Дицинодентия
| Дицинодентия | |
|---|---|

| |
| Скелет Дииктодона | |

| |
| Скелет Пласериаса | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Клэйд : | Сторона синапса |
| Клэйд : | Терапсида |
| Подотряд: | † Аномодонтия |
| Клэйд : | † Чайнозаврия |
| Клэйд : | † Дицинодонтия Оуэн , 1859 г. |
| Клады и роды | |
|
см. « Таксономия ». | |
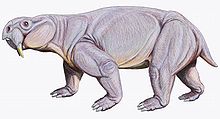
Dicynodontia — вымершая клада аномодонтов не , вымерший тип терапсидов, относящихся к млекопитающим . Дицинодонты были травоядными животными , которые обычно имели пару клыков, отсюда и их название, что означает «два собачьих зуба». Члены группы обладали роговым, обычно беззубым клювом, уникальным среди всех синапсидов . Дицинодонты впервые появились в Южной Пангее в середине перми , ок. 270–260 миллионов лет назад и стали широко распространены по всему миру и стали доминирующими травоядными животными в поздней перми , ок. 260–252 млн лет назад. Они были опустошены пермским вымиранием , которое уничтожило большинство других терапсидов ок. 252 млн лет назад. Они восстановились во время триаса , но вымерли к концу этого периода. Они были наиболее успешными и разнообразными из терапсидов, не относящихся к млекопитающим: известно более 70 родов размером с крысу , от нор размером со слона до браузеров .
Характеристики
[ редактировать ]

дицинодонтов Череп узкоспециализирован, легкий, но прочный, с синапсидными височными отверстиями в задней части черепа, значительно увеличенными для размещения более крупных челюстных мышц. Передняя часть черепа и нижняя челюсть обычно узкие и у всех, за исключением некоторых примитивных форм, беззубые. Вместо этого передняя часть рта оснащена роговым клювом, как у черепах и цератопсовых динозавров . Пища обрабатывалась за счет втягивания нижней челюсти при закрытии рта, производя мощное режущее действие. [ 2 ] что позволило бы дицинодонтам справляться с твердым растительным материалом. Дицинодонты обычно имели пару увеличенных клыкообразных зубов на верхней челюсти, аналогичных бивням, присутствующим у некоторых современных млекопитающих. У самых ранних родов это были просто увеличенные зубы, но в более поздних формах они несколько раз самостоятельно эволюционировали в постоянно растущие зубы, подобные бивням млекопитающих. [ 3 ] Предполагается, что у некоторых дицинодонтов наличие бивней связано с половым диморфизмом . [ 4 ] У некоторых дицинодонтов, таких как Stahleckeria, не было настоящих бивней, а вместо этого они имели клыкообразные расширения по бокам клюва. [ 5 ] [ 6 ] : 139
Тело короткое, сильное, бочкообразное, с сильными конечностями. У крупных родов (таких как Dinodontosaurus ) задние конечности держались прямо, а передние сгибались в локтях. И грудной пояс , и подвздошная кость большие и сильные. Хвост короткий. [ нужна ссылка ]
Следы дицинодонтов Pentasauropus позволяют предположить, что у дицинодонтов были мясистые подушечки на ногах. [ 7 ] Мумифицированная кожа экземпляров листрозавра из Южной Африки имеет многочисленные выпуклости. [ 8 ]
Эндотермия и анатомия мягких тканей
[ редактировать ]Дицинодонтов уже давно подозревали в том, что они теплокровные животные. Их кости хорошо васкуляризированы и имеют гаверсовы каналы , а пропорции тела способствуют сохранению тепла. [ 9 ] У молодых особей кости настолько сильно васкуляризированы, что имеют более высокую плотность каналов, чем у большинства других терапсидов. [ 10 ] Тем не менее, исследования позднего триаса дицинодонтов копролитов парадоксальным образом демонстрируют паттерны пищеварения, более типичные для животных с медленным метаболизмом. [ 11 ]
Совсем недавно открытие остатков волос в пермских копролитах, возможно, подтверждает статус дицинодонтов как эндотермных животных. Поскольку эти копролиты происходят от хищных видов, а переваренные кости дицинодонтов многочисленны, было высказано предположение, что по крайней мере некоторые из этих остатков волос происходят от добычи дицинодонтов. [ 12 ] Новое исследование с использованием химического анализа показало, что до пермского вымирания цинодонты и дицинодонты независимо развили теплую кровь. [ 13 ]
История
[ редактировать ]
Дицинодонты известны с середины 1800-х годов. Южноафриканский геолог Эндрю Геддес Бейн дал первое описание дицинодонтов в 1845 году. В то время Бейн руководил строительством военных дорог при Королевском инженерном корпусе и обнаружил множество окаменелостей рептилий во время своих исследований Южной Африки. Бэйн описал эти окаменелости в письме 1845 года, опубликованном в «Трудах Лондонского геологического общества» , назвав их «двузубыми» из-за двух выдающихся бивней. [ 14 ] В том же году английский палеонтолог Ричард Оуэн назвал два вида дицинодонтов из Южной Африки: Dicynodon Lacerticeps и Dicynodon bainii . Поскольку Бэйн был занят делами Королевского инженерного корпуса, он хотел, чтобы Оуэн более подробно описал его окаменелости. Оуэн не публиковал описание до 1876 года в своем «Описательном и иллюстрированном каталоге ископаемых рептилий Южной Африки в коллекции Британского музея» . [ 15 ] К этому времени было описано гораздо больше дицинодонтов. еще один важный вид, названный Ptychognathus declivis В 1859 году из Южной Африки был назван . В том же году Оуэн назвал группу Dicynodontia. [ 16 ] В своем «Описательном и иллюстрированном каталоге » Оуэн почтил Бэйна, предложив Bidentalia в качестве замены его Dicynodontia. [ 15 ] В последующие годы название Bidentalia быстро вышло из употребления, уступив место популярности Dicynodontia Оуэна. [ 17 ]
Эволюционная история
[ редактировать ]


Дицинодонты впервые появились в средней перми в Южном полушарии, где Южная Африка была центром их известного разнообразия, и претерпели быструю эволюционную радиацию , став глобально распространенными и одними из самых успешных и многочисленных наземных позвоночных в поздней перми . [ 18 ] [ 19 ] За это время они включали большое разнообразие экотипов, в том числе крупных, средних и мелких травоядных, а также кротообразных норных кротов с короткими конечностями. [ 20 ]
Известно, что только четыре линии пережили Великое Вымирание ; первые три представлены каждым одним родом: Myosaurus , Kombuisia и Lystrosaurus , причем последний является наиболее обычным и широко распространенным травоядным животным Инда ( ранний триас ). Ни один из них не просуществовал долго до триаса. Четвертой группой были Kannemeyeriformes , единственные дицинодонты, диверсифицировавшиеся в триасовый период. [ 21 ] Эти коренастые животные размером от свиньи до быка были самыми многочисленными травоядными животными в мире от оленекского до ладинского периода. В Карнии они были вытеснены траверсодонтными цинодонтами и рептилиями -ринхозаврами . периоде В норийском (середина позднего триаса), возможно, из-за увеличения засушливости, они резко сократились, и роль крупных травоядных животных взяли на себя динозавры -зауроподоморфы . [ нужна ссылка ]
Окаменелости размером с азиатского слона дицинодонта Lisowicia bojani , обнаруженные в Польше, указывают на то, что дицинодонты дожили, по крайней мере, до позднего норийского периода или самого раннего рэта (позднего триаса); это животное также было самым крупным известным видом дицинодонтов. [ 22 ] [ 23 ]
Шесть фрагментов ископаемых костей, обнаруженных в Квинсленде , Австралия, были интерпретированы как остатки черепа в 2003 году. Это указывает на то, что дицинодонты выжили в меловой период на юге Гондваны . [ 24 ] Родство этих экземпляров с дицинодонтами было подвергнуто сомнению (включая предположение о том, что они принадлежали баурузуховым крокодилаформам). Агнолина и др. В 2010 году [ 25 ] а в 2019 году Кнутсен и Эрлеманс посчитали, что это ископаемое относится к плио - плейстоценовому возрасту, и переосмыслили его как окаменелость крупного млекопитающего, вероятно, дипротодонтида . [ 26 ]
С упадком и исчезновением каннемейерид больше не было доминирующих крупных синапсидных травоядных до эпохи среднего палеоцена (60 млн лет назад), когда млекопитающие , отдаленные потомки цинодонтов , начали диверсифицироваться после вымирания нептичьих динозавров.
Систематика
[ редактировать ]Таксономия
[ редактировать ]Dicynodontia первоначально была названа английским палеонтологом Ричардом Оуэном . Он был возведен как семейство отряда Anomodontia и включало роды Dicynodon и Ptychognathus . Другие группы Anomodontia включали Gnathodontia , в которую входил Rhynchosaurus (теперь известный как архозавроморф ) и Cryptodontia , в которую входил Oudenodon . Криптодонты отличались от дицинодонтов отсутствием бивней. Несмотря на отсутствие бивней, уденодон теперь классифицируется как дицинодонт, и название Cryptodontia больше не используется. Томас Генри Хаксли пересмотрел Dicynodontia Оуэна как отряд, включающий Dicynodon и Oudenodon . [ 27 ] Позже Dicynodontia была отнесена к подотряду или инфраотряду с более крупной группой Anomodontia, которая классифицируется как отряд. В недавних исследованиях рейтинг Dicynodontia менялся: Ивахненко (2008) считал его подотрядом, Иванчненко (2008) считал его инфраотрядом, а Куркин (2010) считал его отрядом. [ 28 ]
Многие высшие таксоны, включая инфраотряды и семейства, были созданы как средство классификации большого количества видов дицинодонтов. Клювер и Кинг (1983) выделили несколько основных групп внутри Dicynodontia, включая Eodicynodontia (содержащую только Eodicynodon ), Endothiodontia (содержащую только Endothiodontidae ), Pristerodontia ( Pristerodontidae , Cryptodontidae , Aulacephalodontidae , Dicynodontidae , Lystrosauridae и Kannemeyeriidae ), Kingoriamorpha (содержат только Kingoriidae ), Diictodontia ( Diictodontidae , Robertiidae , Cistecephalidae , Emydopidae и Myosauridae ) и Venyukoviamorpha . [ 29 ] Большинство этих таксонов больше не считаются действительными. Каммерер и Ангельчик (2009) предположили, что проблематичная таксономия и номенклатура Dicynodontia и других групп является результатом большого количества противоречивых исследований и тенденции к ошибочному установлению недействительных названий. [ 17 ]






















Филогения
[ редактировать ]Ниже представлена модифицированная кладограмма Angielczyk et al. (2021): [ 30 ]
| Дицинодентия | |
Текущая классификация
[ редактировать ]- Дицинодентия
- Брахипросопус
- Колободеки
- Эодицинодон
- Лантаностегус
- Ньяпхули
- Эндотиодонтия
- Эумантеллиды
- Пилаецефалиды
- Терохелония
- Эмидопоидея
- Биденталия
- Криптодонтия
- Дицинодонтоидеа
- Кунилония
- Таохеодон
- Даптоцефалия
- Делектозавр
- Дицинодон
- Динаномодон
- Перамодон
- Эльф
- Гордония
- Интерпрезозавр
- Катумбия
- турфанодон
- Виваксозавр
- Листрозавриды
- Каннемейериформес
- Ангонизавр
- Динодонтозавриды
- Шансиодонтиды
- Каннемейериииды
- Stahleckeriidae
- ? Сунгеодон
- Вы будете волонтером
- Плацеринии
- Stahleckeriinae
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Рацки, Гжегож; Лукас, Спенсер Г. (2018). «Время вымирания дицинодонтов в свете необычной польской фауны позднего триаса и подхода Кювье к вымиранию». Историческая биология . 32 (4): 1–11. дои : 10.1080/08912963.2018.1499734 . S2CID 91926999 .
- ^ Кромптон, штат Аризона; Хоттон, Н. (1967). «Функциональная морфология жевательного аппарата двух дицинодонтов (Reptilia, Therapsida)». Постилла . 109 : 1–51.
- ^ Уитни, MR; Ангельчик, К.Д.; Пикук, БР; Сидор, Калифорния (2021 г.). «Эволюция синапсидного бивня: данные гистологии дицинодонта-терапсида» . Труды Королевского общества B: Биологические науки . 288 (1961). дои : 10.1098/rspb.2021.1670 . ПМЦ 8548784 . ПМИД 34702071 . S2CID 239890042 .
- ^ Ангельчик, Кеннет Д.; Бенуа, Жюльен; Рубидж, Брюс С. (февраль 2021 г.). Маршрут, Марчелло (ред.). «Новый дицинодонт-цистецефал с клыками (Therapsida, Anomodontia) из аргиллитовой формации верхней перми верхней Мадумабисы, бассейн Луангва, Замбия» . Статьи по палеонтологии 7 (1): 405–446. дои : 10.1002/spp2.1285 . ISSN 2056-2799 . S2CID 210304700 .
- ^ Каммерер, Кристиан Ф.; Ордоньес, Мария де лос Анхелес (01 июня 2021 г.). «Дицинодонты (Therapsida: Anomodontia) Южной Америки» . Журнал южноамериканских наук о Земле . 108 : 103171. Бибкод : 2021JSAES.10803171K . дои : 10.1016/j.jsames.2021.103171 . ISSN 0895-9811 . S2CID 233565963 .
- ^ Колберт, Э.Х. , (1969), Эволюция позвоночных , John Wiley & Sons Inc (2-е изд.)
- ^ Циттон, Пол; Диас-Мартинес, Игнасио; де Вале, Сильвина; Консолье-Гонелла, Карлос (7 августа 2018 г.). «Пятитадактильные следы триаса из группы Лос Менукос (провинция Рио-Негро, Патагония, Аргентина): возможные ограничения автоподиальной позы гондваанских гусеничных мастеров» . ПерДж . 6 : е5358. дои : 10.7717/peerj.5358 . ПМК 6086091 . ПМИД 30123702 .
- ^ Смит, Роджер М.Х.; Бота, Дженнифер; Вильетти, Пиа А. (октябрь 2022 г.). «Тафономия четвероногих, пострадавших от засухи, в бассейне Кару в раннем триасе, Южная Африка» . Палеогеография, Палеоклиматология, Палеоэкология . 604 : 111207. Бибкод : 2022PPP...60411207S . дои : 10.1016/j.palaeo.2022.111207 . S2CID 251781291 .
- ^ Баккер, Роберт Т. (апрель 1975 г.). «Возрождение динозавров». Научный американец . 232 (4): 58–79. Бибкод : 1975SciAm.232d..58B . doi : 10.1038/scientificamerican0475-58 .
- ^ Бота-Бринк, Дженнифер; Ангельчик, Кеннет Д. (2010). «Объясняют ли необычайно высокие темпы роста пермо-триасовых дицинодонтов (Therapsida, Anomodontia) их успех до и после вымирания в конце пермского периода?» . Зоологический журнал Линнеевского общества . 160 (2): 341–365. дои : 10.1111/j.1096-3642.2009.00601.x .
- ^ Байдек, Петр; Овоцкий, Кшиштоф; Недзведский, Гжегож (2014). «Предположительные дицинодонтовые копролиты из верхнего триаса Польши». Палеогеография, Палеоклиматология, Палеоэкология . 411 : 1–17. Бибкод : 2014PPP...411....1B . дои : 10.1016/j.palaeo.2014.06.013 .
- ^ Байдек, Петр; Кварнстрем, Мартин; Овоцкий, Кшиштоф; Сулей, Томаш; Сенников Андрей Г.; Голубев Валерий К.; Недзведский, Гжегож (2016). «Микробиота и остатки пищи, включая возможные свидетельства присутствия волос до млекопитающих в копролитах верхней перми из России». Летайя . 49 (4): 455–477. дои : 10.1111/лет.12156 .
- ^ Рей, Кевин; Амио, Ромен; Фурель, Франсуа; Абдала, Фернандо; Флюто, Фредерик; Джалиль, Нур-Эддин; Лю, Цзюнь; Рубидж, Брюс С.; Смит, Роджер М.Х.; Штайер, Ж. Себастьен; Вильетти, Пиа А; Ван, Сюй; Лекюе, Кристоф (2017). «Изотопы кислорода предполагают повышенный термометаболизм в пределах нескольких пермо-триасовых терапсидных клад» . электронная жизнь . 6 : e28589. дои : 10.7554/eLife.28589 . ПМК 5515572 . ПМИД 28716184 .
- ^ Бейн, А.Г. (1845 г.). «Об открытии ископаемых останков двузубых и других рептилий в Южной Африке» . Труды Лондонского геологического общества . 1 : 53–59. дои : 10.1144/GSL.JGS.1845.001.01.72 . hdl : 2027/uc1.c034667778 . S2CID 128602890 .
- ^ Перейти обратно: а б Оуэн, Р. (1876). Описательный и иллюстрированный каталог ископаемых рептилий Южной Африки в коллекции Британского музея . Лондон: Британский музей. п. 88.
- ^ Оуэн, Р. (1860). «Об отрядах ископаемых и современных рептилий и их распространении во времени». Отчет двадцать девятого собрания Британской ассоциации содействия развитию науки . 1859 : 153–166.
- ^ Перейти обратно: а б Каммерер, CF; Ангельчик, К.Д. (2009). «Предлагаемая высшая таксономия аномодонтных терапсидов» (PDF) . Зоотакса . 2018 : 1–24.
- ^ Куркин А.А. (июль 2011 г.). «Пермские аномодонты: Палеобиогеография и распространение группы» . Палеонтологический журнал . 45 (4): 432–444. дои : 10.1134/S0031030111030075 . ISSN 0031-0301 . S2CID 129331000 .
- ^ Олройд, Саванна Л.; Сидор, Кристиан А. (август 2017 г.). «Обзор глобальной летописи окаменелостей четвероногих в Гуадалупе (средняя пермь)» . Обзоры наук о Земле . 171 : 583–597. Бибкод : 2017ESRv..171..583O . doi : 10.1016/j.earscirev.2017.07.001 . ISSN 0012-8252 .
- ^ Ангельчик, Кеннет Д.; Каммерер, Кристиан Ф. (2018). «5. Синапсиды немлекопитающих: глубокие корни генеалогического древа млекопитающих». Эволюция, разнообразие и систематика млекопитающих . стр. 117–198. дои : 10.1515/9783110341553-005 . ISBN 9783110341553 . S2CID 92370138 .
- ^ Каммерер, Кристиан Ф.; Фрёбиш, Йорг; Ангельчик, Кеннет Д. (31 мая 2013 г.). «О достоверности и филогенетическом положении Eubrachiosaurus Browni , каннемейериформного дицинодонта (Anomodontia) из триасовой Северной Америки» . ПЛОС ОДИН . 8 (5): e64203. Бибкод : 2013PLoSO...864203K . дои : 10.1371/journal.pone.0064203 . ПМЦ 3669350 . ПМИД 23741307 .
- ^ Томаш Сулей; Гжегож Недзведски (2019). «Синапсид позднего триаса размером с слона с прямыми конечностями» . Наука . 363 (6422): 78–80. Бибкод : 2019Sci...363...78S . дои : 10.1126/science.aal4853 . ПМИД 30467179 .
- ^ Сен-Флер, Николас (4 января 2019 г.). «Родственник млекопитающих размером со слона, который пасся рядом с динозаврами» . Нью-Йорк Таймс . Проверено 6 января 2019 г.
- ^ Талборн, Т.; Тернер, С. (2003). «Последний дицинодонт: реликт австралийского мелового периода» . Труды Королевского общества B: Биологические науки . 270 (1518): 985–993. дои : 10.1098/rspb.2002.2296 . JSTOR 3558635 . ПМК 1691326 . ПМИД 12803915 .
- ^ Агнолин, Флорида; Эскурра, доктор медицины; Паис, DF; Солсбери, Юго-Запад (2010 г.). «Переоценка фауны нептичьих динозавров мелового периода из Австралии и Новой Зеландии: доказательства их родства с Гондваной» (PDF) . Журнал систематической палеонтологии . 8 (2): 257–300. дои : 10.1080/14772011003594870 . S2CID 130568551 .
- ^ Эспен М. Кнутсен; Эмма Орлеманс (2019). «Последний дицинодонт? Переоценка таксономических и временных связей спорного австралийского ископаемого». Исследования Гондваны . 77 : 184–203. дои : 10.1016/j.gr.2019.07.011 . S2CID 202908716 .
- ^ Осборн, HF (1904). «Реклассификация Рептилий» . Американский натуралист . 38 (446): 93–115. дои : 10.1086/278383 . S2CID 84492986 .
- ^ Куркин А.А. (2010). «Позднепермские дицинодонты Восточной Европы». Палеонтологический журнал . 44 (6): 72–80. дои : 10.1134/S0031030110060092 . S2CID 131459807 .
- ^ Клювер, Массачусетс; Кинг, GM (1983). «Переоценка взаимоотношений пермских дицинодонтов (Reptilia, Therapsida) и новая классификация дицинодонтов». Анналы Южноафриканского музея . 91 : 195–273.
- ^ Ангельчик, К.Д.; Лю, Дж.; Ян, В. (2021). «Переописание Kunpania scopulusa , двудентального дицинодонта (Therapsida, Anomodontia) из Гваделупы Северо-Западного Китая». Журнал палеонтологии позвоночных . 41 (1): e1922428. Бибкод : 2021JVPal..41E2428A . дои : 10.1080/02724634.2021.1922428 . S2CID 236406006 .
Дальнейшее чтение
[ редактировать ]- Кэрролл, Р.Л. (1988), Палеонтология и эволюция позвоночных , WH Freeman & Co.
- Кокс Б., Сэвидж Р.Дж.Г., Гардинер Б., Харрисон К. и Палмер Д. (1988) Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла , 2-е издание, Marshall Publishing
- Кинг, Джиллиан М. , «Аномодонтия», часть 17 C, Энциклопедия палеогерпетологии , Гуцав Фишер Верлаг, Штутгарт и Нью-Йорк, 1988 г.
- Кинг, Джиллиан М., 1990, Дицинодонты: исследование палеобиологии , Чепмен и Холл, Лондон и Нью-Йорк.





