Овираптор
| Овираптор Временной диапазон: поздний мел ,
~ | |
|---|---|

| |
| Голотип Овираптора | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Динозаврия |
| Клэйд : | Саурисхия |
| Клэйд : | Теропода |
| Семья: | † Овирапториды |
| Подсемейство: | † Овирапторины |
| Род: | † Овираптор Осборн , 1924 год. |
| Типовой вид | |
| † Овираптор филоцератопс Осборн, 1924 год.
| |
| Синонимы | |
| |
Овираптор ( / ˈ oʊ v ɪ r æ p t ər / ; букв. « похититель яиц » ) — род динозавров овирапторидов - , обитавший в Азии в позднемеловой период. Первые останки были собраны в формации Джадохта в Монголии в 1923 году во время палеонтологической экспедиции под руководством Роя Чепмена Эндрюса , а в следующем году род и типовой вид Oviraptor philoceratops были названы Генри Фэйрфилдом Осборном . Название рода отсылает к первоначальной мысли о привычках кражи яиц, а конкретное название было призвано подкрепить эту точку зрения, указывая на предпочтение цератопсов яйцам . Несмотря на то, что к этому роду было отнесено множество экземпляров, Овираптор известен только по одному частичному скелету, рассматриваемому как голотип , а также по гнезду, состоящему из примерно пятнадцати яиц и нескольких небольших фрагментов молоди.
Овираптор представлял собой довольно маленький пернатый овирапторид, его длина оценивалась в 1,6–2 м (5,2–6,6 футов), а вес — 33–40 кг (73–88 фунтов). У него была широкая нижняя челюсть и череп с гребнем. И верхняя, и нижняя челюсти были беззубыми и имели роговой клюв , который использовался во время кормления наряду с прочной морфологией нижних челюстей. Руки были хорошо развиты и удлинены, оканчивались тремя пальцами с изогнутыми когтями. Как и у других овирапторидов, у Овираптора были длинные задние конечности с четырьмя пальцами и редуцированным первым пальцем . Хвост, вероятно, был не очень удлиненным и заканчивался пигостилем , поддерживающим крупные перья.
Первоначальные родственные связи Oviraptor отнес их к неродственным Ornithomimidae в то время были плохо изучены, и первоначальный описатель Генри Осборн . Однако повторные исследования, проведенные Ринченом Барсболдом, доказали, что Oviraptor достаточно отличался, чтобы выделить отдельное семейство Oviraptoridae . При первом описании Овираптор интерпретировался как динозавр, похищающий яйца и поедающий яйца, учитывая тесную связь голотипа с гнездом динозавра. Однако находки многочисленных овирапторозавров в гнездовых позах показали, что этот экземпляр на самом деле высиживал гнездо, а не воровал и не питался яйцами. Более того, сообщалось об обнаружении останков маленького подростка или птенца в сочетании с образцом голотипа, что еще больше подтверждает родительскую заботу .
История открытия
[ редактировать ]
Первые останки овираптора были обнаружены на красноватых песчаниках позднемеловой джадохтинской свиты Монголии , в местонахождении Байн-Дзак (также известном как «Пылающие скалы »), в ходе Третьей среднеазиатской экспедиции в 1923 году. Эту экспедицию возглавил североамериканский натуралист Рой Чепмен Эндрюс и завершилось открытием трёх новых для науки останков теропод, в том числе овираптора . Официально они были описаны североамериканским палеонтологом Генри Фэйрфилдом Осборном в 1924 году, который на основе нового материала назвал роды Oviraptor , Saurornithoides и Velociraptor . Особый род Oviraptor был выделен с типовым видом O. philoceratops на основе голотипа AMNH 6517 , частичного индивида без задней части скелета, но с сильно раздробленным черепом , частичными шейными и спинными позвонками , грудными элементами, включая вилку с левой рука и части кистей, левая подвздошная кость и некоторые ребра . Соответственно, этот экземпляр был найден лежащим над гнездо, содержащее примерно 15 яиц (гнездо, занесенное в каталог как AMNH 6508), череп которого отделен от яиц осадком всего на 10 см (100 мм ) . Учитывая близость обоих экземпляров, Осборн интерпретировал Овираптора как динозавра с привычкой поедать яйца и объяснил, что общее название Oviraptor означает в переводе с латыни «похититель яиц» или «похититель яиц» из-за ассоциации окаменелостей. Видовое название « филоцератопс » означает «любовь к яйцам цератопсов», что также связано с первоначальной мыслью о том, что гнездо принадлежит протоцератопсу или другому цератопсу . Однако Осборн предположил, что название Овираптор могло отражать неправильное представление об этом динозавре. Более того, Осборн обнаружил, что Овираптор похож на неродственных (в то время, однако, считавшихся родственными) быстро бегающих орнитомимидов, основанных на беззубых челюстях, и отнес Овираптора к Ornithomimidae . [1] Осборн ранее сообщал об этом таксоне как «Fenestrosaurus philoceratops» . [2] но позже это было дискредитировано. [1]

В 1976 году монгольский палеонтолог Ринчен Барсболд отметил некоторые несоответствия в таксономическом расположении овираптора и пришел к выводу, что этот таксон сильно отличается от орнитомимид по анатомическим признакам. Исходя из этого, он создал Oviraptoridae , чтобы содержать Oviraptor и его близких родственников. [3] Осборном После первоначального описания Овираптора , яичное гнездо, связанное с голотипом, было признано принадлежащим Протоцератопсу . [4] [5] а овирапториды в значительной степени считались тероподами, питающимися яйцами. [6] Тем не менее, в 1990-х годах открытие многочисленных гнездящихся и птенцов овирапторидов доказало, что Осборн был прав в своих предостережениях относительно названия Овираптор . Эти результаты показали, что овирапториды высиживали и защищали свои гнезда, приседая на них. Эта новая линия доказательств показала, что гнездо, связанное с голотипом Овираптора, принадлежало ему, и особь на самом деле высиживала яйца в момент смерти, а не охотилась на них. [7] [8] [9]
Рекомендуемые образцы
[ редактировать ]
После присвоения названия Oviraptoridae в 1976 году Барсболд отнес к Oviraptor шесть дополнительных экземпляров , включая два конкретных экземпляра под номерами MPC-D 100/20 и 100/21. [3] В 1986 году Барсболд понял, что последние два не принадлежат к этому роду и вместо этого представляют собой новый овирапторид: Conchoraptor . [10] Большинство других экземпляров также вряд ли принадлежат самому овираптору , и их отнесли к другим овирапторадам. [11] [12] Частичная особь, также с яйцами, из формации Баян Мандаху в Монголии, была упомянута в 1996 году Дун Чжимингом и Филипом Дж. Карри , экземпляр IVPP V9608. [9] Однако в 2010 году Николас Р. Лонгрич и два последних палеонтолога выразили неуверенность в отношении этого направления, поскольку существует несколько анатомических различий, таких как пропорции фаланг рук. Они пришли к выводу, что этот экземпляр представляет собой другой неопределенный вид, не относящийся к этому таксону. [13] В 1981 году Барсболд отнес экземпляр MPC-D 100/42 к Овираптору , очень хорошо сохранившейся и довольно полной особи из джадохтинской свиты. [14] Поскольку известные элементы Oviraptor были настолько фрагментарными по сравнению с другими представителями, MPC-D 100/42 стал основным эталоном/изображением этого таксона, который широко известен как Oviraptor philoceratops в научной литературе . [15] [11]

Эта концепция была опровергнута Джеймсом М. Кларком и его коллегами в 2002 году, которые отметили, что этот экземпляр с высоким гребнем имеет больше общих черт черепа с Ситипати , чем с Овираптором , который фактически не сохраняет гребень. может представлять собой второй вид первого или целый новый род. [16] В 1986 году Барсболд описал второй вид овираптора , «O. mongoliensis» , на основе экземпляра MPC-D 100/32a, происходящего из формации Немегт . [10] Однако повторное исследование, проведенное Барсболдом в 1997 году, обнаружило достаточно различий в этом экземпляре, чтобы назвать новый род Rinchenia , но он не описал его формально, и этот новый овирапторид остался как nomen dubium . [17] и его команда внесли поправки, В 2004 году польский палеонтолог Хальшка Осмолска официально назвав таксон Rinchenia mongoliensis . [18] Североамериканский и его коллеги в 2018 году сообщили о палеонтолог Марк А. Норелл новом экземпляре овираптора : AMNH 33092, который состоит из большеберцовой кости и двух плюсневых костей птенца или очень маленькой молоди. AMNH 33092 был обнаружен вместе с голотипом и, вероятно, был частью гнезда. Овираптор теперь известен по голотипу с соответствующими яйцами и молоди/птенцу. [19]
Описание
[ редактировать ]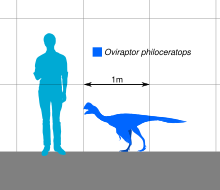
Длина голотипа оценивается в 1,6–2 м (5,2–6,6 футов) и вес от 33 до 40 кг (от 73 до 88 фунтов). [20] [21] [22] Хотя у голотипа в основном отсутствует задняя часть скелета, вполне вероятно, что у Овираптора были две хорошо развитые задние конечности, которые заканчивались тремя функциональными пальцами , первый из которых был рудиментарным , а также относительно уменьшенный хвост . Как видно у родственных овирапторидов , руки были покрыты удлиненными перьями , а хвост заканчивался пигостилем , который, как известно, поддерживает веер из перьев. [23] [24]
Череп
[ редактировать ]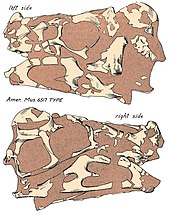
Череп отверстиями Овираптора отверстиями) по сравнению был глубоким и укороченным, с большими ( с другими динозаврами, и в сохранившемся виде его длина составляла около 17,9 см (179 мм). Однако фактическая длина может быть больше, учитывая, что в черепе голотипа отсутствуют несколько областей, таких как предчелюстная кость. Череп-голотип почти полностью лишен гребня, однако верхние поверхности сросшихся теменных и лобных костей указывают на то, что он, вероятно, имел хорошо развитый гребень, поддерживаемый носовой и предчелюстной костями (преимущественно последней) головотрубки . У овираптора была удлиненная верхняя челюсть и зубная кость, что могло привести к более вытянутой морде по сравнению с очень коренастыми челюстями других овирапторидов. Небо сошниками жесткое, расширено ниже линии челюсти и образовано предчелюстными костями, и верхнечелюстными костями . Как и у других овирапторидов, он мог иметь пару зубообразных выступов на нёбе, направленных вниз. Как и у других овирапторидов, ноздри (наружные ноздри) были относительно небольшими и располагались высоко на черепе. У овираптора были беззубые челюсти, которые заканчивались крепким, попугая похожим на , рамфотекой (роговым клювом). Кривизна кончика зубной кости была загнута вниз, но менее выражена, чем у других овирапторидов, таких как Citipati . В целом нижняя челюсть представляет собой короткую и глубокую кость, занимающую 19,5 см (195 мм). [16]
Посткраниальный скелет
[ редактировать ]
голотипа Как и у большинства овирапторид, нервные отростки шейных позвонков были короткими, а нервные дуги имели Х-образную форму. Однако шипы становятся более выраженными в задних позвонках. Зигапофизы первых шейных позвонков расположены параллельно друг другу, а постзигапофизы, по-видимому, существенно не отклоняются от средней линии, в основном сходны с Citipati . Шейные ребра у голотипа срослись с позвонками. [25] [19] Нервные отростки имеют прямоугольную форму в переднем ряду спинных позвонков , если смотреть сбоку, и крупнее, чем отростки шейных позвонков. На переднем спинном позвонке можно найти несколько плевроцелей (небольших воздушных отверстий), похожих на таковые у Хаана . [25]
Вилка поверхности гипоклеидия — направленного вниз овираптора сильно отличается от других овирапторидов наличием срединного киля на передней выступа в центре вилки. Эта кость имеет V-образную форму, округлую в поперечном сечении, сохраняет удлиненный шиповидный гипоклеидий, межключичный угол составляет около 90°. [26] У голотипа скапулокоракоид коракоид слит, однако сильно поврежден. Лопатка . слегка изогнута, ее длина составляет 23 см (230 мм) Овираптор имел относительно удлиненную руку, состоящую из плечевой , лучевой , локтевой и кисти рук . [1] [25]

Формула фаланг овираптора была 2-3-4, как и у большинства других теропод и овирапторидов. На руке Овираптора было три тонких птичьих пальца, каждый из которых оканчивался сплющенными из стороны в сторону и загнутыми когтями ( когтевая кость). В отличие от некоторых овирапторидов, у Овираптора не наблюдалось редукции второго и третьего пальцев относительно первого. [1] [25] [27] У рассматриваемого молодого Oviraptor AMNH 33092 сохраняется материал задних конечностей, включающий правую большеберцовую кость с III и IV плюсневыми костями. Длина его голени составляет 5,87 см (58,7 мм), что указывает на то, что особь значительно меньшего размера, чем голотип. [19] В гнезде AMNH 6508 сохранились яйца элонгатоолитид , длина каждого яйца составляет 14 см (140 мм) (некоторые из них неполные). Тем не менее, существует вероятность того, что тафономическое дробление могло сжать их до 2 см (20 мм). [7] [28]
Классификация
[ редактировать ]Овираптор с орнитомимидами изначально был связан Осборном из-за его беззубого клюва. Осборн также обнаружил сходство с Chirostenotes , который до сих пор считается близким родственником овираптора . [1] В 1976 году Барсболд включающее Oviraptor и его близких родственников, сделав Oviraptor типовым родом Oviraptoridae создал новое семейство , . [3] Во время переописания голотипа черепа в 2002 году Кларком и его коллегами они отметили, что у Овираптора были относительно удлиненные верхняя челюсть и зубная кость. Эти черты менее выражены у производных овирапторид и позволяют предположить, что Овираптор принадлежит к ближнему основанию Овирапторид. [16]
соответствует Приведенная ниже кладограмма анализу, проведенному Грегори Ф. Фанстоном и его коллегами в 2020 году: [29]
| Овирапториды |
| ||||||
Палеобиология
[ редактировать ]Кормление
[ редактировать ]
впервые был описан в 1924 году Осборном, Когда Овираптор первоначально предполагалось, что он был яйцеядным — организмом, питающимся яйцами, — на основании связи голотипа с гнездом, которое, как считалось, принадлежало протоцератопсу . [1] В 1977 году Барсболд предложил гипотезу сокрушающей челюсти. Он утверждал, что сила мощных нижних челюстей и, вероятно, рамфотеки (рогового клюва) была достаточно сильной, чтобы сломать раковины моллюсков, таких как моллюски , которые встречаются в той же геологической формации, что и овираптор . Эти кости составляют часть основной кости верхней челюсти или верхней челюсти , которые сходятся посередине, образуя пару зубцов. Рамфотека и нижние челюсти вместе с выступами нескольких костей неба представляли собой инструмент для прокалывания. Барсболд также предположил, что овирапториды могли вести полуводный образ жизни, основанный на питании моллюсками, высоком расположении носовых полостей, увеличенной мускулатуре хвоста и большем размере первого пальца руки . [30] В тезисах конференции 1990 года Дэвид К. Смит представил остеологическую переоценку овираптора , отвергнув утверждения Барсболда. Он не нашел никаких доказательств того, что передние конечности специализируются на передвижении в воде , а челюсти, вместо того, чтобы сохранять дробящий механизм, сохраняют режущие поверхности. Поскольку череп беззуб , легко сложен и не имеет нескольких сильных мест прикрепления мышц, Смит предположил, что листья могли быть важной частью рациона овираптора . [31] Однако в 1995 году Норелл и его коллеги сообщили о фрагментированных останках ящерицы в полости тела экземпляра-голотипа, что позволяет предположить, что Овираптор был частично плотоядным . [32]

В 2008 году Стиг Олав К. Янсен сравнил череп нескольких видов овирапторидов с черепами птиц и черепах , чтобы выяснить, какие свойства могут предсказать появление рамфотеки. Он обнаружил, что нижние челюсти овирапторидов очень похожи на челюсти попугаев , а верхние челюсти больше похожи на челюсти черепах. На основании этих наблюдений Янсен предположил, что овирапториды были всеядны , поскольку резко развитая рамфотека вместе с выступающими передними конечностями были приспособлены для ловли и разрывания мелкой добычи . Более того, заостренные выступы нёба способствовали удержанию добычи. Янсен отметил, что полностью травоядная диета овирапторидов кажется маловероятной, поскольку у них не было плоских и широких томий (режущих краев нижних челюстей) для жевания , и они не могли двигать нижними челюстями в стороны. Однако он считал нижние челюсти достаточно сильными, чтобы раздробить хотя бы такие элементы, как яйца, орехи или другие твердые семена . [33]
Лонгрич и его коллеги в 2010 году также отвергли гипотезу дурофагов (животных, которые практикуют дробление панциря), учитывая, что у таких животных обычно развиваются зубы с широкой поверхностью дробления. Заостренная форма зубных костей нижних челюстей позволяет предположить, что у овирапторидов вместо этого была рамфотека с острыми краями, которая использовалась для разрезания пищи. Симфизарная область (сращение костей) в передней части зубной кости, возможно, давала некоторую способность к дроблению, но , поскольку это была относительно небольшая область, это, вероятно, не было основной функцией челюстей. Еще одним аргументом против того, что они питались моллюсками, является тот факт, что большинство овирапторидов были обнаружены в отложениях , которые, как интерпретируется, представляют собой в основном засушливые или полузасушливые среды, такие как овираптор в формации Джадохта . Команда также обнаружила, что овирапториды и дицинодонты имеют общие черепные особенности, такие как короткие, глубокие и беззубые челюсти; удлиненные зубные симфизы; удлиненные нижнечелюстные отверстия; и острое небо. К современным животным с челюстями, напоминающими челюсти овирапторидов, относятся попугаи и черепахи ; последняя группа имеет также зубчатые выступы на небной области. Лонгрич и его коллеги пришли к выводу, что из-за сходства овирапторидов с травоядными животными большая часть их рациона, скорее всего, состояла из растительного материала. Челюсти овирапторидов, возможно, были специализированы для переработки пищи, такой как ксерофитная растительность — растительность, приспособленная к среде с небольшим количеством воды, — которая росла бы в их засушливой среде, но это невозможно продемонстрировать, поскольку о ней мало что известно. Палеофлора пустыни Гоби . [13]
Однако в 2018 году Фанстон и его коллеги поддержали гипотезу сокрушающей челюсти. Они отметили, что коренастый рострум и крепкие нижние челюсти овирапторидов на самом деле предполагают сильный и щипающий укус, очень похожий на укус попугаев. Фанстон и его коллеги считали, что эти анатомические особенности овирапторидов соответствуют плодоядной диете, включающей орехи и семена. [34]
Воспроизведение
[ редактировать ]

После описания эмбрионального экземпляра Citipati в 1994 году овирапториды стали более понятными: вместо того, чтобы питаться яйцами, они фактически высиживали гнезда и ухаживали за ними . Этот образец показал, что голотип Овираптора, вероятно, был половозрелой особью, которая погибла, насиживая связанное гнездо с яйцами . [7] Это новое поведение овирапторидов стало более ясным после отчета и краткого описания гнездящегося взрослого экземпляра Ситипати в 1995 году Норелла и его коллег. Особь была найдена на кладке яиц , ее задние конечности симметрично присели по обе стороны гнезда, а передние конечности закрывали периметр гнезда. Эта задумчивая поза сегодня встречается только у современных птичьих динозавров и подтверждает поведенческую связь между последней группой и нептичьими динозаврами . [8] В 1996 году Донг и Карри описали новый гнездящийся экземпляр овирапторида из формации Баян-Мандаху . Он был найден лежащим на вершине гнезда, состоящего примерно из 6 сохранившихся яиц, отложенных в виде насыпи с круглым узором. Поскольку экземпляр был найден над гнездом с передними конечностями, закрывающими яйца, и частично сохранившейся задней лапой недалеко от центра гнезда, Донг и Карри предположили, что он был пойман и похоронен песчаной бурей во время инкубации. Они исключили возможность того, что овирапториды похитили яйца, поскольку они либо поглотили бы гнездо, либо инстинктивно покинули бы его задолго до того, как оно было погребено песчаной бурей или другим метеорологическим явлением . [9]
В 1999 году Кларк и его команда подробно описали ранее сообщенный образец гнездования Ситипати и кратко обсудили образец голотипа Овираптора и его связь с гнездом AMNH 6508. Они отметили, что точное положение, в котором голотип был найден над гнездом, неясно, поскольку они были разделены во время приготовления , и гнездо, по-видимому, не полностью укомплектовано: сохранилось около 15 яиц, два из которых были повреждены. Причем полукруглое расположение гнезда указывает на то, что яйца откладывались парами и не менее чем в три кольца, причем первоначально это гнездо было круглым, похожим на холмик . [28]
Томас П. Хопп и Марк Дж. Орсен в 2004 году проанализировали насиживающее поведение вымерших и современных видов динозавров, включая овирапторидов, чтобы оценить причину удлинения и развития крыльев и рулевых перьев . Учитывая, что наиболее полный гнездовой экземпляр овирапторидов — на тот момент гнездовой экземпляр Citipati 1995 года — был найден в позе, очень напоминающей птичью: передние конечности были почти согнуты, а грудная область, живот и ступни соприкасались с яйца, Хопп и Орсен указали, что длинные перистые перья и перьевой покров, скорее всего, присутствовали при жизни. «Крылья» и хвост овирапторидов обеспечивали защиту яиц и птенцов от климатических факторов, таких как солнечный свет , ветер и осадки . Однако руки этого экземпляра не были сильно сложены, как у некоторых современных птиц, а более вытянуты, напоминая стиль крупных нелетающих птиц , таких как страус . Вытянутое положение руки также похоже на насиживающее поведение этой птицы, которая, как известно, гнездится большими кладками, как овирапториды. Основываясь на положении передних конечностей гнездящихся овирапторид, Хопп и Орсен предположили, что высиживание является древней причиной удлинения крыльев и хвостовых перьев, поскольку существовала большая потребность в обеспечении оптимального защита яиц и молоди. [35]

В 2005 году Тамаки Сато и его команда сообщили о необычном образце овирапторида из формации Наньсюн . Было обнаружено, что у этого нового экземпляра сохранилась в основном тазовая область с двумя яйцами внутри, что указывает на самку . Размер и положение яиц позволяют предположить, что овирапториды сохранили два функциональных яйцевода , но уменьшили количество овулировавших яиц до одного на яйцевод. [36] Дэвид Дж. Варриккио и его коллеги в 2008 году обнаружили, что относительно большой размер яичной кладки овирапторидов и троодонтид наиболее похож на таковой у современных птиц, которые практикуют полигамное самцов спаривание и обширную родительскую заботу , таких как бескилевые птицы, что предполагает схожие привычки. Эта репродуктивная система, скорее всего, представляет собой наследственное состояние современных птиц, а забота обоих родителей (в которой участвуют оба родителя) является более поздним развитием. [37] В 2014 году У. Скотт Персонс и его коллеги предположили, что овирапторозавры были вторично нелетающими, и некоторые черты их хвостов могут указывать на склонность к демонстративному поведению , например, к демонстрации ухаживания . Хвост некоторых овирапторозавров и овирапторидов заканчивался пигостилем — костной структурой на конце хвоста, которая, по крайней мере у современных птиц, используется для поддержки веера из перьев. Кроме того, хвост был особенно мускулистым и обладал выраженной гибкостью, что, возможно, помогало в движениях ухаживания. [24]

В 2018 году Цзы-Руэй Ян и его коллеги определили слои кутикулы на нескольких яичных скорлупах динозавров-манирапторов, в том числе овирапторидов. Эти особые слои состоят из белков , полисахаридов и пигментов , но главным образом из липидов и гидроксиапатита . У современных птиц они служат для защиты яиц от обезвоживания и проникновения микроорганизмов . Поскольку большинство экземпляров овирапторидов было обнаружено в отложениях на основе калише , Ян и его коллеги предположили, что яйца, покрытые кутикулой, могли быть репродуктивной стратегией, адаптированной для повышения успеха их вылупления в таком засушливом климате и окружающей среде. [38]
В 2019 году Ян и его коллеги переоценили гипотезу терморегуляционной контактной инкубации с использованием полных гнезд овирапторидов из формации Наньсюн и представили подробную реконструкцию архитектуры кладки овирапторид. Они отметили, что взрослые особи овирапторидов, находящиеся в гнезде, не обязательно насиживали яйца, поскольку могли представлять собой самку в процессе откладки яиц, а многокольцевая кладка препятствовала достаточной передаче тепла от родителя к внутренним кольцам яиц. Обычное гнездо овирапторидов представляло собой пологий холмик с высокоорганизованной архитектурой: яйца, вероятно, были пигментированными и располагались парами, причем каждая пара располагалась в трех-четырех эллиптических кольцах. Поскольку родитель, вероятно, действовал из центра гнезда, в этом регионе яиц не было. Ян и его коллеги пришли к выводу, что стиль гнездования овирапторидов настолько уникален, что ему не хватает современных аналогов, поэтому использование размножения овирапторидов может быть не лучшим примером для информирования об эволюции репродуктивных стратегий птиц. Однако команда не смогла определить, был ли несовершеннолетний Овираптор AMNH 33092 вылупился из гнезда, связанного с голотипом. [39]
Палеосреда
[ редактировать ]
Овираптор известен из местонахождения Байн-Дзак формации Джадохта в Монголии, формации, которая восходит к позднему мелу примерно 71–75 миллионов лет назад . [40] Палеосреда джадохтинской свиты интерпретируется как полузасушливый климат с песчаными дюнами и аллювиальными условиями, похожими на современную пустыню Гоби . Полузасушливый степной ландшафт был осушён прерывистыми ручьями и иногда подвергался воздействию пыльных и песчаных бурь , а влажность носила сезонный характер. [41] [42] нескольких недолговечных водоемах Хотя в основном считается, что это образование сохранило крайне засушливую среду, в районе Ухаа Толгод сообщалось о , основанных на речных отложениях. [43] Более того, считается, что позже, в кампанскую эпоху и в маастрихтский период , климат сместится в сторону более влажной речной среды, наблюдаемой в формации Немегт . [42]
Джадохтинская свита разделена на нижнюю байн-дзакскую и верхнюю тургругынскую пачки. Известные останки Овираптора были произведены членом Байн Дзак, который также дал динозавров Bainoceratops , Pinacosaurus , Protoceratops , Saurornithoides , Velociraptor , [40] и Хальшкараптор . [44] Дальнейшая фауна динозавров из этого члена включает фауну из местонахождения Ухаа Толгод, состоящую из Апсарависа , Байронозавра , Ситипати , Гобиптерикса , Хана , Хола , Шуувуйи , Цаагана , [43] и Минотавразавр . [45]
Тафономия
[ редактировать ]Поза голотипа Овираптора , а также наличие яиц позволяют предположить, что он был пойман в ловушку над гнездом во время песчаной бури , и захоронение было относительно быстрым, учитывая, что тело не имело возможности полностью разъединиться или быть съеденным хищниками. [1] [9] Палеонтолог Кеннет Карпентер также согласился с тем, что песчаные бури, возможно, были наиболее вероятным событием того, что яйца, найденные в отложениях, были захоронены. [46] Среди элементов череп стал особенно уплощенным и деформированным в процессе окаменения . [16]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с д и ж г Осборн, HF (1924). «Три новых теропода, зона протоцератопса, центральная Монголия». Новитаты Американского музея (144): 1–12. HDL : 2246/3223 . OCLC 40272928 .
- ^ Осборн, HF (1924). «Открытие неизвестного континента». Естественная история . 24 (2): 133–149.
- ^ Jump up to: а б с Barsbold, R. (1976). "О новом позднемеловом семействе мелких теропод Oviraptoridae fam. nov Монголии" [A new Late Cretaceous family of small theropods Oviraptoridae n. fam. in Mongolia]. Doklady Akademii Nauk SSSR . 226 (3): 685−688.
- ^ Кумбс, WP (1989). «Современные аналоги гнездования динозавров и родительского поведения». В Фарлоу, Джо (ред.). Палеобиология динозавров . Специальные статьи Геологического общества Америки. Том. Специальный доклад Геологического общества Америки 238. Колорадо: Боулдер. стр. 21−54. дои : 10.1130/SPE238-p21 . ISBN 0-8137-2238-1 .
- ^ Талборн, РА (1992). «Гнездо динозавра Протоцератопса» . Летайя . 25 (2): 145–149. Бибкод : 1992Лета..25..145Т . дои : 10.1111/j.1502-3931.1992.tb01379.x .
- ^ Карри, Пи Джей; Годфри, SJ; Несов, Л.А. (1993). «Новые экземпляры ценугнатид (Dinosauria: Theropoda) из верхнего мела Северной Америки и Азии». Канадский журнал наук о Земле . 30 (10): 2255–2272. Бибкод : 1993CaJES..30.2255C . дои : 10.1139/e93-196 .
- ^ Jump up to: а б с Норелл, Массачусетс; Кларк, Дж. М.; Дашевег, Д.; Барсболд, Р.; Кьяппе, LM; Дэвидсон, Арканзас; Маккенна, MC; Альтангерель, П.; Новачек, MJ (1994). «Эмбрион тероподного динозавра и сходство яиц динозавра из Пылающих скал» . Наука . 266 (5186): 779–782. Бибкод : 1994Sci...266..779N . дои : 10.1126/science.266.5186.779 . JSTOR 2885545 . ПМИД 17730398 . S2CID 22333224 .
- ^ Jump up to: а б Норелл, Массачусетс; Кларк, Дж. М.; Кьяппе, LM; Дашевег, Д. (1995). «Гнездящийся динозавр» . Природа . 378 (6559): 774–776. Бибкод : 1995Natur.378..774N . дои : 10.1038/378774a0 . S2CID 4245228 .
- ^ Jump up to: а б с д Донг, З.; Карри, Пи Джей (1996). «Об обнаружении скелета овираптора на яичном гнезде в Баян Мандаху, Внутренняя Монголия, Китайская Народная Республика». Канадский журнал наук о Земле . 33 (4): 631–636. Бибкод : 1996CaJES..33..631D . дои : 10.1139/e96-046 .
- ^ Jump up to: а б Barsbold, R. (1986). "Хищные динозавры овирапторы" [Carnivorous dinosaurs oviraptors]. In Vorobyeva, E. I. (ed.). Herpetological research in the Mongolian People's Republic (in Russian). Institute of Evolutionary Morphology and Ecology of Animals: Doklady Akademii Nauk SSSR. pp. 210–223.
- ^ Jump up to: а б Марьянская, Т.; Осмольская, Х.; Вулсан, М. (2002). «Авиаланский статус овирапторозаврии» (PDF) . Acta Palaeontologica Polonica . 47 (1): 97−116.
- ^ Кундрат, М.; Яначек, Дж. (2007). «Краниальная пневматизация и слуховые восприятия овирапторидного динозавра Conchoraptor gracilis (Theropoda, Maniraptora) из позднего мела Монголии» . Naturwissenschaften . 94 (9): 769–778. Бибкод : 2007NW.....94..769R . дои : 10.1007/s00114-007-0258-7 . ПМИД 17530209 . S2CID 40622976 .
- ^ Jump up to: а б Лонгрич, Северная Каролина; Карри, Пи Джей; Донг, З. (2010). «Новый овирапторид (Dinosauria: Theropoda) из верхнего мела Баян Мандаху, Внутренняя Монголия» . Палеонтология . 53 (5): 945–960. Бибкод : 2010Palgy..53..945L . дои : 10.1111/j.1475-4983.2010.00968.x .
- ^ Barsbold, R. (1981). "Беззубые хищные динозавры Монголии" [Toothless carnivorous dinosaurs of Mongolia]. Transactions, Joint Soviet–Mongolian Palaeontological Expedition (in Russian). 15 : 28−39. S2CID 195060070 . Translated paper
- ^ Барсболд, Р.; Марьянска, Т.; Осмольска, Х. (1990). «Овирапторозаврия». В Вейшампеле, Д.Б.; Осмольская, Х.; Додсон, П. (ред.). Динозаврия (1-е изд.). Беркли: Издательство Калифорнийского университета. стр. 249−258. ISBN 9780520067271 .
- ^ Jump up to: а б с д Кларк, Дж. М.; Норелл, Массачусетс; Роу, Т. (2002). «Краниальная анатомия Citipati osmolskae (Theropoda, Oviraptorosauria) и новая интерпретация голотипа Oviraptor philoceratops» (PDF) . Новитаты Американского музея (3364 г.): 1–24. doi : 10.1206/0003-0082(2002)364<0001:CAOCOT>2.0.CO;2 . hdl : 2246/2853 . S2CID 52247789 .
- ^ Барсболд, 1997. Овирапторозаврия. В Карри и Падиане (ред.). Энциклопедия динозавров. 505-509.
- ^ Осмольска, Х.; Карри, Пи Джей; Барсболд, Р. (2004). «Овирапторозаврия» В Вейшампеле, DB; Додсон, П.; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Издательство Калифорнийского университета. стр. 100-1 165−1 ISBN 0520242092 .
- ^ Jump up to: а б с Норелл, Массачусетс; Баланов, А.М.; Барта, Делавэр; Эриксон, генеральный директор (2018). «Второй экземпляр Citipati osmolskae, связанный с гнездом яиц из Ухаа Толгод, Омноговский аймак, Монголия» . Новитаты Американского музея (3899 г.): 1–44. дои : 10.1206/3899.1 . hdl : 2246/6858 . S2CID 53057001 .
- ^ Вернер, Дж.; Грибелер, Э.М. (2013). «Новый взгляд на размножение нептичьих динозавров и их эволюционные и экологические последствия: связь ископаемых свидетельств с аллометрией современных близких родственников» . ПЛОС ОДИН . 8 (8): е72862. Бибкод : 2013PLoSO...872862W . дои : 10.1371/journal.pone.0072862 . ПМК 3749170 . ПМИД 23991160 .
- ^ Пол, GS (2016). Принстонский полевой справочник по динозаврам (2-е изд.). Принстон, Нью-Джерси: Издательство Принстонского университета. п. 178. ИСБН 9780691167664 .
- ^ Кампионе, Невада; Эванс, округ Колумбия (2020). «Точность и точность оценки массы тела нептичьих динозавров» . Биологические обзоры . 95 (6): 1759–1797. дои : 10.1111/brv.12638 . ПМИД 32869488 . S2CID 221404013 .
- ^ Фанти, Ф.; Карри, Пи Джей; Бадамгарав, Д.; Лалуэса-Фокс, К. (2012). «Новые экземпляры немегтомайи из баруунгойотской и нэмегтинской свит (поздний мел) Монголии » ПЛОС ОДИН 7 (2): e31330. Бибкод : 2012PLoSO... 731330F дои : 10.1371/journal.pone.0031330 . ПМЦ 3275628 . ПМИД 22347465 .
- ^ Jump up to: а б Лица, WS; Карри, Пи Джей; Норелл, Массачусетс (2014). «Формы и функции хвоста овирапторозавра» . Acta Palaeontologica Polonica . дои : 10.4202/app.2012.0093 .
- ^ Jump up to: а б с д Баланов, А.М.; Норелл, Массачусетс (2012). «Остеология Хана Маккеннаи (Овирапторозаврия, Теропода)» . Бюллетень Американского музея естественной истории . 372 (372): 1–77. дои : 10.1206/803.1 . hdl : 2246/6413 . OCLC 884738416 . S2CID 86067306 .
- ^ Несбитт, С.Дж.; Тернер, А.Х.; Сполдинг, М.; Конрад, Дж.Л.; Норелл, Массачусетс (2009). «Вилочка теропода» . Журнал морфологии . 270 (7): 856–879. дои : 10.1002/jmor.10724 . ПМИД 19206153 .
- ^ Лю, Дж.; Карри, Пи Джей; Сюй, Л.; Чжан, X.; Пу, Х.; Цзя, С. (2013). «Динозавры-овирапториды размером с курицу из центрального Китая и их онтогенетические последствия» (PDF) . Naturwissenschaften . 100 (2): 165−175. Бибкод : 2013NW....100..165L . дои : 10.1007/s00114-012-1007-0 . ПМИД 23314810 . S2CID 206871470 .
- ^ Jump up to: а б Кларк, Дж. М.; Норелл, Массачусетс; Кьяппе, LM (1999). «Скелет овираптора из позднего мела из Ухаа Толгода, Монголия, сохранившийся в птичьем высиживающем положении над гнездом овирапторидов» . Новитаты Американского музея (3265 г.): 1–36. hdl : 2246/3102 .
- ^ Фанстон, Г.Ф.; Цогтбаатар, К.; Цогтбаатар, К.; Кобаяши, Ю.; Салливан, К.; Карри, Пи Джей (2020). «Новый двупалый динозавр проливает свет на происхождение овирапторозаврии» . Королевское общество открытой науки . 7 (10): 201184. Цифровой код : 2020RSOS....701184F . дои : 10.1098/rsos.201184 . ПМЦ 7657903 . ПМИД 33204472 .
- ^ Барсболд, Р. (1977). «Кинетизм и особенности челюстного аппарата овирапторов (Theropoda, Saurischia)». Советско-Монгольская палеонтологическая экспедиция, Труды . 4 : 37−47. Переведенная статья
- ^ Смит, ДК (1990). «Остеология Oviraptor philoceratops, возможного травоядного теропода из верхнего мела Монголии». Журнал палеонтологии позвоночных . 10 (доп. 003): 42А. дои : 10.1080/02724634.1990.10011841 .
- ^ Норелл, Массачусетс; Гаффни, ES; Дингус, Л. (1995). Обнаружение динозавров в Американском музее естественной истории . Кнопф Инк. 225.
- ^ Янсен, СОК (2008). Морфология клюва овирапторидов на основе современных птиц и черепах (Диссертация). Университет Осло. стр. 1−48. hdl : 10852/11785 .
- ^ Фанстон, Г.Ф.; Мендонка, ЮВ; Карри, Пи Джей; Барсболд, Р.; Барсболд, Р. (2018). «Анатомия, разнообразие и экология овирапторозавров в бассейне Немегта» . Палеогеография, Палеоклиматология, Палеоэкология . 494 : 101–120. Бибкод : 2018PPP...494..101F . дои : 10.1016/j.palaeo.2017.10.023 .
- ^ Хопп, ТП; Орсен, MJ (2004). «Поведение динозавров и происхождение маховых перьев» (PDF) . В Карри, Пи Джей; Коппельхус, Е.Б.; Шугар, Массачусетс; Райт, Дж.Л. (ред.). Пернатые драконы: исследования перехода от динозавров к птицам . Блумингтон: Издательство Университета Индианы. стр. 234−250.
- ^ Сато, Т.; Ченг, Ю.-Н.; У, Х.-С.; Зеленицкий, ДК; Сяо, Ю.-Ф. (2005). «Пара яиц в скорлупе внутри самки динозавра» (PDF) . Наука . 308 (5720): 375. doi : 10.1126/science.1110578 . ПМИД 15831749 . S2CID 19470371 .
- ^ Варриккио, диджей; Мур, младший; Эриксон, генеральный директор; Норелл, Массачусетс; Джексон, Флорида; Борковски, Джей Джей (2008). «Птичья отцовская забота имела происхождение от динозавров» . Наука 322 (5909): 1826–1828 гг. Бибкод : 2008Науч... 322.1826В дои : 10.1126/science.1163245 . ПМИД 19095938 .
- ^ Ян, Т.-Р.; Чен, Ю.-Х.; Виманн, Дж.; Спиринг, Б.; Сандер, премьер-министр (2018). «Кутикула ископаемой яичной скорлупы объясняет экологию гнездования динозавров» . ПерДж . 6 : е5144. дои : 10.7717/peerj.5144 . ПМК 6037156 . ПМИД 30002976 .
- ^ Ян, Т.-Р.; Виманн, Дж.; Сюй, Л.; Ченг, Ю.-Н.; У, Х.-С.; Сандер, премьер-министр (2019). «Реконструкция кладок овирапторидов проливает свет на их уникальную биологию гнездования» . Acta Palaeontologica Polonica . 466 : 581–596. дои : 10.4202/app.00497.2018 .
- ^ Jump up to: а б Дашевег, Д.; Дингус, Л.; Петля, Д.Б.; Свишер III, CC; Дулам, Т.; Суини, MR (2005). «Новое стратиграфическое подразделение, условия осадконакопления и оценка возраста верхнемеловой джадохтинской свиты, южный бассейн Улан-Нура, Монголия» (PDF) . Новитаты Американского музея (3498): 1–31. doi : 10.1206/0003-0082(2005)498[0001:NSSDEA]2.0.CO;2 . hdl : 2246/5667 . S2CID 55836458 .
- ^ Ежикевич, Т. (1997). «Джадохтинская свита». В Карри, Пи Джей; Падиан, К. (ред.). Энциклопедия динозавров . Сан-Диего: Академическая пресса. стр. 188 −191. ISBN 978-0-12-226810-6 .
- ^ Jump up to: а б Чинзориг, Т.; Кобаяши, Ю.; Цогтбаатар, К.; Карри, Пи Джей; Ватабе, М.; Барсболд, Р. (2017). «Первые орнитомимиды (Theropoda, Ornithomimosauria) из верхнемеловой джадохтинской свиты Тёгрогийн Ширэ, Монголия» . Научные отчеты 7 (5835): 5835. Бибкод : 2017NatSR...7.5835C . дои : 10.1038/ s41598-017-05272-6 ПМЦ 5517598 . ПМИД 28724887 .
- ^ Jump up to: а б Дингус, Л.; Петля, БД; Дашевег, Д.; Свишер III, CC; Минджин, К.; Новачек, MJ; Норелл, Массачусетс (2008). «Геология Ухаа Толгода (джадохтинская свита, верхний мел, бассейн Немегта, Монголия)» (PDF ) Новитаты Американского музея (3616 г.): 1–40. дои : 10.1206/442.1 . hdl : 2246/5916 . S2CID 129735494 . [ постоянная мертвая ссылка ]
- ^ Кау, А.; Бейранд, В.; Воетен, DFAE; Фернандес, В.; Таффоро, П.; Штейн, К.; Барсболд, Р.; Цогтбаатар, К.; Карри, Пи Джей; Годфруа, П. (2017). «Синхротронное сканирование выявило экоморфологию земноводных в новой кладе птицеподобных динозавров» . Природа . 552 (7685): 395–399. Бибкод : 2017Natur.552..395C . дои : 10.1038/nature24679 . ПМИД 29211712 . S2CID 4471941 .
- ^ Алиса, Дж.; Лоуэн, М. (2013). «Новый материал минотавразавра из формации Джодокта устанавливает новые таксономические и стратиграфические критерии для этого таксона» . Журнал палеонтологии позвоночных . Программа и тезисы: 76. Архивировано из оригинала 01 ноября 2020 г. Проверено 19 ноября 2020 г.
- ^ Карпенер, К. (1999). «Яйца как окаменелости». Яйца, гнезда и детеныши динозавров: взгляд на размножение динозавров (жизнь прошлого) . Блумингтон: Издательство Университета Индианы. п. 111. ИСБН 9780253334978 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Овираптором, на Викискладе?
СМИ, связанные с Овираптором, на Викискладе?  Данные, связанные с Овираптором , в Wikispecies
Данные, связанные с Овираптором , в Wikispecies - Гнездо овираптора AMNH 6508, фотографии в AMNH
- овираптора Фотография голотипа черепа в AMNH

