Therizinosauridae
| Теризинозавриды Временный диапазон: меловой ,
~ | |
|---|---|

| |
| Конные скелеты Nothronychus McKinleyi и N. Graffami | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Клада : | Динозаурия |
| Клада : | Сауриския |
| Клада : | Теропода |
| Клада : | Манираптора |
| Клада : | † Therizinosauria |
| Суперсемейство: | † Therizinosauroidea |
| Семья: | † Therizinosauridae Малев , 1954 |
| Тип видов | |
| † Therizinosaurus cheloniformis Малев, 1954
| |
| Роды | |
| Синонимы | |
Therizinosauridae (что означает «ящерицы») [ 1 ] является вымершим семейством полученных (продвинутых) теризинозавраиидов динозавров , чьи окаменелости были обнаружены в основном в глуших границах. Несмотря на то, что репрезентативные окаменелости были обнаружены только по всей Азии и Северной Америке , считается, что диапазон Therizinosauridae охватывал большую часть суперконтинента Лорасии на основе нескольких следов и изолированных останков в Европе и Африке. В настоящее время Therizinosauridae включает восемь описанных и названных таксонов.
Therizinosauridae была названа в 1954 году палеонтологом Евгением Малеем после крупных когти, несущих когти, когтя типовых видов Therzinosaurus cheloniformis . Теризинозавриды, как правило, были большими и очень надежно построенными животными , у которых был почти сходящийся план тела с более поздними (и также вымершими) наземными леницами . Самыми крупными родами группы являются Thrizinosaurus и Segonosaurus , которые составляли около 10 м (33 фута) и длиной 7 м (23 фута) соответственно. Физиология теризинозавридов включают широкий и округлый живот, поддерживаемый широким и надежным тазом с толстыми задние льготы, состоящие из очень крепких, четырехмагнических ног, сильной сборки руки с повышенной гибкостью рук, удлиненными когтями рук и высокоэтапным Почти птичьего внутреннего уха. Черты, которые также хорошо известны, включают удлиненную шею, выдающийся кератиновый клюв и просауропод, похожий на зубной ряд, который отличается от всех теропод. Кроме того, по оценкам, теризинозавриды имели более продвинутые перья по сравнению с примитивными теризинозаврами ( Beipiaosaurus или Jianchangosaurus ).
Уникальные и странные особенности группы поощряли исследования палеобиологии и палеоэкологии семьи . Справедливая часть современных исследований сосредоточена на кормлениях этих динозавров, поскольку они считаются лучшими кандидатами на теропод для травоядных . В то время как другие тероподные группы являются полностью плотоядными, члены Therizinosauridae расходились и приняли травоядный и, возможно, всеядный образ жизни. Это еще больше поддерживается их необычной морфологией. Как указано в их морфологии ног и несколько следов из Азии, Африки и Европы, они, вероятно, были ходунками Plantigrade , но может потребоваться дальнейшее обследование. Теризинозавриды были яйцевальщими животными, которые вкладывали в колонии и укладывали яичные сцепления . Фактически, яйца теризинозавра особенно распространены в поздних мелах , в основном в Азии. Oofamily . Dendroolithidae часто приписывают динозаврам теризинозавра Некоторые из первых яиц дендрулита были найдены на Баян Шире и Нанчао Формации .
Их отношения сбивались с толку и неясны в первые годы исследований в основном из -за необычных черт среди членов. Было предложено несколько альтернативных классификаций (например, именование Segonosauridae в 1979 году) до тех пор, пока в 1990 -х годах не были описаны более полные образцы и другие таксоны, что подтвердило их как тероподы . Многие из общих персонажей в группе также показали, что Segnosauridae были младшим синонимом гораздо более раннего семейства Therizinosauridae. Текущий филогенетический консенсус заключается в том, что теризинозавриды эволюционировали из мелких, похожих на птиц манирапторов , и, таким образом, они попадают в каелурозаврианскую кладу, называемую Манираптором . Более того, большинство черт теризинозавридов (таких как структура уха ) были унаследованы меньшими, мудрыми, плотоядными предками. Обширный филогенетический анализ пришел к выводу, что в Манирапторе теризинозавры были первыми из пяти основных групп, которые расходились.
История открытия
[ редактировать ]
Первым окончательным обнаруженным теризинозавридом был Thrizinosaurus . Окаменечные останки были обнаружены в 1918 году во время монгольской полевой экспедиции по образованию Немегта пустыни Гоби . Несколько когтей были обнаружены командой, а затем описали советский палеонтолог Евгений Малев в 1954 году. Когти были необычайно большими, приблизительно 1 м (3,3 фута) длиной, если он восстановлен, очень прямые и уплощенные. Малев считал, что когти принадлежали гигантской морской черепахе и назвали новый род и виды Therizinosaurus cheloniformis , также возводя Therizinosauridae , включающие новые виды. [ 1 ]
В 1970 -х годах было сделано еще больше результатов, включая название большего количества таксонов. Дополнительные образцы Thrizinosaurus были направлены монгольским палеонтологом Ринчен Барсболд в 1976 году. Он описал набор частичных рук из формации Немегта и, учитывая сходства в когтях, он направил этот образец к роду. [ 2 ] Монгольский палеонтолог Альтангел Перл описал и назвал новый род Segonosaurus в 1979 году, основанный на нижних челюстях и большей части задних конечностей. Он также придумал более новые Segonosauridae (ныне синоним Therizinosauridae), чтобы сдержать этот вид. [ 3 ] В том же году палеонтолог Донг Жиминг описал род nanshiungosaurus , но неправильно интерпретировал остатки, связанные с каким -то карликовым сауроподом. [ 4 ] В следующем году Барсболд и Перл придумали семейство Segonosauria (ныне Therizinosauria), чтобы сдержать Segonosauridae и Kin. Кроме того, они назвали и кратко описали Erlikosaurus , нового рода, меньше, чем Segonosaurus . [ 5 ] Смущающе, Перл переписывал Эрликозавр, рассматривая таксона как нового рода и видов в 1981 году. [ 6 ] Кроме того, Перл описал еще один образец Therizinosaurus в 1982 году, на этот раз частичную заднюю конечность. Он направил задницу к роду, основанному на сходстве с Segonosaurus . [ 7 ]
новые теризинозавриды Neimongosaurus и Nothronychus ( N. McKinleyi ), В 2001 году были описаны и названные [ 8 ] [ 9 ] и в следующем году Эрлисавр также был описан и назван. [ 10 ] Хотя новый вид nothronychus ( Н. Граффами ) был назван в 2009 году. [ 11 ] При этом оба вида делают Nothronychus наиболее полным известным теризинозавридом. [ 12 ]
Описание
[ редактировать ]
Теризинозавриды были довольно продвинутыми и надежно построенными животными, которые достигли многотонных измерений, которые делают их среди самых больших манирапторов . Самыми маленькими известными теризинозавридами являются Erlianaurus , Erlikosaurus и Neimongosaurus , которые длились около 3–4 м (9,8–13,1 фута), кроме того, голотип черепа Erlikosaurus составляет около 25 см (250 мм). Segonosaurus и возможный теризинозаврид Suzhousaurus были примерно такого же размера, длиной 6–7 м (20–23 фута) и весом 1,3 т (2900 фунтов). Немного меньших членов представлены Nanshiungosaurus и Nothronychus , оцениваются в длину 5–5,3 м (16–17 футов) и 600–1000 кг (0,60–1,00 т) в веса соответственно. [ 13 ] [ 14 ] Связанный «nanshiungosaurus» Bohlini был одинаковым по размеру (около 6 м (20 футов) длиной), однако его назначение на Nanshiungosaurus маловероятно, и это может или не представлять другой вид или образец более известного Suzhousaurus . [ 14 ] [ 15 ] Thrizinosaurus были крупнейшими представителями, достигнув верхних измерений группы, они выросли до 10 м (33 фута) длиной более 5 т (11 000 фунтов). Эти аспекты, которые делают род среди крупнейших известных теропод . [ 14 ] Экспоненциальные размеры, полученные в Therizinosauridae, по -видимому, были вызваны объемной и специализированной диетой в группе и ранними членами Therizinosauria. [ 16 ]
Впечатления кожи от Beipiaosaurus и Jianchangosaurus указывают на то, что примизитивные теризинозавры были покрыты слоем , похожими на ненормативные , редкие перья, похожие на те, которые наблюдались в компоненте Sinosauropteryx , а также более длинные, более простые, похожие на Quill-перья, которые могли быть использованы на демонстрации. [ 17 ] [ 18 ] [ 19 ] [ 20 ] Однако, хотя и самый ранний известный пример примитивных перьев стадии I встречается среди ранних теризинозавров, считается, что более продвинутые члены обладают более развитыми, птичьими перьями. Несмотря на то, что обычно связаны с полетом, перья, которые покрывали этих животных, не использовались для этой цели, вместо этого они помогали ни в метаболизме , ни в дисплее . [ 21 ] [ 22 ]
Череп
[ редактировать ]


У теризинозавридов было больше специализированных черепов по сравнению с примитивными членами, такими как Falcarius , у которых была универсальная морда без клюва. [ 23 ] У них были относительно вытянутые морды, и наконечник был покрыт рампотекой ( кератиновой клюв), используемого во время кормления. Хотя расширение клюва на дорсальной поверхности морды неизвестно, подсчитано, что он слегка перекрыл полость носа, как у некоторых современных птиц. Устье беззубых (нижняя челюсть) также разработала рампотеку, которую можно увидеть на ( без зубов) переднего конца. Это расширение было поставлено с помощью нервно -сосудистого отверстия (небольших ям), обнаруженных на боковых поверхностях. [ 24 ] теризинозавридов Известные образцы эрликозавра , Neimongosaurus и Segnosaurus сохраняют многочисленные нейроверосудистые отверстия (более печально известный в Erlikosaurus ), что указывает на то, что в жизни присутствовал хорошо развитый клюв. [ 8 ] [ 24 ] [ 25 ] Как верхняя челюсть, так и предчезгилла были зубчатыми, и некоторые виды теризинозавридов имели специализированные, ретудированные зубные зубные ситуации, такие как Segonosaurus и, возможно, Neimongosaurus . [ 25 ] Мозговые мозги известны из трех теризинозавридов: Erlikosaurus , Neimongosaurus и N. McKinleyi . К сожалению, мозговой мозг Neimongosaurus не был описан и не иллюстрирован. [ 12 ] Мозговые кольца направляются на дно, космируются с хорошо развитыми боковыми ориентированными парами, которые сильно пневматизированы и имели выдающееся центральное отверстие . [ 24 ] [ 12 ]
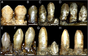



Расточие зубные рты - еще одна фигурная черта в теризинозавридах, которая отличается от всех тероподных групп. По меньшей мере две разные морфологии зубов наблюдаются среди теризинозавридов; Первый представлен относительно гомодонтом , овальными или ланцетными зубами в форме с умеренными грубыми зубцами (зубцами) на коронах (верхняя часть открытой части). Этот тип зубного зуба лучше представлен полным трехмерным голотипом черепа Erlikosaurus , который включает упомянутые символы. [ 24 ] Два изолированных зуба известны из Nothronychus , и они имеют ланцетную форму, симметричны, имеют умеренные зубные зубные зубные камни и сильно напоминают зубцы Erlikosaurus . Кроме того, они, кажется, вытекают из зубного мозга на основе сравнения с последним. [ 12 ] Сохраненный голотип зубного кончика Neimongosaurus сохраняет изверенный зуб , который имеет ланцетную форму с небольшими грубыми зубцами, попадая в этот тип зубных зубов. [ 8 ] Другой тип зубной морфологии - тот, который наблюдается на высокоспециализированном Segonosaurus . В этом таксоне зубы очень гетеродонт , в форме листьев с относительно меньшим количеством зубчатых зубцов, которые заметно развиваются, которые были больше, чем у предыдущих теризинозавридов. Эти зубчатые зубцы состоят из многочисленных сложенных каринов (режущих краев) с зубчатыми передними краями, создавая шероховатую и разрушенную поверхность вблизи основания зубных коронок. [ 25 ] Большинство теризинозавридов, по -видимому, имели низкую скорость замены зубов или не могли их потерять. [ 11 ] [ 26 ]
Посткраниальный скелет
[ редактировать ]
У теризинозавридов плечевой пояс был надежно специализирован, чем у других манирапторов и менее похожими на птицы. Лопатка (лопатка) была очень прямой и сплюснутой на верхнем конце , сливаемой с коракоидной костью, образуя лопатокоракоид. Рядом с области лопатокоракоидного шва на коракоиде присутствует большая отверстия. Коракоид представляет собой широкий и широкий элемент, который немного выпуклый и утолщен вблизи лопатокоракоидного шва. Компания плечевая кость (верхняя рука) была исключительно надежной и гибкой с широкими нижними концами, как видно на гумне теризинозавридов Nothronychus и Therizinosaurus . Не только это, но и мышца бицепса была заметно хорошо развита в Thrizinosaurus . [ 2 ] [ 12 ] В Segonosaurus был сильно построен дельтопекторный гребень ( прикрепление дельтовидной мышцы ). [ 3 ] Antebrachium был относительно прямой у большинства членов, особенно на Therizinosaurus . Снижение запястного блока на теризинозавридах позволило усилить сгибание рук. Ручные фаланги имеют относительно стандартную форму среди манирапторов с формулой цифр I, II и III. [ 10 ] [ 12 ]

Ручные кости ( кости когтя ) пропорционально больше, чем фаланги, сильно сплющенные из стороны в сторону, и повторяются с большей степенью специализации, чем теризинозавра . Большинство теризинозавридов были резко заостренными и повторяющимися изоляциями с очень надежными бугорками ( прикрепление сухожилий сгибателей ). Эти черты лучше видны на Nothronychus и Segonosaurus . [ 3 ] [ 10 ] [ 12 ] Однако в Therizinosaurus ручные костюмы были чрезвычайно удлинены и прямыми с плохими кривыми. Хотя большинство из них являются неполными, в случае восстановления они будут измерять длину около 1 м (3,3 фута), что делает их самыми большими когтями для любого известного наземного животного. Туберклы не так сильно развиты, как у других теризинозавридов, кроме того, у Thrizinosaurus были одни из самых длинных передних конечностей, известных для любых двуногных динозавров: сохранившаяся правая рука в образец IGM 100/15 имеет общую длину 2,4 м (7,9 фута). Полем [ 2 ] [ 27 ]
Позвоночная колонна была высоко пневматизирована (расположенная на воздухе) и относительно хорошо документирована от нескольких плотно сохранившихся элементов среди родов, но образцы Nanshiungosaurus и Nothronychus сохраняют наиболее полную серию позвоночных. [ 4 ] [ 12 ] Теризинозавриды имели большие и надежные шейные позвонки с относительно короткими нервными шипами и уплатыми (вогнутыми на обоих концах) до опишкокуле (вогнутых задних концов), которые были удлинены и имели некоторую степень пневмажности. В спинных позвонках нервные шипы более удлиненные и жесткие. Несколько пневматопоров (небольшие отверстия, ведущие к воздушным карманам) присутствуют на дорсолатеральных поверхностях центра, и они постепенно уменьшаются в размерах. У некоторых видов, таких как Н. Граффами, на спине была узкая конструкция, похожая на горба , о чем свидетельствует очень удлиненные нейронные колючки первых дорсалов. У теризинозавридов крестчик был составлен примерно шестью сакральными позвонками с широким центром, которые имеют округлые аспекты. Каудальные позвонки были коренастыми с жесткими и округленными поперечными проекциями. По сравнению с ранними членами, теризинозавриды имели уменьшение количества хвостах и незначительное сужение шеврона , которое указывает на более короткий и гибкий хвост . [ 3 ] [ 4 ] [ 12 ]

Теризинозавриды имели широкие туловища поддерживаемые широким, похожего на лениво -таз , . Грубная клетка была очень круглой и составленной удлиненными ребрами с довольно надежной капитулой . Наиболее модифицированным элементом в рамках Thrizinosaurid был владение уникальным опиопубическим тазом (лоб и иший, простирающийся назад), что известно, иначе только у птиц и орнитишхов . был Подвздошной подвздошной кости больше, чем уший и лоб , имел очень отклоненные и выраженные подвздошные лезвия. Выдающее расширение подвздошного костюма показывает, что у теризинозавридов были огромные бедра . В нижней части ичийя присутствовал большой процесс обтураторного процесса (похожая на гребень), особенно в Segonosaurus и Nothronychus . Лобик был больше, чем у Ischium, и имел большой лобкий ботинок (широкое расширение в конце). В отличие от любой другой группы Theropod, лоб и иший были прикреплены вместе с твердой структурой с квадратным концом. [ 3 ] [ 4 ] [ 12 ] И бедренная кость , и голень были надежны в конституции. Первый был относительно прямой с хорошо развитыми артикуляциями и бедренной головой. Цвета была немного короче, чем бедра и имела широкий нижний конец, образуя выдающуюся лодыжку . Малоберцовая кость была ущипнута на дне и очень жестко. Как астрагал , так и кальциум были в целом надежными и удлиненными костями, но астрагаларное тело было, по -видимому, уменьшено в этой группе. У Astragalus есть удлиненный боковой мыщел , но медиальный Боковая проекция Astragalus утолщается и распространяется на среднюю линию всего элемента от внутренней стороны до задней части, ведущее пространство для нижней части голени; Это обнажает боковую поверхность голени в фронтальном виде. Кальциан был больше и устойчивым, чем большинство теропод. Он имеет округлую форму с вогнутыми суставными поверхностями и имел подвижную артикуляцию с соседними элементами, такими как астрагал или дистальные ласки. Другим высоко измененным элементом был Тетрадактильные PES : Плеутарь был составлен четырьмя укороченными, полностью функционально плюсневыми . Сновистики III и IV были почти одинаковыми по размеру, второй был слегка узким, а первый был самым коротким. Однако существуют следы плюсневого V, но он сильно уменьшен и не имеет функциональной значимости - как наблюдается на Segonosaurus . Формула фаланга была как и в других манирапторах, IV-4, III-3, II-2 и I-1 (за исключением Unguals). Педали педали были резко заостренными, сплюснуты от стороны в сторону и меньше ручных. [ 7 ] [ 10 ] [ 12 ] Однако в Erlikosaurus они массивны, сочетают в себе жесткую и повторяющуюся форму с надежными бугорками. [ 6 ]
Классификация
[ редактировать ]
Семейство Therizinosauridae была придумана Евгением Малеем в 1954 году, чтобы сдержать загадочный Thrizinosaurus , который интерпретировал этот таксон как представляющий гигантских морских черепах. Родственники Thrizinosaurus были позже найдены, но не признаны как таковые в течение некоторого времени. [ 1 ] С описанием Segonosaurus в 1979 году палеонтолог Altangerel Perle придумал семейство Segonosauridae, чтобы сдержать этот загадочный таксон и предварительно считал эту группу представлять тероподы. Он отметил, что эта новая семья отличалась от Therizinosauridae, основанных на признаках когтя и антебрахий. [ 3 ] Более того, в том же году Перл и Ринчен Барсболд проанализировали структуру таза Segonosaurus и пришли к выводу, что он необычайно отличается от структуры «традиционных» теропод. Основываясь на этих наблюдениях, они предположили, что сегенозавриды должны быть разделены на отдельную группировку, около уровня основного подразделения в подразделении Saurischia . [ 28 ] Параллельно с этим, вновь, а также описанный Nanshiungosaurus был назначен Titanosaurinae Dong Zhiming , основанным на предположении, что это был род Sauropod. [ 4 ] В следующем году и Барсболд, и Перл назвали новую секундозаурию Theropod Segonosauria, содержащую Segonosauridae. В этой новой статье они также описали новые Erlikosaurus и Enigmosaurus (тогда неназванный), отмечая, что сегензавры были более похожи на динозавров теропод, и хотя некоторые из их особенностей напоминали особенности орнитишхов и сауроподов , эти сходства были поверхностными и различными при подробном изучении. [ 5 ]
В 1982 году Перл описал третий образец теризинозавридов Therizinosaurus . Он направил этот материал на род, основанный на поразительных сходствах с образцами Segonosaurus . Кроме того, Перл сравнил передние конечности в этих двух таксонах и пришел к выводу, что они характеризовались удлиненным оружием, возможно, принадлежащим к одной таксономической группе. [ 7 ] Однако, основываясь на остеологических особенностях, в 1984 году Грегори С. Пол предположил, что сегензавры не были тероподами, а поздними мелами -просауроподами , и они представляли переходную форму между просороподами и орнитишами. Например, Segnosaurs напоминали просауроподы в их морфологии черепа и ног, а также были похожи на орнитишки в их морде, лобке и лодыжке. [ 29 ] В 1988 году он предложил сегенозаврную классификацию для теризинозавридов Therizinosaurus . [ 30 ] В обзорной статье о книге «Динозаврия» в 1990 году Барсболд и Тереза Марьянка считают Сегнозаурию загадочной группой сауришек, в которой должно быть изменение позиция. Они, однако, не согласились с Therizinosaurus как таксоном сегнозавров, так как он был известен из материала передней части передней части; Они подтвердили упомянутый материал задних конечностей как сегнозавриан. Наконец, Барсболд и Марьянска отметили поразительное сходство между тазами Наншингозавры и Сегнозавром , такими как опиопубическое состояние и большие подвздошные лезвия. Они пришли к выводу, что первое было частью Segonosauridae. [ 31 ]

С описанием Thrizinosauroids Alxasaurus в 1993 году Дейлом А. Расселом и Донг Чжимингом, сродство группы было довольно ясным. Этот новый таксон представлял собой наиболее полный член и был известен из нескольких образцов с многочисленными чертами теропода. Более того, консервированные задних конечностей в некоторых образцах показали, что назначение задних конечностей сегенозавры теризинозавру было правильным, а «сегенозавры» были фактически тероподами. Рассел и Донг также отметили чрезвычайное сходство между Therizinosauridae и Segonosauridae и считали, что последний был синонимом первого из -за приоритета . Тем не менее, Alxasaurus был довольно более примитивным родом, а суперсемейство Therzinosauroidea была придуманной для его содержания и родственных видов. [ 32 ] Позади этого года Кларк и его коллеги переписали череп голотипа Erlikosaurus и обнаружили больше тероподных признаков, чем когда они впервые описаны. Они пришли к выводу, что теризинозавры с большей вероятностью будут классифицированы как манирапторанские тероподы. [ 33 ] Сама Therizinosauria была построена в 1997 году Руселлом, чтобы сдержать все эти тероподы. Это новое инфрамодерство состояло из Therizinosauroidea и более продвинутых Therizinosauridae. [ 34 ] Семейству Therizinosauridae впервые было дано филогенетическое определение Полом Серено в 1998 году, который определил его как всех динозавров ближе к Erlikosaurus, чем к Ornithomimus . [ 35 ]
В 2010 году Линдсей Занно провела наиболее подробный филогенетический анализ Thrizinosauria к этому моменту. Она сослалась на недоступность, ущерб, потенциальную потерю образцов голотипа, дефицит черепных останков и фрагментарные образцы с небольшим количеством перекрывающихся элементов в качестве наиболее значительных препятствий для разрешения эволюционных отношений внутри группы. Эти факторы влияли на положение Segonosaurus и позиции некоторых других азиатских теризинозавридов; Zanno заявил о более хорошо сохранившихся образцах, и потребуется повторное открытие отсутствующих элементов. Zanno также пересмотрел Therizinosauroidea, чтобы исключить Falcarius и сохранил его в более широкой кладке Therizinosauria, которая стала старшим синонимом Segonosauria. Кроме того, она определила Therizinosauridae как наименее инклюзивная клада, содержащая Erlikosaurus , Nothronychus , Segnosaurus и Therizinosaurus . [ 36 ] Фалькарий и Цзянчангозавр в настоящее время считаются наиболее примитивными теризинозаврами, в то время как теризинозавра считаются более полученными, что эти два таксона, но менее полученные, чем теризинозавриды. [ 19 ] [ 37 ]
Большинство филогенетических анализов восстанавливают семейство Therzinosauridae, содержащие Erlikosaurus , Erliansaurus , Nanshiungosaurus , Neimongosaurus , Nothronychus , Segonosaurus и Therizinosaurus . Тем не менее, Suzhousaurus обычно восстанавливается в группе. [ 38 ] [ 19 ] [ 39 ] [ 37 ] Therizinosauridae, вдоль горстки названных родов, составляют суперсемейство Thrizinosauroidea, и, наконец, Therizinosauroidea находится в пределах более высокого инфрадострада теризинозаврии. [ 19 ] [ 37 ] Хотя консенсус еще не достигнут, было предложено, чтобы теризинозавры были наиболее примитивной кладкой в Манирапторе , а также родственной клады для овираптозавров . [ 40 ] [ 11 ]
The cladogram below follows the 2019 phylogenetic analysis by Scott Hartman and colleagues, which is largely based on the work from Zanno in 2010. While most therizinosaurids are recovered in relatively traditional, well-established positions, the therizinosauroid "Nanshiungosaurus" bohlini was unusually recovered as таксон теризинозавридов (не показан в кладограмме): [ 37 ]

| Therizinosauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Палеобиология
[ редактировать ]Кормление
[ редактировать ]
В 2009 году Zanno и коллеги заявили, что теризинозавры были наиболее широко считающимися кандидатами на травоядные лица среди теропод на основе небольших, плотно упакованных, грубых задумчиков; копья в форме зубов с относительно низкой скоростью замены; хорошо развитый кератиновый клюв; длинная шея для просмотра; относительно маленькие черепа; очень большая способность кишечника, как указано в окружности ребра в стволе и наружные процессы проживания Ильи; и заметное отсутствие курсороальных адаптаций в задних конечностях. Все эти особенности предполагают, что члены этой семьи питаются растительностью , а также предварительно обрабатывают ее в уст, чтобы начать разрыв целлюлозы и лигнина . Возможно, это еще более верно для теризинозавридов, которые, кажется, еще больше эксплуатировали этих персонажей. Одной из наиболее заметных адаптаций в продвинутых теризинозавридах являются четырехжисские ноги, которые имели полностью функциональную первую цифру, несущую вес, которая, вероятно, была адаптирована для медленного стиля жизни. Занно и его коллеги обнаружили, что Ornithomimosauria , Thrizinosauria и Oviraptorosauria имели либо прямые, либо морфологические доказательства травоядных , что означало бы, что эта диета эволюционировала независимо много раз в коелурозаврских тероподах, либо то, что примитивное состояние группы было, по крайней мере, факультативным гербисловием с карнвари. [ 11 ] Череп теризинозавридов также был специализирован, так как он был, вероятно, был ограничен с похожим на клюв фронтом, . Утверждалось, что этот рострум, вероятно, был покрыт кератиновым клювом, адаптацией, которая могла бы помочь повысить стабильность черепа путем смягчения стресса и напряжения, испытываемого черепом во время кормления. [ 41 ] [ 25 ]
Как указывает их соответствующие зубные морфологии, современные теризинозавриды Erlikosaurus и Segonosaurus были разделены нишевыми распределением . Эти дифференциации включают относительно нечеткие и симметричные зубы с умеренными зубцами ( зубцами ) в Erlikosaurus и увеличенными зубцами в Segnosaurus , состоящих из дополнительных каринов и сложенных каринов с зубчатыми передними краями, которые вместе создали шероховаренную, измельченную поверхность вблизи зуба зубчатого зубца, которые вместе создали шероховатую поверхность измельчения рядом с базой зубного зуба, которые вместе создали шероховатую поверхность из -за базы зубного зуба, которые вместе создали шероховаренную, измельчивающуюся поверхность вблизи карины. Короны, которые, по -видимому, были уникальны для Segonosaurus и предполагают, что они потребляли уникальные пищевые ресурсы или использовали высокоспециализированные стратегии кормления, и имели более высокую степень орального обработки пищевых продуктов, чем другие теризинозавриды. [ 25 ] В дополнение к этим морфологическим различиям, в кнопке 2019 года и Занно отмечают, что травоядные динозавры следовали двум основным режимам кормления. Одним из них была обработка пищи в кишечнике, которая характеризуется гранильными черепами и относительно низкими силами укуса. Вторым была оральная пищевая обработка, характеризующаяся функциями, связанными с обширной обработкой, такими как нижние челюсти или зубное зуб. Было обнаружено, что Segnosaurus находится в первом режиме, тогда как Erlikosaurus с большей вероятностью попадает во вторую группу, что указывает на то, что эти два теризинозаврида были функционально разделены и заняты различными нишами. [ 42 ] В том же году Али Набавизаде пришел к выводу, что большинство теризинозавров были в основном ортальскими кормушками (перемещающими их челюсти вверх и вниз, а не к бокам) и поднимали их челюсти изогнуто, в результате чего верхние и нижние зубы каждой стороны сразу связывались друг с другом. [ 43 ]
Тем не менее, у продвинутых поздних меловых теризинозавридов имели относительно слабые силы укуса по сравнению с примизитивными теризинозаврами. В 2013 году палеонтолог Стефан Лаутеншлагер выполнил цифровые реконструкции для черепной мускулатуры в Эрликозавре и обнаружил, что сила укуса Эдмонтозавра была больше, чем для первого. Меньшая сила укуса для Erlikosaurus для раздевания и обрезки лучше подается в листьях , а не для активного жевания. [ 44 ] В более новом исследовании с большим количеством таксонов теризинозавра Lautenschlager обнаружил аналогичные выводы. Эволюционные тенденции в механике челюсти теризинозавров заметили изменение силы укуса во времени, от более высоких сил укуса у ранних членов до меньших в усовершенствованных теризинозавридах. Теризинозавридам помогали преуменьшенные кончики и симфизарные регионы (объединение между костями) зубных сил , а также, вероятно, также клювами. В отличие от этого, прямые и более удлиненные зубные зубные зубные зубные зубные зубные зубные зубные зубные зубцы примизинозавров имели самые высокие величины напряжения и напряжения. Вниз понижающее движение головы в то время как захватывающая растительность было более вероятным, чем боковое или вверх движение, хотя такое поведение будет более вероятным у тезинозавридов с их смягчающими стрессом челюстями. [ 45 ]
Пищу
[ редактировать ]
Среди теризинозавридов передние конечности были увеличены в надежности, и гибкость запястья также была увеличена, при этом предполагаемый охват для кормления, вероятно, удлинялся. Более того, грудное поясное пояс было модифицировано для дальнейшего увеличения вертикального досягаемости, однако, как полагают, снизилась способность животных схватки. Эти адаптации более связаны с помощью их травоядного образа жизни, поскольку они специализируются на способности собирать и собирать растительность. [ 12 ] Более того. В теризинозавридах Neimongosaurus диапазон движения в руках был примерно круглым в суставе гленоид-губерта в плече, и направлялся в сторону и слегка вниз, что расходилось от более овальных, обратно-направленных диапазонов других теропод Полем Эта способность значительно расширять свои руки вперед, возможно, помогла этим теризинозавридам достичь и понять листву. [ 46 ] В 2014 году Lautenschlager проверил биомеханическую функцию множественных когтей теризинозавра. Он отметил, что руки некоторых теризинозавридов (такие как nothronyhus или thrizinosaurus ) были более эффективными при пронзительном или стягивании растительности. Руки должны были бы иметь возможность расширить диапазон животного до такой степени, что голова не может быть достигнута, если бы они использовались для просмотра и стягивания растительности. В родах, где сохраняются элементы как шеи, так и передней передней части, шейки были равны по длине или дольше, чем передние конечности, поэтому тягает растительность, вероятно, только в том случае, если более низкие части длинных ветвей были сняты для доступа к выходу из растительности. Lautenschlager также обнаружил, что когти Thrizinosaurid не использовались бы для копания, что было бы сделано с когтями для ног, потому что, поскольку, как и в других манирапторах, перья на передних конеках были бы мешали этой функции. Кроме того, это действие приводит к более высокому напряжению напряжения на дорсальной области когти - это более очевидно в Теризинозавр . Тем не менее, он не мог ни подтвердить, ни игнорировать, что когтичные когти могли быть полностью использованы для сексуального демонстрации , самообороны, внутривидовой конкуренции , захвата спаривания во время спаривания или стабилизации при приборе пищи . [ 47 ]
Мозг и чувства
[ редактировать ]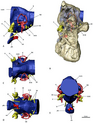
Группа также известна адаптацией к структурам их уха . Структура их внутреннего уха почти птица, с птичьими полукруглыми каналами и расширенной улиткой . Для птиц расширенная улитка позволяет им услышать в рамках увеличенного диапазона частот , что указывает на аналогичную функцию в странных теризинозавридах, а также дает им хороший слух и баланс, что, действительно, являются чертами, лучше связанными с плотоядными тероподами . Кроме того, удлинение улитки, адаптация, которая независимо развивалась в ряде других тероподных групп, как полагают, еще больше улучшает слуховую хватку. Передний мозг теризинозавридов был довольно увеличен и развился с удлиненными обонятельными луковицами . Адаптация к внутреннему уху и переднему мозгу теризинозавридов, вероятно, выполняли ряд функций, таких как хорошо развитые запаха , сложное социальное поведение , повышенная настороженность вокализации несовершеннолетни чувства Пневматические камеры в сенсорных областях в черепах теризинозавридов ( Erlikosaurus или Nothronychus McKinleyi ) указывают на то, что барабанные системы приведут к увеличению и оптимальному низкочастотному звуковому приему, возможно, внедрены . Таково случай с Н. МакКинлейи , который имел среднюю частоту слуха от 1100 до 1450 Гц и возможные верхние пределы между 3000 и 3700 Гц. Особенности включают не только обширную базовую пневматичность, но и развитие базисфеноидной буллы (полая костная структура). Кроме того, ориентация горизонтального полукруглого канала относительно горизонтальной ориентации затылочного мыщелка дала теризинозавридам горизонтальную осанку головки, которая позволила бинокулярному зрению с перекрывающимися визуальными полями. Подавляющее большинство этих чувств также было хорошо разработано в более ранних коелурозаврах и других тероподах, что указывает на то, что теризинозавриды унаследовали многие из этих признаков от их мелких плотоядных манирапторов Диетические цели. [ 48 ] [ 49 ]
Локомоция
[ редактировать ]
В 1964 году Захаров описал и назвал конкретного Ichnogenus Macropodosaurus , который представлен серией с четырьмя точками . Эти треки были найдены в постели сеноманского возраста в Таджикистане , отметив, что они были сделаны двуногим четырехпеханием динозавра , а пальцы ног были перепончаты или, по крайней мере, очень мясистые. Маловероятно, что они были сделаны четвероногим животным, поскольку не было обнаружено никаких ручных следов. Следы имеют длину около 50 см (500 мм) и ширину 30 см (300 мм). Поскольку исследование теризинозавров началось сзади к этим результатам, Захаров не смог определить точный тип динозавра. [ 50 ] Сенников в 2006 году пересмотрел эти следы и пришел к выводу, что динозавр теризинозавридов мог бы сделать эти следы. Он сравнил макроподозавра с сочлененными ногами Erlikosaurus и упомянутым из Therizinosaurus, показывающий, что при сочленении в положении Plantigrade они вписываются в морфологию макроподозавра . Поэтому он считал, что эти треки более связаны с теризинозавридами и был одним из первых в предложении Plantigrade Stance у теризинозавридов. [ 51 ] Дополнительный след макроподозавра был обнаружен в Польше и впервые зарегистрировался в 2008 году. Этот след напоминает треки, описанные Захарова, и, следовательно, назначенный Macropodosaurus sp. Эти треки могут указывать на наличие теризинозавров в Европе . [ 52 ] В 2017 году Masrour с коллегами сообщил, что Macropodosaurus Footprints в том, что сейчас является Марокко . Они похожи на первоначальное описание с некоторой степенью полуплантиградизма. [ 53 ]
Тем не менее, некоторые другие следы могут указывать на более цифровую позицию. Так обстоит случай, снятые в формировании Cantwell , помеченные в цифрах DMNH 2010-07-01, 2013-08-04, 2013-08-06 и 2014-11-05. Эти впечатления составляются четырьмя пальцами с первой цифрой, немного меньшей, чем другие, что объясняется теризинозавридами. По крайней мере, один след, DMNH 2010-07-01, был непосредственно сравнивать с относительно полными правыми PE Erlikosaurus, выявляющими последовательную морфологию. Тем не менее, большая часть следа была сделана (по -видимому) пальцами и подушкой, указывающей на более вертикальное положение плюсневых средств. [ 54 ] [ 55 ] Тем не менее, Botelho и его коллеги также считали позицию Plantigrade для теризинозавридов в 2016 году. Соответственно, члены Therizinosauridae (такие как Neimongosaurus или Nothronychus ) повторно развили полностью функциональную первую цифру, которая артикулирует на Ankle . [ 56 ]
Размножение
[ редактировать ]Гнездование
[ редактировать ]



Некоторые из первых дендрулитидных яиц , которые приписываются теризинозаврам, были сообщены из формаций Баян -Шире и Нанчао в том же году, 1997 года. Они состояли из нескольких яичных сцеплений (группа) со средним составом 7 или более яиц. [ 57 ] [ 58 ] [ 59 ] Кроме того, формация Bayan Shireh вызвала полностью выращенные, специфические таксоны теризинозавридов, такие как Erlikosaurus и Segonosaurus . [ 5 ] [ 25 ] Яичные гнезда из формирования Нанчао оставались неписанными в течение нескольких лет, и только кратко изучалось, но идентифицировали как содержащие окаменелые эмбрионы . [ 60 ] [ 61 ] Однако в 2007 году они были описаны палеонтологом Мартином Кундратом и его коллегами и предварительно идентифицированными как теризинозавриды на основе анатомических особенностей, таких как дорезость без зубов с спаданным краем, зубной зубной зуб Выдающийся дельтопекторный гребень, подвздошной подвздошной кости с расширенным передним концом и удлиненные, резко установленные ручные мануалы. Большинство яиц имеют средний размер 7 см × 9 см (70 мм × 90 мм), и, учитывая эти размеры, они, вероятно, были уложены самкой среднего размера. Хотя было обнаружено несколько яичных сцеплений, было найдено, что было 7 яиц, из которых 3 из них сохраняли эмбрионы. [ 62 ] В 2019 году Хартман и его коллеги были первыми авторами, которые включили эти эмбрионы в филогенетический анализ, и, как и ожидалось, эмбрионы были извлечены в качестве теризинозавридов. [ 37 ]
В реферате на конференции 2013 года палеонтолог Йошицугу Кобаяши и его коллеги сообщили о исключительном месте гнездования динозавров теропод в формировании Джавкхланта , которое содержало не менее 17 яичных сцеплений из того же слоя в районе 22 м на 52 м. Каждая муфта содержала сферические яйца, которые находились в контакте друг с другом и расположены в круговой структуре без центрального отверстия. Основываясь на микроскопических особенностях яичной скорлупы, они идентифицировали яйца как дендрулитиды, которые ранее были приписаны динозаврам теризинозавра. Многочисленные сцепления указывают на то, что некоторые теризинозавриды были колониальными гнездами , и тот факт, что они были обнаружены в одном стратиграфическом слое, предполагает, что они вложенные на участок в одном случае и, следовательно, не проявляли филопатрического поведения . [ 63 ] Это место гнездования было официально описано в 2019 году Кохей Танакой и его коллегами. В этом исчерпывающем описании количество яичных сцеплений было исправлено не менее 15, отметив, что яйца были отложены в сцеплениях, состоящих из 3-30 яиц и вылуплены в тот же сезон единого гнездования. Яйца имеют средний диаметр 13 см (130 мм) с некоторыми изменениями между 10 см (100 мм) до 15 см (150 мм). Основываясь на гладкой внешней поверхности, как внутренних, так и внешних частях, и нескольких пор-нерегулярных пор этих яиц, они были подтверждены, чтобы они были относятся к Dendroolithidae. Приблизительно 50% зоны гнездования было разрушено, и, основываясь на распределении яичных сцеплений, плотность гнезда может быть оценена около 1 гнезда на 10 м. 2 Полем Это указывает на то, что первоначально присутствовало до 32 гнезд. Среда обитания, в которую вложенные родители, была полузасушливая пойма , а яичные сцепления были покрыты в органическом материале во время инкубации, как и некоторые существующие архозавры сегодня ( крокодилы и мегаподные птицы ). Кроме того, ассоциация фрагментов яичной оболочки указывает на то, что многие из сцеплений вылупились до того, как участок был похоронен в результате наводнения. Из 15 сцеплений, по крайней мере, 9 успешно вылупились, что представляет собой показатель успеха в гнездовании не менее 60% для всего участка. [ 64 ]
Выдающийся мест гнездования формирования Джавкхланта указывает на то, что колониальное гнездование сначала развивалось у неавийских видов динозавров, чтобы повысить успех вылупления в экосистемах, подверженных высоким давлению хищничества гнезда (например, это образование). Наконец, хотя яйца дендролититов также приписываются мегалозавроидам , а теризинозавриды не известны из формации Джавкхланта, оно перекрывает формацию Баян -Шире, где другие дендруолитидные яйца и энгмозавр , эрлосозавр и сегенозавр были обнаружены . Более того, примечательно отсутствие мегалозавроидов в этих формациях. [ 64 ]
Онтоген
[ редактировать ]
По меньшей мере четыре эмбриональных этапа развития были зарегистрированы от эмбрионов Нанчао; Кундрат и его коллеги классифицировали их с этапов А до D в зависимости от развития. Стадия А является самым ранним из всех и характеризуется плохим окостенением костей и пористой структурой центра в позвоночнике . На этапах B и C кости становятся немного более сочлененными и окостененными. Похоже, что они соответствуют уровням развития 45–50 и 64-дневными эмбрионами американского аллигатора . Последняя и более продвинутая стадия - это D, где эмбрионы полностью окостяли позвоночные и частично уменьшенный нейроцентральный швар в их шейных позвонках. Кроме того, большинство теризинозаврских персонажей более печально известны на этом этапе, например, достойная предчелюстная деятельность . Наиболее зрелый эмбрион представлен CAGS-01-IG-5, который имел явно более развитое оксификацию, чем птенца аллигатора. Это указывает на то, что эмбриональные теризинозавриды достигли более зрелого скелета, чем другие птенца архозавра в OVO и оставался в яйце в течение более длительного периода, чтобы увеличить свои пропорции, несмотря на продвинутую окостенение. Основываясь на состояниях развития, сохранившихся в эмбрионах, Кундрат и его коллеги предположили грубый инкубационный период от 1,5 до 3 месяцев. [ 62 ]
Учитывая, что у некоторых эмбрионов были очерчились свои кости, и тот факт, что ни один взрослый не был обнаружен в связи с гнездами, птенцы теризинозавридов были очень предварительными (способными к локомоции с рождения) и способными оставлять свои гнезда для кормления в одиночку, независимо от своих родителей. Подземно сконструированные гнезда могут свидетельствовать об отсутствии родительской помощи в течение инкубационного периода, кроме того, скелетная зрелость эмбрионов стадии D была значительно адаптирована, чтобы обеспечить немедленную локомоцию после рождения, потенциально предполагая сверхпериальное поведение. Независимость от их родителей также может быть подтверждена развитием зубов детенышей, которые соответствуют всеядной диете. [ 62 ]
Палеоэкология
[ редактировать ]
Теризинозавриды были очень специализированными травоядами, которые развили конвергентный образ жизни в более поздних наземных лени . В некоторых аспектах они настолько похожи, что эта идея также отражена во имя нескольких видов, таких как Nothronychus (лень когти) или Suzhousaurus megatherioides ( megatherium -like). [ 47 ] Вносяные их анатомические и физиологические признаки, такие как листовые, грубо зубчатые зубы, сильная рука с большими когтями, общеизвестно вытянутая шея и развитие кератинового клюва, расположенного теризинозавридами как травоядные браузеры . с высоким просмотром Более конкретно, теризинозавриды обитали в нишах в своих экосистемах и обычно жили в полузасушливых до водно -болотных мест обитания, состоящих из высокой растительности, как видно из речной лакростиновой обстановки большинства образцов. [ 27 ] [ 55 ] [ 65 ]
Как отражается как минимум 31 следа теризинозавридов на Кантвелл формировании Аляски . , некоторые виды образовали малые стада , что согласуется со сложной структурой мозга и ушей в этих тероподах Совместное появление с хадрозавридами в этой области может также указывать на то, что эти очень разные динозавры извлекли выгоду из экологического взаимодействия, так же, как некоторые животные сегодня собираются по взаимным полезным причинам, таким как приобретение дополненного ресурса или меньшее давление хищничества. Также возможно, что стадо гадрозавридов и теризинозавридов проходило через эту местность в разное время и не сталкивалось друг с другом. Независимо от этих объяснений, трек Thrizinosaurid отражает общительное поведение в этих тероподах. [ 55 ]
Распределение
[ редактировать ]
Хотя подавляющее большинство всех теризинозавридов вышли из Азии (особенно из Китая и Монголии ), [ 36 ] [ 39 ] Относительно полные останки Nothronychus были найдены за пределами Азии в Северной Америке . [ 12 ] Хотя ископаемые записи теризинозавридов указывают на то, что семья процветала вблизи туронской стадии позднего мела , на стадии Барремиц уже было панарасианское распределение теризинозавра. С тех пор, как Лауразия начала развалиться во время мелового периода, возникли две теории, чтобы объяснить, как теризинозавры могли охватывать по всей Лорасии. Одна теория заключается в том, что примитивные члены уже присутствовали как в Азии, так и в Северной Америке до того, как она начала распадаться, предполагая появление для теризинозавров, по крайней мере, позднего триаса. что группа рассеялась между Азией и Северной Америкой через «меловой» беринговой сухопутной мост во время берриасского валангинского Вторая теория заключается в том , этапа. Это последнее предложение подтверждается присутствием теризинозавридов в Северной Америке во время раннего позднего мела, таких как Nothronychus . [ 36 ] Тем не менее, следы теризинозавридов известны из отдаленных мест, таких как Европа или Африка. В Польше было обнаружено несколько четырех треков, и в Марокко были обнаружены серия следов, начиная с позднего мела. Оба результата могут указывать на то, что теризинозавриды были гораздо более диспергированы, чем считалось ранее. [ 52 ] [ 53 ]
Идея сухопутного моста еще больше поддерживается множественным совместным появлением следах Хадразаврида и Теризинозаврида в нижней части Формирования Кантвелла Национального парка Денали , что отражает важный обмен фауны между сухопутными массами . Эти треки показывают доминирующее присутствие гадрозавридов и сниженную концентрацию теризинозавридов в том же месте в потенциально в одно и то же время. Как указывает обнаружение от впечатления, подобного водяным, представляющего один ископаемый лист из того же места, пульс был сделан динозаврами, когда они пересекали мелкий водоем вдали от основных речных каналов, что любопытно, учитывая, что во время Позднее меловое эта часть Северной Америки была полузасушливой средой обитания. Это происшествие в этой единственной местности в нижней части Формирования Кантвелла не было задокументировано в другом месте в Северной Америке, и эти пути представляют собой первую зарегистрированную встречу между общеизвестно различными динозаврами из Северной Америки. Разнообразие iChnotaxa на этом сайте поддерживает идею аналогичных фаун динозавров между Аляской и Азией в поздний меховой период, в частности, с Формирование Немегта, которое имело относительно влажную среду. Фиорилло и его коллеги предположили, что Аляска представляла «ворота» для обмена фауны между двумя континентальными супругами и существованием «мелового» берингового сухопутного моста. Полем [ 55 ]
Смотрите также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а беременный в Maleev, E. A. (1954). "Новый черепахообразный ящер в Монголии" [New turtle like reptile in Mongolia]. Природа [ Priroda ] (in Russian) (3): 106−108. Translated paper .
- ^ Jump up to: а беременный в Барсболд Р. (1976). «Новые данные о Thrizinosaurus (Thirizinosauridae, Theropoda)». Совместная советско-монгольская палеонтологическая экспедиция (на русском языке). 3 : 76–92.
- ^ Jump up to: а беременный в дюймовый и фон Перл А. (1979). «Segonosauridae - novoe semejstvo teropod iz pozdnego mela mongolii» [Segnosauridae - новое семейство теропод из позднего мела Монголии]. Сделки Совместной советско-монгольской палеонтологической экспедиции (на русском языке). 8 : 45–55. Переведенная бумага
- ^ Jump up to: а беременный в дюймовый и Донг, З. (1979). «Окаменелости меловых динозавров в южном Китае» [меловые динозавры Уаанана (Южный Китай)]. В институте палеонтологии позвоночных и палеоантропологии; Нанкин Институт палеонтологии (ред.). Мезозойские и кайнозойские красные кольца на юге Китая (на китайском языке). Пекин: Science Press. С. 342–350. Переведенная бумага
- ^ Jump up to: а беременный в Barsbuld, R.; Перл А. (1980). (PDF)PDF Acta Poolonological Польша 25 (2): 190–1
- ^ Jump up to: а беременный Перл А. (1981). «Novyy Segnozavrid Iz verkhnego mela mongolii» [New Segnosauridae из верхнего мела Монголии]. Сделки Совместной советско-монгольской палеонтологической экспедиции (на русском языке). 15 : 50–59. Переведенная бумага
- ^ Jump up to: а беременный в Перл А. (1982). «Задняя конечность Теризинозавра из верхнего мела Монголии». Проблемы в монгольской геологии (по -русски). 5 : 94–98. Переведенная бумага
- ^ Jump up to: а беременный в Zhang, X.-H.; Сюй, х.; Zhao, Z.-J.; Серено, ПК; Kuang, X.-W.; Тан Л. (2001). «Терризинозавроидный динозавр с длинной шеей из верхнего мелового ирена-дабасу формирования Ней Монгола, Китайская Народная Республика» (PDF) . Повертеблат Паласиатика . 39 (4): 282–290.
- ^ Киркленд, Джи; Wolfe, DG (2001). «Первый окончательный теризинозаврид (Dinosauria; Theropoda) из Северной Америки». Журнал палеонтологии позвоночных . 21 (3): 410–414. doi : 10.1671/0272-4634 (2001) 021 [0410: fdtdtf] 2.0.co; 2 . JSTOR 20061971 . S2CID 85705529 .
- ^ Jump up to: а беременный в дюймовый Сюй, х.; Zhang, Z.-h.; Секрет, ПК; Чжао, X.-J.; Ключ, X.-W.; Хан, Дж.; Тан Л. (2002). ( PDF) . велебр Паласиатическое 40 : 228–2
- ^ Jump up to: а беременный в дюймовый Занно, Ле; Джилетт, DD; Олбрайт, LB; Titus, AL (2009). «Новый североамериканский теризинозаврид и роль травоядных в эволюции хищных динозавров» . Труды Королевского общества б . 276 (1672): 3505–3511. doi : 10.1098/rspb.2009.1029 . JSTOR 30244145 . PMC 2817200 . PMID 19605396 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж k л м Хедрик, BP; Занно, Ле; Вулф, DG; Додсон, П. (2015). «Тлевочный когти: остеология и тафономия Нотрониха МакКинлейи и Н. Граффами (Динозаурия: Теропода) и анатомические соображения для полученных теризинозавридов» . Plos один . 10 (6): E0129449. BIBCODE : 2015PLOSO..1029449H . doi : 10.1371/journal.pone.0129449 . PMC 4465624 . PMID 26061728 .
- ^ Хольц, Тр; Рей, Л.В. (2007). Динозавры: самая полная, современная энциклопедия для любителей динозавров всех возрастов . Случайный дом. ISBN 9780375824197 Полем Список родов для Holtz 2012 Информация о весах
- ^ Jump up to: а беременный в Пол, GS (2016). Полевой гид Принстона по динозаврам (2 -е изд.). Принстон, Нью -Джерси: издательство Принстонского университета. С. 162–168. ISBN 9780691167664 .
- ^ Ты, ч.; Morschhauser, Em; Li, D.; Додсон, П. (2018). «Представление фауны динозавров Мазоншан». Журнал палеонтологии позвоночных . 38 (SUP. 1): 1–11. Bibcode : 2018jvpal..38s ... 1y . doi : 10.1080/02724634.2017.1396995 . S2CID 202867591 .
- ^ Бенсон, RBJ; Кампионе, NE; Каррано, MT; Mannion, PD; Салливан, C.; Upchurch, P.; Эванс, округ Колумбия (2014). «Уровень эволюции массы тела динозавров указывают на 170 миллионов лет устойчивых экологических инноваций на линии птичьего стебля» . PLOS Биология . 12 (6): E1001896. doi : 10.1371/journal.pbio.1001853 . HDL : 10088/25994 . PMC 4011683 . PMID 24802911 .
- ^ Сюй, х.; Тан, З.-Л.; Ван, XL (1999). «Теризинозавроидный динозавр с покровными структурами из Китая». Природа . 339 (6734): 350–354. Bibcode : 1999natur.399..350x . doi : 10.1038/20670 . ISSN 1476-4687 . S2CID 204993327 .
- ^ Сюй, х.; Zheng, x.; Вы, H. (2009). «Новый тип перьев в неавийском тероподе и ранней эволюции перьев» . Труды Национальной академии наук . 106 (3): 832–834. doi : 10.1073/pnas.0810055106 . PMC 2630069 . PMID 19139401 .
- ^ Jump up to: а беременный в дюймовый PU, H.; Kobayashi, Y.; Lü, J.; Сюй, Л.; Wu, y.; Чанг, H.; Zhang, J.; Цзя, С. (2013). «Необычный базальный динозавр теризинозавра с орнитишским стоматологическим расположением из северо -восточного Китая» . Plos один . 8 (5): E63423. BIBCODE : 2013PLOSO ... 863423P . doi : 10.1371/journal.pone.0063423 . PMC 3667168 . PMID 23734177 .
- ^ Li, Q.; Кларк, JA; Гао, К.-Q.; Zhou, C.-F.; Meng, Q.; Li, D.; D'Alba, L.; Shawkey, MD (2014). «Эволюция меланосом указывает на ключевой физиологический сдвиг в пернатых динозаврах» . Природа . 507 (7492): 350–353. Bibcode : 2014natur.507..350L . doi : 10.1038/nature12973 . PMID 24522537 . S2CID 4395833 .
- ^ Dimond, CC; Каюта, RJ; Брукс, JS (2011). «Перья, динозавры и поведенческие сигналы: определение гипотезы визуального дисплея для адаптивной функции перьев в неавийских тероподах». Биос . 82 (3): 58–63. doi : 10.1893/011.082.0302 . JSTOR 23033900 . S2CID 98221211 .
- ^ Макнамара, я; Чжан, Ф.; Кернс, SL; Orr, PJ; Toulouse, A.; Фоли, Т.; Hone, DW E; Роджерс, CS; Бентон, MJ; Джонсон, Д.; Сюй, х.; Zhou, Z. (2018). «Окаменечная кожа выявляет коэволюцию с перьями и метаболизмом у пернатых динозавров и ранних птиц» . Природная связь . 9 (2072): 2072. Bibcode : 2018natco ... 9.2072M . doi : 10.1038/s41467-018-044443-x . ISSN 2041-1723 . PMC 5970262 . PMID 29802246 .
- ^ Zanno, Le (2010). «Остеология Falcarius utahensis (Dinosauria: Theropoda): Характеристика анатомии базальных теризинозавров» . Зоологический журнал Линневого общества . 158 (1): 196–230. doi : 10.1111/j.1096-3642.2009.00464.x .
- ^ Jump up to: а беременный в дюймовый Lautenschlager, S.; Лоуренс, MW; Перл, А.; Занно, Ле; Эмили, младший (2014). «Анатомия черепа Эрликозавра Эндрюси (Dinosauria, Thrizinosauria): новые идеи, основанные на цифровой реконструкции». Журнал палеонтологии позвоночных . 34 (6): 1263–1291. Bibcode : 2014jvpal..34.1263L . doi : 10.1080/02724634.2014.874529 .
- ^ Jump up to: а беременный в дюймовый и фон Занно, Ле; Tsogtbaatar, K.; Chinzorig, T.; Гейтс, для (2016). «Specialiizations анатомии нижней челюсти и зубной промышленности Segonosaurus galbinsis (Ropoda: Therizininosauria) » ПЕРЕЙ 4 : E1885. Doi : 10.7717/ peerj.1 PMC 4824891 . PMID 27069815
- ^ Занно, Ле; Маковицки, PJ (2011). «Травоядные экоморфологии и специализации в эволюции динозавров Theropod» . Труды Национальной академии наук . 108 (1): 232–237. Bibcode : 2011pnas..108..232Z . doi : 10.1073/pnas.1011924108 . PMC 3017133 . PMID 21173263 .
- ^ Jump up to: а беременный Senter, P.; Джеймс, RH (2010). «Высота бедра гигантских динозавров в области теропода Deinocheirus mirificus и Therizinosaurus cheloniformis, а также последствия для музея и палеоэкологии» (PDF) . Бюллетень музея естественной истории Ганма (14): 1–10.
- ^ Barsbold, R.; Перл А. (1979). «Modiphikatsiy Tasa Sayrisziy I Parallelinoe Rasvitie Zishchnich Dinosavrov» [Модификация в таз Сауриша и параллельное развитие хищных динозавров]. Труды совместной советской монгольской палеонтологической экспедиции (на русском языке). 8 : 39–44. Переведенная бумага
- ^ Пол, Г.С. (1984). «Динозавры снезавров: реликвии перехода Prosauropod-ornithischian?». Журнал палеонтологии позвоночных . 4 (4): 507–515. Bibcode : 1984jvpal ... 4..507p . doi : 10.1080/02724634.1984.10012026 . ISSN 0272-4634 . JSTOR 4523011 .
- ^ Пол, Г.С. (1988). Хищные динозавры мира . Нью -Йорк: Саймон и Шустер. С. 185–283. ISBN 9780671619466 .
- ^ Barsbold, R.; Maryyanska, T. (1990). "Saurischia sedis mutilis: segonosauria" В Вайшампеле, дБ; Осмольска, H.; Додсон, П. (ред.). Динозаврия (1 -е изд.). Беркли: Университет Калифорнийской прессы. Стр. 408–4 ISBN 9780520067271 .
- ^ Рассел, да; Донг, З. (1993). «Природство нового теропода из пустыни Альса, внутренней Монголии, Китайской Народной Республики». Канадский журнал наук о Земле . 30 (10): 2107–2127. Bibcode : 1993cajes..30.2107r . doi : 10.1139/e93-183 .
- ^ Кларк, JM; Перл, А.; Норэлл, М. (1994). «Череп Эрликозавра Эндрюси, позднего мела -сеназавра (Theropoda, Therizinosauridae) из Монголии». Американский музей новичок (3115): 1–39. HDL : 2246/3712 .
- ^ Рассел, Д.А. (1997). "Therizinosasuria" В Керри, PJ; Священник, К. (ред.). Энциклопедия динозавров Сан -Диего: академическая пресса. стр. 729–3 ISBN 978-0-12-226810-6 .
- ^ Серено, ПК (1998). «Рациональный для филогенетических определений, с применением к таксономии динозавров более высокого уровня (PDF) . Новый ежегодный книга по геологии и палеонтологии - трактаты . 210 (32): 41-83. Doi : 10.1127/njgpa/210/998/41 .
- ^ Jump up to: а беременный в Zanno, Le (2010). Полем Журнал . 8 (4): 503–5 doi : 10.1080/ 1472019.480.48045 S2CID 53405097 .
- ^ Jump up to: а беременный в дюймовый и Hartman, S.; Мортимер, М.; Wahl, WR; Ломакс, доктор; Lippincott, J.; Lovelace, DM (2019). «Новый паравийский динозавр из позднего юрского периода Северной Америки поддерживает позднее приобретение полета птиц» . ПЕРЕЙ . 7 : E7247. doi : 10.7717/peerj.7247 . PMC 6626525 . PMID 31333906 .
- ^ Zanno, LE (2006). «Групный пояс и передние мыши примитивного теризинозавра Falcarius utahensis (Theropoda, Maniraptora): анализ эволюционных тенденций в Therizinosauroidea» . Журнал палеонтологии позвоночных . 26 (3): 636–650. doi : 10.1671/0272-4634 (2006) 26 [636: tpgafo] 2.0.co; 2 . JSTOR 4524610 . S2CID 86166623 .
- ^ Jump up to: а беременный Суды, H.D.; Averianov, A. (2016). «Therizinosauroidea (Dinosauria: Theropoda) из верхнего мела Узбекистана». Мерашные исследования . 59 : 155–178. Bibcode : 2016crres..59..155s . doi : 10.1016/j.cretres.2015.11.003 .
- ^ Смит, DK; Занно, Ле; Сандерс, RK; Deblieux, DD; Киркленд, Джи (2011). «Новая информация о мозгах североамериканского теризинозаврского (Theropoda, Maniraptora) Falcarius utahensis» . Журнал палеонтологии позвоночных . 31 (2): 387–404. Bibcode : 2011jvpal..31..387s . doi : 10.1080/02724634.2011.549442 . JSTOR 25835833 . S2CID 73715992 .
- ^ Lautenschlager, S.; Witmer, LM; Перл, А.; Рэйфилд, EJ (2013). «Эдентулизм, клювы и биомеханические инновации в эволюции динозавров теропод» . ПНА . 110 (51): 20657–20662. Bibcode : 2013pnas..11020657L . doi : 10.1073/pnas.1310711110 . JSTOR 23761610 . PMC 3870693 . PMID 24297877 .
- ^ Кнопка, DJ; Zanno, Le (2019). «Повторная эволюция дивергентных способов травоядных в неавийских динозаврах» . Текущая биология . 30 (1): 158–168.e4. doi : 10.1016/j.cub.2019.10.050 . PMID 31813611 . S2CID 208652510 .
- ^ Набавизаде А. (2019). «Караниальная мускулатура у травоядных динозавров: обзор реконструированных анатомических механизмов разнообразия и кормления» . Анатомическая запись . 303 (4): 1104–1145. doi : 10.1002/ar.24283 . PMID 31675182 . S2CID 207815224 .
- ^ Lautenschlager, S. (2013). «Черноальная миология и силу укуса Эрликозавра Эндрюси: новый подход к цифровым мышечным реконструкциям» . Журнал анатомии . 222 (2): 260–272. doi : 10.1111/joa.12000 . PMC 3632231 . PMID 23061752 .
- ^ Lautenschlager, S. (2017). «Функциональное разделение ниши в Thrizinosauria дает новое понимание эволюции травоядного теропода» (PDF) . Палеонтология . 60 (3): 375–387. Bibcode : 2017Palgy..60..375L . doi : 10.1111/pala.12289 . S2CID 90965431 .
- ^ Burch, SH (2006). «Диапазон движения гленумерного сустава теризинозавра Neimongosaurus Yangi (Dinosauria: Theropoda)» . Журнал палеонтологии позвоночных . 26 (Supp. 3): 46a. doi : 10.1080/02724634.2006.10010069 . S2CID 220413406 .
- ^ Jump up to: а беременный Lautenschlager, S. (2014). «Морфологическое и функциональное разнообразие в когтях теризинозавра и последствия для эволюции тероподного когтя» . Труды Королевского общества б . 28 (1785): 20140497. DOI : 10.1098/rspb.2014.0497 . PMC 4024305 . PMID 24807260 .
- ^ Стефан, Л.; Эмили, младший; Перл, А.; Линдси, Эз; Лоуренс, MW (2012). «Эндокраниальная анатомия Теризинозаврии и ее последствия для сенсорной и когнитивной функции» . Plos один . 7 (12): E52289. BIBCODE : 2012PLOSO ... 752289L . doi : 10.1371/journal.pone.0052289 . PMC 3526574 . PMID 23284972 .
- ^ Смит, DK; Сандерс, RK; Wolfe, DG (2018). «Переоценка основных мягких тканей и пневматичности теризинозаврского Nothronychus McKinleyi (Theropoda; Maniraptora)» . Plos один . 13 (7): E0198155. BIBCODE : 2018PLOSO..1398155S . doi : 10.1371/journal.pone.0198155 . PMC 6067709 . PMID 30063717 .
- ^ Захаров, С.А. (1964). «На сеноманском динозавре, треки которого были найдены в долине Ривер Ширкент». В Реймане, Вм (ред.). Палеонтология Таджикистана (на русском языке). Душанбе: Академия наук Tajik SSR Press. С. 31–35.
- ^ Sennikov, AG (2006). «Чтение треков Segonosaur». Природа (на русском языке). 5 : 58–67.
- ^ Jump up to: а беременный Gierliński, G. D (2015). «Новые следы динозавров из верхнего мела Польши в свете палеогеографического контекста» . Ichnos . 22 (3–4): 220–226. Bibcode : 2015ichno..22..220G . doi : 10.1080/10420940.2015.1063489 . S2CID 128598690 .
- ^ Jump up to: а беременный Masrour, Moussa; Лкебир, Нура; Pérez-Lorente, Félix (2017). «Анза палеохнологическая площадка. Покойный меховой. Марокко. Часть II. Проблемы больших дорожных путей динозавров и первого африканского макроподозавра. Журнал африканских наук о Земле . 134 : 776–793. Bibcode : 2017jafes.134..776m . doi : 10.1016/j.jafrearsci.2017.04.019 . ISSN 1464-343X .
- ^ Фиорильо, А. Р; Адамс, Т.Л. (2012). «Трасса теризинозавра из Нижней Формирования Кантвелла (верхний меловой) национального парка Денали, Аляска» . Палаис . 27 (6): 395–400. Bibcode : 2012Palai..27..395f . doi : 10.2110/palo.2011.p11-083r . S2CID 129517554 .
- ^ Jump up to: а беременный в дюймовый Фиорильо, Ар; Маккарти, PJ; Kobayashi, Y.; Tomsich, CS; Тайкоски, RS; Lee, Y.-N.; Танака, Т.; Noto, CR (2018). «Необычная ассоциация треков Hadrosaur и Thrizinosaur в поздних меловых скалах национального парка Денали, Аляска» . Научные отчеты . 8 (11706): 11706. Bibcode : 2018natsr ... 811706f . doi : 10.1038/s41598-018-30110-8 . PMC 6076232 . PMID 30076347 .
- ^ Botelho, JH; Smith-Paredes, D.; Soto-Acuña, S.; Нуньес-Леон, Д.; Palma, V.; Варгас, АО (2016). «Большой рост проксимальных плюсневых костюмов у эмбрионов птиц и эволюция положения Hallux в схватке». Журнал экспериментальной зоологии Часть B: молекулярная эволюция и развитие . 328 (1–2): 106–118. doi : 10.1002/jez.b.22697 . HDL : 10533/232005 . ISSN 1552-5015 . PMID 27649924 . S2CID 4952929 .
- ^ Ariunchimeg, Y. (1997). «Результаты исследований яиц динозавров». Монголия -японская палеонтологическая экспедиция . Аннотация Сообщения: 13.
- ^ Watabe, M.; Ariunchimeg, y.; Бринкман Д. (1997). «Яичные гнезда динозавров и их осадочная среда в местности Бейн -Шир (поздний меловой), Восточный Гоби». Монголия -японская палеонтологическая экспедиция . Аннотация Сообщения: 11.
- ^ Мэннинг, TW; Джойси, Ка; Круикшанк, Ари (1997). «Наблюдения за микроструктурами в яйцах динозавров из провинции Хэнань, Китайской Республики Народ». В Вольберге, DL; Пень, e.; Розенберг, Р.Д. (ред.). Dinofest International: Материалы симпозиума, проведенного в Университете штата Аризона . Пенсильвания: Академия естественных наук. С. 287–290.
- ^ Carpener, K. (1999). «Эмбрион и вылупление». Яйца, гнезда и детские динозавры: взгляд на размножение динозавра (жизнь прошлого) . Блумингтон: издательство Университета Индианы. п. 209. ISBN 9780253334978 .
- ^ Kundrát, M.; Круикшанк, Ари; Мэннинг, TW; Nodds, J. (2005). «Восхождение эмбриологии динозавров: обучение на исключительных образцах». Симпозиум парка Динозавров . Расширенные тезисы: 53–55.
- ^ Jump up to: а беременный в Kundrát, M.; Круикшанк, Ари; Мэннинг, TW; Nodds, J. (2007). «Эмбрионы теризинозавра теропод из верхнего мела Китая: диагностика и анализ моделей окостенения» . Acta Zoologica . 89 (3): 231–251. doi : 10.1111/j.1463-6395.2007.00311.x .
- ^ Kobayashi, Y.; Lee, y.; Barsbold, R.; Zelenitsky, D.; Танака, К. (2013). «Первая запись в гнездовой колонии динозавров из Монголии выявляет гнездование теризинозавра». В Максвелле, E.; Миллер-Камп, Дж.; Anemone, R. (Eds.). 73 -е ежегодное собрание общества палеонтологии позвоночных . Лос -Анджелес. п. 155
{{cite book}}: CS1 Maint: местоположение отсутствует издатель ( ссылка ) - ^ Jump up to: а беременный Танака, К.; Kobayashi, Y.; Зеленицкий, DK; Терриен, Ф.; Lee, Y.-N.; Barsbold, R.; Кубота, К.; Lee, H.-J.; Tsogtbaatar, C.; Idersaikhan, D. (2019). «Исключительное сохранение места гнездования динозавров позднего мела от Монголии выявляет колониальное гнездование в неавийском тероподе» . Геология . 47 (9): 843–847. Bibcode : 2019geo .... 47..843t . doi : 10.1130/g46328.1 . S2CID 198412503 .
- ^ Owocki, K.; Кремер, Б.; Cotte, M.; Bocherens, H. (2020). «Диетические предпочтения и климат, выведенные из изотопов кислорода и углерода зубной эмали из Tarbosaurus Bataar (Nemegt Fomaration, верхняя меловая, Монголия)». Палеогеография, палеоклиматология, палеоэкология . 537 : 109190. Bibcode : 202020pp ... 53709190O . doi : 10.1016/j.palaeo.2019.05.012 .









