1] Название C. primaevus (Leidy 1854) позже было переименовано в Canis indianensis (Leidy 1869), когда Лейди узнала, что название C. primaevus ранее использовалось британским натуралистом Брайаном Хоутоном Ходжсоном для обозначения волка . [5]

В 1876 году зоолог Джоэл Асаф Аллен обнаружил останки Canis Mississippiensis (Allen 1876) и связал их с C. dirus (Leidy 1858) и Canis indianensis (Leidy 1869). Поскольку этих трех экземпляров было найдено так мало, Аллен счел, что лучше оставить каждый экземпляр в списке под его предварительным названием, пока не будет найдено больше материала, раскрывающего их родство. [6] В 1908 году палеонтолог Джон Кэмпбелл Мерриам начал извлекать многочисленные окаменелые фрагменты костей большого волка из смоляных ям Ранчо Ла Бреа. К 1912 году он нашел достаточно полный скелет, чтобы можно было официально распознать эти и ранее найденные экземпляры под названием C. dirus (Leidy 1858). Поскольку правила номенклатуры предусматривали, что название вида должно быть самым старым названием, когда-либо применявшимся к нему. [12] Поэтому Мерриам выбрал название образца Лейди 1858 года — C. dirus . [13] В 1915 году палеонтолог Эдвард Трокселл выразил свое согласие с Мерриамом, когда объявил C. indianensis синонимом C. dirus . [14] В 1918 году, после изучения этих окаменелостей, Мерриам предложил объединить их названия в отдельный род Aenocyon (от ainos , «ужасный» и cyon , «собака»), чтобы стать Aenocyon dirus , [2] но в то время не все были согласны с тем, чтобы этого вымершего волка поместили в новый род, отдельный от рода Canis . [15] Canis ayersi (Sellards 1916) и Aenocyon dirus (Merriam 1918) были признаны синонимами C. dirus палеонтологом Эрнестом Лунделиусом в 1972 году. [16] все вышеперечисленные таксоны были объявлены синонимами C. dirus в 1979 году. По словам палеонтолога Рональда М. Новака, [17]
В 1984 году исследование Бьорна Куртена выявило географическое разнообразие популяций лютого волка и предложило два подвида: Canis dirus guildayi (названный Куртеном в честь палеонтолога Джона Э. Гилдея) для экземпляров из Калифорнии и Мексики, у которых были более короткие конечности и более длинные. зубы и Canis dirus dirus для экземпляров к востоку от Североамериканского континентального водораздела , у которых были более длинные конечности и более короткие зубы. [3] [18] [19] [20] Куртен назвал верхнюю челюсть, найденную в Пещере Отшельника, штат Нью-Мексико, представителем номинального подвида C. d. дирус . [3]
В 2021 году исследование ДНК показало, что лютоволк представляет собой сильно отличающуюся линию по сравнению с современными волкоподобными собаками , и этот вывод согласуется с ранее предложенной таксономической классификацией лютого волка как рода Aenocyon (древнегреческий: «ужасный волк»). волк»), предложенный Мерриамом в 1918 году. [21]
Эволюция
[ редактировать ]В Северной Америке семейство псовых возникло 40 миллионов лет назад. [22] [23] и подсемейство собак Caninae около 32 миллионов лет назад. [24] предки лисьеподобных Vulpini и собакообразных Canini От Caninae 9 миллионов лет назад произошли . Эта группа была впервые представлена Эвционом , и в основном койотоподобным Эвционом дависи , широко распространённым по Северной Америке. [25] От Canini 6–5 миллионов лет назад возникла Cerdocyonina , сегодня представленная южноамериканскими псовыми. [26] Его сестра, волкообразная Canina, появилась на свет 5 миллионов лет назад, однако, вероятно, они возникли еще 9 миллионов лет назад. [25] Около 7 миллионов лет назад представители рода Canis распространились в Евразию и Африку, а Эвцион дал начало первому представителю рода Canis в Европе. [27] Около 4–3 миллионов лет назад C. chihliensis , первый представитель семейства Canis размером с волка , возник в Китае и распространился, дав начало другим волкоподобным представителям по всей Евразии и Африке. Представители рода Canis позже распространились в Северную Америку. [26]
Ужасный волк эволюционировал в Северной Америке. [26] [21] Однако его наследственное происхождение обсуждается, существуют две конкурирующие теории. Первая теория основана на морфологии ископаемых , которая указывает на то, что распространение рода Canis за пределы Евразии привело к появлению лютого волка. [26] Вторая теория основана на данных ДНК, которые указывают на то, что ужасный волк произошел от наследственной линии, зародившейся в Америке и отделенной от рода Canis . [21]
Морфологические доказательства
[ редактировать ]| Дивергенция ужасных волков по морфологии | ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Эволюционное расхождение лютого волка на основе морфологии [26] [28] |
Морфологические данные, основанные на ископаемых останках, указывают на то, что распространение рода Canis за пределы Евразии привело к появлению ужасного волка. [26] [28]
В 1974 году Роберт А. Мартин предположил, что крупный североамериканский волк C.armbrusteri (волк Армбрустера) был C. lupus . [29] Новак, Куртен и Анналиса Берта предположили, что C. dirus не произошел от C. lupus . [17] [30] [31] В 1987 году была выдвинута новая гипотеза, согласно которой популяция млекопитающих могла дать начало более крупной форме, называемой гиперморфом, в периоды, когда еды было в изобилии, но когда еды позже стало не хватать, гиперморф либо адаптировался к меньшей форме, либо вымер. Эта гипотеза может объяснить большие размеры тела, обнаруженные у многих млекопитающих позднего плейстоцена по сравнению с их современными собратьями. И вымирание, и видообразование – процесс, посредством которого новый вид отделяется от более старого – могут происходить одновременно в периоды экстремальных климатических явлений. [32] [33] Глория Д. Гуле согласилась с Мартином, предположив далее, что эта гипотеза может объяснить внезапное появление C. dirus в Северной Америке и, судя по сходству в форме их черепов, что C. lupus дал начало C. dirus гиперморфу из-за к обилию дичи, стабильной среде и крупным конкурентам. [34]
Трое палеонтологов Сяомин Ван , Ричард Х. Тедфорд и Рональд М. Новак предполагают, что C. dirus произошел от Canis Armbrusteri . [26] [28] при этом Новак заявляет, что оба вида возникли в Америке. [35] и что экземпляры, найденные в Камберлендской пещере, штат Мэриленд , по-видимому, представляют собой C. Armbrusteri, перешедшую в C. dirus . [36] [37] Новак полагал, что Canis edwardii был первым волком, появившимся в Северной Америке, и, по-видимому, он близок к линии, от которой произошли C.armbrusteri и C.dirus . [38] Тедфорд считает, что ранний волк из Китая, Canis chihliensis , возможно, был предком как C. Armbrusteri , так и серого волка C. lupus . [39] Внезапное появление C.armbrusteri в средних широтах Северной Америки в раннем плейстоцене 1,5 миллиона лет назад вместе с мамонтом позволяет предположить, что это был иммигрант из Азии, [28] Серый волк C. lupus появился в Берингии позже в плейстоцене и проник в средние широты Северной Америки во время последнего ледникового периода вместе со своей берингийской добычей. [26] [28] [37] В 2010 году Франсиско Превости предположил, что C. dirus является сестринским таксоном C. lupus . [40]
C. dirus жил в период от позднего плейстоцена до раннего голоцена , 125 000–10 000 лет назад ( за несколько лет до настоящего времени ), в Северной и Южной Америке. [3] Большинство окаменелостей из восточной части C. d. dirus датируются 125 000–75 000 YBP, но западные C. d. Окаменелости гульдайи не только меньше по размеру, но и более поздние; таким образом, было высказано предположение, что C. d. guildayi происходит от C. d. дирус . [3] [20] Однако существуют спорные экземпляры C. dirus , возраст которых составляет 250 000 лет назад. Ископаемые экземпляры C. dirus, обнаруженные в четырех местах в районе Хей-Спрингс округа Шеридан, штат Небраска , были названы Aenocyon dirus nebrascensis (Frick 1930, неописанный), но Фрик не опубликовал их описания. Позже Новак назвал этот материал C.armbrusteri ; [41] затем, в 2009 году, Тедфорд официально опубликовал описание образцов и отметил, что, хотя они демонстрировали некоторые морфологические характеристики как C. Armbrusteri , так и C. dirus , он называл их только C. dirus . [39]
Ископаемое, обнаруженное в Конном зале пещеры Саламандр в Блэк-Хилс в Южной Дакоте, возможно, может быть C. dirus ; если это так, то эта окаменелость является одним из самых ранних зарегистрированных экземпляров. [19] [42] Он был внесен в каталог как Canis cf. С. дирус [43] (где ср. по латыни означает совещаться, неуверенный). Окаменелость лошади, найденная в Конном зале, дает урановой серии датировку по 252 000 лет назад , а Canis cf. dirus относится к тому же периоду. Предполагалось, что экземпляр [19] [43] C.armbrusteri и C.dirus имеют некоторые общие характеристики ( синапоморфии ), которые предполагают происхождение последнего от первого. Летопись окаменелостей предполагает, что C. dirus возник около 250 000 лет назад на открытой местности среднего континента, а затем распространился на восток и вытеснил своего предка C. Armbrusteri . [28] Таким образом, первое появление C. dirus составит 250 000 YBP в Калифорнии и Небраске, а затем и в остальной части США, Канаде, Мексике, Венесуэле, Эквадоре, Боливии и Перу. [39] но идентичность этих самых ранних окаменелостей не подтверждена. [44]
В Южной Америке экземпляры C. dirus, датированные поздним плейстоценом, были обнаружены вдоль северного и западного побережья, но ни один из них не был обнаружен в Аргентине, районе, где обитали Canis gezi и Canis nehringi . [39] Учитывая их сходство и временные рамки, предполагается, что C. gezi был предком Canis nehringi . Одно исследование показало, что C. dirus произошел более эволюционно по сравнению с C. nehringi и имел больший размер и конструкцию нижних коренных зубов для более эффективного хищничества. [45] По этой причине некоторые исследователи предположили, что C. dirus мог возникнуть в Южной Америке. [46] [19] [31] Тедфорд предположил, что C.armbrusteri был общим предком как северных, так и южноамериканских волков. [39] Более поздние исследования пришли к выводу, что C. dirus и C. nehringi были одним и тем же видом. [40] [47] и что C. dirus мигрировал из Северной Америки в Южную Америку, что сделало его участником Великого американского обмена . [40] В 2018 году исследование показало, что Canis gezi не относится к роду Canis и должен быть отнесен к подтрибе Cerdocyonina , однако род не был предложен. [47]
Обнаружение в 2020 году предполагаемой окаменелости лютого волка на северо-востоке Китая указывает на то, что ужасные волки, возможно, пересекали Берингию , когда она существовала. [48]
ДНК-доказательства
[ редактировать ]| Кладограмма, показывающая взаимоотношения между живыми и вымершими волкообразными псовыми на основе ДНК [примечание 1] |
| На основе данных яДНК, указывающих на то, что ужасный волк разветвился 5,7 миллиона лет назад. [21] |




Данные ДНК указывают на то, что ужасный волк произошел от наследственной линии, которая возникла в Америке и была отделена от рода Canis . [21]
В 1992 г. была предпринята попытка извлечь последовательность митохондриальной ДНК из скелетных останков A. d. guildayi, чтобы сравнить его родство с другими видами Canis . Попытка не увенчалась успехом, поскольку эти останки были извлечены из ям Ла Бреа, и смолу невозможно было удалить из костного материала. [51] В 2014 году попытка извлечь ДНК колумбийского мамонта из смоляных ям также потерпела неудачу: исследование пришло к выводу, что органические соединения асфальта пронизывают кости всех древних образцов из ям Ла Бреа, что затрудняет извлечение образцов ДНК. [52]
В 2021 году исследователи секвенировали ядерную ДНК (из ядра клетки), взятую из пяти окаменелостей ужасных волков, датируемых 13 000–50 000 лет назад. Последовательности показывают, что ужасный волк представляет собой сильно разошедшуюся линию, которая в последний раз имела общего предка с волкоподобными собаками 5,7 миллиона лет назад. В ходе исследования также были измерены многочисленные образцы скелетов лютого волка и серого волка, которые показали, что их морфология очень схожа, что привело к теории о том, что люто-волк и серый волк имели тесное эволюционное родство. Был сделан вывод, что морфологическое сходство между лютыми волками и серыми волками обусловлено конвергентной эволюцией . Известно, что представители волкообразных собак гибридизуются друг с другом, но исследование не смогло обнаружить никаких признаков генетического смешения пяти образцов ужасных волков с современными североамериканскими серыми волками и койотами, а также их общим предком. Это открытие указывает на то, что линии волков и койотов развивались изолированно от линии ужасных волков. [21]
Исследование предполагает раннее происхождение линии ужасных волков в Америке и что эта географическая изоляция позволила им развить определенную степень репродуктивной изоляции с момента их расхождения 5,7 миллиона лет назад. Койоты, волки, серые волки и вымерший ксеноцион эволюционировали в Евразии и сравнительно недавно, в позднем плейстоцене, распространились в Северную Америку, поэтому смешения с лютым волком не произошло. Длительная изоляция линии лютого волка подразумевает, что другие американские ископаемые таксоны, в том числе C. Armbrusteri и C. edwardii , также могут принадлежать к линии лютого волка. Результаты исследования согласуются с ранее предложенной таксономической классификацией лютого волка как рода Aenocyon . [21]
Радиоуглеродное датирование
[ редактировать ]Возраст большинства местонахождений лютоволков определяется исключительно биостратиграфией , но биостратиграфия является ненадежным индикатором в пределах асфальтовых отложений. [53] [54] Некоторые участки были датированы радиоуглеродом , при этом образцы ужасных волков из ям Ла Бреа были датированы следующими календарными годами: 82 экземпляра датированы 13 000–14 000 лет назад; 40 экз. возрастом 14–16 тыс . лет назад; 77 экз. возрастом 14–18 тыс . лет назад; 37 экз. возрастом 17–18 тыс . лет назад; 26 экз. возрастом 21–30 тыс . лет назад; 40 экз. возрастом 25–28 тыс . лет назад; и 6 экз. возрастом 32–37 тыс. лет назад. [44] : Т1 Образец из пещеры Паудер-Милл-Крик, штат Миссури, был датирован 13 170 лет назад. [19]
Описание
[ редактировать ]
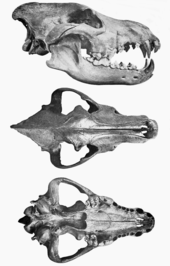
Средние пропорции ужасных волков были аналогичны пропорциям двух современных североамериканских волков: юконского волка ( Canis lupus pambasileus ). [55] [13] и северо-западный волк ( Canis lupus occidentalis ). [55] Самые крупные северные волки сегодня имеют высоту в плечах до 38 дюймов (97 см) и длину тела 69 дюймов (180 см). [56] : 1 Некоторые экземпляры ужасных волков с Ранчо Ла Бреа меньше этого, а некоторые больше. [13]
У ужасного волка были меньшие ступни и большая голова по сравнению с северным волком того же размера тела. Длина черепа могла достигать 310 мм (12 дюймов) и более, с более широким небом , лобной областью и скуловыми дугами по сравнению с юконским волком. Такие размеры делают череп очень массивным. Его сагиттальный гребень был выше, передняя часть носа значительно выступала назад, а задние концы носовых костей заходили относительно далеко назад в череп. Связанный скелет лютого волка с ранчо Ла Бреа найти сложно, потому что смола позволяет костям разбираться во многих направлениях. Были собраны части позвоночного столба , и выяснилось, что он похож на позвоночник современного волка с таким же количеством позвонков. [13]
Географические различия у ужасных волков не были обнаружены до 1984 года, когда исследование останков скелетов показало различия в некоторых черепно-зубных особенностях и пропорциях конечностей между экземплярами из Калифорнии и Мексики ( A.d. ) и экземплярами , guildayi найденными с востока Континентальный водораздел ( A. d. dirus ). Сравнение размеров конечностей показывает, что задние конечности A. d. guildayi были на 8% короче юконского волка из-за значительно более коротких голеней и плюсен , а также из-за того, что передние конечности были короче из-за их немного более коротких нижних костей. [57] [58] Обладая сравнительно более легкими и меньшими конечностями и массивной головой, A. d. гильдайи не были так хорошо приспособлены к бегу, как лесные волки и койоты. [58] [13] А. д. dirus обладал значительно более длинными конечностями, чем A. d. гильдайи . Передние конечности были на 14% длиннее, чем у A. d. guildayi за счет длины плечевых костей на 10% , лучевых костей на 15% и пястных костей на 15% . Задние конечности были на 10% длиннее, чем у A. d. guildayi и голеней на 10% из-за длиннее бедер и плюсневых костей на 15% . А. д. dirus по длине конечностей сравним с юконским волком. [57] Самый А.д. крупный dirus femur был найден в Кэрролл-Кейв, штат Миссури, и имел размер 278 мм (10,9 дюйма). [20]

| Переменная конечности | А. д. гильдайи [58] | Юконский волк [58] | ОБЪЯВЛЕНИЕ. дирус [57] |
|---|---|---|---|
| Плечевая кость (верхняя часть передней ноги) | 218 мм (8,6 дюйма) | 237 мм (9,3 дюйма) | 240 мм (9,4 дюйма) |
| Радиус (нижняя часть передней ноги) | 209 мм (8,2 дюйма) | 232 мм (9,1 дюйма) | 240 мм (9,4 дюйма) |
| Пястная кость (передняя стопа) | 88 мм (3,4 дюйма) | 101 мм (4,0 дюйма) | 101 мм (4,0 дюйма) |
| Бедренная кость (верхняя часть задней части ноги) | 242 мм (9,5 дюйма) | 251 мм (9,9 дюйма) | 266 мм (10,5 дюйма) |
| Большеберцовая кость (нижняя часть задней ноги) | 232 мм (9,1 дюйма) | 258 мм (10,2 дюйма) | 255 мм (10,0 дюйма) |
| Плюсневая кость (задняя стопа) | 93 мм (3,7 дюйма) | 109 мм (4,3 дюйма) | 107 мм (4,2 дюйма) |
А. д. guildayi , по оценкам, весил в среднем 60 кг (132 фунта), а A. d. dirus весил в среднем 68 кг (150 фунтов), некоторые экземпляры были крупнее, [20] но они не могли превышать 110 кг (243 фунта) из-за ограничений скелета. [59] Для сравнения, средний вес юконского волка составляет 43 кг (95 фунтов) для самцов и 37 кг (82 фунта) для самок. Индивидуальный вес юконских волков может варьироваться от 21 кг (46 фунтов) до 55 кг (121 фунт). [60] с одним юконским волком весом 79,4 кг (175 фунтов). [56] : 1 Эти цифры показывают, что средний лютоволк по размеру аналогичен самому большому современному серому волку. [20]
Останки полного самца A. dirus иногда легко идентифицировать по сравнению с другими экземплярами Canis, поскольку бакулюм (кость полового члена) лютого волка сильно отличается от таковой у всех других ныне живущих псовых . [19] [57]
Приспособление
[ редактировать ]
Было показано, что экологические факторы, такие как тип среды обитания, климат, специализация добычи и конкуренция хищников, сильно влияют на краниодентальную пластичность серого волка , которая представляет собой адаптацию черепа и зубов под воздействием окружающей среды. [62] [63] [64] Точно так же ужасный волк был гиперплотоядным животным с черепом и зубными рядами, приспособленными для охоты на крупную и борющуюся добычу; [65] [66] [67] форма его черепа и морды со временем менялась, а изменения размера его тела коррелировали с колебаниями климата. [68]
Палеоэкология
[ редактировать ]Последний ледниковый период , обычно называемый «Ледниковым периодом», длился 125 000 лет. [69] –14 500 йенских фунтов [70] и был самым последним ледниковым периодом нынешнего ледникового периода , который произошел в последние годы плейстоценовой эры. [69] Ледниковый период достиг своего пика во время последнего ледникового максимума , когда ледниковые щиты начали наступать с 33 000 YBP и достигли максимального предела в 26 500 YBP. Дегляциация началась в Северном полушарии примерно в 19 000 лет назад, а в Антарктиде примерно в 14 500 лет назад, что согласуется с данными о том, что талая ледниковая вода была основным источником резкого повышения уровня моря (14 500 лет назад). [70] Доступ в северную часть Северной Америки был заблокирован Висконсинским оледенением . Ископаемые свидетельства из Америки указывают на вымирание в основном крупных животных, называемых плейстоценовой мегафауной , ближе к концу последнего оледенения. [71]
Прибрежная южная Калифорния с 60 000 YBP до конца последнего ледникового максимума была более прохладной и с более сбалансированным запасом влаги, чем сегодня. Во время последнего ледникового максимума средняя годовая температура снизилась с 11 ° C (52 ° F) до 5 ° C (41 ° F) градусов, а годовое количество осадков уменьшилось со 100 см (39 дюймов) до 45 см (18 в). [72] Этот регион не пострадал от климатических последствий Висконсинского оледенения и, как полагают, был убежищем ледникового периода для животных и чувствительных к холоду растений. [73] [74] [75] К 24 000 лет назад обилие дуба и чапараля уменьшилось, но увеличилось количество сосен, образовав открытые парковые зоны, похожие на сегодняшние прибрежные горные /можжевеловые леса. После 14 000 лет назад численность хвойных пород уменьшилась, а численность современных прибрежных растительных сообществ, включая дубовые леса, чапараль и кустарники прибрежного шалфея , увеличилась. Равнина Санта-Моника расположена к северу от города Санта-Моника и простирается вдоль южного подножия гор Санта-Моника , и на 28 000–26 000 лет назад на ней преобладали прибрежные заросли шалфея, а на возвышенностях - кипарисы и сосны. Горы Санта-Моники поддерживали сообщество чапараля на своих склонах и изолировали прибрежные секвойи и кизил в защищенных каньонах, а также речные сообщества, включавшие иву, красный кедр и платан. Эти растительные сообщества предполагают зимние осадки, аналогичные тем, которые наблюдаются в современной прибрежной южной Калифорнии, но присутствие прибрежных секвой, обнаруженных сейчас в 600 километрах (370 миль) к северу, указывает на более прохладный, влажный и менее сезонный климат, чем сегодня. Эта среда поддерживала крупных травоядных животных, которые были добычей ужасных волков и их конкурентов. [72]
Добыча
[ редактировать ]
Ряд образцов животных и растений, которые попали в ловушку и затем были сохранены в смоляных ямах, были удалены и изучены, чтобы исследователи могли узнать о прошлом. Смолистые ямы Ранчо Ла Бреа, расположенные недалеко от Лос-Анджелеса в Южной Калифорнии, представляют собой совокупность ям с отложениями липкого асфальта, время отложения которых различается от 40 000 до 12 000 YBP. Начиная с 40 000 YBP, захваченный асфальт под давлением метана выносился на поверхность через трещины, образуя просачивания, которые могут занимать несколько квадратных метров и иметь глубину 9–11 м (30–36 футов). [53] Большое количество окаменелостей ужасных волков было обнаружено в смоляных ямах Ла Бреа. [26] Из смоляных ям было извлечено более 200 000 экземпляров (в основном фрагментов). [20] останки варьируются от смилодона до белок, беспозвоночных и растений. [53] Период времени, представленный на ямках, включает последний ледниковый максимум, когда глобальные температуры были на 8 ° C (14 ° F) ниже, чем сегодня, переход плейстоцен-голоцен ( Бёллинг-Аллерёд интервал ), старейшее похолодание дриаса , похолодание молодого дриаса от от 12 800 до 11 500 лет назад, а вымирание американской мегафауны - 12 700 лет назад, когда вымерло 90 родов млекопитающих весом более 44 кг (97 фунтов). [54] [68]
Изотопный анализ может быть использован для идентификации некоторых химических элементов, что позволит исследователям сделать выводы о рационе видов, обитающих в ямах. Изотопный анализ костного коллагена , выделенного из образцов Ла Бреа, свидетельствует о том, что ужасный волк Смилодон и американский лев ( Panthera atrox ) боролись за одну и ту же добычу. Их добычей были «вчерашний верблюд» ( Camelops hesternus ), плейстоценовый бизон ( Bison antiquus ), «карликовая» вилорогая ( Capromeryx major ), западная лошадь ( Equus occidentalis ) и аборигенный «пасущийся» наземный ленивец ( Paramylodon harlani ). на луга Северной Америки. Колумбийский мамонт ( Mammuthus columbi ) и американский мастодонт ( Mammut americanum ) были редкостью в Ла Бреа. Лошади по-прежнему питались смешанно, а вилороги - смешанно, но во время последнего ледникового максимума и связанного с ним изменения растительности верблюды и бизоны были вынуждены в большей степени полагаться на хвойные породы. [72]
Исследование изотопных данных окаменелостей лютого волка Ла-Бреа, датированных 10 000 лет назад, свидетельствует о том, что лошадь была важным видом добычи в то время и что ленивец, мастодонт, бизон и верблюд были менее распространены в рационе лютого волка. [65] [73] Это указывает на то, что ужасный волк не был специалистом по добыче и в конце позднего плейстоцена, перед своим исчезновением, он охотился или питался наиболее доступными травоядными животными. [73]
Зубной ряд и сила прикуса
[ редактировать ]
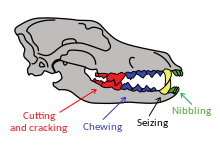
По сравнению с рода Canis зубным рядом представителей , ужасный волк считался наиболее эволюционно развитым (продвинутым) волкоподобным видом в Америке. Ужасного волка можно было отличить отдельно от всех других видов Canis по наличию: «P2 с задним бугорком; P3 с двумя задними бугорками; M1 с местаскилидом, энтокристом, энтоконулидом и поперечным гребнем, простирающимся от метаконида до гиперконуляра. полка М2 с энтокристедом и энтоконулидом». [30]
Исследование расчетной силы укуса клыков у большой выборки живых и ископаемых хищников-млекопитающих с поправкой на массу тела показало, что для плацентарных млекопитающих сила укуса клыков (в ньютонах на килограмм массы тела) составляла Самый крупный из них — ужасный волк (163), за ним среди современных псовых следуют четыре гиперплотоядных, которые часто охотятся на животных крупнее себя: африканская охотничья собака (142), серый волк (136), волк (112) и динго (108). Сила укуса хищников имела аналогичную тенденцию с собаками. На размер самой крупной добычи хищника сильно влияют его биомеханические ограничения. Морфология лютого волка была похожа на морфологию его ныне живущих родственников, и если предположить, что лютоволк был общественным охотником, то его высокая сила укуса по сравнению с современными псовыми позволяет предположить, что он охотился на относительно крупных животных. Оценка силы укуса пятнистой гиены, поедающей кости (117), бросила вызов распространенному предположению, что для поглощения костей необходима высокая сила укуса клыков и хищников. [67]
Исследование размеров черепа и челюстных мышц лютоволков не выявило существенных различий с современными серыми волками по всем показателям, кроме 4 из 15. Верхний зубной ряд был таким же, за исключением того, что лютоволк имел большие размеры, а у P4 было относительно большее и массивное лезвие, которое улучшало режущие способности хищника. Челюсть лютого волка имела относительно более широкую и массивную височную мышцу, способную генерировать немного большую силу укуса, чем у серого волка. Из-за расположения челюстей у лютого волка было меньше височных рычагов, чем у серого волка на нижнем хищническом (m1) и нижнем p4 уровне, но функциональное значение этого не известно. Нижние премоляры были относительно немного крупнее, чем у серого волка. [66] а лютоволк m1 был намного крупнее и обладал большей режущей способностью. [13] [31] [66] Клыки ужасного волка имели большую силу изгиба, чем у живых псовых того же размера, и были похожи на клыки гиен и кошачьих. [78] Все эти различия указывают на то, что лютоволк был способен наносить более сильные укусы, чем серый волк, и благодаря своим гибким и более закругленным клыкам был лучше приспособлен для борьбы с добычей. [65] [66]
| Зуб переменный | волчанка современная Североамериканский [79] | волчанка Ла Бреа [79] | Волчанка Берингия [79] | тяжело, тяжело Сангамонская эпоха [3] [65] (125 000–75 000 фунтов стерлингов) | тяжело, тяжело Поздний Висконсин [3] [65] (50 000 йен.) | Дирус Гильдайи [3] [65] (40 000–13 000 юаньских фунтов) |
|---|---|---|---|---|---|---|
| длина м1 | 28.2 | 28.9 | 29.6 | 36.1 | 35.2 | 33.3 |
| ширина м1 | 10.7 | 11.3 | 11.1 | 14.1 | 13.4 | 13.3 |
| m1 тригониды длина | 19.6 | 21.9 | 20.9 | 24.5 | 24.0 | 24.4 |
| длина п4 | 15.4 | 16.6 | 16.5 | 16.7 | 16.0 | 19.9 |
| ширина p4 | - | - | - | 10.1 | 9.6 | 10.3 |
| длина p2 | - | - | - | 15.7 | 14.8 | 15.7 |
| ширина р2 | - | - | - | 7.1 | 6.7 | 7.4 |
Поведение
[ редактировать ]В Ла-Бреа хищных птиц и млекопитающих привлекли мертвые или умирающие травоядные животные, которые застряли в болоте, а затем эти хищники сами оказались в ловушке. [53] [80] По оценкам, отлов травоядных происходил раз в пятьдесят лет. [80] и на каждый экземпляр останков травоядных животных, найденных в ямах, приходилось примерно десять хищников. [53] А. д. guildayi — самый распространенный хищник, встречающийся в Ла- Бреа, за ним следует смилодон . [54] [68] Останки ужасных волков в смоляных ямах превосходят по численности останки серых волков в соотношении пять к одному. [44] Считается, что во время последнего ледникового максимума прибрежная Калифорния с климатом немного прохладнее и влажнее, чем сегодня, была убежищем. [73] а сравнение частоты появления останков ужасных волков и других хищников в Ла- Бреа с другими частями Калифорнии и Северной Америки указывает на значительно большую численность; следовательно, более высокая численность ужасных волков в регионе Ла-Бреа не отражает более широкую территорию. [81] Если предположить, что лишь немногие из кормившихся хищников оказались в ловушке, вполне вероятно, что в этих случаях довольно значительные группы ужасных волков кормились вместе. [82]

Разница между самцом и самкой вида, за исключением их половых органов, называется половым диморфизмом , и в этом отношении среди псовых существует небольшая разница. Исследование останков лютого волка, датированных 15 360–14 310 лет назад и взятых из одной ямы, в ходе которого основное внимание уделялось длине черепа, размеру клыков и длине нижних коренных зубов, показало небольшой диморфизм, аналогичный диморфизму серого волка, что указывает на то, что ужасные волки жили моногамными парами. . [82] Их большой размер и очень плотоядные зубы подтверждают предположение, что ужасный волк был хищником, питавшимся крупной добычей. [82] [83] [84] Чтобы убить копытных, крупнее их самих, африканская дикая собака, волк и серый волк полагаются на свои челюсти, поскольку они не могут использовать передние конечности для борьбы с добычей, и работают вместе как стая, состоящая из альфа-пары и их потомков от текущий и предыдущие годы. Можно предположить, что лютые волки жили стаями сородичей, которых возглавляла альфа-пара. [82] Крупные и социальные хищники могли бы успешно защищать трупы добычи, застрявшие в смоляных ямах, от более мелких одиночных хищников и, следовательно, с большей вероятностью сами бы оказались в ловушке. Многие A.d. Останки guildayi и Smilodon , найденные в смоляных ямах, позволяют предположить, что оба были социальными хищниками. [81] [85]
Все социальные наземные хищники-млекопитающие охотятся в основном на наземных травоядных млекопитающих, масса тела которых аналогична совокупной массе членов социальной группы, нападающих на животное-жертву. [59] [86] Большой размер лютого волка обеспечивает предполагаемый размер добычи от 300 до 600 кг (от 660 до 1320 фунтов). [20] [83] [84] Анализ стабильных изотопов костей лютого волка свидетельствует о том, что они предпочитали потреблять жвачных животных, таких как бизоны, а не других травоядных животных, но переходили к другой добыче, когда еды становилось мало, и иногда питались выброшенными на берег китами вдоль тихоокеанского побережья, когда они были доступны. [20] [66] [87] Стая лесных волков может убить лося массой 500 кг (1100 фунтов), который является их любимой добычей. [20] [56] : 76 и вполне возможна стая ужасных волков, сбившая бизона. [20] Хотя некоторые исследования показали, что из-за поломки зубов лютоволк, должно быть, грыз кости и мог быть падальщиком, его широкое распространение и более изящные конечности лютоволка указывают на хищника. Как и современный серый волк, лютоволк, вероятно, использовал свои коренные зубы после хищения, чтобы получить доступ к костному мозгу, но больший размер лютоволка позволял ему разгрызать более крупные кости. [66]
Поломка зуба
[ редактировать ]
Поломка зубов связана с поведением хищника. [88] Исследование девяти современных хищников показало, что каждый четвертый взрослый человек ломал зубы, причем половина из этих поломок приходилась на клыки. Больше всего поломок произошло у пятнистой гиены, которая поедает всю добычу, включая кости; наименьшее количество поломок наблюдалось у африканской дикой собаки , а серый волк занимал промежуточное положение между ними. [89] [88] Поедание кости увеличивает риск случайного перелома из-за относительно высоких и непредсказуемых напряжений, которые она создает. Чаще всего ломаются клыки, за ними следуют премоляры, хищные коренные зубы и резцы. Клыки – это зубы, которые чаще всего ломаются из-за своей формы и функции, что подвергает их изгибающим нагрузкам, непредсказуемым как по направлению, так и по величине. Риск перелома зуба также выше при убийстве крупной добычи. [89]
Исследование ископаемых останков крупных хищников из ям Ла Бреа, датированных 36 000–10 000 лет назад, показывает, что уровень поломки зубов составляет 5–17% у лютого волка, койота, американского льва и смилодона по сравнению с 0,5–2,7% у десяти современных хищников. . Более высокая частота переломов наблюдалась во всех зубах, но частота переломов клыков была такой же, как у современных хищников. [ нужны разъяснения ] Ужасный волк чаще ломал резцы по сравнению с современным серым волком; таким образом, было высказано предположение, что ужасный волк во время кормления прижимал свои резцы ближе к кости. Окаменелости ужасных волков из Мексики и Перу демонстрируют аналогичную картину разрушения. Исследование 1993 года показало, что более высокая частота поломки зубов среди хищников плейстоцена по сравнению с ныне живущими хищниками не была результатом охоты на более крупную дичь, о чем можно было предположить, исходя из большего размера первых. Когда добычи мало, конкуренция между хищниками возрастает, заставляя их есть быстрее и, следовательно, потреблять больше костей, что приводит к поломке зубов. [68] [88] [90] Поскольку их добыча вымерла около 10 000 лет назад, вымерли и эти хищники плейстоцена, за исключением койота (который является всеядным ) . [88] [90]
Более позднее исследование ям Ла-Бреа сравнило поломку зубов у ужасных волков в два периода времени. В одной яме находились ископаемые лютые волки возрастом 15 000 лет назад, а в другой - 13 000 лет назад. Результаты показали, что у 15 000 люто волков YBP было в три раза больше поломок зубов, чем у 13 000 люто волков YBP, чья поломка соответствовала таковой у девяти современных хищников. Исследование пришло к выводу, что между 15 000 и 14 000 YBP доступность добычи была меньше или конкуренция была выше для ужасных волков, и что к 13 000 YBP, когда виды-жертвы приближались к исчезновению, конкуренция хищников снизилась, и, следовательно, частота поломки зубов у ужасных волков также увеличилась. отклоненный. [90] [91]
К хищникам относятся как стайные охотники , так и одиночные охотники. Одиночный охотник зависит от мощного укуса клыков, чтобы подчинить свою добычу, и, таким образом, демонстрирует сильный нижнечелюстной симфиз . Напротив, стайный охотник, который наносит много более мелких укусов, имеет сравнительно более слабый нижнечелюстной симфиз. Таким образом, исследователи могут использовать силу нижнечелюстного симфиза у ископаемых экземпляров плотоядных животных, чтобы определить, каким типом охотника они были – стайным или одиночным – и даже как они поедали свою добычу. Нижние челюсти псовых укреплены позади хищных зубов, что позволяет животным раскалывать кости пост-хищными зубами (коренные зубы M2 и M3). Исследование показало, что профиль опоры нижней челюсти лютого волка был ниже, чем у серого волка и красного волка, но очень похож на профиль койота и африканской охотничьей собаки. Слабая в дорсо-вентральном направлении симфизарная область (по сравнению с премолярами P3 и P4) лютого волка указывает на то, что он наносил неглубокие укусы, как и его современные родственники, и поэтому был стайным охотником. Это говорит о том, что ужасный волк, возможно, имел обработанную кость, но не был так хорошо приспособлен к ней, как серый волк. [92] Тот факт, что частота переломов у лютого волка в позднем плейстоцене снизилась до уровня его современных родственников. [88] [91] предполагает, что снижение конкуренции позволило лютому волку вернуться к пищевому поведению, предполагающему меньшее потребление костей, поведению, для которого он лучше всего подходил. [90] [92]
Результаты исследования зубных микроизносов на зубной эмали у экземпляров видов плотоядных животных из ям Ла Бреа, включая ужасных волков, позволяют предположить, что эти хищники не испытывали пищевого стресса непосредственно перед своим вымиранием. Данные также показали, что степень использования туш (т.е. количество потребляемого по отношению к максимально возможному количеству, включая измельчение и потребление костей) была меньше, чем у крупных хищников сегодня. Эти результаты показывают, что поломка зубов была связана с охотничьим поведением и размером добычи. [93]
Влияние климата
[ редактировать ]Предыдущие исследования показали, что изменения размера тела ужасного волка коррелируют с колебаниями климата. [68] [94] В более позднем исследовании сравнивалась морфология краниодентов ужасного волка из четырех ям Ла Бреа, каждая из которых представляет четыре разных периода времени. Результаты свидетельствуют об изменении размера ужасного волка, износа и поломки зубов, формы черепа и формы морды с течением времени. Размер тела ужасного волка уменьшился между началом последнего ледникового максимума и ближе к его концу, во время теплого колебания Аллерёда . Признаки пищевого стресса (нехватка пищи, приводящая к снижению потребления питательных веществ) наблюдаются в меньшем размере тела, черепах с более крупным основанием черепа и более короткой мордой (неотения формы и размера), а также в большей степени поломки и износа зубов. У ужасных волков возрастом 17 900 лет назад были обнаружены все эти особенности, что указывает на пищевой стресс. Ужасные волки, датированные 28 000 лет назад, также в некоторой степени демонстрировали многие из этих особенностей, но были самыми крупными изученными волками, и было высказано предположение, что эти волки также страдали от пищевого стресса и что волки до этой даты были еще больше по размеру. [68] Питательный стресс, вероятно, приведет к усилению силы укуса, что приведет к более полному поеданию туш и растрескиванию костей. [68] [95] и с изменениями формы черепа для улучшения механических преимуществ. Климатические записи Северной Америки показывают циклические колебания во время ледникового периода, которые включали быстрое потепление с последующим постепенным похолоданием, называемые событиями Дансгаарда-Эшгера . Эти циклы вызвали бы повышение температуры и засушливости, а в Ла- Бреа вызвали бы экологический стресс и, следовательно, продовольственный стресс. [68] Подобная тенденция была обнаружена и у серого волка, который в бассейне Санта-Барбары изначально представлял собой массивную, мощную и, возможно, конвергентную эволюцию с ужасным волком, но к началу голоцена был заменен более изящными формами. [35] [34] [68]
Информация об ужасном волке основана на размерах черепа. [68] Переменная 28 000 йен. 26 100 йенских фунтов 17 900 йен. 13 800 йен. Размер тела крупнейший большой самый маленький средний/маленький Поломка зуба высокий низкий высокий низкий Износ зубов высокий низкий высокий низкий Форма морды укорочение, самое большое основание черепа средний самое короткое и самое большое основание черепа средний Форма зубного ряда крепкий – – хрупкий ДЕЛАТЬ событие номер 3 или 4 никто неточные данные неточные данные
Конкуренты
[ редактировать ]
Незадолго до появления ужасного волка в Северную Америку вторгся подрод Xenocyon Canis ( предок азиатского волка и африканской охотничьей собаки), который был таким же большим, как ужасный волк, и более плотоядным. Летопись окаменелостей показывает, что они редки, и предполагается, что они не могли конкурировать с недавно выведенным ужасным волком. [96] Анализ стабильных изотопов свидетельствует о том, что ужасный волк Смилодон и американский лев боролись за одну и ту же добычу. [72] [93] Среди других крупных хищников были вымерший североамериканский гигантский короткомордый медведь ( Arctodus simus ), современная пума ( Puma concolor ), плейстоценовый койот ( Canis latrans ) и плейстоценовый серый волк, который был более массивным и крепким, чем сегодня. Эти хищники, возможно, конкурировали с людьми, охотившимися за аналогичной добычей. [93]
Образцы, идентифицированные по морфологии как берингийские волки ( C. lupus ) и радиоуглеродному датированию 25 800–14 300 лет назад, были обнаружены в пещере Natural Trap у подножия гор Бигхорн в Вайоминге , на западе США. Это место находится прямо к югу от места, которое в то время было разделом между Лаврентидским ледниковым щитом и Кордильерским ледниковым щитом . Возможно, существовал временный канал между ледниками, который позволял этим крупным аляскинским прямым конкурентам лютого волка, которые также были приспособлены для охоты на мегафауну, проникать к югу от ледниковых щитов. Останки лютого волка отсутствуют к северу от 42° северной широты в Северной Америке, следовательно, этот регион был доступен для распространения берингийских волков на юг вдоль линии ледника. Насколько широко они были тогда распространены, неизвестно. Они также вымерли в конце позднего плейстоцена, как и ужасный волк. [44]
После прибытия в восточную Евразию лютому волку, вероятно, пришлось бы столкнуться с конкуренцией со стороны самого доминирующего и широко распространенного хищника этой территории — восточного подвида пещерной гиены ( Crocuta crocuta ultima ). Конкуренция с этим видом, возможно, привела к очень низкому уровню популяций лютого волка в Евразии, что привело к нехватке ископаемых останков лютого волка в этой хорошо изученной ископаемой фауне. [97]
Диапазон
[ редактировать ]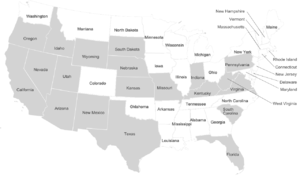
Останки ужасного волка были обнаружены в самых разных средах обитания, включая равнины, луга и некоторые лесные горные районы Северной Америки, засушливые саванны Южной Америки и, возможно, степи Восточной Азии. Высота участков варьируется от уровня моря до 2255 м (7400 футов). [19] Расположение этих ископаемых останков позволяет предположить, что ужасные волки жили преимущественно на открытых низинах вместе со своей добычей — крупными травоядными. [46] Останки ужасного волка не часто встречаются в высоких широтах Северной Америки. [19] Отсутствие окаменелостей использовалось как доказательство того, что лютые волки не мигрировали на восток через Берингию до тех пор, пока в 2020 году не были обнаружены останки азиатского лютого волка. [97]
В Соединенных Штатах окаменелости ужасных волков были обнаружены в Аризоне, Калифорнии, Флориде, Айдахо, Индиане, Канзасе, Кентукки, Миссури, Небраске, Нью-Мексико, Орегоне, Пенсильвании, Южной Каролине, Южной Дакоте, Техасе, Юте, Вирджинии, на Западе. Вирджиния, Вайоминг, [19] и Невада. [98] Идентичность окаменелостей, обнаруженных севернее Калифорнии, не подтверждена. [44] Было пять сообщений о неподтвержденных окаменелостях ужасного волка к северу от 42 ° северной широты в Фоссил-Лейк, штат Орегон (125 000–10 000 лет назад), водохранилище Американский водопад, Айдахо (125 000–75 000 лет назад), пещере Саламандр, Южная Дакота (250 000 лет назад), и четыре близко сгруппированных участка в северной Небраске (250 000 YBP). [44] Это предполагает ограничение ареала обитания ужасных волков из-за температуры, добычи или среды обитания. [44] Основные места добычи ископаемых A. d. dirus расположены к востоку от Скалистых гор и включают пещеру Фризенхан недалеко от Сан-Антонио, штат Техас; Пещера Кэрролл, недалеко от Ричленда, штат Миссури; и Реддик, Флорида . [20]

Места в Мексике, где были собраны останки ужасных волков, включают Эль -Седасо в Агуаскальентесе , муниципалитет Комонду в Южной Нижней Калифорнии , Эль- Седраль в Сан-Луис-Потоси , карьер Эль-Тахо недалеко от Текискиака , штат Мехико Вальсекильо в Пуэбле Халиско , , озеро Чапала в в штате Мехико , пещера Лолтун . Юкатан , Потресито в Синалоа , пещера Сан-Хосесито возле Арамберри в Нуэво-Леоне Терапа в Соноре. и Образцы из Therapa были подтверждены как A. d. гильдайи . [65] Находки в пещере Сан-Хосесито и Эль- Седасо содержат наибольшее количество особей из одного местонахождения.
В Южной Америке ужасные волки датируются возрастом менее 17 000 лет назад и зарегистрированы в шести местах: Муако в западном Фалькон штате в Венесуэле , провинция Талара в Перу , штат Монагас в восточной Венесуэле , департамент Тариха в Боливии , Атакама пустыня Чили и Эквадор . [99] [100] [19] [101] Если ужасный волк возник в Северной Америке, этот вид, вероятно, распространился в Южную Америку через Андский коридор. [19] [102] предполагаемый путь миграции млекопитающих умеренного пояса из Центральной Америки в Южную Америку из-за благоприятных прохладных, сухих и открытых мест обитания, которые иногда характеризовали этот регион. Скорее всего, это произошло во время ледникового периода, поскольку тогда путь состоял из открытых засушливых регионов и саванны, тогда как в межледниковые периоды он состоял бы из влажных тропических лесов. [19] [103]
В 2020 году ископаемая нижняя челюсть, позже проанализированная как принадлежащая лютому волку, была найдена в окрестностях Харбина на северо-востоке Китая. Окаменелость была таксономически описана и датирована 40 000 лет назад. Это открытие бросает вызов предыдущим теориям о том, что низкие температуры и ледяные щиты в северных широтах Северной Америки могут стать барьером для ужасных волков. Эта теория основывалась на том, что окаменелости ужасных волков не были обнаружены выше 42° широты в Северной Америке. Предполагается, что ужасный волк преследовал мигрирующую добычу из средних широт Северной Америки, а затем через Берингию в Евразию. [97]
Вымирание
[ редактировать ]
Во время четвертичного вымирания около 12 700 YBP вымерло 90 родов млекопитающих весом более 44 килограммов (97 фунтов). [54] [68] Считается, что исчезновение крупных хищников и падальщиков было вызвано исчезновением добычи мегатравоядных, от которой они зависели. [104] [105] [19] [88] Причина вымирания мегафауны дискутируется [93] но это объясняется воздействием изменения климата , конкуренцией с другими видами, включая чрезмерную эксплуатацию вновь прибывшими охотниками-людьми, или комбинацией того и другого. [93] [106] В одном исследовании предлагается изучить несколько моделей вымирания, поскольку так мало известно о биогеографии лютого волка, его потенциальных конкурентах и добыче, а также о том, как все эти виды взаимодействовали и реагировали на изменения окружающей среды, произошедшие во время вымирания. [19]
Данные древней ДНК и радиоуглерода показывают, что местные генетические популяции были заменены другими представителями того же вида или другими представителями того же рода. [107] И ужасный волк, и берингийский волк вымерли в Северной Америке, оставив процветать только менее плотоядной и более грациозной форме волка. [79] который, возможно, превзошел ужасного волка. [108] Одно исследование предполагает раннее происхождение линии ужасных волков в Америке, что привело к их репродуктивной изоляции, так что, когда койоты, волки, серые волки и ксеноционы распространились в Северную Америку из Евразии в позднем плейстоцене, не могло быть никакого смешения с ужасный волк. Серые волки и койоты, возможно, выжили благодаря своей способности гибридизироваться с другими псовыми, такими как домашняя собака, и приобретать черты, устойчивые к болезням, приносимым таксонами, прибывшими из Евразии. Репродуктивная изоляция, возможно, помешала лютому волку приобрести эти черты. [21] Исследование 2023 года зафиксировало высокую степень субхондральных дефектов суставных поверхностей у экземпляров лютого волка и смилодона из ям Ла-Бреа-Тар, которые напоминали рассекающий остеохондроз . Поскольку современные собаки с этим заболеванием являются инбредными , исследователи предположили, что то же самое могло произойти с доисторическими видами, когда они были на грани исчезновения, но предупредили, что необходимы дополнительные исследования, чтобы определить, имело ли это место и с образцами из других частей Земли. Америки. [109]
Останки ужасного волка, имеющие самый молодой геологический возраст, датируются 9440 лет назад в пещере Бринюлфсон, округ Бун, штат Миссури . [31] [108] 9 860 YBP в Ранчо Ла Бреа, Калифорния, и 10 690 YBP в Ла Мирада, Калифорния . [108] Останки ужасного волка были датированы радиоуглеродом до 8200 лет назад в Уайтуотер-Дроу в Аризоне . [106] [110] Однако один автор заявил, что радиоуглеродное датирование карбоната кости ненадежно. [19] Все эти даты некалиброваны , и фактический возраст останков, вероятно, старше. В Южной Америке самые последние останки в Таларе , Перу, датируются 9030 ± 240 YBP (также некалиброванные), тогда как самые последние останки «C. nehringi» из Лухана , Аргентина, старше, чем самый последний стратиграфический разрез этого места. датируется 10–11 000 YBP. [111]
См. также
[ редактировать ]Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Перейти обратно: а б с д Лейди, Дж. (1858). «Уведомление об останках вымерших позвоночных из долины реки Ниобрара, собранных во время исследовательской экспедиции 1857 года в Небраске под командованием лейтенанта Г.К. Уоррена, главного инженера США, доктором Ф.В. Хейденом, геологом Экспедиция, Труды» . Труды Академии естественных наук Филадельфии . 10:21 .
- ^ Перейти обратно: а б Мерриам, Дж. К. (1918). «Заметка о систематическом положении волков группы Canis dirus» . Бюллетень кафедры геологии Калифорнийского университета . 10 :533.
- ^ Перейти обратно: а б с д и ж г час я Куртен, Б. (1984). «Географическая дифференциация ранчолабрейского лютого волка ( Canis dirus Leidy) в Северной Америке». В Генуэе, ХХ; Доусон, MR (ред.). Вклад в палеонтологию четвертичных позвоночных: том в память о Джоне Э. Гилдее . Специальное издание 8. Музей естественной истории Карнеги . стр. 218–227 . ISBN 978-0-935868-07-4 .
- ^ Перейти обратно: а б Лейди, Дж. (1854 г.). «Объявление о некоторых ископаемых костях, обнаруженных г-ном Фрэнсисом А. Линке на берегу реки Огайо, штат Индиана» . Труды Академии естественных наук Филадельфии . 7 (7): 200.
- ^ Перейти обратно: а б Лейди, Дж. (1869). «Вымершая фауна млекопитающих Дакоты и Небраски, включая описание некоторых родственных форм из других местностей, а также краткий обзор останков млекопитающих Северной Америки» . Журнал Академии естественных наук Филадельфии . 7 : 368.
- ^ Перейти обратно: а б Аллен, Дж. А. (1876 г.). «Описание некоторых останков вымершего вида волка и вымершего вида оленей из главного региона верхней части Миссисипи» . Американский научный журнал . с3-11 (61): 47–51. Бибкод : 1876AmJS...11...47A . дои : 10.2475/ajs.s3-11.61.47 . hdl : 2027/hvd.32044107326068 . S2CID 88320413 .
- ^ АМЕГИНО, Ф. 1902. Заметки о некоторых млекопитающих.новые или малоизвестные окаменелости из долины Тариха.Анналы Национального музея Буэнос-Айреса, 3-я серия, 1: 225–261.
- ^ Селлардс, Э.Х. (1916). «Человеческие останки и связанные с ними окаменелости плейстоцена Флориды» . Годовой отчет Геологической службы Флориды . 8 : 152.
- ^ Фрик, К. (1930). «Замерзшая фауна Аляски». Естественная история (30): 71–80.
- ^ От греческого αἰνος ( ainós ) «ужасный» + κύων ( kúōn ) «собака» и латинского dirus «угрожающий»
- ^ Пейджский музей. «Просмотр коллекций на ранчо Ла Бреа» . Фонд Музея естественной истории округа Лос-Анджелес . Архивировано из оригинала 25 января 2017 года . Проверено 19 декабря 2016 г.
- ^ МКЗН (2017). «Кодекс онлайн (см. главу 6, статью 23.1)» . Международный кодекс зоологической номенклатуры онлайн . Международная комиссия по зоологической номенклатуре. Архивировано из оригинала 24 мая 2009 г. Проверено 17 апреля 2017 г.
- ^ Перейти обратно: а б с д и ж Мерриам, Дж. К. (1912). «Фауна Ранчо Ла Бреа, Часть II. Canidae» . Мемуары Калифорнийского университета . 1 : 217–273.
- ^ Трокселл, Э.Л. (1915). «Окаменелости позвоночных Рок-Крик, Техас» . Американский научный журнал . 189 (234): 613–618. Бибкод : 1915AmJS...39..613T . дои : 10.2475/ajs.s4-39.234.613 .
- ^ Стивенсон, Марк (1978). «9» . Ин Холл, Роберта Л.; Шарп, Генри С. (ред.). Волк и человек: параллельная эволюция . Нью-Йорк: Академическая пресса. п. 180. ИСБН 978-0-12-319250-9 . Проверено 1 мая 2017 г.
- ^ Лунделиус, Э.Л. (1972). «Ископаемые позвоночные, фауна Инглсайда позднего плейстоцена, округ Сан-Патрисио, Техас». Бюро экономической геологии . Отчет о расследовании № 77: 1–74.
- ^ Перейти обратно: а б Новак 1979 , стр. 106–108.
- ^ Ван, X. (1990). «Останки ужасного волка плейстоцена из реки Канзас с заметками об ужасных волках в Канзасе». Периодические статьи Музея естественной истории Канзасского университета . 137 : 1–7.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Дандас, Р.Г. (1999). «Четвертичные находки лютого волка Canis dirus в Северной и Южной Америке» (PDF) . Борей . 28 (3): 375–385. Бибкод : 1999Борея..28..375D . дои : 10.1111/j.1502-3885.1999.tb00227.x . S2CID 129900134 . Архивировано из оригинала (PDF) 27 сентября 2020 г. Проверено 28 августа 2015 г.
- ^ Перейти обратно: а б с д и ж г час я дж к Энионг, Уильям; Роман, Крис (2006). «Новые оценки массы тела Canis dirus , вымершего лютого волка плейстоцена». Журнал палеонтологии позвоночных . 26 : 209–212. doi : 10.1671/0272-4634(2006)26[209:NBMEFC]2.0.CO;2 . S2CID 83702167 .
- ^ Перейти обратно: а б с д и ж г час я Перри, Анджела Р.; Митчелл, Кирен Дж.; Мутон, Алиса; Альварес-Карретеро, Сандра; Халм-Биман, Ардерн; Хейл, Джеймс; Джеймисон, Александра; Мичен, Джули; Лин, Одри Т.; Шуберт, Блейн В.; Амин, Карли; Антипина Екатерина Евгеньевна; Бовер, Пере; Брейс, Селина; Карманьини, Альберто; Карё, Кристиан; Саманьего Каструита, Хосе А.; Чаттерс, Джеймс К.; Добни, Кейт; Два короля, Марио; Эвин, Аллоуэн; Гобер, Филипп; Гопалакришнан, Шьям; Гауэр, Грэм; Хейнигер, Холли; Хелген, Кристофер М.; Капп, Джош; Косинцев Павел А.; Линдерхольм, Анна; Озга, Эндрю Т.; Прессли, Саманта; Салис, Александр Т.; Сареми, Недда Ф.; Шоу, Колин; Скерри, Кэтрин; Тараненко Дмитрий Евгеньевич; Томпсон, Мэри; Саблин Михаил Владимирович; Кузьмин Ярослав В.; Коллинз, Мэтью Дж.; Синдинг, Миккель-Хольгер С.; Гилберт, М. Томас П .; Стоун, Энн К.; Шапиро, Бет; Ван Валкенбург, Блэр ; Уэйн, Роберт К.; Ларсон, Грегер; Купер, Алан; Франц, Лоран А.Ф. (2021). «Ужасные волки были последними представителями древней линии псовых Нового Света» . Природа . 591 (7848): 87–91. Бибкод : 2021Natur.591...87P . дои : 10.1038/s41586-020-03082-x . ПМИД 33442059 . S2CID 231604957 .
- ^ Ван и Тедфорд 2008 , стр. 20
- ^ Ван, Сяомин (2008). «Как собаки стали править миром» . Журнал естественной истории . Том. июль/август . Проверено 24 мая 2014 г.
- ^ Ван и Тедфорд 2008 , стр. 49
- ^ Перейти обратно: а б Тедфорд, Ван и Тейлор, 2009 г. , стр. 4.
- ^ Перейти обратно: а б с д и ж г час я Ван и Тедфорд 2008 , стр. 148–150
- ^ Ван и Тедфорд 2008 , стр. 143–144
- ^ Перейти обратно: а б с д и ж Тедфорд, Ван и Тейлор, 2009 , стр. 181.
- ^ Мартин, РА; Уэбб, SD (1974). «Млекопитающие позднего плейстоцена из фауны Логова Дьявола, округ Леви». В Уэббе, С.Д. (ред.). Плейстоценовые млекопитающие Флориды . Гейнсвилл: Университетское издательство Флориды . стр. 114–145 . ISBN 978-0-8130-0361-0 .
- ^ Перейти обратно: а б Берта 1988 , стр. 50
- ^ Перейти обратно: а б с д Куртен, Б.; Андерсон, Э. (1980). «11-Карнавора». Плейстоценовые млекопитающие Северной Америки . Издательство Колумбийского университета , Нью-Йорк. стр. 168–172. ISBN 978-0-231-03733-4 .
- ^ Гейст, Валериус (1998). Олени мира: их эволюция, поведение и экология (1-е изд.). Механиксбург, Пенсильвания: Stackpole Books . п. 10. ISBN 978-0-8117-0496-0 . Проверено 1 мая 2017 г.
- ^ Гейст, Валериус (1987). «О видообразовании млекопитающих ледникового периода, с особым упором на оленьих и капридов». Канадский журнал зоологии . 65 (5): 1067–1084. дои : 10.1139/z87-171 .
- ^ Перейти обратно: а б Гуле, Г.Д. (1993). Сравнение временных и географических вариаций черепа неарктических, современных, голоценовых и позднеплейстоценовых серых волков ( Canis lupus ) и избранных Canis (магистерская диссертация). Университет Манитобы, Виннипег. стр. 1–116.
- ^ Перейти обратно: а б Новак 1979 , стр. 102.
- ^ Новак, Рональд М.; Федерофф, Николас Э. (Бруско) (2002). «Систематический статус итальянского волка Canis lupus ». Акта Териологика . 47 (3): 333–338. дои : 10.1007/BF03194151 . S2CID 366077 .
- ^ Перейти обратно: а б Р. М. Новак (2003). «Эволюция и систематика 9-волков». В Мехе, Л. Дэвид; Бойтани, Луиджи (ред.). Волки: поведение, экология и охрана . Издательство Чикагского университета . стр. 242–243. ISBN 978-0-226-51696-7 .
- ^ Новак 1979 , стр. 84.
- ^ Перейти обратно: а б с д и Тедфорд, Ван и Тейлор, 2009 г. , стр. 146–148.
- ^ Перейти обратно: а б с Превости, Франциско Дж. (2010). «Филогения крупных вымерших южноамериканских псовых (Mammalia, Carnivora, Canidae) с использованием подхода «полных доказательств»» . Кладистика . 26 (5): 456–481. дои : 10.1111/j.1096-0031.2009.00298.x . ПМИД 34875763 . S2CID 86650539 . См. страницу 472.
- ^ Новак 1979 , стр. 93.
- ^ Лунделиус, EL; Белл, CJ (2004). «7 Бланканский, ирвингтонский и ранчолабрейский возрасты млекопитающих» (PDF) . В Майкле О. Вудберне (ред.). Позднемеловые и кайнозойские млекопитающие Северной Америки: биостратиграфия и геохронология . Издательство Колумбийского университета. п. 285. ИСБН 978-0-231-13040-0 . Архивировано из оригинала (PDF) 8 июня 2017 года . Проверено 1 мая 2017 г.
- ^ Перейти обратно: а б Мид, Дж.И.; Манганаро, К.; Репеннинг, Калифорния; Агенброуд, LD (1996). «Ранние ранчолабрейские млекопитающие из пещеры Саламандр, Блэк-Хиллз, Южная Дакота» . В Стюарте, КМ; Сеймур, КЛ (ред.). Палеоэкология и палеосреда позднекайнозойских млекопитающих, дань уважения карьере К.С. (Руфуса) Чёрчера . Университет Торонто Пресс , Торонто, Онтарио, Канада. стр. 458–482. ISBN 978-0-8020-0728-5 . Архивировано из оригинала 10 января 2015 г.
- ^ Перейти обратно: а б с д и ж г Мичен, Джули А.; Браник, Александрия Л.; Фрай, Трент Дж. (2016). «Вымерший морфотип берингийского волка, обнаруженный в континентальной части США, имеет значение для миграции и эволюции волков» . Экология и эволюция . 6 (10): 3430–8. Бибкод : 2016EcoEv...6.3430M . дои : 10.1002/ece3.2141 . ПМК 4870223 . ПМИД 27252837 .
- ^ Берта 1988 , стр. 113
- ^ Перейти обратно: а б Новак 1979 , стр. 116–117.
- ^ Перейти обратно: а б Красный, Ян; Дуда, Павел; Робовский, Ян; Окринова, Изабела; Павелкова Ржичанкова, Вера (2018). «Филогения Caninae (Carnivora): сочетание морфологии, поведения, генов и окаменелостей». Зоологика Скрипта . 47 (4): 373–389. дои : 10.1111/zsc.12293 . S2CID 90592618 .
- ^ Руководитель, Дэн; Ян, Янгешань; Ли, Цян; Ни, Сицзюнь (октябрь 2020 г.). «Окаменелость позднего плейстоцена из северо-восточного Китая является первой находкой лютого волка (Carnivora: Canis dirus) в Евразии» . Четвертичный интернационал . 591 : 87–92. Бибкод : 2021QuInt.591...87L . дои : 10.1016/j.quaint.2020.09.054 . S2CID 224877090 – через ResearchGate.
- ^ Кёпфли, К.-П.; Поллинджер, Дж.; Годиньо, Р.; Робинсон, Дж.; Леа, А.; Хендрикс, С.; Швейцер, РМ; Тельманн, О.; Сильва, П.; Фан, З.; Юрченко А.А.; Добрынин П.; Макунин А.; Кэхилл, Дж.А.; Шапиро, Б.; Альварес, Ф.; Брито, Джей Си; Геффен, Э.; Леонард, Дж.А.; Хелген, КМ; Джонсон, МЫ; О'Брайен, SJ; Ван Валкенбург, Б.; Уэйн, РК (17 августа 2015 г.). «Общегеномные данные показывают, что африканские и евразийские золотые шакалы являются разными видами» . Современная биология . 25 (16): 2158–65. Бибкод : 2015CBio...25.2158K . дои : 10.1016/j.cub.2015.06.060 . ПМИД 26234211 .
- ^ АЛЬВАРЕС, Франциско; Богданович, Веслав; Кэмпбелл, Лиз А.Д.; ГОДИНЬО, Рэйчел; Хатлауф, Дженнифер; Джала, Ядвендрадев В.; Китченер, Эндрю К.; Кёпфли, Клаус-Петер; Крофель, Миха; Мельман, Патрисия Д.; Сенн, Хелен; Силлеро-Зубири, Клаудио; Виранта, Суви; Верхан, Джеральдин (2019). «Виды Old World Canis с таксономической неопределенностью: выводы и рекомендации семинара. CIBIO. Вайран, Португалия, 28–30 мая 2019 г.» (PDF) . Группа специалистов МСОП/SSC по собакам . Проверено 6 марта 2020 г.
- ^ Янчевский, Д.Н.; Юки, Н.; Гилберт, округ Колумбия; Джефферсон, GT; О'Брайен, SJ (1992). «Молекулярно-филогенетический вывод на основе окаменелостей саблезубых кошек с Ранчо Ла Бреа» . Труды Национальной академии наук Соединенных Штатов Америки . 89 (20): 9769–73. Бибкод : 1992PNAS...89.9769J . дои : 10.1073/pnas.89.20.9769 . ПМЦ 50214 . ПМИД 1409696 .
- ^ Голд, Дэвид А.; Робинсон, Жаклин; Фаррелл, Эслинг Б.; Харрис, Джон М.; Тельманн, Олаф; Джейкобс, Дэвид К. (2014). «Попытка экстракции ДНК из колумбийского мамонта Ранчо Ла Бреа ( Mammuthus columbi ): Перспективы древней ДНК из асфальтовых отложений» . Экология и эволюция . 4 (4): 329–36. Бибкод : 2014EcoEv...4..329G . дои : 10.1002/ece3.928 . ПМЦ 3936381 . ПМИД 24634719 .
- ^ Перейти обратно: а б с д и Сток, К. (1992). Ранчо Ла Бреа: записи плейстоценовой жизни в Калифорнии . Научная серия (7-е изд.). Музей естественной истории округа Лос-Анджелес. стр. 1–113. ISBN 978-0-938644-30-9 .
- ^ Перейти обратно: а б с д О'Киф, Франция; Фет, Е.В.; Харрис, Дж. М. (2009). «Компиляция, калибровка и синтез радиоуглеродных дат фауны и флоры, Ранчо Ла Бреа, Калифорния» (PDF) . Вклад в науку . 518 : 1–16. дои : 10.5962/стр.226783 . S2CID 128107590 . Архивировано из оригинала (PDF) 3 июля 2017 г. Проверено 28 июня 2017 г.
- ^ Перейти обратно: а б Тедфорд, Ван и Тейлор, 2009 , стр. 325.
- ^ Перейти обратно: а б с Мех, Л. Дэвид (1966). Волки острова Рояль . Фауна Серия 7. Фауна национальных парков США. ISBN 978-1-4102-0249-9 . Проверено 1 мая 2017 г.
- ^ Перейти обратно: а б с д Хартстон-Роуз, Адам; Дандас, Роберт Г.; Бойд, Бриттин; Лонг, Райан С.; Фаррелл, Эслинг Б.; Шоу, Кристофер А. (15 сентября 2015 г.). Джон М. Харрис (ред.). «Бакула ранчо Ла Бреа». Вклад в науку . Научная серия 42 (специальный том под названием «Ла Бреа и за его пределами: Палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия Музея естественной истории раскопок округа Лос-Анджелес на Ранчо ла Бреа). Музей естественной истории округа Лос-Анджелес: 53–63.
- ^ Перейти обратно: а б с д Сток, Честер; Лэнс, Джон Ф. (1948). «Относительная длина элементов конечностей Canis dirus » . Бюллетень Академии наук Южной Калифорнии . 47 (3): 79–84.
- ^ Перейти обратно: а б Соркин, Борис (2008). «Биомеханическое ограничение массы тела у наземных хищников-млекопитающих». Летайя . 41 (4): 333–347. Бибкод : 2008Лета..41..333С . дои : 10.1111/j.1502-3931.2007.00091.x .
- ^ «Серый волк (на Юконе)» (PDF) . Окружающая среда Юкон . Правительство Канады. 2017 . Проверено 18 апреля 2017 г. .
- ^ Ранчо ла Бреа. Реставрация Часа. Р. Найт. Мурал для амер. Музейный зал Человека. Береговой хребет на заднем плане, Старый Болди слева [1]
- ^ Перри, Анджела (2016). «Волк в собачьей шкуре: первоначальное приручение собаки и плейстоценовая вариация волка». Журнал археологической науки . 68 : 1–4. Бибкод : 2016JArSc..68....1P . дои : 10.1016/j.jas.2016.02.003 .
- ^ Леонард, Дженнифер (2014). «Экология стимулирует эволюцию серых волков» (PDF) . Эволюционно-экологические исследования . 16 : 461–473.
- ^ Флауэр, Люси, Огайо; Шреве, Даниэль К. (2014). «Исследование палеодиетической изменчивости европейских псовых плейстоцена». Четвертичные научные обзоры . 96 : 188–203. Бибкод : 2014QSRv...96..188F . doi : 10.1016/j.quascirev.2014.04.015 .
- ^ Перейти обратно: а б с д и ж г Ходнетт, Джон-Пол; Мид Джим; Баэз, А. (март 2009 г.). «Ужасный волк, Canis dirus (Mammalia; Carnivora; Canidae), из позднего плейстоцена (ранчолабрей) Восточно-Центральной Соноры, Мексика». Юго-западный натуралист . 54 (1): 74–81. дои : 10.1894/CLG-12.1 . S2CID 84760786 .
- ^ Перейти обратно: а б с д и ж Анионг, В.; Бейкер, А. (2006). «Черепно-лицевая морфология и пищевое поведение Canis dirus , вымершего лютого волка плейстоцена». Журнал зоологии . 269 (3): 309–316. дои : 10.1111/j.1469-7998.2006.00043.x .
- ^ Перейти обратно: а б Роу, С.; МакГенри, К.; Томасон, Дж. (2005). «Клуб укусов: Сравнительная сила укуса крупных кусающих млекопитающих и прогноз хищнического поведения ископаемых таксонов» . Труды Королевского общества B: Биологические науки . 272 (1563): 619–25. дои : 10.1098/рспб.2004.2986 . ПМК 1564077 . ПМИД 15817436 .
- ^ Перейти обратно: а б с д и ж г час я дж к О'Киф, Ф.Робин; Биндер, Венди Дж.; Фрост, Стивен Р.; Садлиер, Редьярд В.; Ван Валкенбург, Блэр (2014). «Краниальная морфометрия лютого волка Canis dirus на ранчо Ла Бреа: временная изменчивость и ее связь с питательным стрессом и климатом» . Электронная палеонтология . 17 (1): 1–24.
- ^ Перейти обратно: а б Межправительственная группа экспертов по изменению климата (ООН) (2007 г.). «Четвертый оценочный доклад МГЭИК: Изменение климата, 2007 г. – Палеоклиматическая перспектива» . Нобелевский фонд . Архивировано из оригинала 30 октября 2015 г. Проверено 11 декабря 2016 г.
- ^ Перейти обратно: а б Кларк, штат Пенсильвания; Дайк, А.С.; Шакун, доктор медицинских наук; Карлсон, А.Е.; Кларк, Дж.; Вольфарт, Б. ; Митровица, JX; Хостетлер, Юго-Запад; Маккейб, AM (2009). «Последний ледниковый максимум». Наука . 325 (594 1): 710–4. Бибкод : 2009Sci...325..710C . дои : 10.1126/science.1172873 . ПМИД 19661421 . S2CID 1324559 .
- ^ Элиас, SA; Шриве, Д. (2016). «Позднеплейстоценовое вымирание мегафауны». Справочный модуль по системам Земли и наукам об окружающей среде (PDF) . стр. 3202–3217. дои : 10.1016/B978-0-12-409548-9.10283-0 . ISBN 978-0-12-409548-9 . S2CID 130031864 . Архивировано из оригинала (PDF) 20 декабря 2016 г.
- ^ Перейти обратно: а б с д Колтрейн, Джоан Бреннер; Харрис, Джон М.; Серлинг, Туре Э.; Элерингер, Джеймс Р.; Диринг, Мария-Дениз; Уорд, Джой; Аллен, Джули (2004). «Биогеохимия стабильных изотопов Ранчо Ла Бреа и ее значение для палеоэкологии позднего плейстоцена прибрежной южной Калифорнии». Палеогеография, Палеоклиматология, Палеоэкология . 205 (3–4): 199–219. Бибкод : 2004PPP...205..199C . дои : 10.1016/j.palaeo.2003.12.008 .
- ^ Перейти обратно: а б с д Фокс-Доббс, К.; Бамп, Дж. К.; Петерсон, Р.О.; Фокс, ДЛ; Кох, PL (2007). «Переменные стабильных изотопов, характерные для плотоядных животных, и вариации в экологии кормления современных и древних популяций волков: тематические исследования на острове Рояль, Миннесота и Ла Бреа» (PDF) . Канадский журнал зоологии . 85 (4): 458–471. дои : 10.1139/Z07-018 . Архивировано из оригинала (PDF) 9 августа 2017 года . Проверено 6 мая 2017 г.
- ^ Моратто, Майкл Дж. (1984). Калифорнийская археология . Academic Press , Орландо, Флорида. п. 89. ИСБН 978-0-12-506180-3 . Проверено 1 мая 2017 г.
- ^ Джонсон, Дональд Ли (1977). «Позднечетвертичный климат прибрежной Калифорнии: свидетельства рефугиума ледникового периода». Четвертичные исследования . 8 (2): 154–179. Бибкод : 1977QuRes...8..154J . дои : 10.1016/0033-5894(77)90043-6 . S2CID 129072450 .
- ^ Колледж искусств и наук (2016). «Бассейн с смолой на ранчо ла Бреа. Реставрация Брюса Хорсфолла для WB Scott» . Университет Кейс Вестерн Резерв . Проверено 24 декабря 2016 г.
- ^ Мерриам, Дж. К. (1911). Фауна Ранчо Ла Бреа . Том. 1. Калифорнийский университет – Беркли. стр. 224–225 .
- ^ Валкенбург, Б. Ван; Рафф, CB (1987). «Сила клыков и убийственное поведение крупных хищников». Журнал зоологии . 212 (3): 379–397. дои : 10.1111/j.1469-7998.1987.tb02910.x .
- ^ Перейти обратно: а б с д Леонард, Дж.А.; Вила, К; Фокс-Доббс, К.; Кох, Польша; Уэйн, РК; Ван Валкенбург, Б. (2007). «Вымирание мегафауны и исчезновение специализированного волка-эктоморфа» (PDF) . Современная биология . 17 (13): 1146–50. Бибкод : 2007CBio...17.1146L . дои : 10.1016/j.cub.2007.05.072 . hdl : 10261/61282 . ПМИД 17583509 . S2CID 14039133 . Архивировано из оригинала (PDF) 28 декабря 2016 года . Проверено 6 мая 2017 г.
- ^ Перейти обратно: а б Маркус, LF; Бергер, Р. (1984). «Значение радиоуглеродных дат для Ранчо Ла Бреа». В Мартине, PS; Кляйн, Р.Г. (ред.). Четвертичные вымирания . Университет Аризоны Пресс, Тусон. стр. 159–188. ISBN 978-0-8165-0812-9 .
- ^ Перейти обратно: а б МакХорс, Брианна К.; Оркатт, Джон Д.; Дэвис, Эдвард Б. (2012). «Хищная фауна Ранчо Ла Бреа: средняя или аномальная?». Палеогеография, Палеоклиматология, Палеоэкология . 329–330: 118–123. Бибкод : 2012PPP...329..118M . дои : 10.1016/j.palaeo.2012.02.022 .
- ^ Перейти обратно: а б с д Ван Валкенбург, Блэр; Сакко, Тайсон (2002). «Половой диморфизм, социальное поведение и внутриполовая конкуренция у крупных хищников плейстоцена». Журнал палеонтологии позвоночных . 22 : 164–169. doi : 10.1671/0272-4634(2002)022[0164:SDSBAI]2.0.CO;2 . S2CID 86156959 .
- ^ Перейти обратно: а б Ван Валкенбург, Б.; Кёпфли, К.-П. (1993). «Черепная и зубная адаптация псовых к хищничеству». В Данстоне, Северная Каролина; Горман, Дж.Л. (ред.). Млекопитающие как хищники . Оксфорд: Издательство Оксфордского университета. стр. 15–37.
- ^ Перейти обратно: а б Ван Валкенбург, Б. (1998). «Упадок североамериканских хищников в позднем плейстоцене». В Сондерсе, Джей-Джей; Стили, ЧБ; Барышников Г.Ф. (ред.). Четвертичная палеозоология Северного полушария . Научные статьи Государственного музея Иллинойса, Спрингфилд. стр. 357–374. ISBN 978-0-89792-156-5 .
- ^ Карбоне, К.; Мэддокс, Т.; Фанстон, Пи Джей; Миллс, MGL; Гретер, ГФ; Ван Валкенбург, Б. (2009). «Параллели между воспроизведением и просачиванием смолы плейстоцена позволяют предположить социальность вымершего саблезубого кота Смилодона » . Письма по биологии . 5 (1): 81–5. дои : 10.1098/rsbl.2008.0526 . ПМЦ 2657756 . ПМИД 18957359 .
- ^ Эрл, М. (1987). «Гибкая масса тела социальных хищников». Американский натуралист . 129 (5): 755–760. дои : 10.1086/284670 . S2CID 85236511 .
- ^ Фокс-Доббс, К.; Кох, Польша; Клементц, Монтана (2003). «Обед в Ла Бреа: изотопная реконструкция режима питания Smilodon Fatalis и Canis dirus во времени». Журнал палеонтологии позвоночных . 23 (3, Дополнение): 51А. дои : 10.1080/02724634.2003.10010538 . S2CID 220410105 .
- ^ Перейти обратно: а б с д и ж Ван Валкенбург, Блэр; Гертель, Фриц (1993). «Тяжелые времена в Ла-Бреа: поломка зубов у крупных хищников позднего плейстоцена» (PDF) . Наука . Новая серия. 261 (5120): 456–459. Бибкод : 1993Sci...261..456V . дои : 10.1126/science.261.5120.456 . ПМИД 17770024 . S2CID 39657617 .
- ^ Перейти обратно: а б Ван Валкенбург, Б. (1988). «Частота поломки зубов у крупных хищных млекопитающих». Американский натуралист . 131 (2): 291–302. дои : 10.1086/284790 . S2CID 222330098 .
- ^ Перейти обратно: а б с д Ван Валкенбург, Блэр (2008). «Цены хищничества: перелом зубов у плейстоценовых и современных хищников» . Биологический журнал Линнеевского общества . 96 : 68–81. дои : 10.1111/j.1095-8312.2008.01108.x .
- ^ Перейти обратно: а б Биндер, Венди Дж.; Томпсон, Элисия Н.; Ван Валкенбург, Блэр (2002). «Временные изменения переломов зубов у ужасных волков Ранчо Ла Бреа». Журнал палеонтологии позвоночных . 22 (2): 423–428. doi : 10.1671/0272-4634(2002)022[0423:TVITFA]2.0.CO;2 . S2CID 85799312 .
- ^ Перейти обратно: а б Терриен, Франсуа (2005). «Профили силы нижней челюсти современных хищников и последствия для пищевого поведения вымерших хищников». Журнал зоологии . 267 (3): 249–270. дои : 10.1017/S0952836905007430 .
- ^ Перейти обратно: а б с д и ДеСантис, LRG; Шуберт, Б.В.; Шмитт-Линвилл, Э.; Унгар, П.; Донохью, С.; Хаупт, Р.Дж. (15 сентября 2015 г.). Джон М. Харрис (ред.). «Текстуры стоматологической микроодежды хищников из смоляных ям Ла-Бреа, Калифорния, и возможные последствия их вымирания» . Вклад в науку . Научная серия 42 (специальный том под названием «Ла Бреа и за его пределами: Палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия Музея естественной истории раскопок округа Лос-Анджелес на Ранчо ла Бреа). Музей естественной истории округа Лос-Анджелес: 37–52.
- ^ О'Киф, Франция (2008). «Реакция популяции лютого волка Canis dirus на изменение климата в верхнем плейстоцене». Журнал палеонтологии позвоночных . 28 :122А. дои : 10.1080/02724634.2008.10010459 . S2CID 220405736 .
- ^ Ценг, Чжицзе Джек; Ван, Сяомин (2010). «Краниальная функциональная морфология ископаемых собак и адаптация к дурофагии у Borophagus и Epicyon (Carnivora, Mammalia)». Журнал морфологии . 271 (11): 1386–98. дои : 10.1002/jmor.10881 . ПМИД 20799339 . S2CID 7150911 .
- ^ Ван и Тедфорд 2008 , стр. 60
- ^ Перейти обратно: а б с Дэн, руководитель; Ян, Янгешань; Лия, Цян; Ния, Сицзюнь (1 октября 2020 г.). «Окаменелость позднего плейстоцена из северо-восточного Китая является первой находкой лютого волка (Carnivora: Canis dirus) в Евразии». Четвертичный интернационал . 591 : 87–92. Бибкод : 2021QuInt.591...87L . дои : 10.1016/j.quaint.2020.09.054 . S2CID 224877090 .
- ^ Скотт, Эрик; Спрингер, Кэтлин Б. (2016). «Первые находки Canis dirus и Smilodon Fatalis из позднеплейстоценовой местной фауны Туле-Спрингс, верхняя часть Лас-Вегас-Уош, Невада» . ПерДж . 4 : е2151. дои : 10.7717/peerj.2151 . ПМЦ 4924133 . ПМИД 27366649 .
- ^ Тедфорд, Ван и Тейлор 2009 , с. 146.
- ^ Каро, Франсиско Дж.; Лабарка, Рафаэль; Провости, Франциско Дж.; Вильявисенсио, Наталья; Ярпа, Габриэла М.; Эррера, Кэтрин А.; Корреа-Лау, Жаклин; Латорре, Клаудио; Санторо, Калоджеро М. (2023). «Первая запись Aenocyon dirus (Leidy, 1858) (Carnivora, Canidae) из верхнего плейстоцена пустыни Атакама, север Чили». Журнал палеонтологии позвоночных . 42 (4). дои : 10.1080/02724634.2023.2190785 . S2CID 258757704 .
- ^ Руис-Рамони, Дамиан; Ван, Сяомин; Ринкон, Асканио Д. (2022). «Псы (Caninae) из прошлого Венесуэлы» . Амегинян . 59 . дои : 10.5710/AMGH.16.09.2021.3448 . S2CID 240576546 .
- ^ Берта 1988 , стр. 119
- ^ Уэбб, SD (1991). «Экогеография и великий американский обмен». Палеобиология . 17 (3): 266–280. Бибкод : 1991Pbio...17..266W . дои : 10.1017/S0094837300010605 . S2CID 88305955 .
- ^ Грэм, RW; Мид, Дж.И. (1987). «Колебания окружающей среды и эволюция фауны млекопитающих во время последней дегляциации в Северной Америке». В Раддимане, штат Вирджиния; Райт, HE (ред.). Северная Америка и прилегающие океаны во время последней дегляциации . Геологическое общество Америки K – 3, Боулдер, Колорадо. стр. 371–402. ISBN 978-0-8137-5203-7 .
- ^ Барноский, А.Д. (1989). «Вымирание в позднем плейстоцене как парадигма повсеместного вымирания млекопитающих». В Доноване, Стивен К. (ред.). Массовые вымирания: процессы и доказательства . Издательство Колумбийского университета , Нью-Йорк. стр. 235–255. ISBN 978-0-231-07091-1 .
- ^ Перейти обратно: а б Браник, Александрия Л.; Мичен, Джули А.; О'Киф, Ф. Робин (15 сентября 2015 г.). Джон М. Харрис (ред.). «Микроэволюция формы челюсти у лютого волка Canis dirus на ранчо Ла Бреа». Вклад в науку . Научная серия 42 (специальный том под названием «Ла Бреа и за его пределами: Палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия Музея естественной истории раскопок округа Лос-Анджелес на Ранчо ла Бреа). Музей естественной истории округа Лос-Анджелес: 23–32.
- ^ Купер, А. (2015). «Резкое потепление привело к смене мегафауны в позднем плейстоцене Голарктики» . Наука . 349 (6248): 602–6. Бибкод : 2015Sci...349..602C . дои : 10.1126/science.aac4315 . ПМИД 26250679 . S2CID 31686497 .
- ^ Перейти обратно: а б с Андерсон, Элейн (1984). «Глава 2: Кто есть кто в плейстоцене» . У Пола С. Мартина; Ричард Г. Кляйн (ред.). Четвертичные вымирания: доисторическая революция . Тусон: Издательство Университета Аризоны . п. 55. ИСБН 978-0-8165-1100-6 .
- ^ Шмёкель, Хью; Фаррелл, Эслинг; Балиси, Майрин Ф. (2023). «Субхондральные дефекты, напоминающие рассекающий остеохондроз, на суставных поверхностях вымершего саблезубого кота Smilodon Fatalis и лютого волка Aenocyon dirus » . ПЛОС ОДИН 18 (7): e0287656. Бибкод : 2023PLoSO..1887656S . дои : 10.1371/journal.pone.0287656 . ПМЦ 10337945 . ПМИД 37436967 .
- ^ Хестер, Джим Дж. (1960). «Позднее плейстоценовое вымирание и радиоуглеродное датирование». Американская древность . 26 (1): 58–77. дои : 10.2307/277160 . JSTOR 277160 . S2CID 161116564 .
- ^ Превости, Ф.Дж., Тонни, Э.П., и Бидегейн, Дж.К. (2009). «Стратиграфический ареал крупных псовых (Carnivora, Canidae) в Южной Америке и его значение для четвертичной биостратиграфии». Четвертичный Интернационал , 210 (1–2), 76–81.
Цитируемые работы
[ редактировать ]- Берта, А. (1988). «Четвертичная эволюция и биогеография крупных южноамериканских Canidae (Mammalia: Carnivora)» . Публикации Калифорнийского университета по геологическим наукам (132). ISBN 978-0-520-09960-9 .
- Новак, Рональд М. (1979). Североамериканский четвертичный пес . 6. Монография Музея естественной истории Канзасского университета. дои : 10.5962/bhl.title.4072 . ISBN 978-0-89338-007-6 . Проверено 1 мая 2017 г.
- Тедфорд, Ричард Х .; Ван, Сяомин ; Тейлор, Берил Э. (2009). «Филогенетическая систематика североамериканских ископаемых собак (Carnivora: Canidae)» (PDF) . Бюллетень Американского музея естественной истории . 325 : 1–218. дои : 10.1206/574.1 . hdl : 2246/5999 . S2CID 83594819 .
- Ван, Сяомин ; Тедфорд, Ричард Х. (2008). Собаки: их ископаемые родственники и история эволюции . Издательство Колумбийского университета , Нью-Йорк. стр. 1–232. ISBN 978-0-231-13529-0 .
Внешние ссылки
[ редактировать ]- Для юных читателей – «Лютый волк» Марка Заблудоффа, Маршалла Кавендиша, 2009 г.
- Эвансвилльский ужасный волк
- Информация об ужасном волке из Музея штата Иллинойс.
- Канина (подтриба)
- Доисторические собаки
- Плейстоценовые хищники
- Голоценовые вымирания
- Первые появления в плейстоцене
- Плейстоценовые млекопитающие Северной Америки
- Плейстоценовые млекопитающие Южной Америки
- Плейстоценовые млекопитающие Азии
- Ископаемые таксоны, описанные в 1858 году.
- Млекопитающие, описанные в 1858 г.
- Высшие хищники