Берингийский волк
| Берингийский волк | |
|---|---|

| |
| Две модели берингийских волков, созданные палеохудожниками, работающими в Интерпретативном центре Юконской Берингии. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Плотоядный |
| Семья: | Псовые |
| Род: | Канис |
| Разновидность: | С. волчанка |
| Население: | † Берингийский волк |
Берингийский волк — вымершая популяция волка ( Canis lupus ), жившая во времена ледникового периода . Он населял территорию современной Аляски , Юкона и северной части Британской Колумбии . Некоторые из этих волков дожили до голоцена . Берингийский серого волка и был всесторонне изучен с использованием волк является экоморфом ряда научных методов, что дало новую информацию о его видах добычи и пищевом поведении. Было установлено, что эти волки морфологически отличаются от современных североамериканских волков и генетически являются базальными для большинства современных и вымерших волков. Берингийскому волку не присвоена классификация подвидов, и его связь с вымершим европейским пещерным волком ( Canis lupus spelaeus ) не ясна.
Берингийский волк по размеру был похож на современного аляскинского внутреннего волка ( Canis lupus pambasileus ) и других серых волков позднего плейстоцена , но был более крепким, с более сильными челюстями и зубами, более широким небом и более крупными хищными зубами по сравнению с размером черепа. По сравнению с берингийским волком, более южный лютоволк ( Aenocyon dirus ) был того же размера, но тяжелее и с более крепким черепом и зубными рядами. Уникальная адаптация черепа и зубных рядов берингийского волка позволила ему производить относительно большую силу укуса, бороться с крупной борющейся добычей и, следовательно, сделала возможным хищничество и добычу мусора в плейстоценовой мегафауне . Берингийский волк охотился чаще всего на лошадей и степных зубров, а также на карибу, мамонта и лесного овцебыка.
В конце ледникового периода, с исчезновением холодных и засушливых условий и исчезновением большей части добычи, берингийский волк вымер. Вымирание его добычи объясняют воздействием изменения климата , конкуренцией с другими видами, включая людей, или комбинацией обоих факторов. Местные генетические популяции были заменены другими представителями того же вида или того же рода . Из североамериканских волков выжил только предок современного североамериканского серого волка. Останки древних волков со схожими черепами и зубными рядами найдены в Западной Берингии (северо-восток Сибири). В 2016 году исследование показало, что некоторые волки, ныне живущие в отдаленных уголках Китая и Монголии, имеют общего предка по материнской линии с одним экземпляром восточноберингийского волка возрастом 28 000 лет.
Таксономия
[ редактировать ]Начиная с 1930-х годов представители Американского музея естественной истории работали с Колледжем Аляски и Исследовательской компанией Фэрбенкса над сбором образцов, обнаруженных в ходе гидравлических выемок золота недалеко от Фэрбенкса , Аляска. Чайлдс Фрик был научным сотрудником по палеонтологии Американского музея и работал в районе Фэрбенкса. В 1930 году он опубликовал статью, в которой содержался список «вымерших плейстоценовых млекопитающих Аляски и Юкона». В этот список вошел один экземпляр того, что он считал новым подвидом, который он назвал Aenocyon dirus alaskensis – аляскинский ужасный волк. [ 1 ] Американский музей назвал их типичными видами плейстоцена в Фэрбенксе. [ 2 ] Однако ни типового образца, ни описания, ни точного местоположения предоставлено не было, а поскольку ужасные волки не были обнаружены так далеко на севере, это название позже было предложено как nomen nudum (недействительное). палеонтологом Рональдом М. Новаком [ 3 ] Между 1932 и 1953 годами двадцать восемь волчьих черепов были обнаружены в ручьях Эстер, Криппл, Инжиниринг и Литл-Эльдорадо, расположенных к северу и западу от Фэрбенкса. Считалось, что черепам 10 000 лет. Геолог и палеонтолог Теодор Галуша, который помог собрать коллекции ископаемых млекопитающих Фрика в Американском музее естественной истории, работал над черепами волков в течение ряда лет и отметил, что по сравнению с современными волками они были «короткомордыми». . [ 4 ] Палеонтолог Стэнли Джон Олсен продолжил работу Галуши с короткомордыми волчьими черепами и в 1985 году на основании их морфологии классифицировал их как Canis lupus ( серый волк ). [ 5 ]
Серые волки были широко распространены по всей Северной Америке как в плейстоцене, так и в исторический период. [ 6 ] В 2007 году Дженнифер Леонард провела исследование, основанное на генетическом, морфологическом и стабильном изотопном анализе семидесяти четырех особей берингийского волка с Аляски и Юкона, которое выявило генетические связи, виды добычи и пищевое поведение доисторических волков и поддержало классификацию. этого волка как C. lupus . [ 7 ] [ 8 ] Леонард не присвоил этим экземплярам классификацию подвидов , который называл их «восточно-берингийскими волками». [ 9 ] Подвид, возможно, не был выделен, поскольку родство между берингийским волком и вымершим европейским пещерным волком ( C. l. spelaeus ) не ясно. Берингия когда-то была территорией суши, которая охватывала Чукотское и Берингово моря , соединяя Евразию с Северной Америкой. Восточная Берингия включала территорию современной Аляски и Юкона. [ 10 ]
Родословная
[ редактировать ]Базальный волк
[ редактировать ]Последовательности ДНК можно нанести на карту, чтобы выявить филогенетическое дерево , отражающее эволюционные отношения, где каждая точка ветвления представляет собой расхождение двух линий от общего предка. На этом дереве термин базальный используется для описания линии, образующей ветвь, расходящуюся ближе всего к общему предку. [ 11 ] Генетическое секвенирование волков показало, что берингийский волк является базальным по отношению ко всем другим серым волкам, за исключением современного индийского серого волка и гималайского волка . [ 8 ]
По состоянию на 2020 год самые старые из известных неповрежденных останков волка принадлежат мумифицированному щенку возрастом 56 000 лет назад, который был извлечен из вечной мерзлоты вдоль небольшого притока Ласт-Чанс-Крик недалеко от Доусон-Сити , Юкон, Канада. Анализ ДНК показал, что он принадлежал к кладе берингийских волков , что самый последний общий предок этой клады датируется 86 700–67 500 лет назад и что эта клада была базальной для всех других волков, за исключением гималайского волка. [ 12 ]
Различные генетические типы серого волка
[ редактировать ]| Филогенетическое древо с указанием сроков в годах. Canis lupus [ а ] | |||||||||||||||||||||||||||
|
Гаплотип — это группа генов , обнаруженных в организме и унаследованных вместе от одного из родителей. [ 13 ] [ 14 ] Гаплогруппа — это группа схожих гаплотипов, которые имеют одну мутацию, унаследованную от общего предка. [ 15 ] Митохондриальная ДНК (мДНК) передается по материнской линии и может возникать тысячи лет назад. [ 15 ] В исследовании 2005 года сравнили последовательности митохондриальной ДНК современных волков с последовательностями тридцати четырех особей, датированных периодом с 1856 по 1915 год. Было обнаружено, что историческая популяция обладает вдвое большим генетическим разнообразием , чем современные волки. [ 16 ] [ 17 ] это предполагает, что разнообразие мДНК волков, искорененных на западе США, более чем в два раза превышало разнообразие современной популяции. [ 17 ] В исследовании 2007 года сравнили последовательности мДНК современных волков с последовательностями мДНК берингийских волков. Двадцать берингийских волков дали шестнадцать гаплотипов, которые не удалось обнаружить у современных волков, по сравнению с семью гаплотипами, которые были обнаружены у тридцати двух современных волков Аляски и Юкона. Это открытие указывает на то, что берингийские волки генетически отличались от современных волков. [ 16 ] [ 18 ] и обладали большим генетическим разнообразием, и что когда-то в Северной Америке существовала более крупная популяция волков, чем сегодня. [ 8 ] Современные аляскинские волки произошли не от берингийских волков, а от евразийских волков, мигрировавших в Северную Америку во время голоцена. [ 6 ]
| Филогенетическое древо волков | ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||
| Упрощенная филогения мДНК современных волков и вымерших берингийских волков [ 8 ] [ 19 ] |
В исследовании 2010 года сравнили последовательности мДНК современных волков с последовательностями мДНК 24 экземпляров древних волков из Западной Европы, датированных периодом от 44 000 до 1200 лет назад (YBP). Исследование показало, что последовательности можно разделить на две гаплогруппы. [ 8 ] [ 16 ] Гаплогруппы 1 и 2 можно было найти среди волков по всей Евразии, но только гаплогруппу 1 можно было найти в Северной Америке. Образцы древних волков из Западной Европы отличались от современных волков от 1 до 10 мутаций, и все они принадлежали к гаплогруппе 2, что указывает на ее преобладание в этом регионе на протяжении более 40 000 лет, как до, так и после Последнего ледникового максимума . Сравнение нынешних и прошлых частот показывает, что в Европе гаплогруппа превосходила численность гаплогруппы 2 за последние несколько тысяч лет 1, но в Северной Америке гаплогруппа 2, включая берингийского волка, вымерла и была заменена гаплогруппой 1 после последнего ледникового максимума. . [ 19 ] [ 20 ] Однако исследование 2016 года не подтвердило существование двух гаплогрупп волков. [ 21 ]
Сценарий, согласующийся с данными о филогенетике, размере ледникового щита и глубине уровня моря, заключается в том, что в позднем плейстоцене уровень моря был самым низким. Единственная волна волчьей колонизации Северной Америки началась с открытия Берингова сухопутного моста длиной 70 000 YBP. Это закончилось закрытием Юконского коридора, который проходил вдоль границы между ледниковым щитом Лаврентида и ледниковым щитом Кордильер на высоте 23 000 лет назад во время максимума позднего ледникового периода. Поскольку волки фигурировали в летописи окаменелостей Северной Америки, генетическое происхождение современных волков можно проследить лишь 80 000 лет назад. [ 22 ] [ 23 ] волчьи гаплотипы, которые уже были в Северной Америке, были заменены этими захватчиками либо посредством конкурентного вытеснения , либо посредством генетического примеси . Замена в Северной Америке основной популяции волков более поздней согласуется с результатами более ранних исследований. [ 8 ] [ 23 ] [ 19 ]

Берингийские волки морфологически и генетически сопоставимы с европейскими волками позднего плейстоцена. [ 24 ] Одно исследование показало, что древние волки по всей Евразии имели последовательность мДНК, идентичную шести берингийским волкам (что указывает на общего предка по материнской линии). В число этих волков входили волк из палеолитической стоянки Нерубайское-4, недалеко от Одессы , Украина, датированный 30 000 лет назад, волк из палеолитической стоянки Заскальная-9 в Заскальной на Крымском полуострове , датированный 28 000 лет назад, и «алтайская собака» из Горный Алтай в Центральной Азии датируется 33 000 лет назад. Другой волк из пещеры Выпускек , в Чехии датированный 44 000 лет назад, имел последовательность мДНК, идентичную двум берингийским волкам (что указывает на другого общего предка по материнской линии). [ 8 ] Берингийские волки филогенетически связаны с отдельной группой из четырех современных европейских гаплотипов мДНК, что указывает на то, что как древние, так и современные североамериканские волки произошли из Евразии. [ 8 ] Из этих четырех современных гаплотипов один был обнаружен только у итальянского волка и один - только у волков в Румынии. [ 25 ] Эти четыре гаплотипа, как и гаплотипы берингийских волков, относятся к гаплогруппе 2 мДНК. [ 19 ] Древние экземпляры волков с похожим черепом и зубными рядами были найдены в Западной Берингии (северо-восток Сибири), на Таймырском полуострове , на Украине и в Германии, где европейские экземпляры классифицируются как Canis lupus spelaeus – пещерный волк. [ 26 ] Берингийские волки и, возможно, волки гигантской степи были приспособлены к охоте на ныне вымершие виды благодаря своей уникальной морфологии черепа и зубов. [ 27 ] Этот тип серого волка, приспособленный для охоты на мегафауну, получил название мегафаунального волка . [ 28 ]
Вполне возможно, что панмиктическая (случайно спаривающаяся) популяция волков с потоком генов , охватывающим Евразию и Северную Америку, существовала до закрытия ледниковых щитов. [ 23 ] [ 19 ] [ 29 ] после чего южные волки обособились, и севернее листов существовал только берингийский волк. Сухопутный мост был затоплен морем на 10 000 YBP, а ледниковые щиты отступили на 12 000–6 000 YBP. [ 23 ] Берингийский волк вымер, а южные волки распространились сквозь тающие ледниковые щиты и вновь заселили северную часть Северной Америки. [ 23 ] [ 29 ] Все североамериканские волки произошли от тех, которые когда-то были изолированы к югу от ледниковых щитов. Однако большая часть их разнообразия была позже утрачена в двадцатом веке из-за искоренения. [ 23 ] [ 17 ]
Описание
[ редактировать ]
Олсен описал короткомордые волчьи черепа следующим образом:
Пропорции черепов этих волков различаются в ростральной области. Область черепа, расположенная впереди подглазничного отверстия , заметно укорочена и сужена с боков в некоторых черепах... Вогнутость рострума , если смотреть сбоку, очевидна на всех черепах с коротким лицом, идентифицированных как Canis lupus из золотые прииски Фэрбенкса. Затылочный экземпляров и надзатылочный гребни заметно уменьшены по сравнению с таковыми у средних C. lupus . Затылочный выступ этих гребней, характерный для волков, примерно одинаков в обеих группах C. lupus ... Исследование большой серии недавних черепов волков из Аляски не выявило особей с такими же вариациями, как у черепов из Фэрбенкса. золотые поля. [ 5 ]
Берингийский волк по размеру был похож на современного аляскинского внутреннего волка ( C. l. pambasileus ). [ 8 ] Самые крупные северные волки сегодня имеют высоту в плечах не более 97 см (38 дюймов) и длину тела не более 180 см (71 дюйм). [ 30 ] Средний вес юконского волка составляет 43 кг (95 фунтов) для самцов и 37 кг (82 фунта) для самок. Индивидуальный вес юконских волков может варьироваться от 21 до 55 кг (от 46 до 121 фунта). [ 31 ] с одним юконским волком весом 79,4 кг (175 фунтов). [ 30 ] Берингийские волки также по размеру были похожи на волков позднего плейстоцена, чьи останки были найдены в смоляных ямах Ла Бреа в Лос-Анджелесе , Калифорния. [ 8 ] Эти волки, называемые волками Ранчо Ла Бреа ( Canis lupus ), физически не отличались от современных серых волков, единственными отличиями были более широкая бедренная кость и более длинный бугр большеберцовой кости (место прикрепления четырехглавых мышц и подколенных сухожилий ), что указывает на то, что они имел сравнительно более мощные мышцы ног для быстрого разбега перед погоней. [ 32 ] Берингийский волк был более крепким и обладал более сильными челюстями и зубами, чем ранчо Ла Бреа или современные волки. [ 8 ] [ 16 ]
В позднем плейстоцене более южный лютоволк ( Aenocyon dirus ) имел ту же форму и пропорции, что и юконский волк. [ 33 ] [ 34 ] но подвид лютого волка A. dirus guildayi , по оценкам, весил в среднем 60 кг (130 фунтов), а подвид A. dirus dirus - в среднем 68 кг (150 фунтов), причем некоторые экземпляры были крупнее. [ 35 ] Ужасный волк был тяжелее берингийского волка и обладал более крепким черепом и зубными рядами. [ 8 ]
Приспособление
[ редактировать ]Адаптация – это эволюционный процесс, посредством которого организм становится более приспособленным к жизни в окружающей среде. [ 36 ] Генетические различия между популяциями волков тесно связаны с типом их среды обитания, и волки рассеиваются преимущественно в пределах той среды обитания, в которой они родились. [ 27 ] Было показано, что экологические факторы, такие как тип среды обитания, климат, специализация добычи и конкуренция хищников, сильно влияют на краниодентальную пластичность серого волка , которая представляет собой адаптацию черепа и зубов под влиянием окружающей среды. [ 27 ] [ 37 ] [ 38 ] В позднем плейстоцене различия между местными средами обитания способствовали появлению целого ряда экотипов волков , которые генетически, морфологически и экологически отличались друг от друга. [ 37 ] Термин экоморф используется для описания узнаваемой связи морфологии организма или вида с использованием ими окружающей среды. [ 39 ] Экоморф берингийского волка демонстрирует эволюционную краниодентальную пластичность, не наблюдавшуюся ни у прошлых, ни у современных серых волков Северной Америки. [ 8 ] и был хорошо адаптирован к богатой мегафауне среде позднего плейстоцена. [ 8 ] [ 9 ]
Палеоэкология
[ редактировать ]
Последний ледниковый период , обычно называемый «Ледниковым периодом», длился 125 000 лет. [ 40 ] –14 500 йенских фунтов [ 41 ] и был самым последним ледниковым периодом нынешнего ледникового периода , который произошел в последние годы плейстоценовой эры. [ 40 ] Ледниковый период достиг своего пика во время последнего ледникового максимума, когда ледниковые щиты начали продвигаться на 33 000 YBP и достигли своего максимального предела в 26 500 YBP. Дегляциация началась в Северном полушарии примерно в 19 000 лет назад, а в Антарктиде примерно в 14 500 лет назад, что согласуется с данными о том, что талая ледниковая вода была основным источником резкого повышения уровня моря 14 500 лет назад. [ 41 ] и Беринговский сухопутный мост был окончательно затоплен около 11 000 YBP. [ 42 ] Ископаемые свидетельства со многих континентов указывают на вымирание крупных животных, называемых плейстоценовой мегафауной , ближе к концу последнего оледенения. [ 43 ]
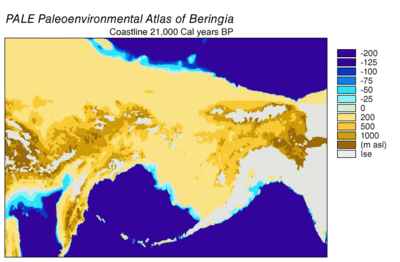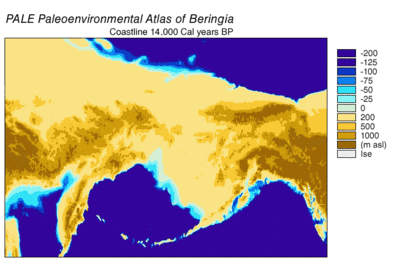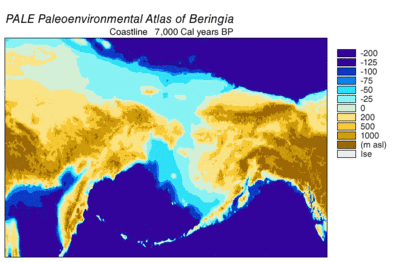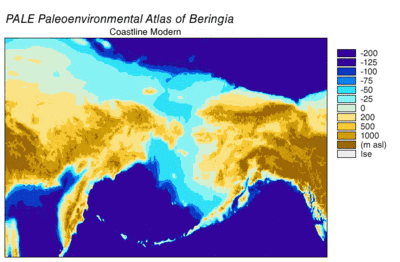
Во время ледникового периода обширная, холодная и сухая гигантская степь простиралась от арктических островов на юг до Китая и от Испании на восток через Евразию и через Берингов мост на Аляску и Юкон, где она была заблокирована Висконсинским оледенением . Сухопутный мост существовал потому, что уровень моря был ниже из-за того, что большая часть воды на планете была заперта в ледниках по сравнению с сегодняшним днем. Таким образом, флора и фауна Берингии были больше связаны с флорой и фауной Евразии, чем с флорой и фауной Северной Америки. [ 44 ] [ 45 ] В восточной Берингии с 35 000 YBP в северных арктических районах температура была на 1,5 ° C (2,7 ° F) выше, чем сегодня, но в южных субарктических регионах было на 2 ° C (3,6 ° F) холоднее. В 22 000 лет назад, во время последнего ледникового максимума, средняя летняя температура была на 3–5 ° C (5,4–9,0 ° F) ниже, чем сегодня, с колебаниями от 2,9 ° C (5,2 ° F) ниже на полуострове Сьюард до 7,5 °. C (13,5 ° F) на Юконе прохладнее. [ 46 ]
Берингия получила больше влаги и прерывистого морского облачного покрова из северной части Тихого океана, чем остальная часть Мамонтовой степи, включая засушливые районы по обе стороны от нее. Влага наблюдалась по градиенту с севера на юг, при этом юг получал наибольшую облачность и влагу из-за воздушного потока из северной части Тихого океана. [ 45 ] Эта влага поддерживала кустарниково- тундровую среду обитания, служившую экологическим рефугиумом для растений и животных. [ 44 ] [ 45 ] В этом берингийском рефугиуме растительность восточной Берингии включала отдельные участки лиственничных и еловых лесов с березой и ольхой . [ 47 ] [ 48 ] [ 49 ] [ 50 ] Эта среда поддерживала крупных травоядных животных , которые были добычей берингийских волков и их конкурентов. Степной бизон ( Bison priscus ), юконская лошадь ( Equus lampei ), шерстистый мамонт ( Mammuthus primigenius ) и дикий як ( Bos mutus ) питались травами, осокой и травянистыми растениями. Карибу ( Rangifer tarandus ) и лесной овцебык ( Symbos cavifrons ) питались тундровыми растениями, включая лишайники, грибы и мхи. [ 10 ]
Добыча
[ редактировать ]
Изотопный анализ может использоваться, чтобы позволить исследователям сделать выводы о рационе изучаемых видов. Два изотопных анализа костного коллагена , выделенного из останков волков позднего плейстоцена, обнаруженных в Берингии и Бельгии, показывают, что волки из обоих регионов охотились в основном на плейстоценовую мегафауну. [ 8 ] [ 19 ] [ 51 ] который стал редким в начале голоцена 12 000 лет назад. [ 19 ] [ 52 ] Берингийский волк охотился чаще всего на лошадей и степных зубров. [ 8 ] [ 18 ] В период, предшествовавший последнему ледниковому максимуму (50 000–23 000 YBP), они также ели лесного овцебыка, а после этого они также ели мамонта. Анализ подтверждает вывод о том, что эти волки были способны убивать и расчленять крупную добычу. [ 8 ]
В другом анализе стабильных изотопов выяснилось, что половина берингийских волков специализируются на овцебыках и карибу, а другая половина — либо на лошадях и бизонах, либо на универсалах. Было обнаружено, что два волка из полноледникового периода (23 000–18 000 лет назад) были специалистами по мамонтам, но неясно, было ли это связано с падальщиками или хищничеством. Анализ окаменелостей других хищников из региона Фэрбенкс на Аляске показал, что мамонт редко встречался в рационе других хищников Берингии. [ 10 ]
Анализ стабильных изотопов мумифицированного детеныша берингийского волка возрастом 56 000 лет назад, который был обнаружен возле реки Клондайк, показал, что большая часть его рациона - и, следовательно, рациона его матери - была основана на водных, а не животных ресурсах. В качестве водных ресурсов было предложено использовать лосося. [ 12 ]
Зубной ряд
[ редактировать ]Исследование Canis зубного ряда , проведенное в 2007 году , показывает, что по сравнению с современным серым волком и плейстоценовым волком Ла Бреа, берингийский волк обладал большими хищными зубами. [ 8 ] и короткое, широкое небо по сравнению с размером черепа. [ 6 ] [ 8 ] Длина ряда премоляров M1, M2 и m1 (нижний хищный) берингийского волка была длиннее, премоляр P4 (верхний хищный) длиннее и шире, а коренные зубы длиннее, чем у двух других типов волков. берингийского волка Короткий и широкий рострум увеличивал силу укуса клыками, одновременно укрепляя череп от стрессов, вызванных борющейся добычей. Сегодня сравнительно глубокие челюсти, подобные челюстям берингийского волка, можно встретить у пятнистой гиены , раскалывающей кости , и у тех псовых, которые приспособлены к поимке крупной добычи. [ 8 ] Берингийские волки обладали краниодентальной морфологией, которая была более специализированной, чем современные серые волки и волки Ранчо Ла Бреа, для захвата, расчленения и поедания костей очень крупных мегатравоядных животных. [ 8 ] [ 18 ] эволюционировав таким образом из-за присутствия мегафауны. [ 53 ] Их более сильные челюсти и зубы указывают на гиперплотоядный образ жизни. [ 8 ] [ 16 ]
Признанным признаком одомашнивания является наличие скученности зубов, при которой ориентация и выравнивание зубов описываются как соприкасающиеся, перекрывающиеся или вращающиеся. Однако исследование 2017 года показало, что у 18% особей берингийского волка наблюдается скученность зубов по сравнению с 9% у современных волков и 5% у домашних собак. Эти экземпляры появились еще до прибытия людей, поэтому возможность скрещивания с собаками невозможна. Исследование показывает, что скученность зубов может быть естественным явлением у некоторых экоморфов волков и не может использоваться для дифференциации древних волков от ранних собак. [ 54 ]

| Зуб переменный | современная Северная Америка | Ранчо Ла Бреа | Восточная Берингия |
|---|---|---|---|
| длина ряда премоляров | 63.4 | 63.6 | 69.3 |
| ширина нёба | 64.9 | 67.6 | 76.6 |
| Длина P4 | 25.1 | 26.3 | 26.7 |
| Ширина P4 | 10.1 | 10.6 | 11.4 |
| Длина М1 | 16.4 | 16.5 | 16.6 |
| Длина М2 | 8.7 | 8.9 | 9.2 |
| длина м1 | 28.2 | 28.9 | 29.6 |
| m1 тригониды длина | 19.6 | 21.9 | 20.9 |
| ширина м1 | 10.7 | 11.3 | 11.1 |
Поломка зуба
[ редактировать ]
Поломка зубов связана с поведением хищника. [ 55 ] Нижние челюсти псовых укреплены позади хищных зубов, что позволяет им раскалывать кости постхищными зубами (коренные зубы M2 и M3). Исследование показало, что современный серый волк обладает большей поддержкой по сравнению со всеми другими существующими псовыми и вымершим ужасным волком. Это указывает на то, что серый волк лучше приспособлен к раскалыванию костей, чем другие псовые. [ 56 ] По сравнению с современными североамериканскими серыми волками среди берингийских волков было гораздо больше особей с умеренно и сильно изношенными зубами и со значительно большим количеством сломанных зубов. Частота переломов у волков варьировалась от минимум 2%, обнаруженных у северного скалистого волка (Canis lupus Incremotus), до максимума, составляющего 11%, обнаруженного у берингийских волков. Распределение переломов по зубному ряду также различается: у берингийских волков частота переломов резцов , хищных и коренных зубов гораздо выше. Похожая картина наблюдалась у пятнистых гиен, что позволяет предположить, что увеличение количества переломов резцов и хищников отражает привычное потребление костей, поскольку кости прогрызаются резцами, а затем раскалываются хищными и коренными зубами. [ 8 ] Риск перелома зубов также выше при захвате и поедании крупной добычи. [ 57 ] [ 58 ]
Конкуренты
[ редактировать ]Помимо берингийского волка, среди других берингийских хищников были евразийский пещерный лев ( Panthera spelaea ), ятаган-зубая кошка ( сыворотка Homotherium) , гигантский короткомордый медведь ( Arctodus simus ) и всеядный бурый медведь ( Ursus arctos ). . [ 10 ] Берингийским волкам пришлось бы конкурировать за туши крупных травоядных со стороны грозного гигантского короткомордого медведя-падальщика. [ 59 ] Кроме того, люди достигли пещер Блуфиш на территории Юкон примерно на 24 000 лет назад, где порезы были обнаружены на экземплярах юконской лошади, степного бизона, карибу ( Rangifer tarandus ), вапити ( Cervus canadensis ) и овцы Далла ( Ovis dalli ). [ 60 ]
Исследование 1993 года показало, что более высокая частота поломки зубов среди хищников плейстоцена по сравнению с ныне живущими хищниками не была результатом охоты на более крупную дичь, о чем можно было предположить, исходя из большего размера первых. Когда добычи мало, конкуренция между хищниками возрастает, заставляя их есть быстрее и потреблять больше костей, что приводит к поломке зубов. [ 55 ] [ 61 ] [ 62 ] По сравнению с современными волками, высокая частота переломов зубов у берингийских волков указывает на более высокое потребление туш из-за более высокой плотности хищников и усиления конкуренции. [ 8 ] Это предложение было оспорено в 2019 году, когда исследование поведения современных волков за последние 30 лет показало, что, когда добычи было меньше, частота переломов зубов увеличивалась более чем вдвое. Это говорит о том, что крупные хищники плейстоцена переживали больше периодов ограниченной доступности пищи по сравнению с их современными собратьями. [ 63 ]
Диапазон
[ редактировать ]
Останки берингийских волков были найдены на Аляске и даже на востоке, вплоть до Юкона в Канаде. [ 9 ] Образцы, идентифицированные по морфологии черепа. [ 9 ] и морфология конечностей [ 64 ] Берингийские волки были найдены в естественной пещере-ловушке у подножия гор Бигхорн в Вайоминге, США. Они были датированы радиоуглеродом между 25 800 и 14 300 YBP, и это место находится прямо к югу от того, что в то время было границей между Лаврентидским ледниковым щитом и Кордильерским ледниковым щитом. Это говорит о том, что между ледниками существовал временный канал от 25 800 YBP. [ 9 ] до наступления ледников 16 000–13 000 YBP. [ 9 ] [ 65 ] Предполагается, что миграция берингийского волка на юг была результатом преследования видов-жертв, поскольку в этой пещере также находились экземпляры степных зубров, мигрировавших из Берингии и ставших добычей волков. [ 9 ] [ 66 ] и овцебык, который, как известно, является важным видом добычи берингийского волка. [ 9 ] [ 10 ] лютые волки отсутствовали к северу от 42° с.ш.; В позднем плейстоцене следовательно, этот регион был бы доступен берингийским волкам для расширения на юг. Нет никаких свидетельств расширения за пределы этого региона. [ 9 ]
Вымирание
[ редактировать ]
Вымирание — результат ликвидации географического ареала вида с сокращением численности его популяции до нуля. Факторы, влияющие на биогеографический ареал и размер популяции, включают конкуренцию, взаимодействие хищник-жертва, переменные физической среды и случайные события. [ 67 ]
Фенотип вымер
[ редактировать ]Фенотип — это любая наблюдаемая и измеримая характеристика организма , включающая в себя любые морфологические, поведенческие и физиологические признаки. [ 68 ] на эти характеристики влияют гены и окружающая среда. [ 69 ] Гигантская степь просуществовала без изменений 100 000 лет, пока не закончилась около 12 000 лет назад. [ 45 ] произошло Вымирание американской мегафауны 12 700 лет назад, когда вымерло 90 родов млекопитающих весом более 44 килограммов (97 фунтов). [ 70 ] [ 61 ] Считается, что исчезновение крупных хищников и падальщиков было вызвано исчезновением добычи мегатравоядных, от которой они зависели. [ 71 ] [ 72 ] Причина вымирания этой мегафауны дискутируется. [ 58 ] но это объясняется воздействием изменения климата , конкуренцией с другими видами, включая людей, или комбинацией обоих факторов. [ 58 ] [ 73 ] Для тех млекопитающих, у которых есть современные представители, древние ДНК и радиоуглеродные данные показывают, что местные генетические популяции были заменены другими представителями того же вида или другими представителями того же рода. [ 74 ]

Послеледниковые изменения окружающей среды на всей территории восточной Берингии привели к массовым изменениям растительности, региональному исчезновению большей части мегафауны и появлению Homo sapiens . [ 46 ] Крупные хищники позднего плейстоцена, которые были более плотоядными, чем их конкуренты, столкнулись с большей уязвимостью перед вымиранием. Берингийский пещерный лев, саблезубая кошка и короткомордый медведь вымерли одновременно со своей крупной мегафауной. всеядный , койот, американский черный медведь бурый медведь, пума и рысь Выжили . И берингийский волк, и лютоволк вымерли в Северной Америке, оставив процветать только менее плотоядной и более грациозной форме волка. [ 8 ] Одна из теорий вымирания утверждает, что берингийский волк был вытеснен и заменен предком современного серого волка. [ 9 ]
Радиоуглеродное датирование останков скелетов 56 берингийских волков показало непрерывную популяцию, насчитывающую более 50 800 YBP. [ 21 ] до 12 500 YBP, за ним следует один волк по цене 7 600 YBP. Это указывает на то, что их популяция начала сокращаться после 12 500 YBP. [ 8 ] хотя добыча мегафауны все еще была доступна в этом регионе до 10 500 YBP. [ 75 ] Время появления этого последнего образца подтверждается обнаружением ДНК мамонта и лошади из отложений, датированных 10 500–7 600 лет назад, из внутренних районов Аляски. [ 75 ] и степной бизон возрастом 5400 лет назад с Юкона. [ 76 ] Время исчезновения лошадей в Северной Америке и минимальная численность популяции североамериканских бизонов совпадают с исчезновением всей гаплогруппы волков в Северной Америке, что указывает на то, что исчезновение их добычи вызвало исчезновение этого волчьего экоморфа. [ 16 ] [ 18 ] Это привело к значительной потере фенотипического и генетического разнообразия внутри вида. [ 8 ]
Гаплотип не вымер
[ редактировать ]Есть части Центральной Евразии, где окружающая среда считается стабильной на протяжении последних 40 000 лет. [ 77 ] В 2016 году в исследовании сравнивались последовательности мДНК древних образцов волков с последовательностями современных волков, включая образцы из отдаленных регионов Северной Америки, России и Китая. Один древний гаплотип, который когда-то существовал как на Аляске (Восточная Берингия, 28 000 лет назад), так и в России (пещера Медвежья, Печорский район, Северный Урал, 18 000 лет назад), был общим для современных волков, обитающих в Монголии и Китае (что указывает на общего предка по материнской линии). ). Исследование показало, что генетическое разнообразие волков прошлого было утрачено в начале голоцена на Аляске, в Сибири и Европе и что совпадение с современными волками ограничено. Исследование не подтвердило две гаплогруппы волков, которые были предложены в более ранних исследованиях. Для древних волков Северной Америки вместо модели вымирания/замены, указанной в других исследованиях, это исследование обнаружило существенные доказательства узкого места (сокращения) популяции , при котором разнообразие древних волков было почти потеряно в начале голоцена. В Евразии утрату многих древних линий невозможно просто объяснить, и, по-видимому, она происходила медленно с течением времени по неясным причинам. [ 21 ]
Потомки
[ редактировать ]В 2021 году анализ мДНК современных и вымерших североамериканских волкообразных собак показал, что берингийский волк был предком южной волчьей клады, в которую входят мексиканский волк и вымерший волк Великих равнин . Мексиканский волк — самый предок серых волков, обитающих сегодня в Северной Америке. Современный койот появился около 10 000 лет назад. Самая генетически базальная клада мДНК койота возникла еще до позднего ледникового максимума и представляет собой гаплотип, который можно найти только у восточного волка . Это означает, что большой волкоподобный плейстоценовый койот был предком восточного волка. Далее, еще один древний гаплотип, обнаруженный у восточного волка, можно обнаружить только у мексиканского волка. Исследование предполагает, что смесь плейстоценового койота и берингийского волка привела к появлению восточного волка задолго до прибытия современных койотов и современных волков. [ 78 ]
Примечания
[ редактировать ]- ^ Полный набор вспомогательных ссылок см. в примечании (a) в филодереве в разделе « Эволюция волка # Волкоподобные псовые».
Ссылки
[ редактировать ]- ^ Фрик, К. (1930). «Замерзшая фауна Аляски». Естественная история (30): 71–80. hdl : 2246/6354 .
- ^ Чейни, Р.; Мейсон, Х. (1936). «Плейстоценовая флора Фэрбенкса, Аляска» . Американский музей Novitates (887): 5. Архивировано из оригинала 03 февраля 2018 г. Проверено 3 февраля 2018 г.
- ^ Новак, Рональд М. (1979). Североамериканский четвертичный пес . 6. Монография Музея естественной истории Канзасского университета. п. 110. дои : 10.5962/bhl.title.4072 . ISBN 978-0-89338-007-6 . Проверено 31 января 2018 г.
- ^ Олсен, Стэнли Дж. (2001). «II.G.8-Одомашнивание:Собаки» . В Кипле, Кеннет Ф.; Орнелас, Кримхильд Коне (ред.). Кембриджская всемирная история еды . Том. 1. Издательство Кембриджского университета. стр. 513–514. ISBN 978-0-521-40214-9 .
- ^ Jump up to: а б Стэнли Дж. Олсен (1985). Происхождение домашней собаки: Летопись окаменелостей – Глава 2 . Издательство Университета Аризоны. п. 22.
- ^ Jump up to: а б с Кох, Пол Л.; Фокс-Доббс, Кена; Ньюсом, Сет В. (2017). «6» . В Дитле, Грегори П.; Флесса, Карл В. (ред.). Природоохранная палеобиология: наука и практика . Издательство Чикагского университета. стр. 110–114. ISBN 978-0-226-50672-2 .
- ^ Смит, Элисон Дж. (2012). «12 свидетельств изменения окружающей среды на основе наземной и пресноводной палеоэкологии» . В Джоне Мэтьюзе (ред.). Справочник SAGE по изменению окружающей среды . Том. 1. МУДРЕЦ. п. 272. ИСБН 978-0-85702-360-5 .
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и С аа аб Леонард, Дженнифер А.; Вила, Карлес; Фокс-Доббс, Кена; Кох, Пол Л.; Уэйн, Роберт К.; Ван Валкенбург, Блэр (2007). «Вымирание мегафауны и исчезновение специализированного экоморфа волка» (PDF) . Современная биология . 17 (13): 1146–50. Бибкод : 2007CBio...17.1146L . дои : 10.1016/j.cub.2007.05.072 . hdl : 10261/61282 . ПМИД 17583509 . S2CID 14039133 . Архивировано из оригинала (PDF) 28 декабря 2016 г. Проверено 28 августа 2015 г.
- ^ Jump up to: а б с д и ж г час я дж к Мичен, Джули А.; Браник, Александрия Л.; Фрай, Трент Дж. (2016). «Вымерший морфотип берингийского волка, обнаруженный в континентальной части США, имеет значение для миграции и эволюции волков» . Экология и эволюция . 6 (10): 3430–8. Бибкод : 2016EcoEv...6.3430M . дои : 10.1002/ece3.2141 . ПМК 4870223 . ПМИД 27252837 .
- ^ Jump up to: а б с д и Фокс-Доббс, Кена; Леонард, Дженнифер А.; Кох, Пол Л. (2008). «Плейстоценовая мегафауна восточной Берингии: палеоэкологические и палеоэкологические интерпретации стабильных изотопов углерода и азота, а также радиоуглеродных записей» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 261 (1–2): 30–46. Бибкод : 2008PPP...261...30F . дои : 10.1016/j.palaeo.2007.12.011 .
- ^ Рис, Джейн Б.; Мейерс, Ноэль; Урри, Лиза А.; Каин, Майкл Л.; Вассерман, Стивен А.; Минорский, Петр Васильевич; Джексон, Роберт Б.; Кук, Бернард Н. (2015). «26-Филогения и древо жизни» . Версия Campbell Biology для Австралии и Новой Зеландии (10-е изд.). Пирсон Австралия. стр. 561–562. ISBN 978-1-4860-0704-2 .
- ^ Jump up to: а б Мичен, Джули; Вуллер, Мэтью Дж.; Барст, Бенджамин Д.; Функ, Джульетта; Крэнн, Карли; Хит, Джесс; Кассат-Джонстон, Молли; Шапиро, Бет; Холл, Элизабет; Хьюитсон, Сьюзен; Зазула, Грант (2020). «Мумифицированный щенок серого волка плейстоцена» . Современная биология . 30 (24): Р1467–Р1468. Бибкод : 2020CBio...30R1467M . дои : 10.1016/j.cub.2020.11.011 . ПМИД 33352124 . S2CID 229346357 .
- ^ Кокс, CB; Мур, Питер Д.; Ладл, Ричард (2016). Биогеография: экологический и эволюционный подход . Уайли-Блэквелл. п. 106. ИСБН 978-1-118-96858-1 .
- ^ Редакционная коллегия (апрель 2012 г.). Краткий научный словарь . Издательство V&S. ISBN 978-93-81588-64-2 .
- ^ Jump up to: а б Арора, Девендер; Сингх, Аджит; Шарма, Викрант; Бхадурия, Харвендра Сингх; Патель, Рам Бахадур (2015). «Hgs Db : База данных гаплогрупп для понимания миграции и оценки молекулярного риска» . Биоинформация . 11 (6): 272–5. дои : 10.6026/97320630011272 . ПМК 4512000 . ПМИД 26229286 .
- ^ Jump up to: а б с д и ж Миклоши, Адам (2015). Поведение, эволюция и познание собак . Оксфордская биология (2-е изд.). Издательство Оксфордского университета. стр. 106–107. ISBN 978-0-19-954566-7 .
- ^ Jump up to: а б с Леонард, Дженнифер А.; Вила, Карлес; Уэйн, Роберт К. (2005). «Утраченное наследие: генетическая изменчивость и размер популяции истребленных серых волков в США ( Canis lupus )». Молекулярная экология . 14 (1): 9–17. дои : 10.1111/j.1365-294X.2004.02389.x . ПМИД 15643947 . S2CID 11343074 .
- ^ Jump up to: а б с д Терви, Сэм (2009). Голоценовые вымирания . Издательство Оксфордского университета. п. 257. ИСБН 978-0-19-953509-5 .
- ^ Jump up to: а б с д и ж г Пилот Малгожата; Браницкий, Войцех; Енджеевский, Влодзимеж; Гощиньский, Яцек; Енджеевска, Богумила; Дикий, Игорь; Шквыря, Марина; Цингарская, Елена (2010). «Филогеографическая история серых волков Европы» . Эволюционная биология BMC . 10 (1): 104. Бибкод : 2010BMCEE..10..104P . дои : 10.1186/1471-2148-10-104 . ПМЦ 2873414 . ПМИД 20409299 .
- ^ Рэнди, Этторе (2011). «Генетика и сохранение волков Canis lupus в Европе». Обзор млекопитающих . 41 (2): 99–111. дои : 10.1111/j.1365-2907.2010.00176.x .
- ^ Jump up to: а б с Эрсмарк, Эрик; Ключ, Корнеля ФК; Чан, Ивонн Л.; Синдинг, Миккель-Хольгер С.; Фейн, Стивен Р.; Илларионова Наталья Александровна; Оскарссон, Маттиас; Улен, Матиас; Чжан, Я-Пин; Дален, Любовь; Саволайнен, Питер (2016). «От прошлого к настоящему: филогеография волков и демографическая история на основе региона митохондриального контроля» . Границы экологии и эволюции . 4 . дои : 10.3389/fevo.2016.00134 . См. страницу 5 с таблицей S3, связь между Clu108 (Россия 18 000) и Clu109 (Аляска 28 000) с Clu8,9,10,22 (Китай).
- ^ Тельманн, О.; Шапиро, Б.; Кюи, П.; Шунеманн, В.Дж.; Сойер, СК; Гринфилд, ДЛ; Гермонпре, МБ; Саблин, М.В.; Лопес-Хиральдес, Ф.; Сандей-Рура, X.; Напиерала, Х.; Урпманн, Х.-П. Лопонте, DM; Акоста, А.А.; Гимш, Л.; Шмитц, РВ; Уортингтон, Б.; Буйкстра, Дж. Э.; Дружкова А.; Графодатский А.С.; Оводов, Н.Д.; Уолберг, Н.; Фридман, АХ; Швейцер, РМ; Кёпфли, К.- П.; Леонард, Дж.А.; Мейер, М.; Краузе, Дж.; Паабо, С.; и др. (2013). «Полные митохондриальные геномы древних псовых предполагают европейское происхождение домашних собак» (PDF ) Наука 342 (6160): 871–4. Бибкод : 2013Sci...342..871T . дои : 10.1126/science.1243650 . hdl : 10261/88173 . ПМИД 24233726 . S2CID 1526260 .
- ^ Jump up to: а б с д и ж Кобльмюллер, Стефан; Вила, Карлес; Лоренте-Галдос, Белен; Дабад, Марк; Рамирес, Оскар; Маркес-Боне, Томас; Уэйн, Роберт К.; Леонард, Дженнифер А. (2016). «Целые митохондриальные геномы освещают древние межконтинентальные расселения серых волков ( Canis lupus )» (PDF) . Журнал биогеографии . 43 (9): 1728. Бибкод : 2016JBiog..43.1728K . дои : 10.1111/jbi.12765 . hdl : 10261/153364 . S2CID 88740690 .
- ^ Жермонпре, Митье; Саблин Михаил Владимирович; Депре, Вивиан; Хофрейтер, Михаэль; Лазничкова-Галетова, Мартина; Стивенс, Рианнон Э.; Стиллер, Матиас (2013). «Палеолитические собаки и раннее одомашнивание волка: ответ на комментарии Крокфорда и Кузьмина (2012)». Журнал археологической науки . 40 (1): 786–792. Бибкод : 2013JArSc..40..786G . дои : 10.1016/j.jas.2012.06.016 .
- ^ Вила, С; Аморим, ИК; Леонард, Дж.А.; Посада, Д; Кастровьехо, Дж; Петруччи-Фонсека, Ф; Крэндалл, Калифорния; Эллегрен, Х; Уэйн, РК (1999). «Филогеография митохондриальной ДНК и история популяции серого волка canis lupus» (PDF) . Молекулярная экология . 8 (12): 2089–103. Бибкод : 1999MolEc...8.2089V . дои : 10.1046/j.1365-294x.1999.00825.x . hdl : 10261/58565 . ПМИД 10632860 . S2CID 31680635 . См. Таблицу 1.
- ^ Барышников Геннадий Ф.; Мол, Дик; Тихонов, Алексей Н (2009). «Находки позднеплейстоценовых хищников на полуострове Таймыр (Россия, Сибирь) с палеоэкологическим контекстом» (PDF) . Российский журнал териологии . 8 (2): 107–113. doi : 10.15298/rusjtheriol.08.2.04 . Проверено 20 мая 2017 г.
- ^ Jump up to: а б с Леонард, Дженнифер (2014). «Экология стимулирует эволюцию серых волков» (PDF) . Эволюционно-экологические исследования . 16 : 461–473. Архивировано из оригинала (PDF) 15 апреля 2016 г. Проверено 21 мая 2016 г.
- ^ Вулперт, Стюарт (14 ноября 2013 г.). «Собаки, вероятно, возникли в Европе более 18 000 лет назад, сообщают биологи Калифорнийского университета в Лос-Анджелесе» . Отдел новостей Калифорнийского университета в Лос-Анджелесе . Проверено 10 декабря 2014 г.
- ^ Jump up to: а б Хофрейтер, Майкл (2007). «Плейстоценовые вымирания: преследование выживших» . Современная биология . 17 (15): 609–11. Бибкод : 2007CBio...17.R609H . дои : 10.1016/j.cub.2007.06.031 . ПМИД 17686436 . S2CID 14268297 .
- ^ Jump up to: а б Мех, Л. Дэвид (1966). Волки острова Рояль . Фауна Серия 7. Фауна национальных парков США. п. 1. ISBN 978-1-4102-0249-9 . Проверено 1 мая 2017 г.
- ^ «Серый волк (на Юконе)» (PDF) . Окружающая среда Юкон . Правительство Канады. 2017 . Проверено 18 апреля 2017 г. .
- ^ Мичен, Дж.А.; Сэмюэлс, JX (2012). «Эволюция койотов (Canis latrans) в ответ на вымирание мегафауны» . Труды Национальной академии наук . 109 (11): 4191–6. Бибкод : 2012PNAS..109.4191M . дои : 10.1073/pnas.1113788109 . ПМК 3306717 . ПМИД 22371581 .
- ^ Тедфорд, Ричард Х.; Ван, Сяомин; Тейлор, Берил Э. (2009). «Филогенетическая систематика североамериканских ископаемых собак (Carnivora: Canidae)» (PDF) . Бюллетень Американского музея естественной истории . 325 : 1–218. дои : 10.1206/574.1 . hdl : 2246/5999 . S2CID 83594819 .
- ^ Мерриам, Дж. К. (1912). «Фауна Ранчо Ла Бреа, Часть II. Canidae» . Мемуары Калифорнийского университета . 1 : 217–273.
- ^ Энионг, Уильям; Роман, Крис (2006). «Новые оценки массы тела Canis dirus, вымершего плейстоценового лютого волка». Журнал палеонтологии позвоночных . 26 : 209–212. doi : 10.1671/0272-4634(2006)26[209:NBMEFC]2.0.CO;2 . S2CID 83702167 .
- ^ Добжанский, Феодосий (1968). «О некоторых фундаментальных понятиях дарвиновской биологии». У Добжанского Феодосия; Хехт, Макс К.; Стир, Уильям К. (ред.). Эволюционная биология . Том. 2. Эпплтон-Сентьюри-Крофтс . стр. 1–34. дои : 10.1007/978-1-4684-8094-8_1 . ISBN 978-1-4684-8096-2 . OCLC 24875357 .
- ^ Jump up to: а б Перри, Анджела (2016). «Волк в собачьей шкуре: первоначальное приручение собаки и плейстоценовая вариация волка». Журнал археологической науки . 68 : 1–4. Бибкод : 2016JArSc..68....1P . дои : 10.1016/j.jas.2016.02.003 .
- ^ Флауэр, Люси, Огайо; Шреве, Даниэль К. (2014). «Исследование палеодиетической изменчивости европейских псовых плейстоцена». Четвертичные научные обзоры . 96 : 188–203. Бибкод : 2014QSRv...96..188F . doi : 10.1016/j.quascirev.2014.04.015 .
- ^ Грант, Питер Р. (2017). «V1.10 — Адаптивное излучение» . В Бауме, Дэвид А.; Футуйма, Дуглас Дж.; Хоекстра, Хопи Э.; Ленски, Ричард Э.; Мур, Аллен Дж.; Пайхель, Кэтрин Л.; Шлютер, Дольф; Уитлок, Майкл С. (ред.). Принстонский путеводитель по эволюции . Издательство Принстонского университета. п. 561. ИСБН 978-0-691-17587-4 .
- ^ Jump up to: а б Межправительственная группа экспертов по изменению климата (ООН) (2007 г.). Четвертый оценочный доклад МГЭИК: Изменение климата, 2007 г. – Палеоклиматическая перспектива (Отчет). Нобелевский фонд.
- ^ Jump up to: а б Кларк, штат Пенсильвания; Дайк, А.С.; Шакун, доктор медицинских наук; Карлсон, А.Е.; Кларк, Дж.; Вольфарт, Б. ; Митровица, JX; Хостетлер, Юго-Запад; Маккейб, AM (2009). «Последний ледниковый максимум». Наука . 325 (5941): 710–4. Бибкод : 2009Sci...325..710C . дои : 10.1126/science.1172873 . ПМИД 19661421 . S2CID 1324559 .
- ^ Якобссон, Мартин; Пирс, Кристоф; Кронин, Томас М.; Бэкман, Ян; Андерсон, Лейф Г.; Барриентос, Наталья; Бьорк, Йоран; Коксалл, Хелен; Де Бур, Агата; Майер, Ларри А.; Мёрт, Карл-Магнус; Нильссон, Йохан; Рэттрей, Джейн Э.; Странн, Кристиан; Семилетов Игорь; О'Риган, Мэтт (2017). «Послеледниковое затопление Берингийского сухопутного моста, датированное 11 000 кал лет назад, на основе новых геофизических и осадочных данных» . Климат прошлых обсуждений : 1. doi : 10.5194/cp-2017-11 .
- ^ Элиас, SA; Шриве, Д. (2016). «Вымирание мегафауны позднего плейстоцена ☆». Справочный модуль по системам Земли и наукам об окружающей среде . дои : 10.1016/B978-0-12-409548-9.10283-0 . ISBN 978-0-12-409548-9 . S2CID 130031864 .
- ^ Jump up to: а б Элиас, С; Крокер, Б. (2008). «Берингов мост: влагобарьер на пути расселения степно-тундровой биоты?» (PDF) . Четвертичные научные обзоры . 27 (27–28): 2473. Бибкод : 2008QSRv...27.2473E . doi : 10.1016/j.quascirev.2008.09.011 .
- ^ Jump up to: а б с д Дейл Гатри, R (2001). «Происхождение и причины возникновения гигантской степи: история облачного покрова, зубных ямок шерстистых млекопитающих, пряжек и Берингии наизнанку». Четвертичные научные обзоры . 20 (1): 549–574. Бибкод : 2001QSRv...20..549D . дои : 10.1016/S0277-3791(00)00099-8 .
- ^ Jump up to: а б Элиас, SA; Бригам-Гретт, Дж. (2007). «ЛЕДНИЧЕСТВА | События позднего плейстоцена в Берингии» (PDF) . Энциклопедия четвертичной науки . стр. 1057–1066. дои : 10.1016/B0-44-452747-8/00132-0 . ISBN 978-0-444-52747-9 . Проверено 2 мая 2017 г.
- ^ Хоффекер, Дж. Ф.; Элиас, С.А. (2007). Экология человека Берингии . Издательство Колумбийского университета. п. 57. ИСБН 978-0-231-13060-8 .
- ^ Бригам-Гретт, Дж .; Ложкин А.В.; Андерсон, премьер-министр; Глушкова О.Ю. (2004). «Палеоэкологические условия в Западной Берингии до и во время последнего ледникового максимума». В Мэдсене, Д.Б. (ред.). Проникновение в Америку, Северо-Восточную Азию и Берингию перед последним ледниковым максимумом . Университет Юты Пресс. стр. 29–61. S2CID 7209566 .
- ^ Шер, А.В.; Кузьмина, С.А.; Кузнецова, ТВ; Сулержицкий, Л.Д. (2005). «Новое представление о вейкселианской среде и климате Восточно-Сибирской Арктики, полученное на основе ископаемых насекомых, растений и млекопитающих» (PDF) . Четвертичные научные обзоры . 24 (5–6): 533. Бибкод : 2005QSRv...24..533S . doi : 10.1016/j.quascirev.2004.09.007 .
- ^ Андерсон, Патрисия М.; против Ложкина, Анатолий (2001). «Интерстадиальный комплекс 3-го этапа (Каргинский / средний Висконсинанский интервал) Берингии: вариации палеообстановки и значение для палеоклиматических интерпретаций». Четвертичные научные обзоры . 20 (1): 93–125. Бибкод : 2001QSRv...20...93A . дои : 10.1016/S0277-3791(00)00129-3 .
- ^ Жермонпре, Митье; Саблин Михаил Владимирович; Стивенс, Рианнон Э.; Хеджес, Роберт Э.М.; Хофрейтер, Михаэль; Стиллер, Матиас; Депре, Вивиан Р. (2009). «Ископаемые собаки и волки из палеолитических памятников Бельгии, Украины и России: остеометрия, древняя ДНК и стабильные изотопы». Журнал археологической науки . 36 (2): 473. Бибкод : 2009JArSc..36..473G . дои : 10.1016/j.jas.2008.09.033 .
- ^ Хофрейтер, Михаэль; Барнс, Ян (2010). «Утраченное разнообразие: все ли виды крупных млекопитающих Голарктики — всего лишь реликтовые популяции?» . БМК Биология . 8:46 . дои : 10.1186/1741-7007-8-46 . ПМК 2858106 . ПМИД 20409351 .
- ^ Стюарт, младший (2009). «Эволюционное последствие индивидуалистической реакции на изменение климата» . Журнал эволюционной биологии . 22 (12): 2363–75. дои : 10.1111/j.1420-9101.2009.01859.x . ПМИД 19888939 . S2CID 20955499 .
- ^ Амин, Карли; Халм-Биман, Ардерн; Эвин, Аллоуэн; Жермонпре, Митье; Бриттон, Кейт; Кучки, Томас; Ларсон, Грегер; Добни, Кейт (2017). «Основной подход к оценке надежности скученности зубов нижней челюсти как маркера приручения собаки» . Журнал археологической науки . 85 : 41–50. Бибкод : 2017JArSc..85...41A . дои : 10.1016/j.jas.2017.06.014 .
- ^ Jump up to: а б Ван Валкенбург, Блэр; Гертель, Фриц (1993). «Тяжелые времена в Ла-Бреа: поломка зубов у крупных хищников позднего плейстоцена» (PDF) . Наука . Новая серия. 261 (5120): 456–459. Бибкод : 1993Sci...261..456V . дои : 10.1126/science.261.5120.456 . ПМИД 17770024 . S2CID 39657617 .
- ^ Терриен, Франсуа (2005). «Профили силы нижней челюсти современных хищников и последствия для пищевого поведения вымерших хищников». Журнал зоологии . 267 (3): 249–270. дои : 10.1017/S0952836905007430 .
- ^ Ван Валкенбург, Б (1988). «Частота поломки зубов у крупных хищных млекопитающих». Являюсь. Нат . 131 (2): 291–302. дои : 10.1086/284790 . S2CID 222330098 .
- ^ Jump up to: а б с ДеСантис, LRG; Шуберт, Б.В.; Шмитт-Линвилл, Э.; Унгар, П.; Донохью, С.; Хаупт, Р.Дж. (15 сентября 2015 г.). Джон М. Харрис (ред.). «Текстуры стоматологической микроодежды хищных животных из смоляных ям Ла-Бреа, Калифорния, и возможные последствия их вымирания» . Вклад в науку . Научная серия 42 (специальный том под названием «Ла Бреа и за его пределами: Палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия Музея естественной истории раскопок округа Лос-Анджелес на Ранчо ла Бреа). Музей естественной истории округа Лос-Анджелес: 37–52 . Проверено 21 июля 2019 г.
- ^ Крокфорд, Сьюзен Дж.; Кузьмин, Ярослав В. (2012). «Комментарии к Жермонпре и др., Журнал археологической науки 36, 2009 г. «Ископаемые собаки и волки из палеолитических памятников в Бельгии, Украине и России: остеометрия, древняя ДНК и стабильные изотопы», а также Жермонпре, Лазкичкова-Галетова и Саблин, Журнал археологической науки 39, 2012 «Палеолитические черепа собак на стоянке Граветтский Пржедмости, Чехия» «. Журнал археологической науки . 39 (8): 2797. Бибкод : 2012JArSc..39.2797C . дои : 10.1016/j.jas.2012.04.033 .
- ^ Буржон, Лориан; Берк, Ариана; Хайэм, Томас (2017). «Самое раннее присутствие человека в Северной Америке датировано периодом последнего ледникового максимума: новые радиоуглеродные даты из пещер Блюфиш, Канада» . ПЛОС ОДИН . 12 (1): e0169486. Бибкод : 2017PLoSO..1269486B . дои : 10.1371/journal.pone.0169486 . ПМЦ 5218561 . ПМИД 28060931 .
- ^ Jump up to: а б О'Киф, Ф.Робин; Биндер, Венди Дж.; Фрост, Стивен Р.; Сэдлиер, Редьярд В.; Ван Валкенбург, Блэр (2014). «Краниальная морфометрия лютого волка Canis dirus на ранчо Ла Бреа: временная изменчивость и ее связь с питательным стрессом и климатом» . Электронная палеонтология . 17 (1): 1–24.
- ^ Ван Валкенбург, Блэр (2008). «Цены хищничества: перелом зубов у плейстоценовых и современных хищников» . Биологический журнал Линнеевского общества . 96 : 68–81. дои : 10.1111/j.1095-8312.2008.01108.x .
- ^ Ван Валкенбург, Блэр; Петерсон, Рольф О.; Смит, Дуглас В.; Сталер, Дэниел Р.; Вучетич, Джон А. (2019). «Частота переломов зубов у серых волков отражает доступность добычи» . электронная жизнь . 8 . doi : 10.7554/eLife.48628 . ПМК 6759352 . ПМИД 31549963 .
- ^ Томия, Сусуму; Мичен, Джули А. (2018). «Посткраниальное разнообразие и недавнее экоморфное обеднение североамериканских серых волков» . Письма по биологии . 14 (1): 20170613. doi : 10.1098/rsbl.2017.0613 . ПМК 5803591 . ПМИД 29343558 .
- ^ Ласель, Денис; Лориоль, Бернар; Зазула, Грант; Галеб, Басам; Уттинг, Николас; Кларк, Ян Д. (2013). «Время наступления и базальное состояние Лаврентидского ледникового щита во время последнего ледникового максимума в горах Ричардсон, СЗТ». Четвертичные исследования . 80 (2): 274. Бибкод : 2013QuRes..80..274L . дои : 10.1016/j.yqres.2013.06.001 . S2CID 140156890 .
- ^ Шапиро, Б.; Драммонд, Эй Джей; Рамбо, А; Уилсон, MC; Матеус, ЧП; Шер, А.В.; Пибус, Огайо; Гилберт, Монтана; Барнс, я; Бинладен, Дж; Виллерслев, Э; Хансен, Эй Джей; Барышников, Г.Ф.; Бернс, Дж.А.; Давыдов С ; Драйвер, Джей Си; Фрёзе, Д.Г.; Харрингтон, Чехия; Кедди, Дж; Косинцев П.; Кунц, ML; Мартин, LD; Стивенсон, Р.О.; Сторер, Дж.; Тедфорд, Р.; Зимов С ; Купер, А. (2004). «Взлет и падение берингийского степного зубра» (PDF ) Наука 306 (5701): 1561–5. Бибкод : 2004Sci... 306.1561S дои : 10.1126/science.1101074 . PMID 15567864 . S2CID 27134675 .
- ^ Стэнли, Стивен М. (1987). Вымирание . Научная американская библиотека. п. 242.
- ^ Пайхель, Кэтрин Л. (2017). «V.12 – Генетика фенотипической эволюции» . В Бауме, Дэвид А.; Будущее, Дуглас Дж.; Хоекстра, Хопи Э.; Ленски, Ричард Э.; Мур, Аллен Дж.; Пайхель, Кэтрин Л.; Шлютер, Дольф; Уитлок, Майкл С. (ред.). Принстонский путеводитель по эволюции . Издательство Принстонского университета. п. 454. ИСБН 978-0-691-17587-4 .
- ^ Штраус, Шэрон Ю. (2017). «III.15 – Адаптация к биотической среде» . В Бауме, Дэвид А.; Футуйма, Дуглас Дж.; Хоекстра, Хопи Э.; Ленски, Ричард Э.; Мур, Аллен Дж.; Пайхель, Кэтрин Л.; Шлютер, Дольф; Уитлок, Майкл С. (ред.). Принстонский путеводитель по эволюции . Издательство Принстонского университета. п. 300. ИСБН 978-0-691-17587-4 .
- ^ О'Киф, Франция; Фет, Е.В.; Харрис, Дж. М. (2009). «Компиляция, калибровка и синтез радиоуглеродных дат фауны и флоры, Ранчо Ла Бреа, Калифорния» . Вклад в науку . 518 : 1–16. дои : 10.5962/стр.226783 . S2CID 128107590 . Проверено 21 июля 2019 г.
- ^ Грэм, RW; Мид, Дж.И. (1987). «Колебания окружающей среды и эволюция фауны млекопитающих во время последней дегляциации в Северной Америке». В Раддимане, штат Вирджиния; Райт, HE (ред.). Северная Америка и прилегающие океаны во время последней дегляциации . Геологическое общество Америки. стр. 371–402. ISBN 978-0-8137-5203-7 .
- ^ Барноский, А.Д. (1989). «Вымирание в позднем плейстоцене как парадигма повсеместного вымирания млекопитающих». В Доноване, Стивен К. (ред.). Массовые вымирания: процессы и доказательства . Издательство Колумбийского университета. стр. 235–255. ISBN 978-0-231-07091-1 .
- ^ Браник, Александрия Л.; Мичен, Джули А.; О'Киф, Ф. Робин (15 сентября 2015 г.). Джон М. Харрис (ред.). «Микроэволюция формы челюсти у лютого волка Canis dirus на ранчо Ла Бреа». Вклад в науку . Научная серия 42 (специальный том под названием «Ла Бреа и за его пределами: Палеонтология сохранившихся в асфальте биот» в ознаменование 100-летия Музея естественной истории раскопок округа Лос-Анджелес на Ранчо ла Бреа). Музей естественной истории округа Лос-Анджелес: 23–32.
- ^ Купер, А. (2015). «Резкое потепление привело к смене мегафауны позднего плейстоцена Голарктики». Наука . 349 (6248): 602–6. Бибкод : 2015Sci...349..602C . дои : 10.1126/science.aac4315 . ПМИД 26250679 . S2CID 31686497 .
- ^ Jump up to: а б Хейл, Дж.; Фрёзе, Д.Г.; Макфи, RDE; Робертс, Р.Г.; Арнольд, LJ; Рейес, А.В.; Расмуссен, М.; Нильсен, Р.; Брук, BW; Робинсон, С.; Демуро, М.; Гилберт, MTP; Мунк, К.; Остин, Джей-Джей; Купер, А.; Барнс, И.; Моллер, П.; Виллерслев, Э. (2009). «Древняя ДНК показывает позднее выживание мамонта и лошади во внутренних районах Аляски» . Труды Национальной академии наук . 106 (52): 22352–7. Бибкод : 2009PNAS..10622352H . дои : 10.1073/pnas.0912510106 . ПМЦ 2795395 . ПМИД 20018740 .
- ^ Зазула, Грант Д.; Холл, Элизабет; Заяц, П. Грегори; Томас, Кристиан; Мэтьюз, Рольф; Ла Фарж, Кэтрин; Мартель, Андре Л.; Хайнцман, Питер Д.; Шапиро, Бет (2017). «Степной зубр среднего голоцена и палеосреды из лугов Верслюс, Уайтхорс, Юкон, Канада». Канадский журнал наук о Земле . 54 (11): 1138–1152. Бибкод : 2017CaJES..54.1138Z . doi : 10.1139/cjes-2017-0100 . hdl : 1807/78639 . S2CID 54951935 .
- ^ Павелкова Ржичанкова, Вера; Робовский, Ян; Ригерт, Ян (2014). «Экологическая структура современной и последней ледниковой фауны млекопитающих Северной Евразии: на примере Алтае-Саянского рефугиума» . ПЛОС ОДИН . 9 (1): e85056. Бибкод : 2014PLoSO...985056P . дои : 10.1371/journal.pone.0085056 . ПМЦ 3890305 . ПМИД 24454791 .
- ^ Уилсон, Пол Дж.; Ратледж, Линда Ю. (2021). «Учитывая плейстоценовые североамериканские волки и койоты в истории происхождения восточных собак » . Экология и эволюция . 11 (13): 9137–9147. Бибкод : 2021EcoEv..11.9137W . дои : 10.1002/ece3.7757 . ПМЦ 8258226 . ПМИД 34257949 .
Внешние ссылки
[ редактировать ]- Нижняя челюсть берингийского волка , датированная 31 700 лет назад, демонстрирует большую острую нижнюю хищную часть - Музей Севера, Университет Аляски (база данных Arctos)
- Нижняя челюсть берингийского волка, датированная 31 700 лет назад – вид экземпляра выше с другой стороны.
- Записки Берингийских исследований – Древние северные волки Правительство Юкона
- Млекопитающие ледникового периода Юкона Правительство Юкона