Парамилодон
| Парамилодон | |
|---|---|

| |
| Скелет в музее смолистых ям Ла Бреа | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | волосатый |
| Семья: | † Милодонтиды |
| Подсемейство: | † Милодонтины |
| Племя: | † Милодонтини |
| Род: | † Парамилодон Браун , 1903 год. |
| Типовой вид | |
| † Парамилодон харлани ( Оуэн , 1840 г.)
| |
Paramylodon — вымерший род наземных ленивцев семейства Mylodontidae , эндемичный для Северной Америки в период от плиоцена до плейстоцена , живший примерно 4,9 млн лет назад — 12 000 лет назад.
Внутри рода признаны только два вида: Paramylodon harlani, также известный как наземный ленивец Харлана, известный от раннего плейстоцена до позднего плейстоцена ( ирвингтон - ранчолабрей ), и более ранний плиоцен -раннеплейстоценовый ( бланканский ) вид Paramylodon garbanii, хотя размещение последний представитель этого рода подвергался сомнению некоторыми авторами. Первые находки окаменелостей относятся к началу 1830-х годов. Они восходят к Ричарду Харлану , в честь которого был назван вид. Род Paramylodon был введен Барнумом Брауном в начале 20 века. Спустя более 150 лет после описания первого вида находки, которые сейчас относят к Paramylodon, неоднократно относили к другим родам, сначала к Mylodon , а с 1950-х годов все чаще к Glossotherium . Paramylodon имеет множество общих черт, которые предполагают близкое родство с Glossotherium . Только с 1990-х годов оба рода стали считаться отдельными, а Glossotherium обитает только в Южной Америке. , а парамилодон населял Северную Америку.
Вид Paramylodon harlani известен по останкам, найденным по всей Северной Америке, а многочисленные останки известны из смоляных ям Ла Бреа в Калифорнии.
Как и у некоторых других милодонтидов, у парамилодона были встроены остеодермы в коже . Парамилодон обитал на открытых ландшафтах, а иногда и в горных районах, и питался травоядными или смешанными кормами. Известны сохранившиеся следы. Морфология передних конечностей позволила предположить, что парамилодон мог рыть норы.
Как и другие наземные ленивцы, парамилодон вымер около 14–12 000 лет назад в результате вымирания мегафауны большинства крупных млекопитающих в Америке в позднем плейстоцене. Парамилодон перекликался во времени с палеоиндейцами , самыми ранними человеческими обитателями Америки, которые, возможно, охотились на парамилодона . Его исчезновение может быть результатом изменения климата, охоты или сочетания обоих факторов.
Таксономия
[ редактировать ]Paramylodon — вымерший род ленивцев из вымершего семейства Mylodontidae . Mylodontidae сгруппированы вместе с современными двупалыми ленивцами семейства Choloepodidae и вымершими Scelidotheriidae в надсемейство Mylodontoidea, причем первое семейство является их ближайшими ныне живущими родственниками. [ 1 ]
Парамилодон обычно считается близким родственником как Mylodon , так и Glossotherium . [ 2 ] [ 3 ] [ 4 ] Напротив, исследование, представленное в 2019 году Лучано Варелой и другими участвующими учёными и включающее многочисленные ископаемые формы всего подотряда ленивцев, частично поставило под сомнение эту точку зрения. В этом исследовании Paramylodon и Glossotherium было обнаружено, что тесно связаны, Mylodon , с другой стороны, составляет основу продвинутых клад милодонтов и лестодонов с некоторыми формами из северной части Южной Америки. [ 5 ] В том же году более подробный филогенетический анализ милодонтов был опубликован исследовательской группой под руководством Альберто Боскаини . Согласно исследованию, Paramylodon , Glossotherium и Mylodon образуют более близкие отношения внутри Mylodontinae. [ 6 ] Эта точка зрения также находит поддержку в вышеупомянутых биохимических данных, также представленных в 2019 году. [ 1 ] Подробный морфологический анализ, опубликованный еще в 2009 году Робертом К. Макафи, также предполагает, что парамилодон и глоссотерий очень тесно связаны и имеют близкого общего предка. [ 7 ]
Ниже представлено филогенетическое древо Mylodontidae, основанное на работе Boscaini et al. . 2019 г., показано положение парамилодона . [ 6 ]
История исследований
[ редактировать ]Открытия в Северной и Южной Америке.
[ редактировать ]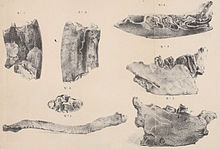

История исследований Paramylodon сложна и характеризуется более чем 150-летней путаницей с Mylodon и Glossotherium . История начинается с первых открытий Ричарда Харлана (1796-1843) в Биг-Боун-Лик в округе Бун в американском штате Кентукки в 1831 году, которые включали правую нижнюю челюсть и ключицу . Харлан признал, что это останки вымершего ленивца, и отнес их к Megalonyx , который в то время уже был известен в Северной Америке, а внутри рода — к виду Megalonyx laqueatus , который он установил незадолго до этого. [ 8 ] [ 9 ] Первоначально находки хранились в Нью-Йорке, но сейчас утеряны. [ 10 ]
Между 1831 и 1836 годами Чарльз Дарвин совершил свое новаторское путешествие на корабле HMS Beagle в Южную Америку и привез оттуда большое количество окаменелостей. Затем их изучил Ричард Оуэн , один из самых важных исследователей викторианской эпохи, и результаты опубликовали. В первой публикации об останках млекопитающих в целом в 1840 году он представил род Mylodon с видом Mylodon darwinii . Род и вид были основаны на нижней челюсти Дарвина, найденной в Пунта-Альта в аргентинской Буэнос-Айрес провинции . В качестве особой характеристики выделялось в общей сложности четыре коренных зуба в каждом зубном ряду. В то же время Оуэн также отметил сходство в строении зубов между нижней челюстью Харлана и Mylodon darwinii . Сделав это вывод, он отказался от названия, придуманного Харланом, Megalonyx laqueatus , и создал новый вид, Mylodon harlani . [ 11 ] Название рода Paramylodon состоит из греческих слов παρα (para означает «рядом» или «рядом»), μύλη ( «myle » « моляр ») и ὀδούς ( «odús » «зуб»), что переводится как «коренной зуб». Два года спустя Харлан прокомментировал использование этого названия, поскольку, по его мнению, оно не описывало каких-либо выдающихся характеристик животного и могло означать любое вымершее млекопитающее, поскольку почти у всех из них были задние коренные зубы. [ 12 ]
В том же 1842 году Оуэн представил подробное описание скелета милодонта, добытого из пойм реки Рио-де-ла-Плата к северу от Буэнос-Айреса ; он установил для него новый вид Mylodonrobustus . [ 13 ] Таким образом, на тот момент род Mylodon состоял из трех видов, два из которых встречались в Южной Америке и один в Северной Америке. Кроме того, должно оказаться проблематичным то, что Оуэн определил Mylodon darwinii как типовой вид рода, хотя, как он признал, это был второй известный и описанный вид после Mylodon harlani . Соответственно, Mylodon harlani фактически имел бы право на статус типового вида . были отнесены разные типовые виды В дальнейшем к Mylodon , например, Иоганнес Теодор Рейнхардт в 1879 году считал его Mylodonrobustus , Рихард Лидеккер в 1887 году, напротив, считал его Mylodon harlani . [ 14 ] [ 7 ] [ 15 ]
Парамилодон и милодон - глоссотериев проблема
[ редактировать ]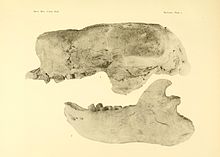


Внутри рода Paramylodon только один вид — P. harlani признан . Другой вид, P. nebrascensis , был описан в 1903 году Барнумом Брауном на основе частичного скелета из Хей-Спринг в Небраске , [ 16 ] но уже в 1920-х годах был синонимом типового вида . Лишь десять лет спустя Гловер Моррилл Аллен создал вид Mylodon garmani с помощью еще одного частичного скелета из реки Ниобрара в Небраске. [ 17 ] но это также считается синонимом Paramylodon harlani . То же самое относится и к нескольким видам, названным Эдвардом Дринкером Коупом еще в 1870-х и 1890-х годах, таким как Mylodon sodalis и Mylodon sulcidens . [ 10 ] Первоначальное подразделение на два подвида: P. h. harlani для робуста и P. h. tenuiceps для изящной формы, как предположил Честер Сток в 1917 году, [ 18 ] [ 19 ] сегодня больше не пропагандируется. [ 20 ] Однако вид « Glossotherium » chapadmalense является проблематичным. Этот вид был первоначально идентифицирован в 1925 году Лукасом Краглиевичем по почти неповреждённому черепу длиной 39 см с нижней челюстью из слоев среднего плиоцена к востоку от Мирамара в аргентинской провинции Буэнос-Айрес . [ 21 ] Он имеет сходство с Glossotheriumrobustum , но также обладает индивидуальными отличиями, которые могут оправдывать его собственный родовой статус; название « Eumylodon » (которое Краглиевич уже использовал для Eumylodon chapadmalense например, было предложено в 1925 году). Таким образом, эта форма могла быть общим предком Glossotherium и Paramylodon . Однако применимо ли это также к североамериканским находкам из плиоцена Флориды и Мексики, впервые перечисленным под тем же названием вида Джесси С. Робертсоном в 1976 году, или они ближе к Paramylodon , в настоящее время неясно из-за отсутствия сравнительных исследований. [ 7 ] Частично ранние останки милодонтов также указаны как P. garbanii , видовое название, данное в 1986 году некоторым плиоценовым останкам нижней челюсти и конечностей из Арройо Э.И. Танке в мексиканском штате Гуанахуато (под научным названием Glossotherium garbanii ). [ 22 ] [ 23 ] Однако этот вид не полностью признан, и другие авторы считают его синонимом Glossotheridium/Glossotherium chapadmalense . [ 24 ]
В 1903 году Барнум Браун (1873-1963) ввел родовое название Paramylodon . Для этой цели он использовал частичный скелет из Хей-Спринг в Небраске , обнаруженный в 1897 году во время экспедиции Американского музея естественной истории . К роду он отнес Paramylodon nebrascensis как вид. В качестве определяющего отличия от североамериканского Mylodon harlani , которого Браун считал типовым видом Mylodon , он указал отсутствие передних клыковидных зубов на верхней челюсти. Таким образом, в плейстоцене Северной Америки в это время были признаны два самостоятельных представителя Mylodonts. [ 16 ] [ 10 ]
Позднее Честер Сток (1892-1950) на основе своих исследований материала находок на ранчо Ла Бреа указал, что особенность отсутствия верхних передних зубов весьма вариабельно развита у Mylodon harlani . Поэтому в качестве синонима он заменил P. nebrascensis на Mylodon harlani в 1917 году. [ 18 ] [ 19 ] Однако в 1928 году Лукас Краглиевич ограничил находки в Северной Америке только Paramylodon и, таким образом, отделил этот род от его южноамериканских представителей. [ 25 ] мнение, которое восемь лет спустя поддержал Анхель Кабрера, но в последующий период оно не нашло большого отклика среди большинства исследователей. Краглиевич тем же шагом также пересмотрел Glossotherium как независимый род, который следует отличать от Mylodon . [ 25 ] Glossotherium также первоначально был помещен Оуэном в его статье 1840 года об открытиях Дарвина на основе фрагмента черепа из Арройо Саранди на юго-западе современного Уругвая , но только два года спустя он объединил его с Mylodon . [ 11 ] [ 13 ]
В дальнейшем, после Краглиевича и Кабреры, Glossotherium по принципу приоритета наименования превратился в таксон «мусорной корзины» , в который поместили многочисленные близкородственные формы. Джордж Гейлорд Симпсон выразил в своей общей таксономии млекопитающих 1945 года, что если бы парамилодон не мог быть четко отделен от глоссотерия , то глоссотерий был бы предпочтительнее из-за того, что он имел приоритет над парамилодоном . [ 26 ] С последующим полным включением Paramylodon в этот род, осуществленным Робертом Хоффстеттером в 1952 году, Glossotherium оказался среди немногих форм ленивцев, встречавшихся в Южной и Северной Америке, но в результате он также обладал высокой изменчивостью. Многочисленные исследователи ХХ века придерживались точки зрения синонимии двух родов ленивцев. [ 27 ] Однако в 1995 году Х. Грегори Макдональд снова отделил североамериканский парамилодон от южноамериканского глоссотерия . При этом он отметил, что на сегодняшний день не было проведено исследований, которые бы продемонстрировали, что эти два рода на самом деле являются синонимами. Фактически, присутствие Paramylodon в Северной Америке скорее отличало бы его от Glossotherium , который, в свою очередь, известен только из Южной Америки. [ 10 ] В дальнейшем было представлено несколько исследований черепов, которые четко отличали два рода и плюс Mylodon друг от друга. [ 7 ] [ 28 ] Некоторые недавние исследования поставили под сомнение принадлежность вида P. garbanii к этому роду. [ 29 ]
Описание
[ редактировать ]
Парамилодон имел длину около 3 метров (9,8 футов), высоту в плечах 1,8 метра и весил до 1,5 метрических тонн. [ 30 ] Он известен из месторождений Северной Америки, в том числе в Мексике и США, а также на юге, вплоть до Гватемалы . Для парамилодона характерны кожные косточки — маленькие кости, встроенные в кожу, что, по-видимому, добавляет животному определенную степень защиты. Эта характеристика также свойственна южноамериканскому милодону . [ 31 ] [ 32 ] [ 33 ] [ 34 ] [ 35 ] [ 36 ]
парамилодона Численность в первую очередь обусловлена находками на ранчо Ла Бреа в Калифорнии . Извлеченный там материал от нескольких десятков человек послужил основой многочисленных расследований, на которых во многом основаны следующие описательные сведения. [ 18 ] [ 37 ] [ 19 ] Этот род был средним представителем Mylodontidae . Полностью реконструированный скелет из Ранчо Ла Бреа имеет общую длину 279 см, из которых около 118 см занимает хвост. В плечах достигает высоты 112 см, а в тазе 122 см. Парамилодон весит около 1,39 тонны , но раньше милодонтиды были значительно меньше. [ 38 ] В целом парамилодон представлял собой животное крепкого телосложения. Он характеризовался удлиненным черепом, короткой шеей, коротким и компактным телом с широким тазом, сильными конечностями и хвостом. [ 38 ]
Характеристики черепа и зубных рядов
[ редактировать ]Характеристики черепа
[ редактировать ]
Череп парамилодона был длинным и узким. Его типичная общая длина составляла от 42,9 до 49,8 см (от 0,429 до 0,498 м). Особенно большой череп имел размеры 54,0 см. [ 39 ] На простом виде он имел довольно прямоугольную форму со средней шириной у затылочной кости 18,8 см, за глазами 12,2 см и рыла 14 см. Типичным для многих милодонтов было постоянно расширяющееся вперед рыло. Череп, однако, оказался в целом намного уже, чем у Glossotherium сопоставимого размера , причем последний имел куполообразную выпуклость на передней линии при виде сбоку, чего не было у Paramylodon . Однако череп Paramylodon , за исключением средней области (куполообразная выпуклость у Glossotherium ), был в среднем выше, его размеры составляли около 13,8 см на затылке и 13 см на рыле. Носовая кость латерально соприкасалась с верхней челюстью. В результате внутренняя часть носа была закрыта по бокам и открыта только спереди, а высота стала примерно такой же, как и ширина, что было связано с общим более узким черепом. Диастема , типичная для ленивцев, лишь слабо соединялась с верхней челюстью. У лобной кости носовая кость выступала далеко назад, так что шов между двумя костями черепа имел скорее V-образную форму. Кроме того, лобная кость представляла собой самую крупную кость всего черепа. Сильный существовал теменный гребень между теменными костями , но он казался значительно уже, чем у Glossotherium . Скуловые дуги снова вторично закрылись, в отличие от большинства ленивцев. Передняя дуга, берущая начало от скуловой кости и направленная назад, имела три отростка: один ориентирован вверх, один вниз и средний горизонтально. Задний отдел дуги, прикрепленный к височной кости , имел пальцевидную форму и присоединялся к срединному отростку переднего отдела дуги. На нижней стороне черепа небная кость выступала гораздо дальше назад, у Paramylodon чем у Glossotherium , что вызвано более длинным расширением кости за последним коренным зубом. Как и у многих милодонтов, обе стороны костей крыльев были заметно вздуты. Однако у Paramylodon это было не так очевидно, как у Glossotherium , поэтому расширенные структуры располагались гораздо дальше друг от друга через базальный феноид клиновидной кости . [ 18 ] [ 19 ] [ 7 ] Особенности, которые связывают Glossotherium и Paramylodon , включают, например, строение зубного ряда с передними клыкообразными зубами и строение зубов, например, второго моляра, или положение костного шва между небной костью и верхней челюстью возле самого заднего зуба. Напротив, Mylodon более отчетливо расходится: у него уменьшенный зубной ряд, более просто устроенные зубы и переднее костное соединение между небной и верхней челюстью. [ 7 ]
Стоматологические характеристики
[ редактировать ]Нижняя челюсть достигала длины от 31,5 до 43,6 см (от 12,4 до 17,2 дюйма). Он был массивным и широким. Горизонтальное тело кости непрерывно увеличивалось в высоту от переднего к заднему ряду, а ниже самого заднего зуба его высота составляла до 10,5 см. [ 40 ] Крепкий симфиз вырос до 11 см (4,3 дюйма) и был типичным для милодонтов. Он вытянут вперед, что свойственно почти всем ленивцам. Это ложковидное расширение симфиза не так отчетливо выступало вбок у Paramylodon, как у Glossotherium , так что боковые края были скорее прямыми и менее отчетливо изогнутыми, чем у последнего. Ширина симфиза в переднем отделе до 15 см. Суставной отросток лишь незначительно выступал над жевательной плоскостью, а венечный отросток располагался значительно выше. Его передний край проходил по прямой линии у Paramylodon , отклоняясь от изогнутого дизайна у Glossotherium . Зубной ряд состоял, как обычно у ленивцев, из 5 зубов на верхнюю половину челюсти и 4 зубов на нижнюю половину челюсти, всего образовалось 18 зубов. Передние зубы имели клыковидную (caniniform) форму, остальные — коренные (molariform). Строение зубного ряда ленивцев считается филогенетически примитивным. Однако у более поздних представителей У парамилодона верхние клыковидные зубы часто были редуцированы, так что зубной ряд тогда состоял всего из 16 зубов. Подобная редукция зубов у Glossotherium не известна . У Mylodon , напротив, самые передние зубы верхнего зубного ряда также уже не были развиты, но нижние клыковидные зубы напоминали задние коренные зубы. имели овальное поперечное сечение Клыкообразные зубы у Paramylodon и были загнуты назад. Однако они не достигали таких размеров, как у Glossotherium или даже у Lestodon . короткая диастема До заднего ряда зубов существовала . Коренные зубы, похожие на коренные, имели плоскую форму с несколько приподнятым краем. Они имели двудольную форму с сильным срединным сужением, за исключением первого моляра верхней челюсти, который имел более прямоугольную форму и образовывал самый длинный зуб на верхней челюсти со средней длиной 3,7 см (1,5 дюйма). лопастная структура была гораздо более заметной, чем у Glossotherium . Все зубы обычно отсутствовали эмаль , вместо этого состоящая из более твердого варианта дентина ( ортодентина ) с дополнительным наружным слоем зубного цемента. Доля ортодентина у парамилодона достигала 28%. [ 41 ] Длина верхнего ряда зубов составляла в среднем 14,4 см, из них задние коренные зубы занимали 12,6 см. Из-за расширенного вперед рыла зубные ряды расходились друг от друга. [ 18 ] [ 19 ] [ 7 ]
Скелетные характеристики
[ редактировать ]
Обширный материал находок на Ранчо Ла Бреа позволяет провести полную реконструкцию скелета тела. Позвоночник состоял из 7 шейных, 16 грудных, 8–9 поясничных, крестцовых и 21 хвостового позвонка. была Плечевая кость массивной, длина — 46 см, голова особо явно не выделялась. Выступающий костный гребень (дельтопекторальный пах) прикреплен к стержню плечевой кости, но в верхней части он менее выражен, чем у Glossotherium . Нижний конец сустава широко выступал вбок. Энтепикондилярное отверстие , иногда встречающееся у некоторых ленивцев, здесь не развито. Локтевая кость имела значительно расширенный верхний суставной отросток — локтевой отросток . Он вырос примерно до 20 см в длину, а вся кость достигла 40 см в длину. Конструкция локтевой кости оказалась короче и крепче, чем у Glossotherium , стержень был широким и суженным сверху спереди и сзади. Точно так же радиус был коротким и массивным, длиной 29,6 см. Самая длинная кость представлена бедра около 54,6 см. Очень короткие экземпляры с Ранчо Ла Бреа имели длину всего 51 см, очень длинные — 58 см. Плоский и широкий дизайн, типичный для наземных ленивцев, поражал, так что кость выглядела почти как доска. Голова лишь слегка возвышалась над поверхностью и имела более внутреннее положение. Стержень был слегка повернут внутрь, а третий вертел как место прикрепления мышц, появившийся у Лестодона не был виден , у Парамилодона . При длине 24,6 см большеберцовая кость была значительно короче бедренной. Это типичная особенность милодонтов, у преимущественно поздних представителей которых нижний отдел задней ноги часто достигал лишь примерно половины длины верхней. [ 42 ] В случае Paramylodon длина голени составляла 45% длины бедра. Его стержень был уплощен, как и бедренная кость, и также имел небольшой изгиб. Верхний конец сустава выступал латерально, ширина здесь достигала примерно трёх четвертей длины всей кости. Малоберцовая кость не срасталась с большеберцовой костью, достигая 26,3 см в длину. [ 19 ] [ 16 ]

Руки и ноги имели сходное строение, как и у других крупных милодонтов Glossotherium и Lestodon , отклонения присутствуют в деталях. Всего на руке было пять лучей (от I до V), и только три внутренних луча (от I до III) имели когти. Пястная кость первого луча срослась с многоугольной костью и образовала единое целое, что часто наблюдается у наземных ленивцев (так называемый пястно-запястный комплекс или MCC). Пястные кости третьего-пятого лучей были массивными и имели длину более 10 см, причем наиболее прочной конструкцией обладал IV луч. У I луча пальца первые две фаланги дополнительно срослись , у II и III лучей имелось в каждом случае по три фаланги, из которых первые две имели, однако, явно уменьшенную длину. Соответствующие концевые члены трех внутренних лучей имели расширенные когтевые отростки, что предполагает соответственно большие когти. Длина варьировала от внутренней (I) до наружной (III) от 7,5 см до 15,4 см и до 17,4 см, высота – от 2,9 до 5,7 см. Наружные пальцы без когтей имели сильно уменьшенные в размерах фаланги. Подножье парамилодона Всего у было четыре луча (от II до V), самый внутренний луч был полностью редуцирован. Когти существовали здесь только на II и III пальцах ног, которые также были наиболее развиты. Однако плюсны здесь имели довольно короткую длину — 3,6 и 6,5 см соответственно, у наружных лучей они становились длиной более 11,0 см каждый и были очень массивными. Как и у двух других милодонтов, второй луч имел всего две фаланги, так как первая и вторая фаланги были слиты в одно целое, соответствующее руке. В отличие от Glossotherium и Lestodon , третий луч Paramylodon также часто состоял только из двух конечностей. Соответствующие концевые фаланги с когтями имели аналогичную руке чрезвычайно прочную конструкцию, только когтевой отросток здесь составлял около 8,5 см на втором и 11,1 см на третьем луче и достигал в каждом случае высоты 3,3 и 3,9 см. Наружные лучи обладали противоположными к этому снова сильно редуцированными концевыми членами. [ 37 ] [ 19 ]
Остеодермы
[ редактировать ]Милодонты — единственная известная линия ленивцев, представители которой имели костные пластинки, так называемые остеодермы , образующиеся в коже, аналогичные современным броненосцам . Однако в отличие от последних они не образовывали у милодонтов прочного костного панциря, а были довольно разбросаны, о чем свидетельствуют находки остатков кожи милодонтов. Несколько сотен остеодерм Paramylodon известны с Ранчо Ла Бреа, [ 43 ] [ 44 ] [ 19 ] кроме того, среди прочего, также в виде плотного слоя на плите из государственного парка Анза-Боррего в Калифорнии. [ 45 ] и из Хайле 15А, богатой окаменелостями известняковой трещины во Флориде . Костные пластинки имели округлую или овальную форму, иногда неправильной формы, длину от 5 до 30 мм. Они имели шероховатую поверхность с неровными углублениями, тогда как нижняя сторона была гладкой и выпуклой. На поперечном разрезе они имели компактную структуру, состоящую из многочисленных пучков волокон, смешанных с твердыми костными пластинками ( остеомами ). В принципе костные пластинки милодонтов имели более простое строение, чем у других ксенартранов . [ 46 ]
Распространение и важные находки ископаемых.
[ редактировать ]
Места обитания и возраст образцов (неполные):
- Участок в заливе Санта-Клара, Сонора, Мексика ~ 4,9–1,8 млн лет назад.
- Местонахождение Три-Бриттон, округ Хендри, Флорида ~ 2,1 млн лет назад — 700 000 лет назад.
- Туринская яма (или яма Эллиотта), округ Монона, штат Айова ~ 1,8–300 000 лет назад.
- Пещера Порт-Кеннеди, округ Монтгомери, Пенсильвания ~ 1,8 млн лет назад — 300 000 лет назад.
- Свалка Фэрмид, округ Мадера, Калифорния ~ 1,8 млн лет назад — 300 000 лет назад.
- Место Рио-де-ла-Пасион, Петен , Гватемала ~ 100 000–11 000 лет назад.
- Место Фанно-Крик, Туалатин , Орегон ~ 15 000–10 000 лет назад.
- Место V Медисин-Хат, Альберта , Канада ~ 1,8 млн лет назад — 11 000 лет назад. [ 47 ]
Обзор и первые события
[ редактировать ]Парамилодон был эндемичен и распространен в Северной Америке и, возможно, также в Центральной Америке . Древнейшие находки, явно относящиеся к этому роду, известны из нижнего плейстоцена . Более старые формы милодонтов происходят из верхнего плиоцена Мексики . [ 24 ] и из американского штата Флорида . Из последнего следует отметить частичный скелет с участка Хайле 15А, расщелины, заполненной отложениями в известняке в округе Алачуа , возраст которой оценивается от 2,1 до 1,8 миллиона лет. Этих ранних представителей обычно называют « Glossotherium» chapadmalensis . [ 27 ] хотя положение внутри рода Glossotherium оспаривается. [ 10 ] [ 7 ] Лишь немногим моложе находки богатой окаменелостями местной фауны Эль-Гульфо из устья реки Колорадо в мексиканском штате Сонора . Они уже помещены в парамилодон и датируются 1,8–1,6 миллионами лет назад. [ 48 ] В целом ископаемые останки нижнего и среднего плейстоцена относительно редки и происходят примерно из 20 местонахождений Северной Америки. Они распространены в основном в южных и центральных районах нынешних Соединенных Штатов и северной Мексики, но также разбросаны по западной части континента вплоть до провинции Альберта в Канаде . Они встречаются как в равнинных, так и в горных местах, причем самая высокая точка обнаружения достигает высоты около 2900 м в Колорадо . [ 49 ] [ 10 ] [ 50 ] Одним из наиболее значительных памятников того периода является яма Лейзи Шелл в округе Хиллсборо во Флориде, где, как сообщается, возраст нескольких черепов и посткраниальных элементов скелета составляет около 1,2 миллиона лет. [ 10 ]
Остатки позднего плейстоцена
[ редактировать ]
Находочный материал верхнего плейстоцена гораздо более обширен и поступает из более чем 100 местонахождений, в Калифорнии только парамилодон известен более чем из 60 местонахождений. Распространение рода аналогично нижнему плейстоцену, но кроме того он встречается несколько восточнее Среднего Запада , например, в Айове . [ 51 ] Самая северная находка этого рода находится в Секиме в штате Вашингтон на 48,1 ° северной широты; на юге этот род также известен по всей Мексике. Некоторые находки теперь указывают на то, что парамилодон , возможно, также жил в Гватемале и Сальвадоре . [ 49 ] [ 10 ] [ 52 ] Среди прочего, находки молодой и взрослой особи были обнаружены на мосту Стивенсона в отложениях ручья Путах-Крик в округе Йоло , Калифорния, и относятся к началу последнего ледникового периода . [ 53 ] Два почти полных скелета были обнаружены в Шонто и Ричвилле в Аризоне , среди немногих известных находок в штате. В целом, ископаемые останки парамилодона очень редки на плато Колорадо на юго-западе США и, кроме того, на северо-западе Мексики, что, возможно, связано с более сухим климатом в этой области в то время. [ 24 ] [ 54 ]
Однако выдающееся мировое значение имеют находки из смоляных ям Ранчо Ла Бреа в южной Калифорнии. Отсюда происходит обширная ископаемая фауна возрастом от 45 000 до 14 000 лет до настоящего времени. Первые находки были обнаружены еще во второй половине XIX века, но гораздо более значительный материал получен благодаря целенаправленным научным исследованиям начала XX века, включавшим в общей сложности более 100 задокументированных памятников. [ 18 ] [ 19 ] [ 55 ] Яркой особенностью фаунистического спектра является необычное доминирование хищников над травоядными. Скорее всего, хищников в большем количестве привлекли животные, застрявшие в асфальте , а затем сами ставшие жертвами естественных ловушек. Среди ленивцев Paramylodon , Megalonyx и Nothrotheriops — три из четырех родов, зарегистрированных в Северной Америке, при этом Eremotherium известен только из восточной части Соединенных Штатов. Тем не менее, парамилодон представляет собой, безусловно, самого многочисленного наземного ленивца в Ла-Бреа: его насчитывается более 70 особей, и среди находок примечательны только 30 черепов. [ 20 ] [ 56 ] Еще один очень обширный комплекс окаменелостей присутствует в местной фауне озера Даймонд-Вэлли в Даймонд-Вэлли и долине Доменигони в округе Риверсайд, а также в южной Калифорнии. Материал добывался при строительстве озера Даймонд-Вэлли с середины 1990-х годов и в настоящее время включает более 100 000 экземпляров более чем 100 таксонов из более чем 2600 различных местностей. В отличие от Ранчо Ла Бреа, здесь доминируют крупные травоядные, а доля крупных хищников невелика. Таким образом, можно сделать вывод о ненарушенном характере фаунистического сообщества. Парамилодон зарегистрирован примерно в 280 отдельных находках, что составляет около 8% от общей фауны млекопитающих. Таким образом, наземный ленивец является пятым по численности представителем млекопитающих в местной фауне озера Алмазной долины после бизонов, лошадей, мастодонта Mammut pacificus и верблюда Camelops . Напротив, два других ленивца Megalonyx и Nothrotheriops , которые также встречаются на территории Ранчо Ла Бреа, гораздо менее многочисленны, и вместе составляют 0,5% от всех обнаружений. Возраст местной фауны озера Даймонд-Вэлли соответствует возрасту ранчо Ла Бреа по данным радиоуглеродное датирование . [ 57 ] [ 58 ]

Палеобиология
[ редактировать ]
Изменение размеров тела и половой диморфизм
[ редактировать ]Как и многие другие группы животных, парамилодон претерпел заметное увеличение размеров тела в ходе своей филогении . Вес представителей нижнего плейстоцена оценивается примерно в 915 кг, однако поздние представители верхнего плейстоцена, вероятно, достигали массы тела до 1,39 т. Основой для соответствующих оценок веса являются бедренные кости , длина которых составляет 48,4 и 54,6 см соответственно. Самые ранние формы из плиоцена , положение которых в пределах рода Paramylodon , однако, широко обсуждается, имели общую массу около 310 кг при длине бедра 35,5 см. Принимая во внимание этих ранних представителей, вес парамилодона увеличился в 4,5 раза за добрые 2,5 миллиона лет. [ 38 ] Особенно поразительно то, что особенно в позднем плейстоцене, во времена Последнего ледникового периода с его чрезвычайно выраженными климатическими колебаниями, практически не наблюдается изменений в размерах, как показывают исследования многочисленных находок на ранчо Ла Бреа, датируемых от 45 000 до 10 000 лет назад. Настоящее время указывает. Это объясняется высокой гибкостью рода по отношению к окружающей среде и, следовательно, высокой приспособляемостью. Однако это предположение игнорирует тот факт, что, согласно правилу Бергмана, все более прохладные условия должны приводить к увеличению размеров тела . [ 59 ]
На основании обширной летописи окаменелостей позднего плейстоцена у Paramylodon можно выделить два морфотипа: изящный вариант и крепкий вариант. Морфотипы не отражаются на общих размерах черепов, а касаются главным образом их выражения, например, соотношения ширины. Также различия можно обнаружить, например, в затылочной кости , которая у робастного варианта вертикальна, а у более изящного — наклонена назад. Таким образом, суставные поверхности для прикрепления шейного отдела позвоночника у последних более выражены, чем у первых. Дальнейшие отклонения обнаруживаются в формировании клыковидных зубов, которые, если они есть, у крепких особей заканчиваются остро, а у изящных – тупо. Возможно, оба морфотипа не являются видами или таксономическими вариациями в смысле подвидов, как первоначально предполагалось, поскольку часто встречаются в одном и том же местонахождении. Скорее, они являются скорее выражением внутривидового полового диморфизма . Однако в настоящее время невозможно отнести морфотип к определенному полу. В 30 известных находках черепов на Ранчо Ла Бреа соотношение крепкого и изящного составляет 3:1; на водохранилище Падение Америки в Айдахо с тремя черепами соотношение составляет 2:1; и в Инглсайде в Техас , тоже с тремя черепами, соотношение 3:0. [ 20 ] Примечательно, что половой диморфизм не отражается на посткраниальном скелете и, следовательно, как отмечалось ранее на примере черепа, не возникает размерного диморфизма в виде значительных различий в длине костей конечностей. Напротив, Eremotherium , который в то же время был также распространен в Северной Америке, но принадлежит к отряду Megatheriidae , как известно, имеет выраженную разницу в размерах между полами. [ 60 ]
Передвижение
[ редактировать ]
В целом для наземных ленивцев предполагается четвероногое передвижение. Однако из-за смещения центра тяжести тела далеко назад они, очевидно, могли переходить и в двуногое положение, сохраняя при этом возможность поддерживать себя с помощью мощного - в отличие от современных древесных ленивцев - очень длинного хвоста. . Задняя стопа парамилодона повернута внутрь, так что основная нагрузка при этом приходится на внешний луч (V). В результате возникает педолатеральная походка, характерная для многочисленных наземных ленивцев, что потребовало значительной перестройки формы и расположения костей предплюсны относительно друг друга, особенно таранной и пяточной костей . У Paramylodon внешний край стопы был немного выгнут вверх, образуя более или менее прямой край, а пяточная кость почти на всю длину соприкасалась с землей. Это согласуется с другими милодонтами, но сильно отличается от близкородственных Scelidotheriidae , у которых была сильно изогнутая ступня, и только задний конец пяточной кости касался земли. [ 61 ] [ 62 ] Еще одна отличительная особенность — задние конечности. Здесь опорно-двигательная система отличается чрезвычайно коротким нижним отделом. У парамилодона нижний отдел достигает менее 50% верхнего. Подобный план построения, присущий почти всем милодонтам, предполагает довольно медленное и громоздкое передвижение. Для сравнения, у мегатерий были значительно более длинные секции нижних конечностей. [ 63 ]
Следы окаменелостей , свидетельствующие о передвижении наземных ленивцев, сохраняются редко. Для парамилодона такие отчетливые шагающие уплотнения были продемонстрированы в тюрьме штата Невада недалеко от Карсон-Сити . Следы были обнаружены еще во второй половине XIX века при добыче песчаника и первоначально, в 1882 году, были интерпретированы как свидетельства существования гигантских людей. Однако к следующему году Отниэль Чарльз Марш признал связь с вымершими наземными ленивцами и начал искать создателя шагающих тюленей среди милодонтов, костные остатки которых также сохранились из того же места. [ 64 ] , были обнаружены многочисленные следы других млекопитающих, таких как мамонты , лошади , лоси , а также хищники Всего в Карсон-Сити, помимо парамилодона , а также птицы . Ступенчатые уплотнения распределены на площади около 8000 м2. 2 Сегодня они в основном прикрыты тюремным строительством, но хорошо задокументированы слепками. Всего у Paramylodon можно было наблюдать десять следов , состоящих из 15–20 отдельных поочередно отпечатавшихся следов. Каждый отдельный след имеет длину от 47 до 51 см и ширину около 20 см. По очертаниям они действительно напоминают человеческие следы, но гораздо более изрезаны по бокам. [ 65 ] Дальнейшие исследования показали, что форма отпечатков очень хорошо соответствует форме стопы парамилодона и что очертания, напоминающие человеческие, обусловлены повернутой наружу стопой ленивца. [ 18 ] [ 66 ] Боковое расстояние между следами составляет около 60 см, что примерно соответствует расстоянию между двумя тазобедренными суставами таза Paramylodon , равно как и длина шага около 146 см, что, в свою очередь, соответствует известной длине задних ног 95 см. см. Поразительно, но при этом сохранились почти исключительно следы задних ног, что первоначально также интерпретировалось как двуногое передвижение животных, аналогичное соответствующим следам окаменелостей Megatherium в Южной Америке. Однако было установлено, что отдельные уплотнения протектора задних лап перекрывают уплотнения передних лап. Поскольку передняя стопа намного меньше задней и располагается иначе, она производит гораздо меньшее впечатление. В отдельных случаях сохранились следы, из которых видно, что передняя часть стопы прикрыта задней. По анатомическим причинам, таким как согнутые ноги при ходьбе, положение и ориентация задней стопы к ноге и тому подобное, постоянное прямохождение крупных наземных ленивцев маловероятно. Длина шага Парамилодон предполагает среднюю скорость от 1,8 до 2,2 м/с, что соответствует установленной скорости мегатерия. [ 24 ]
Исследования лопаток как молодых, так и взрослых особей показывают значительное изменение формы: от более округлой у первых до преимущественно яйцевидной формы у вторых. Онтогенетический отпечаток сравним с таковым у современных древесных ленивцев. Сходство лопаток молодых представителей парамилодона и детенышей современных ленивцев позволяет предположить сходное поведение. Соответственно, детеныши парамилодона все еще обладали различными способностями к лазанию и, возможно, цеплялись за своих матерей во время транспортировки. [ 67 ]
Диета
[ редактировать ]Привычки питания
[ редактировать ]Милодонты обычно считаются травоядными (зерноядными) из-за их зубов с высокой коронкой и плоскими жевательными поверхностями, похожими на зубы современных травоядных. [ 40 ] Однако отсутствие эмали на зубах ленивцев затрудняет сравнение. О зерноядном рационе судили по особенному формированию зубов. [ 19 ] Анализ жевательного аппарата парамилодона показал, что пища измельчается преимущественно при жевательных движениях вперед, назад и в стороны, о чем также свидетельствуют соответствующие следы перетирания. Этому не противоречат клыковидные передние зубы, которые, если и сформированы, то довольно маленькие. широко развит Нижнечелюстной сустав у парамилодона и имеет неспециализированную поверхность, прилегающая к нему суставная ямка на черепе выглядит неглубокой, что характерно для травоядных животных с их вращательными жевательными движениями. Однако на внутренней стороне имеется дополнительная, почти вертикальная суставная фасетка, закрепленная во впадине на внешней стенке кости крыла . Это имело тенденцию ограничивать слишком сильные боковые жевательные движения. Расположение жевательной мышцы позволяло парамилодону открывать пасть всего на 22°, что значительно меньше, чем у двупалых ленивцев , имеющих, однако, сравнительно более длинные клыковидные зубы. В целом строение жевательного аппарата скорее свидетельствует о предпочтении смешанного растительного питания. [ 68 ] [ 69 ]
Пищевое поведение
[ редактировать ]Строение морды, которая не такая широкая, как у Lestodon , указывает на травяную диету, аналогичную ныне живущим белым носорогам , но значительно шире, чем у Eremotherium и Megatherium , которые оба были браузерами, а это означает, что они ели листья деревьев и кустарников, что сравнимо с таковым у черного носорога , который также является браузером. [ 70 ] [ 69 ] Удлиненный симфиз нижней челюсти выступает далеко за пределы носовой области. , нет Поскольку окостенения носовой перегородки , как у Mylodon , здесь следует предполагать энергичное развитие хряща. Кроме того, язык мог выполнять и опорную функцию во время кормления. Благодаря положению подъязычной кости , которая была смещена далеко назад в черепе, и ее прочной конструкции с сильными мышечными креплениями, подбородочно-подъязычная мышца , например, была особенно сильной и длинной, так что в ней можно было разместить очень подвижный язык. [ 69 ] Поскольку копролиты не известны, у Paramylodon в отличие от Mylodon , остатки пищи не могут быть определены непосредственно. Кроме того, из-за отсутствия эмали детальное изотопное исследование редко возможно. Следовательно, применение таких методов требует отличной сохранности окаменелостей; в случае Paramylodon это было выполнено на зубной кости нескольких зубов из стоянки верхнего плейстоцена в Инглсайде, штат Техас. Результаты, полученные при этом с использованием изотопов углерода , находятся между диапазонами современных травоядных животных, специализирующихся на жестком выпасе или мягком питании растений, тем самым пропагандируя смешанный рацион, но, возможно, с более сильным уклоном в сторону трав. Таким образом, полученные к настоящему времени результаты хорошо согласуются с открытым ландшафтом, в котором обитал парамилодон . Однако существует также гипотеза, что представитель ленивца мог копать корни. Об этом говорят, например, сильные передние ноги, имевшие мощную плечевую кость, широко выступающую на нижнем суставном конце, короткий локтевая кость с длинным расширенным локтевым отростком, образующим массивную мускулатуру предплечья, и несколько уплощенными когтями, что делает их очень подходящими для рытья. Кроме того, очевидны различия в зубном ряду между ранними и поздними представителями рода. Например, формы нижнего плейстоцена обладали еще менее высокими зубными коронками и соответственно более низкой нижней челюстью, а формы верхнего плейстоцена имели значительно более высокие зубы и более массивную нижнюю челюсть. Возможно, это отражает все более сильную адаптацию парамилодона с течением времени. [ 10 ]
Социальное поведение
[ редактировать ]Большинство находок Paramylodon состоят из одиночных особей, массовые скопления, как, например, на Ранчо Ла Бреа, представляют собой скопления на протяжении нескольких тысячелетий. Поэтому можно предположить, что парамилодон , как и современные древесные представители, жил одиночно и в большинстве случаев образовывал материнско-детенышевые группы. Пищеварительная система была, вероятно, устроена так же, как и у современных ленивцев, так что и у парамилодона преобладал довольно медленный обмен веществ с длительным временем прохождения пищи, которая хорошо переваривалась. Предполагаемое медленное передвижение предполагает, что они не участвовали в более крупных сезонных миграциях - в отличие от многочисленных других травоядных, населяющих открытые ландшафты. Согласно этому, животные вели относительно малоподвижный образ жизни. Яркой особенностью многочисленных местонахождений с парамилодоном является частое их совместное присутствие с колумбийским мамонтом и бизоном . Однако оба представителя крупных травоядных млекопитающих демонстрировали разный образ жизни со своими стадными формированиями и длительными миграциями, а также отклонениями в системе пищеварения. Поэтому, Парамилодон, вероятно, использовал другую экологическую нишу , чтобы избежать прямой конкуренции с другими мегатравоядными ландшафтами Северной Америки степными . [ 49 ]
Вымирание
[ редактировать ]Как и большинство других наземных ленивцев, парамилодон вымер в конце позднего плейстоцена в результате вымирания позднеплейстоценовой мегафауны . Однако, в отличие от многих других родов, имеется мало радиометрических данных о Paramylodon, измеренных непосредственно из ископаемого материала. Одна из самых молодых известных калиброванных радиоуглеродных дат относится к местности Гипсовая пещера в Неваде и датируется 14 603–14 213 годами ранее . [ 71 ] [ 72 ] Самая молодая дата с Ранчо Ла Бреа составляет около 13 995–13 763 кал. лет до настоящего времени, самая молодая прямая дата для этого вида по состоянию на 2023 год. [ 73 ] Однако известны еще некоторые возможные недавние находки, немногие из которых были обнаружены на археологических памятниках, связанных с ранним заселением человека на североамериканском континенте. более 130 остеодермов На участке Обри Кловиса в северо-центральном Техасе было зарегистрировано . Почвенный субстрат вокруг находок радиометрически датирован возрастом 12 860 лет назад. Задокументированные там же каменные артефакты , насчитывающие около 9800 штук, также можно отнести к культуре Хлодвига на основании точки Хлодвига . Останки парамилодона , однако, не имеют прямого отношения к ранним поселенцам, поскольку были найдены в близлежащем водоеме, за исключением единственной костной пластинки. [ 74 ] [ 75 ] Также известны останки, одновременные с артефактами культуры Кловис, с памятника Киммсвик в Миссури, хотя на них нет следов порезов. [ 76 ] Из немногих распространенных на сегодняшний день находок неясно, привела ли прямая охота к их исчезновению. [ 74 ] [ 75 ]
На то, что ранние колонизаторы Северной Америки взаимодействовали, преследовали и, возможно, охотились на крупных наземных ленивцев, свидетельствуют следы из Национального памятника Уайт-Сэндс в Нью-Мексико . Здесь несколько сотен следов более крупных ленивцев связаны со следами человека на берегу бывшего озера. В некоторых случаях они перекрываются, а в одном случае человеческие следы лежат внутри следа ленивца. Яркой особенностью пересечения следов ленивца и человека является резкая смена направления первого, что предполагает прямое противостояние причинителей. Однако на этом месте не обнаружено никаких ископаемых останков, а возраст следов до сих пор был датирован лишь косвенно (от 15 560 до 10 000 лет назад). Кроме того, более точно следы ленивцев не определены. Они демонстрируют сильные различия в размерах, которые можно отнести либо к животным разного возраста, либо к разным видам. В рассматриваемый период, помимо парамилодона , нотротериопса , более мелкого наземного ленивца из группа Notthrotheriidae и Megalonyx , крупный род Megalonychidae . В регионе встречаются [ 77 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Саманта Прессли, Грэм Дж. Слейтер, Фрэнсис Пухос, Аналия М. Форазиепи, Роман Фишер, Келли Моллой, Миган Маки, Джеспер В. Олсен, Александр Крамарц, Маттиас Тальоретти, Фернандо Скалья, Максимилиано Лескано, Хосе Луис Ланата, Джон Саутон, Роберт Феранец, Джонатан Блох, Адам Хайдук, Фабиана М. Мартин, Родольфо Салас Джисмонди, Марсело Регеро, Кристиан де Муизон, Алекс Гринвуд, Брайан Т. Чейт, Кирсти Пенкман, Мэтью Коллинз и Росс Д.Е. Макфи: Палеопротеомика решает отношения с ленивцами. Экология природы и эволюция 3, 2019, стр. 1121–1130.
- ^ Годен, Тимоти Дж. (2004). «Филогенетические отношения между ленивцами (млекопитающие, ксенартра, тихоходки): краниодентальные данные» . Зоологический журнал Линнеевского общества . 140 (2): 255–305. дои : 10.1111/j.1096-3642.2003.00100.x . ISSN 1096-3642 . S2CID 38722942 .
- ^ Ринкон, Асканио Д.; Солорсано, Андрес; Макдональд, Х. Грегори; Флорес, Моника Нуньес (01 июня 2017 г.). «Baraguatherium takumara, Gen. et Sp. Nov., самый ранний милодонтоидный ленивец (ранний миоцен) из северной части Южной Америки» . Журнал эволюции млекопитающих . 24 (2): 179–191. дои : 10.1007/s10914-016-9328-y . ISSN 1573-7055 . S2CID 254703922 .
- ^ Брамбилла, Лучано; Ибарра, Дамиан Альберто (2 ноября 2018 г.). «Archaeomylodon sampedrinensis, gen. et sp. nov., новый милодонтин из среднего плейстоцена Пампейского региона, Аргентина» . Журнал палеонтологии позвоночных . 38 (6): e1542308. Бибкод : 2018JVPal..38E2308B . дои : 10.1080/02724634.2018.1542308 . ISSN 0272-4634 . S2CID 91874640 .
- ^ Варела, Лучано; Тамбуссо, П. Себастьян; Макдональд, Х. Грегори; Фаринья, Ричард А. (01 марта 2019 г.). «Филогения, макроэволюционные тенденции и историческая биогеография ленивцев: выводы из байесовского морфологического анализа часов» . Систематическая биология . 68 (2): 204–218. doi : 10.1093/sysbio/syy058 . ISSN 1063-5157 . ПМИД 30239971 .
- ^ Jump up to: а б Боскаини, Альберто; Пужос, Франсуа; Годен, Тимоти Дж. (ноябрь 2019 г.). «Переоценка филогении Mylodontidae (Mammalia, Xenarthra) и расхождение милодонтиновых и лестодонтиновых ленивцев». Зоологика Скрипта . 48 (6): 691–710. дои : 10.1111/zsc.12376 . S2CID 201194980 .
- ^ Jump up to: а б с д и ж г час Макафи, Роберт К. (2009). « Переоценка черепных признаков Glossotherium и Paramylodon (Mammalia: Xenarthra: Mylodontidae) » . Зоологический журнал Линнеевского общества . 155 (4): 885–903. дои : 10.1111/j.1096-3642.2008.00468.x . ISSN 0024-4082 .
- ^ Ричард Харлан: Описание челюстей, зубов и ключицы Megalonyx laqueatus. Ежемесячный американский журнал геологии и естествознания, 1, 1831 г., стр. 74–76.
- ^ Ричард Харлан: Описание челюстей, зубов и ключицы Megalonyx laqueatus. Медико-физические исследования, 1835, С. 334–336.
- ^ Jump up to: а б с д и ж г час я Х. Грегори Макдональд: Гравиградные ксенартраны из раннего плейстоцена, карьер Лейси Шелл 1А, округ Хиллсборо, Флорида. Бюллетень Музея естественной истории Флориды 37, 1995, S. 345–373.
- ^ Jump up to: а б Ричард Оуэн: Ископаемые млекопитающие. В книге Чарльза Дарвина (Hrsg.): Зоология путешествия HMS Beagle под командованием капитана Фицроя в период с 1832 по 1836 год. Часть I. Ископаемые млекопитающие. Лондон, 1840 г., С. 12–111 (С. 63–73).
- ^ Ричард Харлан: Уведомление о двух новых млекопитающих из Брансуик-Кана, Джорджия; с наблюдениями за ископаемыми четвероногими животными Соединенных Штатов. Американский журнал науки и искусства 43, 1842, S. 141–144 ( [1] )
- ^ Jump up to: а б Ричард Оуэн: Описание скелета вымершего гигантского ленивца Mylodonrobustus, Owen, с наблюдениями по остеологии, природному сходству и вероятным местам обитания мегатериоидных четвероногих в целом. Лондон, 1842 г., С. 1–176.
- ^ Ричард Лидеккер: Каталог ископаемых млекопитающих в Британском музее естественной истории. Часть V. Лондон, 1887, С. 1–345 (С. 106).
- ^ Хуан Карлос Ферникола, Серхио Ф. Вискайно и Херардо Де Юлиис: Ископаемые млекопитающие, собранные Чарльзом Дарвином в Южной Америке во время его путешествий на борту HMS Beagle. Revista de la Asociación Geológica Argentina 64 (1), 2009, S. 147–159.
- ^ Jump up to: а б с Барнум Браун: новый род наземных ленивцев из плейстоцена Небраски. Бюллетень Американского музея естественной истории, 29, 1903 г., S. 569–583.
- ^ Гловер М. Аллен: Новый Милодон. Воспоминания Музея сравнительной зоологии 40 (7), 1913, С. 319–346.
- ^ Jump up to: а б с д и ж г Честер Сток: Дальнейшие наблюдения за строением черепа ленивцев-милодонтов с ранчо Ла Бреа. Публикации Калифорнийского университета, Бюллетень факультета геологии 10, 1917 г., стр. 165–178.
- ^ Jump up to: а б с д и ж г час я дж Честер Сток: кайнозойские тяжелозубые неполнозубые животные западной части Северной Америки с особым упором на плейстоценовых Megalonychinae и Mylodontidae с Ранчо Ла Бреа. Карнеги. Вашингтонский институт 331, 1925, С. 1–206.
- ^ Jump up to: а б с Х. Грегори Макдональд: Половой диморфизм в черепе наземного ленивца Харлана. Вклад в науку 510, 2006, С. 1–9.
- ^ Лукас Краглиевич: Четыре новых гравиграды фауны арауканы "Чападмаленсе". Анналы Национального музея естественной истории Буэнос-Айреса 33, 1925, S. 215–235.
- ^ Гэри С. Морган: Фауна позвоночных и геохронология Великого американского биотического обмена в Северной Америке. Бюллетень Музея естественной истории и науки Нью-Мексико, 44, 2008 г., стр. 93–140.
- ^ Марисоль Монтеллано-Баллестерос и Оскар Карранса-Кастанеда: Описание милодонтида из раннего Бланкано центральной таблицы Мексики. Институт геологии Национального автономного университета Мексики, журнал 6, 1986 г., стр. 193–203.
- ^ Jump up to: а б с д Х. Грегори Макдональд: Ископаемая Ксенартра из Мексики: обзор. В: Марисоль Монтеллано Баллестерос и Хоакин Арройо Кабралес (Hrsg.): Достижения в палеомастозоологических исследованиях в Мексике. Серия археологии, Национальный институт антропологии и истории, Кордова, 2002 г., стр. 227–248.
- ^ Jump up to: а б Лукас Краглиевич: « Mylodon darwini» Owen — это генотип вида «Mylodon» Ow. Physis: Журнал Аргентинского общества естественных наук, 9, 1928, S. 169–185.
- ^ Джордж Гейлорд Симпсон: Принципы классификации и классификация млекопитающих. Бюллетень Американского музея естественной истории 85, 1945, С. 1–350 (С. 71).
- ^ Jump up to: а б Джесси С. Робертсон: Последние млекопитающие плиоцена из Хайле XVA, округ Алачуа, Флорида. Бюллетень Музея естественной истории Флориды. 20, 1976, С. 111–186.
- ^ Диего Брандони, Бренда С. Ферреро и Эрнесто Брунетто: Mylodon darwini Owen (Xenarthra, Mylodontinae) из позднего плейстоцена Месопотамии, Аргентина, с замечаниями об индивидуальной изменчивости, палеобиологии, палеобиогеографии и палеоокружающей среде. Журнал палеонтологии позвоночных 30 (5), 2010, с. 1547–1558.
- ^ Брамбилла, Лучано; Толедо, Марсело Хавьер; Аро, Джозеф Август; Агилар, Хосе Луис (ноябрь 2019 г.). «Новый морфотип остеодермы (Xenarthra, Mylodontidae) из среднего плейстоцена Аргентины» . Журнал южноамериканских наук о Земле . 95 : 102298. Бибкод : 2019JSAES..9502298B . дои : 10.1016/j.jsames.2019.102298 . S2CID 201317034 .
- ^ Джефферсон, Джордж Т. (декабрь 1988 г.). «Крупные травоядные млекопитающие позднего плейстоцена: последствия для ранних моделей охоты человека в Южной Калифорнии» . Бюллетень Академии наук Южной Калифорнии . 87 (3): 89–103 . Проверено 27 мая 2023 г.
- ^ Морено, Франческо П.; Вудворд, Артур Смит (1899). «На части кожи млекопитающего, названного Neomylodon listai , из пещеры недалеко от бухты Консуэло, залив Последней Надежды, Патагония». Труды Зоологического общества : 144–156.
- ^ Отто Норденшельд (при участии других авторов): Научные результаты шведской экспедиции в Магеллановы земли 1895-1897 годов под руководством доктора Отто Норденшельда. Том II: Зоология. Стокгольм, 1899, стр. 1–170 (особенно стр. 149–170).
- ^ Лопес-Мендоса, Патрисио; Мена-Ларрен, Франциско (декабрь 2011 г.). «Кожные кости вымерших наземных ленивцев и их роль в тафономических исследованиях пещер: случай Баньо Нуэво-1 (Центральная Андская Патагония, Чили)» . Мексиканский журнал геологических наук . 28 (3): 519–532.
- ^ Вильгельм Бранко: Применение рентгеновских лучей в палеонтологии. Трактаты Королевской прусской академии наук, Берлин, 1906 г., стр. 1–55.
- ^ Хилл, Роберт В. (декабрь 2006 г.). «Сравнительная анатомия и гистология ксенартрановых остеодерм». Журнал морфологии . 267 (12): 1441–1460. дои : 10.1002/jmor.10490 . ПМИД 17103396 . S2CID 22294139 .
- ^ Макдональд, Х. Грегори (декабрь 2018 г.). «Обзор присутствия остеодерм у ленивцев: значение остеодерм как плезиоморфного признака ксенартры». Журнал эволюции млекопитающих . 25 (4): 485–493. дои : 10.1007/s10914-017-9415-8 . S2CID 38600428 .
- ^ Jump up to: а б Честер Сток: Структура стоп Mylodon harlani. Публикации Калифорнийского университета. Вестник геологического отдела № 10 (16), 1917, с. 267–286.
- ^ Jump up to: а б с Х. Грегори Макдональд: Палеоэкология вымерших ксенартранов и Великий американский биотический обмен. Бюллетень Музея естественной истории Флориды 45 (4), 2005 г., с. 313–333.
- ^ TDA Cockerell: ископаемый наземный ленивец из Колорадо. Исследования Университета Колорадо, 6, 1909, S. 309–312.
- ^ Jump up to: а б М. Сусана Барго, Херардо де Юлиис и Серхио Ф. Вискайно: Гипсодонтия наземных ленивцев плейстоцена. Польский палеонтологический журнал 51 (1), 2006, С. 53–61.
- ^ Калтхофф, Даниэла К. (2011). «Микроструктура твердых тканей зубов у ископаемых и современных ксенартранов (Mammalia: Folivora и Cingulata)» . Журнал морфологии . 272 (6): 641–661. дои : 10.1002/jmor.10937 . ПМИД 21456028 . S2CID 23933914 .
- ^ Ринкон, AD; Макдональд, Х.Г.; Солорсано, А.; Флорес, Миннесота; Руис-Рамони, Д. (2015). «Новый загадочный милодонтоидный ленивец позднего миоцена из северной части Южной Америки» . Королевское общество открытой науки . 2 (2): 140256. Бибкод : 2015RSOS....240256R . дои : 10.1098/rsos.140256 . ПМЦ 4448802 . ПМИД 26064594 .
- ^ Джон К. Мерриам: Недавние открытия млекопитающих четвертичного периода в Южной Калифорнии. Наука24, 1906, с. 248–250.
- ^ Уильям Дж. Синклер: Кожные кости парамилодона из отложений асфальта Ранчо ла Бреа, недалеко от Лос-Анджелеса, Калифорния. Труды Американского философского общества 49 (195), 1910, S. 191–195.
- ^ Макдональд, Х. Грегори (2017). «Обзор присутствия остеодерм у ленивцев: значение остеодерм как плезиоморфного признака ксенартры» . Журнал эволюции млекопитающих . 25 (4): 485–493. дои : 10.1007/s10914-017-9415-8 . ISSN 1064-7554 . S2CID 254697023 .
- ^ Хилл, Роберт В. (2006). «Сравнительная анатомия и гистология ксенартрановых остеодерм» . Журнал морфологии . 267 (12): 1441–1460. дои : 10.1002/jmor.10490 . ISSN 0362-2525 . ПМИД 17103396 . S2CID 22294139 .
- ^ База данных палеобиологии: коллекция Medicine Hat Unit V.
- ^ Фред В. Кроксен III, Кристофер А. Шоу и Дэвид Р. Сассман: геология плейстоцена и палеонтология дельты реки Колорадо в заливе Санта-Клара, Сонора, Мексика. В: Роберт Э. Рейнольдс (Hrsg): Дикое, живописное и быстрое путешествие по желобу реки Колорадо. Путеводитель по симпозиуму по пустыне 2007 года и отрывки из его докладов. Калифорнийский государственный университет, 2007, с. 84–89.
- ^ Jump up to: а б с Х. Грегори Макдональд и Стив Пеликан: Мамонты и милодонты: экзотические виды с двух разных континентов в фауне плейстоцена Северной Америки. Четвертичный Интернационал 142/143, 2006, С. 229–241.
- ^ Х. Грегори Макдональд, Роберт Г. Дандас и Джеймс К. Чаттерс: Таксономия, палеоэкология и тафономия наземных ленивцев (Xenarthra) из местности на свалке Фэрмид (плейстоцен: ирвингтон) округа Мадера, Калифорния. Четвертичные исследования 79, 2013, с. 215–227.
- ^ Х. Грегори Макдональд: Наземный ленивец Харлана (Paramylodon harlani) (Xenarthra: Mylodontidae) из позднего плейстоцена (ранчолабрей) Айовы. Журнал Академии наук Айовы 119 (1-4), 2012 г., стр. 16–21.
- ^ Карбо-Шанона, Жерар; Хименес-Идальго, Эдвард; Хименес-Морено, Франсиско Х.; Бенитес-Гальвес, Анри (01 апреля 2021 г.). «Новая находка Paramylodon harlani (Owen 1840) (Xenarthra, Pilosa, Mylodontidae) из позднего плейстоцена Вальсекильо, Пуэбла, с комментариями по его палеобиогеографии и палеоэкологии в Мексике» (PDF ) Бюллетень Мексиканского геологического общества . 73 (1):А100720. дои : 10.18268/BSGM2021v73n1a100720 . S2CID 235517769 .
- ^ Роберт Г. Дандас и Лаура М. Каннингем: наземный ленивец Харлана (Glossotherium harlani) и колумбийский мамонт (Mammuthus columbi) из Стивенсон-Бридж, округ Йоло, Калифорния. ПалеоБиос 15 (3), 1993, С. 47–62.
- ^ Х. Грегори Макдональд, Ларри Д. Агенброуд и Кэрол Манганаро Хейден: позднеплейстоценовый милодонт-ленивец Paramylodon harlani (Mammalia: Xenarthra) из Аризоны. Юго-западный натуралист 49 (2), 2014, С. 229–238.
- ^ Энтони Р. Фриша, Блэр ван Валкенбург, Лилиан Спенсер и Джон Харрис: Хронология и пространственное распределение костей крупных млекопитающих в яме 91, ранчо Ла Бреа. Палео 23 (1), 2008 г., стр. 35–4.
- ^ Лесли Ф. Маркус: Перепись многочисленных крупных млекопитающих плейстоцена на Ранчо Ла Бреа. Вклад в науку 38, 1960, С. 1–11.
- ^ Кэтлин Спрингер, Эрик Скотт, Дж. Кристофер Сейджбил и Линдон К. Мюррей: Местная фауна озера Даймонд-Вэлли: позвоночные позднего плейстоцена из внутренней части Южной Калифорнии. Бюллетень Музея Северной Аризоны 65, 2009 г., стр. 217–236.
- ^ Кэтлин Спрингер, Эрик Скотт, Дж. Кристофер Сейджбил и Линдон К. Мюррей: Динамика фауны крупных млекопитающих позднего плейстоцена из внутренней части южной Калифорнии: местная фауна озера Даймонд-Вэлли. Четвертичный Интернационал 217, 2010, С. 256–265.
- ^ Дональд Р. Протеро и Кристина Р. Раймонд: Стазис наземных ленивцев позднего плейстоцена (Paramylodon harlani) из смоляных ям Ранчо Ла Бреа, Калифорния. Бюллетень Музея естественной истории и науки Нью-Мексико, 53, 2011 г., стр. 624–628.
- ^ Дональд Р. Протеро и Кристина Р. Раймонд: Вариации и половой диморфизм размеров у плейстоценовых наземных ленивцев (Xenarthra). Бюллетень 44 Музея естественной истории и науки Нью-Мексико, 2008 г., стр. 331–333.
- ^ Х. Грегори Макдональд: Биомеханические выводы о передвижении наземных ленивцев: интеграция морфологических данных и данных о следах. Бюллетень 42 Музея естественной истории и науки Нью-Мексико, 2007 г., стр. 201–208.
- ^ Х. Грегори Макдональд: Эволюция педолатеральной стопы у наземных ленивцев: закономерности изменений в астрагале. Журнал эволюции млекопитающих 19, 2012, с. 209–215.
- ^ Де Юлиис, Жерар; Ре, Уильям Х.; Вискаино, Серджио Ф. (25 марта 2004 г.). «Мегатериина Черного Быка (Mammalia, Xenarthra): новый вид Pyramiodontherium и обзор Plesiomegatherium » Журнал палеонтологии позвоночных . 24 (1): 214–227. Бибкод : 2004JVPal..24..214D . дои : 10.1671/17.1 . ISSN 0272-4634 . S2CID 85178982 .
- ^ Отниэль Чарльз Марш: О предполагаемых человеческих следах, недавно найденных в Неваде. Американский журнал науки 152, 1883, S. 139–140.
- ^ Джозеф ЛеКонте: Следы Карсона. Природа 1883, С. 101–102.
- ^ Честер Сток: Происхождение предполагаемых человеческих следов в Карсон-Сити, Невада. Наука 51, 1920, с. 514
- ^ Грасс, Энди Д. (2 января 2019 г.). «Вывод о различном поведении между взрослыми и молодыми особями гигантских наземных ленивцев на основе морфологии лопаток» . Журнал палеонтологии позвоночных . 39 (1): e1569018. Бибкод : 2019JVPal..39E9018G . дои : 10.1080/02724634.2019.1569018 . ISSN 0272-4634 . S2CID 132603649 .
- ^ Вирджиния Л. Неаполь: Механизм питания плейстоценового наземного ленивца Glossotherium. Вклад в науку Музей естественной истории округа Лос-Анджелес 415, 1989, стр. 1–23.
- ^ Jump up to: а б с М. Сусана Барго и Серджио Ф. Вискаино: Палеобиология наземных ленивцев плейстоцена (Xenarthra, Tardigrada): биомеханика, морфогеометрия и экоморфология применительно к жевательному аппарату. Амегиниана 45 (1), 2008, С. 175–196.
- ^ М. Сусана Барго, Нестор Толедо и Серхио Ф. Вискайно: Морда южноамериканских наземных ленивцев плейстоцена (Xenarthra, Tardigrada). Журнал морфологии 267, 2006, с. 248–263.
- ^ Гилмор, Дэниел Макгоуэн (2011). Хронология и экология позднеплейстоценовой мегафауны в Северной долине Уилламетт, штат Орегон (Массачусетс). Портлендский государственный университет.
- ^ Стюарт, Энтони Дж. «Глава 6. Северная Америка: мастодонты, наземные ленивцы и саблезубые кошки». Исчезнувшие гиганты: Затерянный мир ледникового периода . Издательство Чикагского университета. стр. 67–112.
- ^ О'Киф, Ф. Робин; Данн, Риган Э.; Вайцель, Элик М.; Уотерс, Майкл Р.; Мартинес, Лиза Н.; Биндер, Венди Дж.; Саутон, Джон Р.; Коэн, Джошуа Э.; Мичен, Джули А.; ДеСантис, Лариса Р.Г.; Кирби, Мэтью Э.; Геццо, Елена; Колтрейн, Джоан Б.; Фуллер, Бенджамин Т.; Фаррелл, Эслинг Б. (18 августа 2023 г.). «Искоренение мегафауны до раннего дриаса на ранчо Ла Бреа связано с изменением состояния, вызванным пожарами» . Наука . 381 (6659): eabo3594. дои : 10.1126/science.abo3594 . ISSN 0036-8075 . ПМИД 37590347 .
- ^ Jump up to: а б К. Рид Ферринг: Археология и палеоэкология стоянки Обри Кловис (41DN479), округ Дентон, штат Техас. Центр экологической археологии, факультет географии, Университет Северного Техаса, Дентон, 2002, С. 1–276.
- ^ Jump up to: а б Дональд К. Грейсон и Дэвид Дж. Мельцер: Охота на Хлодвига и вымирание крупных млекопитающих: критический обзор доказательств. Журнал мировой предыстории 16 (4), 2002, с. 313–359.
- ^ Редмонд, Брайан Г.; Макдональд, Х. Грегори; Гринфилд, Хаскель Дж.; Берр, Мэтью Л. (март 2012 г.). «Новые доказательства эксплуатации человеком наземного ленивца Джефферсона (Megalonyx jeffersonii) в позднем плейстоцене из северного Огайо, США» . Мировая археология . 44 (1): 75–101. дои : 10.1080/00438243.2012.647576 . ISSN 0043-8243 . S2CID 161436888 .
- ^ Бустос, Дэвид; Джейкуэй, Джексон; Урбан, Томми М.; Холлидей, Вэнс Т.; Фенерти, Брендан; Райхлен, Дэвид А.; Будка, Марцин; Рейнольдс, Салли К.; Аллен, Брюс Д.; С любовью, Дэвид В.; Сантуччи, Винсент Л. (06 апреля 2018 г.). «Следы сохраняют охоту в конце плейстоцена? Взаимодействие человека и ленивца в Северной Америке» . Достижения науки . 4 (4): eaar7621. Бибкод : 2018SciA....4.7621B . дои : 10.1126/sciadv.aar7621 . ISSN 2375-2548 . ПМЦ 5916513 . ПМИД 29707640 .
Внешние ссылки
[ редактировать ]- Мир ленивцев: онлайн-библиография
- Музей Пейджа: информация о наземных ленивцах из смоляных ям Ла Бреа
Пилосан генерирует | |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||


- Доисторические ленивцы
- Доисторические плацентарные роды
- Плиоценовые ксенартраны
- Плейстоценовые ксенартраны
- Плиоценовые млекопитающие Северной Америки
- Плейстоценовые млекопитающие Северной Америки
- Бланкан
- Ирвингтонский
- Ранчолабриан
- Плейстоцен Канада
- Палеонтология в Альберте
- Окаменелости Канады
- Плейстоценовая Калифорния
- Геология округа Лос-Анджелес, Калифорния
- Палеонтология в Калифорнии
- Плейстоценовая Мексика
- Окаменелости Мексики
- Плейстоценовая Гватемала
- Окаменелости Гватемалы
- Ископаемые таксоны описаны в 1903 году.
- Таксоны, названные Барнумом Брауном