Ископаемое

Ископаемое ( от классического латинского ископаемых , букв. « полученное путем раскопок » ) [1] Это любые сохранившиеся останки, отпечатки или следы некогда живого существа из прошлой геологической эпохи . Примеры включают кости , ракушки , экзоскелеты , каменные отпечатки животных или микробов , предметы, сохранившиеся в янтаре , волосы , окаменелое дерево и ДНК остатки . Совокупность окаменелостей известна как летопись окаменелостей . Хотя летопись окаменелостей неполна, многочисленные исследования показали, что имеется достаточно информации, чтобы дать хорошее представление о модели разнообразия жизни на Земле. [2] [3] [4] Кроме того, запись может предсказать и заполнить такие пробелы, как открытие Тиктаалика в арктике Канады . [5]
Палеонтология включает изучение окаменелостей: их возраста, способа образования и эволюционного значения. Образцы обычно считаются окаменелостями, если их возраст превышает 10 000 лет. [6] Возраст самых старых окаменелостей составляет около 3,48 миллиарда лет. [7] [8] [9] до 4,1 миллиарда лет. [10] [11] Наблюдение в XIX веке того, что определенные окаменелости были связаны с определенными пластами горных пород , привело к признанию геологической шкалы времени и относительного возраста различных окаменелостей. Развитие методов радиометрического датирования в начале 20 века позволило ученым количественно измерить абсолютный возраст горных пород и находящихся в них окаменелостей.
Существует множество процессов, которые приводят к окаменению , включая перминерализацию , отливки и плесень, аутигенную минерализацию , замещение и рекристаллизацию, сжатие, карбонизацию и биоиммурацию.
Окаменелости различаются по размеру: от бактерий размером один микрометр (1 мкм). [12] динозаврам . и деревьям длиной в несколько метров и весом во многие тонны В окаменелостях обычно сохраняется только часть умершего организма, обычно та часть, которая была частично минерализована в течение жизни, например кости и зубы позвоночных животных или хитиновый или известковый экзоскелет беспозвоночных . Окаменелости также могут состоять из следов, оставленных организмом при его жизни, например, следов животных или фекалий ( копролиты ). Эти типы окаменелостей называются следами окаменелостей или ихноокаменелостями , в отличие от окаменелостей тела . Некоторые окаменелости являются биохимическими и называются хемофоссилиями или биосигнатурами .
| Часть серии о |
| Палеонтология |
|---|
 |
|
Палеонтологический портал Категория |
История обучения
Сбор окаменелостей датируется, по крайней мере, началом письменной истории. Сами окаменелости называются летописью окаменелостей. Летопись окаменелостей была одним из первых источников данных, лежащих в основе изучения эволюции , и продолжает иметь отношение к истории жизни на Земле . Палеонтологи изучают летопись окаменелостей, чтобы понять процесс эволюции и то, как отдельные виды развивались .
Древние цивилизации

Окаменелости были видимы и распространены на протяжении большей части естественной истории, поэтому документально подтвержденное взаимодействие человека с ними восходит к письменной истории или даже раньше.
есть много примеров палеолитических В Европе каменных ножей с ископаемыми иглокожими , установленными точно на рукоятке, относящимися к Homo heidelbergensis и неандертальцам . [13] Эти древние народы также просверливали отверстия в центре этих круглых ископаемых раковин, по-видимому, используя их в качестве бус для ожерелий.
Древние египтяне собирали окаменелости видов, напоминающих кости современных видов, которым они поклонялись. Бог Сет ассоциировался с гиппопотамом , поэтому в храмах этого божества хранились окаменелые кости бегемоподобных видов. [14] Пятилучевые ископаемые раковины морских ежей ассоциировались с божеством Сопду , Утренней звездой, эквивалентом Венеры в римской мифологии. [13]
Окаменелости, по-видимому, внесли непосредственный вклад в мифологию многих цивилизаций, включая древних греков. Классический греческий историк Геродот писал о местности недалеко от Гипербореи , где грифоны охраняли золотые сокровища. действительно велась добыча золота В этом районе , где черепа клювистых протоцератопсов были обычным явлением в виде окаменелостей.
Более поздний греческий ученый Аристотель в конце концов понял, что ископаемые морские ракушки из камней похожи на те, что были найдены на пляже, что указывает на то, что окаменелости когда-то были живыми животными. Ранее он объяснял их с точки зрения выдохов испарения . [15] которую персидский эрудит Авиценна преобразовал в теорию окаменевших жидкостей ( succus lapidificatus ). Признание того, что ископаемые морские ракушки происходят из моря, было основано в 14 веке Альбертом Саксонским и принято в той или иной форме большинством натуралистов к 16 веку. [16]
Римский натуралист Плиний Старший писал о « камнях языка », которые он называл глоссопетрами . Это были ископаемые зубы акулы, которые в некоторых классических культурах считались похожими на языки людей или змей. [17] Он также писал о рогах Аммона , которые представляют собой ископаемые аммониты , откуда группа осьминогов с панцирем в конечном итоге получила свое современное название. Плиний также делает одно из ранее известных упоминаний о жабках , которые до XVIII века считались магическим лекарством от яда, происходящего из голов жаб, но которые представляют собой ископаемые зубы лепидотов , меловой лучепёрой рыбы. [18]
Считается, что у равнинных племен Северной Америки окаменелости, такие как многие неповрежденные окаменелости птерозавров, естественно обнаруженные в этом регионе, связаны с их собственной мифологией о громовой птице . [19]
В доисторической Африке не существует такой прямой мифологической связи, но есть немало свидетельств того, что племена там раскапывали и перемещали окаменелости в церемониальные места, по-видимому, относясь к ним с некоторым почтением. [20]
В Японии зубы ископаемых акул ассоциировались с мифическими тенгу , которые считались острыми, как бритва, когтями этого существа, задокументированными спустя некоторое время после 8-го века нашей эры. [17]
В средневековом Китае ископаемые кости древних млекопитающих, в том числе человека прямоходящего, часто принимали за « кости дракона » и использовали в качестве лекарств и афродизиаков . Кроме того, некоторые из этих ископаемых костей собираются как «предметы искусства» учеными, которые оставили надписи на различных артефактах с указанием времени их добавления в коллекцию. Хорошим примером является известный ученый Хуан Тинцзянь из династии Сун в 11 веке, который хранил особую окаменелость морской ракушки, на которой было выгравировано его собственное стихотворение. [21] В своих «Очерках бассейна снов», династии Сун опубликованных в 1088 году, китайский ученый и официальный представитель Шэнь Го выдвинул гипотезу, что морские окаменелости, найденные в геологическом слое гор, расположенных в сотнях миль от Тихого океана, являются свидетельством того, что доисторическое морское побережье когда-то существовало там и менялось на протяжении веков. времени . [22] [23] Его наблюдения за окаменевшим бамбуком в засушливой северной климатической зоне нынешнего Яньань , провинция Шэньси , Китай, привели его к выдвижению первых идей о постепенном изменении климата из-за того, что бамбук естественным образом растет в районах с более влажным климатом. [23] [24] [25]
В средневековом христианском мире окаменелые морские существа на склонах гор рассматривались как доказательство библейского потопа из Ноева ковчега . Наблюдая за существованием ракушек в горах, древнегреческий философ Ксенофан (ок. 570–478 до н. э.) предположил, что когда-то мир был затоплен великим наводнением, которое похоронило живые существа в засыхающей грязи. [26] [27]
В 1027 году персидский Авиценна объяснил каменистость окаменелостей в «Книге исцеления» :
Если то, что сказано об окаменении животных и растений, верно, то причиной этого (феномена) является мощная минерализующая и окаменевающая сила, которая возникает в определенных каменистых местах или исходит внезапно из земли при землетрясении и оседаниях и окаменевает все, что приходит. вступить с ним в контакт. В сущности, окаменение тел растений и животных не более необычно, чем превращение вод. [28]
С 13-го века до наших дней ученые отмечали, что ископаемые черепа Deinotherium giganteum , найденные на Крите и в Греции, могли быть интерпретированы как черепа циклопов из греческой мифологии и, возможно, являются источником этого греческого мифа. . [29] [30] Их черепа, кажется, имеют единственное отверстие для глаз спереди, как и их современные родственники- слоны , хотя на самом деле это отверстие для их хобота.

В скандинавской мифологии раковины иглокожих (круглая пятичастная пуговица, оставшаяся от морского ежа) ассоциировались с богом Тором и не только включались в громовые камни , изображения молота Тора и последующие кресты в форме молота, когда христианство было принято, но и также держат в домах, чтобы обеспечить защиту Тора. [13]
Из них выросли пастушьи короны из английского фольклора, которые использовались для украшения и в качестве талисманов на удачу и размещались у дверей домов и церквей. [31] В Саффолке другой вид использовался пекарями в качестве талисмана на удачу, которые называли их « сказочными хлебами» , связывая их с буханками хлеба аналогичной формы, которые они пекли. [32] [33]
Ранние современные объяснения
Более научные взгляды на окаменелости возникли в эпоху Возрождения . Леонардо да Винчи согласился с мнением Аристотеля о том, что окаменелости являются остатками древней жизни. [34] : 361 Например, Леонардо заметил несоответствия библейскому повествованию о потопе как объяснению происхождения ископаемых:
Если бы Потоп разнес раковины на расстояние трех-четырехсот миль от моря, он бы смешал их с различными другими природными объектами и свалил бы их в кучу; но даже на таком расстоянии от моря мы видим всех устриц вместе, а также моллюсков, каракатиц и всех других ракушек, собравшихся вместе, найденных все вместе мертвыми; и одиночные раковины встречаются отдельно друг от друга, какими мы видим их каждый день на морских берегах.
И мы находим устриц очень большими семьями, среди которых можно увидеть у некоторых раковины, все еще соединенные вместе, что указывает на то, что они были оставлены там морем и что они еще были живы, когда был прорезан Гибралтарский пролив. В горах Пармы и Пьяченцы можно увидеть множество ракушек и кораллов с дырочками, все еще прилипших к камням... [35]

В 1666 году Николас Стено исследовал акулу и связал ее зубы с «языковыми камнями» древней греко-римской мифологии, заключив, что на самом деле это были не языки ядовитых змей, а зубы каких-то давно вымерших существ. вид акулы. [17]
Роберт Гук (1635–1703) включил микрофотографии окаменелостей в свою «Микрографию» и был одним из первых, кто наблюдал ископаемые формы . Его наблюдения за окаменелостями, которые, как он утверждал, представляют собой окаменелые останки существ, некоторые из которых больше не существовали, были опубликованы посмертно в 1705 году. [36]
Уильям Смит (1769–1839) , английский инженер каналов, заметил, что в породах разного возраста (на основании закона суперпозиции ) сохранились разные совокупности окаменелостей, и что эти совокупности следовали друг за другом в регулярном и определяемом порядке. Он заметил, что камни из отдаленных мест можно сопоставить на основе содержащихся в них окаменелостей. Он назвал это принципом преемственности фауны . Этот принцип стал одним из главных дарвиновских доказательств реальности биологической эволюции.
Жорж Кювье пришел к выводу, что большинство, если не все, окаменелости животных, которые он исследовал, были остатками вымерших видов. Это привело Кювье к тому, что он стал активным сторонником геологической школы мысли, называемой катастрофизмом . Ближе к концу своей статьи 1796 года о живых и ископаемых слонах он сказал:
Все эти факты, согласующиеся между собой и не противоречащие никаким сообщениям, кажутся мне доказательством существования мира, предшествующего нашему, разрушенного какой-то катастрофой. [37]

Интерес к окаменелостям и геологии в целом расширился в начале девятнадцатого века. В Британии открытия Мэри Эннинг окаменелостей, в том числе первого полного ихтиозавра и полного скелета плезиозавра , вызвали общественный и научный интерес. [38]
Линней и Дарвин
Ранние натуралисты хорошо понимали сходства и различия современных видов, что привело Линнея к разработке иерархической системы классификации, которая используется до сих пор. Дарвин и его современники впервые связали иерархическую структуру древа жизни с тогда еще очень скудной летописью окаменелостей. Дарвин красноречиво описал процесс происхождения с модификациями, или эволюцию, в ходе которой организмы либо адаптируются к естественным и изменяющимся воздействиям окружающей среды, либо погибают.
Когда Дарвин писал «Происхождение видов посредством естественного отбора, или сохранение избранных рас в борьбе за жизнь» , древнейшими окаменелостями животных были окаменелости кембрийского периода, возраст которых, как теперь известно, составляет около 540 миллионов лет. Его беспокоило отсутствие более старых окаменелостей из-за последствий для достоверности его теорий, но он выразил надежду, что такие окаменелости будут найдены, отметив, что: «только небольшая часть мира известна точно». Дарвин также размышлял над внезапным появлением многих групп (т.е. типов ) в древнейших известных кембрийских ископаемых слоях. [39]
После Дарвина
Со времен Дарвина летопись окаменелостей увеличилась до 2,3–3,5 миллиардов лет. [40] Большинство этих докембрийских окаменелостей представляют собой микроскопические бактерии или микроокаменелости . Однако сейчас известны макроскопические окаменелости позднего протерозоя. ( Биота Эдиакары также называемая вендской биотой), датируемая 575 миллионами лет назад, в совокупности представляет собой богато разнообразную совокупность ранних многоклеточных эукариот .
Летопись окаменелостей и последовательность фауны составляют основу науки биостратиграфии или определения возраста горных пород на основе залегающих в них окаменелостей. В течение первых 150 лет геологии биостратиграфия и суперпозиция были единственными средствами определения относительного возраста горных пород. Геологическая шкала времени была разработана на основе относительного возраста пластов горных пород, определенного ранними палеонтологами и стратиграфами .
С первых лет двадцатого века методы абсолютного датирования , такие как радиометрическое датирование (включая калия/аргона , аргона/аргона , ряды урана и, для самых недавних окаменелостей, радиоуглеродное датирование ), использовались для проверки относительного возраста, полученного методом окаменелостей и определить абсолютный возраст многих окаменелостей. Радиометрическое датирование показало, что возраст самых ранних известных строматолитов превышает 3,4 миллиарда лет.
Современная эпоха
Летопись окаменелостей — это эволюционная эпопея жизни, которая разворачивалась на протяжении четырех миллиардов лет, пока условия окружающей среды и генетический потенциал взаимодействовали в соответствии с естественным отбором.
Виртуальный музей окаменелостей [41]
Палеонтология объединилась с эволюционной биологией, чтобы разделить междисциплинарную задачу по описанию древа жизни, которое неизбежно ведет назад во времени к докембрийской микроскопической жизни, когда развивались клеточная структура и функции. Глубокое время существования Земли в протерозое и еще глубже в архее «описывается только с помощью микроскопических окаменелостей и тонких химических сигналов». [42] Молекулярные биологи, используя филогенетику , могут сравнивать гомологию аминокислотных или нуклеотидных последовательностей белков (т.е. сходство) для оценки таксономии и эволюционных расстояний между организмами с ограниченной статистической достоверностью. С другой стороны, изучение окаменелостей может более точно определить, когда и в каком организме впервые появилась мутация. Филогенетика и палеонтология работают вместе, проясняя все еще смутные представления науки о появлении жизни и ее эволюции. [43]

Найлсом Элдриджем Исследование Phacops рода трилобитов подтвердило гипотезу о том, что изменения в расположении глазных линз трилобита происходили урывками на протяжении миллионов лет в девонском периоде . [44] Интерпретация Элдриджем летописи окаменелостей Факопа заключалась в том, что окаменели последствия изменений хрусталика, а не быстро происходящий эволюционный процесс. Эти и другие данные побудили Стивена Джея Гулда и Найлза Элдриджа опубликовать свою основополагающую статью о прерывистом равновесии в 1971 году.
Синхротронный рентгеновский томографический анализ ранних кембрийских двусторонних эмбриональных микрокаменелостей позволил по-новому взглянуть на эволюцию многоклеточных животных на самых ранних стадиях. Метод томографии обеспечивает ранее недостижимое трехмерное разрешение на границах окаменения. загадочных двулатерий, червеобразной Markuelia и предполагаемого примитивного протостома Pseudooides Окаменелости двух , позволяют взглянуть на зародышевого листка эмбриональное развитие . Эти эмбрионы возрастом 543 миллиона лет подтверждают появление некоторых аспектов развития членистоногих раньше, чем считалось ранее, в позднем протерозое. Сохранившиеся эмбрионы из Китая и Сибири подверглись быстрой диагенетической фосфатизации, что привело к превосходной сохранности, включая клеточные структуры. [ жаргон ] Это исследование является ярким примером того, как знания, закодированные в летописи окаменелостей, продолжают предоставлять недостижимую иначе информацию о возникновении и развитии жизни на Земле. Например, исследование предполагает, что Markuelia имеет самое близкое родство с червями-приапулидами и соседствует с эволюционным ветвлением Priapulida , Nematoda и Arthropoda . [45] [ жаргон ]
Несмотря на значительные успехи в обнаружении и идентификации палеонтологических образцов, общепринято считать, что летопись окаменелостей крайне неполна. [46] [47] Подходы к измерению полноты летописи окаменелостей были разработаны для многочисленных подмножеств видов, в том числе сгруппированных таксономически, [48] [49] временно, [50] экологически/географически, [51] или в сумме. [52] [53] Это охватывает область тафономии и изучение систематических ошибок в палеонтологических записях. [54] [55] [56]
Знакомства/Возраст
Стратиграфия и оценки

Палеонтология стремится отобразить, как жизнь развивалась в геологическом времени. Существенным препятствием является сложность определения возраста ископаемых. Слои, в которых сохранились окаменелости, обычно не содержат радиоактивных элементов, необходимых для радиометрического датирования . Этот метод — наш единственный способ придать абсолютный возраст камням возрастом более 50 миллионов лет, и его точность может достигать 0,5% или выше. [57] Хотя радиометрическое датирование требует тщательной лабораторной работы, его основной принцип прост: известны скорости распада различных радиоактивных элементов , поэтому отношение радиоактивного элемента к продуктам его распада показывает, как давно радиоактивный элемент был включен в породу. Радиоактивные элементы распространены только в породах вулканического происхождения, поэтому единственные ископаемые породы, которые можно датировать радиометрически, — это слои вулканического пепла, которые могут служить конечными концами промежуточных отложений. [57]
Следовательно, палеонтологи полагаются на стратиграфию для датировки окаменелостей. Стратиграфия – это наука о расшифровке «слоеного пирога», то есть осадочной пластинки. [58] Породы обычно образуют относительно горизонтальные слои, причем каждый слой моложе предыдущего. Если ископаемое находится между двумя слоями, возраст которых известен, утверждается, что возраст ископаемого находится между двумя известными возрастами. [59] Поскольку толщи горных пород не являются непрерывными, а могут быть разбиты разломами или периодами эрозии , очень сложно сопоставить пласты горных пород, которые не находятся непосредственно рядом. Однако для сопоставления изолированных пород можно использовать окаменелости видов, выживавших в течение относительно короткого времени: этот метод называется биостратиграфией . Например, конодонт Eoplacognathus pseudoplanus имеет небольшой ареал в среднем ордовике. [60] Если породы неизвестного возраста имеют следы E. pseudoplanus , то они имеют среднеордовикский возраст. Чтобы быть полезными, такие индексные окаменелости должны быть отличительными, быть распространенными по всему миру и занимать короткий временной интервал. Вводящие в заблуждение результаты получаются, если индексные окаменелости датированы неправильно. [61] Стратиграфия и биостратиграфия вообще могут дать только относительную датировку ( A была до B ), чего часто бывает достаточно для изучения эволюции. Однако в некоторые периоды времени это сложно из-за проблем, связанных с сопоставлением пород одного возраста на разных континентах . [61] Отношения генеалогического древа также помогают сузить дату первого появления родословной. Например, если окаменелости B или C датируются X миллионов лет назад и рассчитанное «генеалогическое древо» говорит, что A был предком B и C, то A, должно быть, развился раньше.
Также можно оценить, как давно разошлись две ныне живущие клады, другими словами, примерно, как давно должен был жить их последний общий предок, предполагая, что мутации ДНК накапливаются с постоянной скоростью. Однако эти « молекулярные часы » ошибочны и обеспечивают лишь приблизительное время: например, они недостаточно точны и надежны для оценки того, когда впервые возникли группы, участвующие в кембрийском взрыве . [62] а оценки, полученные с помощью разных методов, могут отличаться в два раза. [63]
Ограничения
Организмы лишь в редких случаях сохраняются в виде окаменелостей и при самых благоприятных обстоятельствах, и лишь небольшая часть таких окаменелостей была обнаружена. Это иллюстрируется тем фактом, что число видов, известных по летописи окаменелостей, составляет менее 5% от числа известных живых видов, а это позволяет предположить, что количество видов, известных по окаменелостям, должно быть намного меньше 1% всех видов, которые когда-либо жили. [64] Из-за особых и редких обстоятельств, необходимых для окаменения биологической структуры, можно ожидать, что лишь небольшой процент форм жизни будет представлен в открытиях, и каждое открытие представляет собой лишь снимок процесса эволюции. Сам переход может быть проиллюстрирован и подтвержден только переходными окаменелостями, которые никогда не смогут продемонстрировать точную половину пути. [65]
Летопись окаменелостей сильно смещена в сторону организмов с твердыми частями, в результате чего большинство групп организмов с мягким телом практически не играют никакой роли. [64] Он изобилует моллюсками , позвоночными , иглокожими , брахиоподами и некоторыми группами членистоногих . [66]
Сайты
депозиты
Местонахождения окаменелостей исключительной сохранности, иногда включающие сохранившиеся мягкие ткани, известны как Lagerstätten , что по-немецки означает «места хранения». Эти образования могли возникнуть в результате захоронения туш в бескислородной среде с минимальным количеством бактерий, что замедляло разложение. Лагерштеттены охватывают геологический период от кембрийского периода до наших дней . Во всем мире одними из лучших примеров почти идеального окаменения являются кембрийские сланцы Маотяньшань и сланцы Бёрджесс , девонские сланцы Хунсрюк , юрские известняки Зольнхофена и каменноугольные месторождения Мазон-Крик .
Процессы окаменения
Рекристаллизация
Говорят, что ископаемое перекристаллизовалось, если исходные скелетные соединения все еще присутствуют, но в другой кристаллической форме, например, от арагонита до кальцита . [67]
-

-
 Ископаемая раковина Busycon sp., перекристаллизованная кальцитом. из Индрио Пит
Ископаемая раковина Busycon sp., перекристаллизованная кальцитом. из Индрио Пит -

-

-
 Рекристаллизованная раковина двустворчатого моллюска со сторовым кальцитом из формации Берд Спринг.
Рекристаллизованная раковина двустворчатого моллюска со сторовым кальцитом из формации Берд Спринг.





Замена

Замена происходит, когда панцирь, кость или другая ткань заменяется другим минералом. В некоторых случаях минеральное замещение исходной раковины происходит настолько постепенно и в таких мелких масштабах, что особенности микроструктуры сохраняются, несмотря на полную утрату исходного материала:
Перминерализация
Перминерализация — это процесс окаменения, который происходит при захоронении организма. Пустые пространства внутри организма (пространства, заполненные жидкостью или газом при жизни) заполняются богатой минералами грунтовой водой . Минералы выпадают в осадок из грунтовых вод, занимая пустые места. Этот процесс может происходить в очень небольших пространствах, например, внутри клеточной стенки растительной клетки . Мелкомасштабная перминерализация может привести к образованию очень детализированных окаменелостей. [68] Чтобы произошла перминерализация, организм должен покрыться осадком вскоре после смерти, в противном случае останки разрушаются падальщиками или разлагаются. [69] Степень разложения останков при покрытии определяет более поздние детали окаменелости. Некоторые окаменелости состоят только из останков скелета или зубов; другие окаменелости содержат следы кожи , перьев или даже мягких тканей. [70] Это форма диагенеза .
фосфатирование
Он включает в себя процесс окаменения, при котором органическое вещество заменяется обильными кальциево - фосфатными минералами . Образующиеся окаменелости имеют тенденцию быть особенно плотными и иметь темную окраску от темно-оранжевого до черного. [71]
Пиритизация
Эта консервация окаменелостей включает в себя элементы серы и железа . Организмы могут пиритизироваться, находясь в морских отложениях, насыщенных сульфидами железа. По мере разложения органического вещества выделяется сульфид, который вступает в реакцию с растворенным железом в окружающих водах, образуя пирит . Пирит заменяет карбонатный материал раковины из-за недостаточного насыщения карбонатом окружающих вод. Некоторые растения пиритизируются при нахождении в глинистой местности, но в меньшей степени, чем в морской среде. Некоторые пиритизированные окаменелости включают докембрийские микроископаемые, морских членистоногих и растения. [72] [73]
-
 пиритизированного аммоноидея Pleuroceras Solare Образец окаменелости
пиритизированного аммоноидея Pleuroceras Solare Образец окаменелости -
 Пиритизированный экземпляр брахиопода Paraspirifer Bownockeri.
Пиритизированный экземпляр брахиопода Paraspirifer Bownockeri. -
 Пиритизированный Triarthrus eatoni из формации Whetstone Gulf
Пиритизированный Triarthrus eatoni из формации Whetstone Gulf -
 Пиритизированный Furcaster палеозойкус из сланца Хунсрюк
Пиритизированный Furcaster палеозойкус из сланца Хунсрюк -
 Пиритизированный Tornoceras uniangulare из формации Ладловилль.
Пиритизированный Tornoceras uniangulare из формации Ладловилль.





окремнение
При окварцевании осаждение кремнезема из насыщенных водоемов имеет решающее значение для сохранения окаменелостей. Насыщенная минералами вода проникает в поры и клетки какого-то мертвого организма, где превращается в гель . Со временем гель обезвоживается , образуя богатую кремнеземом кристаллическую структуру, которая может быть выражена в форме кварца , халцедона , агата , опала и других, сохраняя первоначальную форму. [74] [75]
-
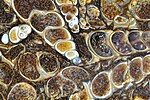 Халцедон заменил ископаемые раковины Elimia tenera включениями остракод.
Халцедон заменил ископаемые раковины Elimia tenera включениями остракод. -
 халцедонизированных брюхоногих моллюсков. Внутренние формы
халцедонизированных брюхоногих моллюсков. Внутренние формы -
 Агатизированные внутренние слепки брюхоногих моллюсков из ловушек Декана.
Агатизированные внутренние слепки брюхоногих моллюсков из ловушек Декана. -
 Агатизированный ископаемый коралл из Флориды
Агатизированный ископаемый коралл из Флориды -
 Ископаемые двустворчатые моллюски заменены опалом из Квинсленда.
Ископаемые двустворчатые моллюски заменены опалом из Квинсленда.





Отливки и формы
В некоторых случаях первоначальные остатки организма полностью растворяются или разрушаются иным образом. Оставшееся в камне отверстие в форме организма называется внешней формой . Если эта пустота позже заполняется осадком, полученный слепок будет напоминать внешний вид организма. Эндокаст , является результатом отложений , , или внутренняя плесень заполняющих внутреннюю часть организма, например, внутреннюю часть двустворчатого моллюска , улитки или полость черепа . [76] Эндокасты иногда называют Штейнкернами , особенно когда двустворчатые моллюски сохраняются таким образом. [77]
-
 Внутренняя плесень (steinkern) Hormotoma sp. из формации Галена
Внутренняя плесень (steinkern) Hormotoma sp. из формации Галена -
 Внутренняя форма брюхоногих моллюсков (штайнкерн) из формации Вентана .
Внутренняя форма брюхоногих моллюсков (штайнкерн) из формации Вентана . -
 Внешняя форма раковины Anomalodonta gigantea из формации Уэйнсвилл.
Внешняя форма раковины Anomalodonta gigantea из формации Уэйнсвилл. -
 Внутренняя плесень (steinkern) Glycymeris alpinus , Австрия.
Внутренняя плесень (steinkern) Glycymeris alpinus , Австрия. -






Аутигенная минерализация
Это особая форма формирования отливок и форм. Если химия правильная, организм (или фрагмент организма) может действовать как ядро для осаждения минералов, таких как сидерит , в результате чего вокруг него образуются узелки. Если это произойдет быстро, до значительного разрушения органической ткани, можно сохранить очень мелкие трехмерные морфологические детали. Конкреции из каменноугольных отложений ископаемых отложений Мейзон-Крик в Иллинойсе, США, являются одними из лучших документированных примеров такой минерализации. [78]
Адпрессия (сжатие-впечатление)
Сжатые окаменелости , такие как ископаемые папоротники, являются результатом химического восстановления сложных органических молекул, составляющих ткани организма. В этом случае ископаемое состоит из исходного материала, хотя и в геохимически измененном состоянии. Это химическое изменение является выражением диагенеза . Часто то, что остается, представляет собой углеродистую пленку, известную как фитолейм, и в этом случае окаменелость называется сжатием. Однако часто фитолейма теряется, и все, что остается, — это отпечаток организма в породе — отпечаток окаменелости. Однако во многих случаях сдавление и вдавление происходят одновременно. Например, когда горная порода расколота, фитолей часто прикрепляется к одной части (сжатие), тогда как аналог остается лишь отпечатком. По этой причине один термин охватывает два способа сохранения: сжатие . [79]
Карбонизация и углефикация
Обугленные или углефицированные окаменелости состоят из органических остатков, которые были восстановлены в основном до химического элемента углерода. Карбонизированные окаменелости состоят из тонкой пленки, которая образует силуэт исходного организма, а исходные органические останки обычно представляли собой мягкие ткани. Угольные окаменелости состоят в основном из угля, а первоначальные органические останки обычно имели древесный состав.
-
![Карбонизированная окаменелость циклоневрального червя, которого когда-то ошибочно приняли за пиявку[80] из силурийской биоты Ваукеша в Висконсине.](//upload.wikimedia.org/wikipedia/commons/thumb/a/ac/Probable_leech_from_the_Waukesha_Biota.jpg/111px-Probable_leech_from_the_Waukesha_Biota.jpg) Карбонизированная окаменелость циклоневрального червя, которого когда-то ошибочно приняли за пиявку [80] из силурийской биоты ваукеша штата Висконсин.
Карбонизированная окаменелость циклоневрального червя, которого когда-то ошибочно приняли за пиявку [80] из силурийской биоты ваукеша штата Висконсин. -

![Карбонизированная окаменелость циклоневрального червя, которого когда-то ошибочно приняли за пиявку[80] из силурийской биоты Ваукеша в Висконсине.](http://upload.wikimedia.org/wikipedia/commons/thumb/a/ac/Probable_leech_from_the_Waukesha_Biota.jpg/111px-Probable_leech_from_the_Waukesha_Biota.jpg)

Сохранение мягких тканей, клеток и молекул
Из-за их древности неожиданным исключением из изменения тканей организма путем химического восстановления сложных органических молекул во время окаменелости стало открытие мягких тканей в окаменелостях динозавров, включая кровеносные сосуды, а также выделение белков и доказательств наличия фрагментов ДНК. . [81] [82] [83] [84] В 2014 году Мэри Швейцер и ее коллеги сообщили о наличии частиц железа ( гетит -aFeO(OH)) в мягких тканях, извлеченных из окаменелостей динозавров. На основании различных экспериментов по изучению взаимодействия железа в гемоглобине с тканью кровеносных сосудов они предположили, что гипоксия раствора в сочетании с хелатированием железа повышает стабильность и сохранность мягких тканей и дает основу для объяснения непредвиденной сохранности ископаемых мягких тканей. [85] Однако более раннее исследование, основанное на восьми таксонах, относящихся к периоду от девона до юры, показало, что во всех этих окаменелостях сохранились достаточно хорошо сохранившиеся фибриллы, которые, вероятно, представляют собой коллаген , и что качество сохранности зависело в основном от расположения коллагена. волокна с плотной упаковкой, способствующей хорошей сохранности. [86] Судя по всему, в этот период времени не было никакой корреляции между геологическим возрастом и качеством сохранности.
Биоиммурация

Биоиммурация происходит, когда скелетный организм перерастает или иным образом поглощает другой организм, сохраняя последний или его отпечаток внутри скелета. [88] Обычно это сидячий скелетный организм, например мшанка или устрица , растущий вдоль субстрата , покрывая других сидячих склеробионтов . Иногда биоиммурированный организм оказывается мягкотелым и затем сохраняется в негативном рельефе как своего рода внешняя форма. Бывают также случаи, когда организм селится поверх живого скелетного организма, который растет вверх, сохраняя поселенца в своем скелете. Биоиммурация известна в летописи окаменелостей ордовика. [89] к Недавнему. [88]
Типы

Индекс
Индексные окаменелости (также известные как направляющие окаменелости, индикаторные окаменелости или зональные окаменелости) — это окаменелости, используемые для определения и идентификации геологических периодов (или стадий фауны). Они исходят из того, что, хотя разные отложения могут выглядеть по-разному в зависимости от условий, при которых они отложились, они могут включать останки одного и того же вида окаменелостей. Чем короче временной диапазон существования вида, тем точнее можно соотнести различные отложения, и поэтому окаменелости быстро развивающихся видов особенно ценны. Лучшие окаменелости-индексы распространены, их легко идентифицировать на видовом уровне и они широко распространены — в противном случае вероятность найти и распознать одну из двух отложений мала.
След
Следы окаменелостей состоят в основном из следов и нор, но также включают копролиты (ископаемые фекалии ) и следы, оставленные в результате питания. [90] [91] Следы окаменелостей особенно важны, поскольку они представляют собой источник данных, который не ограничивается животными с легко окаменелыми твердыми частями, и отражают поведение животных. Многие следы датируются значительно раньше, чем окаменелости тел животных, которые, как считается, были способны их создавать. [92] Хотя точное отнесение следов окаменелостей к их создателям, как правило, невозможно, следы могут, например, служить самым ранним физическим свидетельством появления животных умеренной сложности (сравнимых с дождевыми червями ). [91]
Копролиты классифицируются как следы окаменелостей, а не окаменелости тела, поскольку они свидетельствуют о поведении животного (в данном случае о диете), а не о морфологии. Впервые они были описаны Уильямом Баклендом в 1829 году. До этого они были известны как «ископаемые еловые шишки » и « безоаровые камни». Они служат ценной цели в палеонтологии, поскольку предоставляют прямые доказательства хищничества и питания вымерших организмов. [93] Размер копролитов может варьироваться от нескольких миллиметров до более 60 сантиметров.
-

-
 Копролит хищного динозавра, найденный на юго-западе Саскачевана.
Копролит хищного динозавра, найденный на юго-западе Саскачевана. -
 Плотно упакованные субаэральные или прибрежные тропы ( Climactichnites wilsoni ), оставленные предполагаемым слизнеобразным моллюском на кембрийской приливной равнине.
Плотно упакованные субаэральные или прибрежные тропы ( Climactichnites wilsoni ), оставленные предполагаемым слизнеобразным моллюском на кембрийской приливной равнине.



Переходный
— Переходное ископаемое это любые окаменелые остатки формы жизни, которая демонстрирует черты, общие как для группы предков, так и для группы ее производных потомков. [94] Это особенно важно там, где группа потомков резко отличается по грубой анатомии и образу жизни от группы предков. Из-за неполноты летописи окаменелостей обычно невозможно точно узнать, насколько близко переходное ископаемое находится к точке дивергенции. Эти окаменелости служат напоминанием о том, что таксономические подразделения — это человеческие конструкции, которые задним числом были навязаны континууму вариаций.
Микрофоссилии

Микроископаемые — это описательный термин, применяемый к окаменевшим растениям и животным, размер которых находится на уровне, на котором окаменелость можно проанализировать невооруженным глазом, или ниже него. Обычно применяемая граница между «микро» и «макро» окаменелостями составляет 1 мм. Микрофоссилии могут быть либо полными (или почти полными) организмами сами по себе (например, морские планктеры, фораминиферы и кокколитофоры ), либо составными частями (например, маленькие зубы или споры ) более крупных животных или растений. Микрофоссилии имеют решающее значение как резервуар палеоклиматической информации, а также обычно используются биостратиграфами для корреляции горных пород.
Смола

Ископаемая смола (в просторечии называемая янтарем ) — природный полимер, встречающийся во многих типах пластов по всему миру, даже в Арктике . Самая старая ископаемая смола датируется триасом , хотя большинство из них относится к кайнозою . Считается, что выделение смолы некоторыми растениями является эволюционной адаптацией для защиты от насекомых и заживления ран. Ископаемая смола часто содержит другие окаменелости, называемые включениями, которые были захвачены липкой смолой. К ним относятся бактерии, грибы, другие растения и животные. Животные включения обычно представляют собой мелких беспозвоночных , преимущественно членистоногих, таких как насекомые и пауки, и лишь крайне редко позвоночных, таких как небольшая ящерица. Сохранность включений может быть превосходной, включая небольшие фрагменты ДНК .
Производный или переработанный

Производное . , переработанное или реманиозное ископаемое — это ископаемое, найденное в горной породе, которое накопилось значительно позже, чем когда окаменелое животное или растение умерло [95] Переработанные окаменелости создаются в результате эрозионной эксгумации (освобождения) окаменелостей из горной породы, в которой они первоначально отложились, и их повторного отложения в более молодых осадочных отложениях.
Древесина


Ископаемая древесина – это древесина, сохранившаяся в летописи окаменелостей. Древесина обычно является той частью растения, которая лучше всего сохраняется (и ее легче всего найти). Ископаемая древесина может быть окаменелой , а может и не быть . Ископаемая древесина, возможно, единственная сохранившаяся часть растения; [96] поэтому такая древесина может получить особое ботаническое название . Обычно это включает в себя слово «ксилон» и термин, указывающий на его предполагаемое родство, например, араукариоксилон (древесина араукарии или какого-либо родственного рода), пальмоксилон (древесина неопределенной пальмы ) или кастаноксилон (древесина неопределенного чинкапина ). [97]
субископаемое

Термин «субокаменелость» может использоваться для обозначения останков, таких как кости, гнезда или отложения фекалий , процесс окаменения которых не завершен либо потому, что продолжительность жизни животного слишком коротка, либо потому, что условия, в которых Захороненные останки не были оптимальными для окаменения. [98] Субфоссилии часто находят в пещерах или других убежищах, где они могут сохраняться тысячи лет. [99] Основное значение субископаемых останков по сравнению с ископаемыми останками заключается в том, что первые содержат органический материал, который можно использовать для радиоуглеродного датирования или экстракции и секвенирования ДНК , белка или других биомолекул. Кроме того, соотношения изотопов могут предоставить много информации об экологических условиях, в которых жили вымершие животные. Субфоссилии полезны для изучения истории эволюции окружающей среды и могут иметь важное значение для исследований в области палеоклиматологии .
Субфоссилии часто встречаются в отложениях, таких как отложения озер, океанические отложения и почвы. После отложения физическое и химическое выветривание может изменить состояние сохранности, а небольшие субфоссилии также могут быть проглочены живыми организмами . Субфоссильные останки, относящиеся к мезозойскому периоду , исключительно редки, обычно находятся на поздней стадии разложения и, следовательно, являются предметом многих споров. [100] Подавляющая часть субископаемого материала происходит из четвертичных отложений, включая множество субокаменевших головных капсул хирономид , остракод панцирей , диатомей и фораминифер .

Для останков, таких как морские ракушки моллюсков , которые часто не меняют свой химический состав в течение геологического времени, а иногда даже могут сохранять такие особенности, как исходная цветовая маркировка, в течение миллионов лет, ярлык «субокаменелости» применяется к раковинам, которые считаются им тысячи лет, но они относятся к голоценовому возрасту и, следовательно, недостаточно стары, чтобы принадлежать к эпохе плейстоцена . [101]
Химические окаменелости
Химические окаменелости, или хемофоссилии, — это химические вещества, обнаруженные в горных породах и ископаемом топливе (нефти, угле и природном газе), которые обеспечивают органический признак древней жизни. Молекулярные окаменелости и соотношения изотопов представляют собой два типа химических окаменелостей. [102] Самыми старыми следами жизни на Земле являются окаменелости этого типа, включая аномалии изотопов углерода, обнаруженные в цирконах , которые предполагают существование жизни еще 4,1 миллиарда лет назад. [10] [11]
Строматолиты

Строматолиты — слоистые аккреционные структуры, на мелководье путем захвата, связывания и цементации осадочных зерен биопленками микроорганизмов образующиеся , особенно цианобактерий . [103] Строматолиты представляют собой одни из самых древних ископаемых свидетельств жизни на Земле, датируемые более 3,5 миллиардов лет назад. [104]
В докембрийское время строматолитов было гораздо больше. В то время как более древние архейские ископаемые останки предположительно представляют собой колонии цианобактерий первичными , более молодые (то есть протерозойские ) окаменелости могут быть формами эукариотных хлорофитов ( то есть зеленых водорослей ). Одним из родов строматолитов, очень распространенных в геологических летописях, является Collenia . Самый ранний строматолит подтвержденного микробного происхождения датируется 2,724 миллиарда лет назад. [105]
Открытие 2009 года предоставляет убедительные доказательства того, что микробные строматолиты существовали еще 3,45 миллиарда лет назад. [106] [107]
Строматолиты являются основным компонентом летописи окаменелостей в течение первых 3,5 миллиардов лет жизни, достигнув пика около 1,25 миллиарда лет назад. [106] Впоследствии их численность и разнообразие сократились. [108] который к началу кембрия упал до 20% от своего пика. Наиболее широко поддерживаемое объяснение состоит в том, что строители строматолитов стали жертвами пасущихся существ ( кембрийская революция субстрата ), подразумевая, что достаточно сложные организмы были обычным явлением более 1 миллиарда лет назад. [109] [110] [111]
Связь между численностью травоядных и строматолитов хорошо документирована в молодого ордовика эволюционной радиации ; Численность строматолитов также увеличилась после того, как вымирания в конце ордовика и в конце перми уничтожили морских животных, и упало до более раннего уровня по мере восстановления морских животных. [112] Колебания численности и разнообразия многоклеточных животных , возможно, были не единственным фактором сокращения численности строматолитов. Причиной изменений могли быть такие факторы, как химия окружающей среды. [113]
Хотя сами прокариотические цианобактерии размножаются бесполым путем посредством деления клеток, они сыграли важную роль в подготовке среды для эволюционного развития более сложных эукариотических организмов. цианобактерии (а также экстремофильные гаммапротеобактерии Считается, что ) в значительной степени ответственны за увеличение количества кислорода Земли в первичной атмосфере посредством продолжающегося фотосинтеза . Цианобактерии используют воду , углекислый газ и солнечный свет для создания пищи. Слой слизи часто образуется поверх матов цианобактериальных клеток. В современных микробных матах мусор из окружающей среды обитания может задерживаться внутри слизи, которая может цементироваться карбонатом кальция, образуя тонкие пластинки известняка . Эти пластинки могут со временем срастаться, в результате чего образуется полосатый рисунок, характерный для строматолитов. Купольная морфология биологических строматолитов является результатом вертикального роста, необходимого для непрерывного проникновения солнечного света в организмы для фотосинтеза. Слоистые сферические структуры роста, называемые онколиты подобны строматолитам и также известны из летописи окаменелостей . Тромболиты представляют собой плохо слоистые или неслоистые сгустковые структуры, образованные цианобактериями, распространенными в летописи окаменелостей и в современных отложениях. [105]
Район каньона реки Зебра платформы Кубис в глубоко расчлененных горах Зарис на юго-западе Намибии представляет собой чрезвычайно хорошо обнаженный пример тромболит-строматолит-метазойных рифов, которые развивались в протерозойский период, причем строматолиты здесь лучше развиты в местах, расположенных вверх по падению, в условиях более высоких скоростей течений и большего притока наносов. [114]
Псевдоокаменелости

Псевдоокаменелости — это визуальные узоры в горных породах, возникшие в результате геологических, а не биологических процессов. Их легко принять за настоящие окаменелости. Некоторые псевдоокаменелости, такие как геологические дендритные кристаллы, образуются в результате естественных трещин в породе, которые заполняются просачивающимися минералами. Другими типами псевдоокаменелостей являются почки (круглые формы в железной руде) и моховые агаты , которые выглядят как мох или листья растений. Конкременты , сферические или овоидные конкреции, обнаруженные в некоторых осадочных слоях, когда-то считались яйцами динозавров , и их часто принимают за окаменелости.
Астробиология
Было высказано предположение, что биоминералы могут быть важными индикаторами внеземной жизни и, таким образом, могут играть важную роль в поисках прошлой или настоящей жизни на планете Марс . Кроме того, считается, что органические компоненты ( биосигнатуры ), которые часто связаны с биоминералами, играют решающую роль как в пребиотических, так и в биотических реакциях. [115]
24 января 2014 года НАСА сообщило, что текущие исследования Curiosity и Opportunity марсоходов на Марсе теперь будут направлены на поиск доказательств древней жизни, включая биосферу, основанную на автотрофных , хемотрофных и/или хемолитоавтотрофных микроорганизмах , а также древнюю воду, в том числе речную. -озерные среды ( равнины, связанные с древними реками или озерами ), которые могли быть пригодными для жизни . [116] [117] [118] [119] Поиск доказательств обитаемости , тафономии (связанной с окаменелостями) и органического углерода на планете Марс теперь является основной целью НАСА . [116] [117]
Искусство
Согласно одной из гипотез, коринфская ваза VI века до нашей эры является старейшим художественным свидетельством окаменелостей позвоночных, возможно, миоценового жирафа, в сочетании с элементами других видов. [120] Однако последующее исследование с использованием искусственного интеллекта и экспертные оценки опровергли эту идею, поскольку у млекопитающих нет глазных костей, показанных у нарисованного монстра. Морфологически роспись на вазе соответствует хищной рептилии семейства Varanidae, которая до сих пор обитает в регионах, занимаемых древнегреческой территорией. [121]
Торговля и коллекционирование
Торговля ископаемыми – это практика покупки и продажи ископаемых. Часто это делается незаконно с артефактами, украденными из исследовательских центров, что ежегодно приводит к потере многих важных научных образцов. [122] Проблема весьма выражена в Китае, где было украдено много экземпляров. [123]
Коллекционирование окаменелостей (иногда, в ненаучном смысле, охота за окаменелостями) — это сбор окаменелостей для научных исследований, хобби или получения прибыли. Сбор окаменелостей, практикуемый любителями, является предшественником современной палеонтологии, и многие до сих пор собирают окаменелости и изучают их как любители. Как профессионалы, так и любители собирают окаменелости ради их научной ценности.
Как лекарство
Использование окаменелостей для решения проблем со здоровьем уходит корнями в традиционную медицину и включает использование окаменелостей в качестве талисманов . Конкретное ископаемое, которое можно использовать для облегчения или лечения болезни, часто основано на его сходстве с симптомами или пораженным органом. Полезность окаменелостей в качестве лекарства почти полностью является эффектом плацебо , хотя ископаемый материал, возможно, может обладать некоторой антацидной активностью или поставлять некоторые необходимые минералы . [124] Использование костей динозавров в качестве «костей дракона» сохранилось в традиционной китайской медицине до наших дней, при этом кости динозавров середины мелового периода использовались для этой цели в округе Руян в начале 21 века. [125]
Галерея
-
 Морские окаменелости, найденные высоко в Гималаях. Коллекция настоятеля Дханкара Гомпы , HP, Индия
Морские окаменелости, найденные высоко в Гималаях. Коллекция настоятеля Дханкара Гомпы , HP, Индия -
 Три небольших окаменелости аммонита , каждая примерно 1,5 см в поперечнике.
Три небольших окаменелости аммонита , каждая примерно 1,5 см в поперечнике. -
 Эоценовая ископаемая рыба Priscacara liops из формации Грин-Ривер в Вайоминге
Эоценовая ископаемая рыба Priscacara liops из формации Грин-Ривер в Вайоминге -
 Перминерализованный трилобит . Asaphus kowalewskie
Перминерализованный трилобит . Asaphus kowalewskie -

-
 Ископаемые креветки ( меловой период )
Ископаемые креветки ( меловой период ) -
 Окаменелое дерево в национальном парке Петрифайд-Форест , Аризона.
Окаменелое дерево в национальном парке Петрифайд-Форест , Аризона. -
 Окаменелый конус араукарии мирабилис из Патагонии , Аргентина, датируемый юрским периодом (около 210 млн лет назад )
Окаменелый конус араукарии мирабилис из Патагонии , Аргентина, датируемый юрским периодом (около 210 млн лет назад ) -

-
 силурийского Orthoceras Ископаемое
силурийского Orthoceras Ископаемое -
 Ископаемый цветок эоцена из Флориссана, Колорадо.
Ископаемый цветок эоцена из Флориссана, Колорадо. -
 ежа Окаменелость морского из Англии
ежа Окаменелость морского из Англии -

-
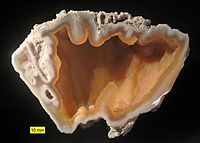 Агатизированный коралл из группы Боярышника ( олигоцен - миоцен ), Флорида . Пример сохранения путем замены.
Агатизированный коралл из группы Боярышника ( олигоцен - миоцен ), Флорида . Пример сохранения путем замены. -
 Окаменелости с пляжей в Балтийском море острова Готланд , помещенные на бумагу с квадратами размером 7 мм (0,28 дюйма).
Окаменелости с пляжей в Балтийском море острова Готланд , помещенные на бумагу с квадратами размером 7 мм (0,28 дюйма). -
 Следы динозавров из национального парка Тороторо в Боливии.
Следы динозавров из национального парка Тороторо в Боливии.
















См. также
- Биоэрозия - эрозия твердых субстратов живыми организмами.
- Криптоспора - окаменевшая спора примитивного растения.
- Эндолит – организм, живущий внутри камня.
- Список парков ископаемых
- Живое ископаемое - организм, напоминающий форму, давно представленную в летописи окаменелостей.
- Палеобиология - изучение органической эволюции с использованием окаменелостей.
- Палеоботаника - Исследование органической эволюции растений на основе окаменелостей.
- Правило Шульца - Связь между износом зубов и продолжительностью жизни ископаемых организмов.
- Зуб акулы – Зубы акулы
- Эффект Синьора-Липпса - систематическая ошибка выборки в летописи окаменелостей затрудняет характеристику вымираний.
Ссылки
- ^ Оксфордский словарь английского языка . Издательство Оксфордского университета. Архивировано из оригинала 11 января 2008 года . Проверено 17 июня 2013 г.
- ^ Яблонски, Дэвид; Рой, Каустув; Валентин, Джеймс В.; Прайс, Ребекка М.; Андерсон, Филип С. (16 мая 2003 г.). «Влияние недавнего времени на историю морского разнообразия» . Наука . 300 (5622): 1133–1135. Бибкод : 2003Sci...300.1133J . дои : 10.1126/science.1083246 . ISSN 1095-9203 . ПМИД 12750517 . S2CID 42468747 . Архивировано из оригинала 15 декабря 2022 года . Проверено 15 декабря 2022 г.
- ^ Сахни, Сарда; Бентон, Майкл Дж.; Ферри, Пол А. (23 августа 2010 г.). «Связь между глобальным таксономическим разнообразием, экологическим разнообразием и распространением позвоночных на суше» . Письма по биологии . 6 (4): 544–547. дои : 10.1098/rsbl.2009.1024 . ПМК 2936204 . ПМИД 20106856 .
- ^ Сахни, Сарда; Бентон, Майкл (2017). «Влияние Притяжения Недавнего времени на летопись окаменелостей четвероногих» (PDF) . Исследования в области эволюционной экологии . 18 :7–23. Архивировано (PDF) из оригинала 15 декабря 2022 года . Проверено 15 декабря 2022 г.
- ^ Эдвард Б. Дэшлер, Нил Х. Шубин и Фариш А. Дженкинс-младший (6 апреля 2006 г.). «Рыба, похожая на девонских четвероногих, и эволюция строения тела четвероногих» (PDF) . Природа . 440 (7085): 757–763. Бибкод : 2006Natur.440..757D . дои : 10.1038/nature04639 . ПМИД 16598249 . Архивировано (PDF) из оригинала 15 декабря 2022 года . Проверено 15 декабря 2022 г.
- ^ «theNAT :: Музей естественной истории Сан-Диего :: Ваша связь с природой в парке Бальбоа :: Часто задаваемые вопросы» . Sdnhm.org. Архивировано из оригинала 10 мая 2012 года . Проверено 5 ноября 2012 г.
- ^ Боренштейн, Сет (13 ноября 2013 г.). «Найдена самая старая окаменелость: познакомьтесь со своей микробной мамой» . Ассошиэйтед Пресс. Архивировано из оригинала 29 июня 2015 года . Проверено 15 ноября 2013 г.
- ^ Ноффке, Нора ; Кристиан, Дэниел; Уэйси, Дэвид; Хейзен, Роберт М. (8 ноября 2013 г.). «Микробно-индуцированные осадочные структуры, фиксирующие древнюю экосистему в формации Дрессер возрастом около 3,48 миллиарда лет, Пилбара, Западная Австралия» . Астробиология . 13 (12): 1103–24. Бибкод : 2013AsBio..13.1103N . дои : 10.1089/ast.2013.1030 . ПМК 3870916 . ПМИД 24205812 .
- ^ Брайан Вастаг (21 августа 2011 г.). «Старейшие «микроокаменелости» вселяют надежду на жизнь на Марсе» . Вашингтон Пост . Архивировано из оригинала 19 октября 2011 года . Проверено 21 августа 2011 г.
Уэйд, Николас (21 августа 2011 г.). «Геологическая группа претендует на древнейшие известные окаменелости» . Нью-Йорк Таймс . Архивировано из оригинала 1 мая 2013 года . Проверено 21 августа 2011 г. - ^ Jump up to: а б Боренштейн, Сет (19 октября 2015 г.). «Намеки на жизнь на ранней Земле, которая считалась пустынной» . Возбуждайте . Йонкерс, Нью-Йорк: Интерактивная сеть Mindspark . Ассошиэйтед Пресс . Архивировано из оригинала 23 октября 2015 года . Проверено 20 октября 2015 г.
- ^ Jump up to: а б Белл, Элизабет А.; Бенике, Патрик; Харрисон, Т. Марк; и др. (19 октября 2015 г.). «Потенциально биогенный углерод сохранился в цирконе возрастом 4,1 миллиарда лет» (PDF) . Учеб. Натл. акад. наук. США . 112 (47): 14518–21. Бибкод : 2015PNAS..11214518B . дои : 10.1073/pnas.1517557112 . ISSN 1091-6490 . ПМЦ 4664351 . ПМИД 26483481 . Архивировано (PDF) из оригинала 6 ноября 2015 года . Проверено 20 октября 2015 г. Раннее издание, опубликованное в Интернете до печати.
- ^ Уэстолл, Фрэнсис; и др. (2001). «Раннеархейские ископаемые бактерии и биопленки в отложениях зеленокаменного пояса Барбертон, Южная Африка, подвергшихся гидротермальному влиянию». Докембрийские исследования . 106 (1–2): 93–116. Бибкод : 2001PreR..106...93W . дои : 10.1016/S0301-9268(00)00127-3 .
- ^ Jump up to: а б с «Коллекционеры доисторических окаменелостей» . Архивировано из оригинала 17 февраля 2019 года . Проверено 16 февраля 2019 г.
- ^ «Древние египтяне собирали окаменелости» . 5 сентября 2016 г. Архивировано из оригинала 10 февраля 2019 г. . Проверено 9 февраля 2019 г.
- ^ Аристотель (1931) [350 г. до н.э. ]. «Книга III часть 6» . Метеорология . Перевод Э. У. Вебстера. Архивировано из оригинала 18 февраля 2014 года . Проверено 20 февраля 2023 г. - из Архива интернет-классики.
- ^ Радвик, MJS (1985). Значение окаменелостей: эпизоды истории палеонтологии . Издательство Чикагского университета . п. 24. ISBN 978-0-226-73103-2 . Архивировано из оригинала 17 марта 2023 года . Проверено 11 октября 2018 г.
- ^ Jump up to: а б с «Хрящевая рыба» . Архивировано из оригинала 30 июля 2017 года . Проверено 16 февраля 2019 г.
- ^ «Ссылки на окаменелости Плиния Старшего» . Архивировано из оригинала 2 января 2019 года . Проверено 16 февраля 2019 г.
- ^ Мэр Адриенн (24 октября 2013 г.). Ископаемые легенды первых американцев . Издательство Принстонского университета. ISBN 978-1-4008-4931-4 . Архивировано из оригинала 17 марта 2023 года . Проверено 18 октября 2019 г. - через Google Книги.
- ^ «Откуда мы знаем, что древние африканские люди ценили окаменелости и камни» . 29 января 2019 года. Архивировано из оригинала 10 февраля 2019 года . Проверено 9 февраля 2019 г.
- ^ «Окаменелость возрастом 400 миллионов лет появилась на выставке вместе со стихотворением Хуан Тинцзяня» People's Daily Net (на традиционном китайском языке, 17 мая 2013 г.). Архивировано из оригинала 12 июня 2018 г. Проверено 7 июня 2018 г.
- ^ Сивин, Натан (1995). Наука в Древнем Китае: исследования и размышления . Брукфилд, Вермонт: VARIORUM, Ashgate Publishing. III, с. 23
- ^ Jump up to: а б Нидэм, Джозеф. (1959). Наука и цивилизация в Китае: Том 3, Математика и науки о небе и Земле . Издательство Кембриджского университета . стр. 603–618.
- ^ Чан, Алан Кам-люн и Грегори К. Кланси, Хуэй-Чье Лой (2002). Исторические перспективы восточноазиатской науки, технологий и медицины . Сингапур: Издательство Сингапурского университета . п. 15. ISBN 9971-69-259-7 .
- ^ Рафферти, Джон П. (2012). геологические науки; Геология: формы рельефа, минералы и горные породы . Нью-Йорк: Образовательное издательство Britannica, стр. 6. ISBN 9781615305445
- ^ Десмонд, Адриан. «Открытие морских нарушений и объяснение ископаемых в древности», American Journal of Science , 1975, том 275: 692–707.
- ^ Рафферти, Джон П. (2012). геологические науки; Геология: формы рельефа, минералы и горные породы . Нью-Йорк: Образовательное издательство Britannica, стр. 5–6. ISBN 9781615305445 .
- ^ Алистер Кэмерон Кромби (1990). Наука, оптика и музыка в средневековой и ранней современной мысли . Международная издательская группа «Континуум». стр. 108–109. ISBN 978-0-907628-79-8 . Архивировано из оригинала 17 марта 2023 года . Проверено 11 октября 2018 г.
- ^ «Миф о Циклопе, вызванный «одноглазыми» окаменелостями?» . Национальное географическое общество . 5 февраля 2003 г. Архивировано из оригинала 17 февраля 2019 г. Проверено 16 февраля 2019 г.
- ^ «8 типов воображаемых существ, «обнаруженных» в окаменелостях» . 19 мая 2015 г. Архивировано из оригинала 16 февраля 2019 г. Проверено 16 февраля 2019 г.
- ^ «Фольклор ископаемых иглокожих» . 4 апреля 2017 года. Архивировано из оригинала 17 февраля 2019 года . Проверено 16 февраля 2019 г.
- ^ Макнамара, Кеннет Дж. (2007). «Пастучьи короны, сказочные хлебы и громовые камни: мифология ископаемых ежей в Англии» . Геологическое общество, Лондон, специальные публикации . 273 (1): 279–294. Бибкод : 2007GSLSP.273..279M . дои : 10.1144/ГСЛ.СП.2007.273.01.22 . S2CID 129384807 . Архивировано из оригинала 21 февраля 2019 года . Проверено 16 февраля 2019 г.
- ^ «Археологические иглокожие! Сказочные хлебы и громовые камни!» . 12 января 2009 г. Архивировано из оригинала 17 февраля 2019 г. . Проверено 16 февраля 2019 г.
- ^ Бокон, Андреа (2010). «Леонардо да Винчи, отец-основатель ихногии». ПАЛЕОС . 25 (5/6). Общество осадочной геологии SEPM: 361–367. Бибкод : 2010Палай..25..361B . дои : 10.2110/palo.2009.p09-049r . JSTOR 40606506 . S2CID 86011122 .
- ^ да Винчи, Леонардо (1956) [1938]. Записные книжки Леонардо да Винчи . Лондон: Рейнал и Хичкок. п. 335. ИСБН 978-0-9737837-3-5 . [ постоянная мертвая ссылка ]
- ^ Брессан, Дэвид. «18 июля 1635 года: Роберт Гук – последний виртуоз глупой науки» . Сеть блогов Scientific American . Архивировано из оригинала 12 февраля 2018 года . Проверено 11 февраля 2018 г.
- ^ «Кювье» . palaeo.gly.bris.ac.uk . Архивировано из оригинала 25 мая 2014 года . Проверено 3 ноября 2008 г.
- ^ «Мэри Эннинг» . Музей Лайм-Реджиса . Архивировано из оригинала 22 августа 2018 года . Проверено 21 августа 2018 г.
- ^ Дарвин, Чарльз (1872), « Глава X: О несовершенстве геологических данных », «Происхождение видов » , Лондон: Джон Мюррей.
- ^ Schopf JW (1999) Колыбель жизни: открытие самых ранних окаменелостей Земли, Princeton University Press, Принстон, Нью-Джерси.
- ^ «Виртуальный музей окаменелостей - окаменелости в геологическом времени и эволюции» . Архивировано из оригинала 8 марта 2007 года . Проверено 4 марта 2007 г.
- ^ Нолл, А. (2003) Жизнь на молодой планете. (Издательство Принстонского университета, Принстон, Нью-Джерси)
- ^ Донован, СК; Пол, CRC, ред. (1998). «Обзор полноты летописи окаменелостей». Адекватность летописи окаменелостей . Нью-Йорк: Уайли. стр. 111–131. ISBN 978-0-471-96988-4 .
- ^ Форти, Ричард , Трилобит!: Свидетель эволюции . Альфред А. Кнопф, Нью-Йорк, 2000 г.
- ^ Донохью, PCJ; Бенгтсон, С; Донг, Х; Гостлинг, Нью-Джерси; Хульдтгрен, Т; Каннингем, Дж.А.; Инь, С; Юэ, З; Пэн, Ф; и др. (2006). «Синхротронная рентгеновская томографическая микроскопия ископаемых эмбрионов». Природа . 442 (7103): 680–683. Бибкод : 2006Natur.442..680D . дои : 10.1038/nature04890 . ПМИД 16900198 . S2CID 4411929 .
- ^ Фут, М.; Сепкоски, Джей-Джей-младший (1999). «Абсолютные меры полноты летописи окаменелостей». Природа . 398 (6726): 415–417. Бибкод : 1999Natur.398..415F . дои : 10.1038/18872 . ПМИД 11536900 . S2CID 4323702 .
- ^ Бентон, М. (2009). «Полнота летописи окаменелостей» . Значение . 6 (3): 117–121. дои : 10.1111/j.1740-9713.2009.00374.x . S2CID 84441170 .
- ^ Жлиобайте, И.; Фортелиус, М. (2021). «О калибровке полного счетчика ископаемых млекопитающих» . Палеобиология . 48 : 1–11. дои : 10.1017/pab.2021.22 . S2CID 238686414 .
- ^ Эйтинг, Т.П.; Ганнелл, Г.Г. (2009). «Глобальная полнота летописи окаменелостей летучих мышей». Журнал эволюции млекопитающих . 16 (3): 151–173. дои : 10.1007/s10914-009-9118-x . S2CID 5923450 .
- ^ Броклхерст, Н.; Апчерч, П.; Мэннион, Полиция; О'Коннор, Дж. (2012). «Полнота летописи окаменелостей мезозойских птиц: значение для ранней эволюции птиц» . ПЛОС ОДИН . 7 (6): e39056. Бибкод : 2012PLoSO...739056B . дои : 10.1371/journal.pone.0039056 . ПМЦ 3382576 . ПМИД 22761723 .
- ^ Реталлак, Г. (1984). «Полнота летописи горных пород и окаменелостей: некоторые оценки с использованием ископаемых почв». Палеобиология . 10 (1): 59–78. Бибкод : 1984Pbio...10...59R . дои : 10.1017/S0094837300008022 . S2CID 140168970 .
- ^ Бентон, MJ; Сторрс, Г.Вм. (1994). «Проверка качества летописи окаменелостей: палеонтологические знания улучшаются». Геология . 22 (2): 111–114. Бибкод : 1994Geo....22..111B . doi : 10.1130/0091-7613(1994)022<0111:TTQOTF>2.3.CO;2 .
- ^ Голландия, СМ; Пацковски, М.Э. (1999). «Модели для моделирования летописи окаменелостей». Геология . 27 (6): 491–494. Бибкод : 1999Geo....27..491H . doi : 10.1130/0091-7613(1999)027<0491:MFSTFR>2.3.CO;2 .
- ^ Кох, К. (1978). «Предвзятость в опубликованных летописях окаменелостей». Палеобиология . 4 (3): 367–372. Бибкод : 1978Pbio....4..367K . дои : 10.1017/S0094837300006060 . S2CID 87368101 .
- ^ Синьор, военнопленный III; Липпс, Дж. Х. (1982). «Ошибка выборки, закономерности постепенного вымирания и катастрофы в летописи окаменелостей». В Серебре, LT; Шульц, PH (ред.). Геологические последствия ударов крупных астероидов и комет о Землю . Специальные статьи Геологического общества Америки. Том. 190. стр. 291–296. дои : 10.1130/SPE190-p291 . ISBN 0-8137-2190-3 .
- ^ Вилена, Д.А.; Смит, AB (2013). «Пространственное смещение в записях морских окаменелостей» . ПЛОС ОДИН . 8 (10): е74470. Бибкод : 2013PLoSO...874470V . дои : 10.1371/journal.pone.0074470 . ПМЦ 3813679 . ПМИД 24204570 .
- ^ Jump up to: а б Мартин, МВт; Гражданкин Д.В.; Боуринг, ЮАР; Эванс, ПАПА; Федонкин, М.А.; Киршвинк, Дж. Л. (5 мая 2000 г.). «Возраст неопротерозойского двулатерального тела и следы окаменелостей, Белое море, Россия: значение для эволюции многоклеточных животных». Наука . 288 (5467): 841–5. Бибкод : 2000Sci...288..841M . дои : 10.1126/science.288.5467.841 . ПМИД 10797002 . S2CID 1019572 .
- ^ Пуфаль, ПК; Гримм, К.А.; Абед, AM и Садака, RMY (октябрь 2003 г.). «Верхнемеловые (кампанские) фосфориты в Иордании: значение для формирования южно-тетического фосфоритового гиганта». Осадочная геология . 161 (3–4): 175–205. Бибкод : 2003SedG..161..175P . дои : 10.1016/S0037-0738(03)00070-8 .
- ^ «Геологическое время: радиометрическая шкала времени» . Геологическая служба США. Архивировано из оригинала 21 сентября 2008 года . Проверено 20 сентября 2008 г.
- ^ Лёфгрен, А. (2004). «Фауна конодонтов в среднеордовикской зоне Eoplacognathus pseudoplanus Балтоскандии». Геологический журнал . 141 (4): 505–524. Бибкод : 2004GeoM..141..505L . дои : 10.1017/S0016756804009227 . S2CID 129600604 .
- ^ Jump up to: а б Гелинг, Джеймс; Йенсен, Сорен; Дрозер, Мэри; Мироу, Пол; Нарбонн, Гай (март 2001 г.). «Роясь под базальным кембрийским GSSP, Форчун-Хед, Ньюфаундленд». Геологический журнал . 138 (2): 213–218. Бибкод : 2001ГеоМ..138..213Г . дои : 10.1017/S001675680100509X . S2CID 131211543 .
- ^ Хаг, Лос-Анджелес; Роджер, Эй Джей (2007). «Влияние отбора проб окаменелостей и таксонов на анализ древнего молекулярного датирования» . Молекулярная биология и эволюция . 24 (8): 889–1897. дои : 10.1093/molbev/msm115 . ПМИД 17556757 .
- ^ Петерсон, Кевин Дж.; Баттерфилд, Нью-Джерси (2005). «Происхождение Eumetazoa: проверка экологических предсказаний молекулярных часов на основе летописи окаменелостей протерозоя» . Труды Национальной академии наук . 102 (27): 9547–52. Бибкод : 2005PNAS..102.9547P . дои : 10.1073/pnas.0503660102 . ПМЦ 1172262 . ПМИД 15983372 .
- ^ Jump up to: а б Протеро, Дональд Р. (2007). Эволюция: о чем говорят окаменелости и почему это важно . Издательство Колумбийского университета . стр. 50–53 . ISBN 978-0-231-51142-1 .
- ^ Исаак, М. (5 ноября 2006 г.). «Заявление CC200: переходных окаменелостей нет» . Архив TalkOrigins . Архивировано из оригинала 27 февраля 2009 года . Проверено 30 апреля 2009 г.
- ^ Донован, СК; Пол, CRC, ред. (1998). Адекватность летописи окаменелостей . Нью-Йорк: Уайли. п. 312. ИСБН 978-0-471-96988-4 . [ нужна страница ]
- ^ Протеро 2013 , стр. 8–9.
- ^ Протеро, Дональд Р. (2013). Оживление окаменелостей: введение в палеобиологию (Третье изд.). Нью-Йорк: Издательство Колумбийского университета. п. 8. ISBN 978-0-231-15893-0 .
- ^ Протеро 2013 , стр. 12–13.
- ^ Протеро 2013 , с. 16.
- ^ Уилби, П.; Бриггс, Д. (1997). «Таксономические тенденции в разрешении деталей, сохранившихся в ископаемых фосфатированных мягких тканях». Геобиос . 30 : 493–502. Бибкод : 1997Geobi..30..493W . дои : 10.1016/S0016-6995(97)80056-3 .
- ^ Уэйси, Д. и др. (2013) Наномасштабный анализ пиритизированных микрокаменелостей показывает дифференциальное гетеротрофное потребление в кремне Ганфлинта ~ 1,9 млрд лет PNAS 110 (20) 8020-8024 два : 10.1073/pnas.1221965110
- ^ Рэйсуэлл, Р. (1997). Геохимическая основа применения стабильных изотопов серы для пиритизации ископаемых. Журнал Геологического общества 154, 343–356.
- ^ Олер, Джон Х. и Шопф, Дж. Уильям (1971). Искусственные микрофоссилии: экспериментальные исследования перминерализации сине-зеленых водорослей в кремнеземе. Наука . 174, 1229–1231.
- ^ Гетц, Аннет Э.; Монтенари, Майкл; Костин, Гелу (2017). «Кремнение и сохранение органического вещества в анисовом Мусшелкалке: последствия для динамики бассейна центральноевропейского моря Мушелкалк» . Центральноевропейская геология . 60 (1): 35–52. Бибкод : 2017CEJGl..60...35G . дои : 10.1556/24.60.2017.002 . ISSN 1788-2281 .
- ^ Протеро 2013 , стр. 9–10.
- ^ «Определение Штейнкерна» . Мерриам-Вебстер . Архивировано из оригинала 13 мая 2021 года . Проверено 13 мая 2021 г.
ископаемое, состоящее из каменистой массы, попавшей в полый природный объект (например, раковину двустворчатого моллюска) в виде грязи или осадка, консолидировалось и осталось в виде слепка после растворения плесени
- ^ Протеро 2013 , с. 579.
- ^ Шут, Швейцария; Клил, CJ (1986). «Палеоботаника в музеях» . Геологический куратор . 4 (9): 553–559. дои : 10.55468/GC865 . S2CID 251638416 .
- ^ Брэдди, Саймон Дж.; Гасс, Кеннет К.; Тесслер, Майкл (4 сентября 2023 г.). «Не первая пиявка: необычный червь из раннего силура Висконсина» . Журнал палеонтологии . 97 (4): 799–804. Бибкод : 2023JPal...97..799B . дои : 10.1017/jpa.2023.47 . ISSN 0022-3360 . S2CID 261535626 .
- ^ Филдс Х (май 2006 г.). «Шокирующий динозавр. Исследуя тираннозавра возрастом 68 миллионов лет, Мэри Швейцер наткнулась на удивительные признаки жизни, которые могут радикально изменить наш взгляд на древних зверей» . Смитсоновский журнал . Архивировано из оригинала 18 января 2015 года.
- ^ Швейцер М.Х., Виттмайер Дж.Л., Хорнер Дж.Р., Топорски Дж.К. (25 марта 2005 г.). «Сосуды мягких тканей и сохранение клеток у тираннозавра рекса». Наука . 307 (5717): 1952–5. Бибкод : 2005Sci...307.1952S . дои : 10.1126/science.1108397 . ПМИД 15790853 . S2CID 30456613 .
- ^ Швейцер М.Х., Чжэн В., Клеланд Т.П., Берн М. (январь 2013 г.). «Молекулярный анализ остеоцитов динозавров подтверждает наличие эндогенных молекул». Кость . 52 (1): 414–23. дои : 10.1016/j.bone.2012.10.010 . ПМИД 23085295 .
- ^ Эмбери Дж., Милнер AC, Уоддингтон Р.Дж., Холл Р.К., Лэнгли М.Л., Милан AM (2003). «Идентификация белкового материала в кости динозавра игуанодона» . Исследование соединительной ткани . 44 (Приложение 1): 41–6. дои : 10.1080/03008200390152070 . ПМИД 12952172 . S2CID 2249126 .
- ^ Швейцер М.Х., Чжэн В., Клеланд Т.П., Гудвин М.Б., Ботман Э., Тейл Э., Маркус М.А., Факра СК (ноябрь 2013 г.). «Роль химии железа и кислорода в сохранении мягких тканей, клеток и молекул в глубоком прошлом» . Труды Королевского общества . 281 (1774): 20132741. doi : 10.1098/rspb.2013.2741 . ПМЦ 3866414 . ПМИД 24285202 .
- ^ Зильберберг, Л.; Лаурин, М. (2011). «Анализ органического матрикса ископаемых костей методом просвечивающей электронной микроскопии». Comptes Рендус Палевол . 11 (5–6): 357–366. дои : 10.1016/j.crpv.2011.04.004 .
- ^ Палмер, Ти Джей; Уилсон, Массачусетс (1988). «Паразитизм ордовикских мшанок и происхождение псевдоборингов». Палеонтология . 31 : 939–949.
- ^ Jump up to: а б Тейлор, П.Д. (1990). «Сохранение мягкотелых и других организмов путем биоиммурации: обзор». Палеонтология . 33 : 1–17.
- ^ Уилсон, Массачусетс; Палмер, Ти Джей; Тейлор, П.Д. (1994). «Самое раннее сохранение мягких окаменелостей путем биоиммурации эпибионтов: верхний ордовик Кентукки». Летайя . 27 (3): 269–270. Бибкод : 1994Letha..27..269W . дои : 10.1111/j.1502-3931.1994.tb01420.x .
- ^ «Что такое палеонтология?» . Музей палеонтологии Калифорнийского университета. Архивировано из оригинала 16 сентября 2008 года . Проверено 17 сентября 2008 г.
- ^ Jump up to: а б Федонкин, М.А.; Гелинг, Дж.Г.; Грей, К.; Нарбонн, генеральный директор; Викерс-Рич, П. (2007). Восстание животных: эволюция и разнообразие царства животных . Джу Пресс. стр. 213–216. ISBN 978-0-8018-8679-9 . Архивировано из оригинала 17 марта 2023 года . Проверено 14 ноября 2008 г.
- ^ например Сейлахер, А. (1994). «Насколько достоверна стратиграфия Круцианы?». Международный журнал наук о Земле . 83 (4): 752–758. Бибкод : 1994ГеоРу..83..752С . дои : 10.1007/BF00251073 . S2CID 129504434 .
- ^ «копролиты» . Словарь.com . Архивировано из оригинала 17 декабря 2008 года . Проверено 29 февраля 2012 г.
- ^ Херрон, Скотт; Фриман, Джон К. (2004). Эволюционный анализ (3-е изд.). Река Аппер-Седл, Нью-Джерси: Pearson Education. п. 816. ИСБН 978-0-13-101859-4 . Архивировано из оригинала 17 марта 2023 года . Проверено 11 октября 2018 г.
- ^ Нойендорф, Клаус К.Е.; Американский геологический институт (2005). Глоссарий геологии . Springer Science & Business Media. ISBN 978-0-922152-76-6 . Архивировано из оригинала 17 марта 2023 года . Проверено 7 июня 2020 г.
- ^ Эд Штраус (2001). «Окаменелый лес из Западного Вашингтона» . Архивировано из оригинала 11 декабря 2010 года . Проверено 8 апреля 2011 г.
- ^ Уилсон Николс Стюарт; Гар В. Ротвелл (1993). Палеоботаника и эволюция растений (2-е изд.). Издательство Кембриджского университета. п. 31. ISBN 978-0-521-38294-6 .
- ^ «Коллекции субокаменелостей» . Южно-Австралийский музей. Архивировано из оригинала 17 апреля 2021 года . Проверено 28 августа 2020 г.
- ^ «Коллекции субокаменелостей» . Южно-Австралийский музей. Архивировано из оригинала 17 июня 2011 года . Проверено 23 января 2014 г.
- ^ Петерсон, Джозеф Э.; Ленчевски, Мелисса Э.; Шерер, Рид П. (октябрь 2010 г.). Степанова, Анна (ред.). «Влияние микробных биопленок на сохранность первичных мягких тканей у ископаемых и современных архозавров» . ПЛОС ОДИН . 5 (10): 13А. Бибкод : 2010PLoSO...513334P . дои : 10.1371/journal.pone.0013334 . ПМЦ 2953520 . ПМИД 20967227 .
- ^ Ананд, Конкала (2022 г.). Зоология: распространение, эволюция и развитие животных . ИЗДАТЕЛЬСКИЙ ДОМ АГ. п. 42. ИСБН 9789395936293 .
- ^ «Химические или молекулярные окаменелости» . petrifiedwoodmuseum.org . Архивировано из оригинала 20 апреля 2014 года . Проверено 15 сентября 2013 г.
- ^ Верховая езда, Р. (2007). «Термин строматолит: к существенному определению» . Летайя . 32 (4): 321–330. дои : 10.1111/j.1502-3931.1999.tb00550.x . Архивировано из оригинала 2 мая 2015 года.
- ^ «Строматолиты, древнейшие окаменелости» . Архивировано из оригинала 9 марта 2007 года . Проверено 4 марта 2007 г.
- ^ Jump up to: а б Лепот, Кевин; Бензерара, Карим; Браун, Гордон Э.; Филиппо, Паскаль (2008). «Микробное влияние на образование строматолитов возрастом 2,7 миллиарда лет». Природа Геонауки . 1 (2): 118–21. Бибкод : 2008NatGe...1..118L . дои : 10.1038/ngeo107 .
- ^ Jump up to: а б Оллвуд, Эбигейл К.; Гротцингер, Джон П.; Нолл, Эндрю Х.; Берч, Ян В.; Андерсон, Марк С.; Коулман, Макс Л.; Каник, Исик (2009). «Контроль развития и разнообразия раннеархейских строматолитов» . Труды Национальной академии наук . 106 (24): 9548–9555. Бибкод : 2009PNAS..106.9548A . дои : 10.1073/pnas.0903323106 . ПМК 2700989 . ПМИД 19515817 .
- ^ Шопф, Дж. Уильям (1999). Колыбель жизни: открытие самых ранних окаменелостей Земли . Принстон, Нью-Джерси: Издательство Принстонского университета. стр. 87–89. ISBN 978-0-691-08864-8 . Архивировано из оригинала 17 марта 2023 года . Проверено 11 октября 2018 г.
- ^ Макменамин, MAS (1982). «Докембрийские конические строматолиты из Калифорнии и Соноры». Бюллетень Палеонтологического общества Южной Калифорнии . 14 (9 и 10): 103–105.
- ^ Макнамара, KJ (20 декабря 1996 г.). «Датирование происхождения животных» . Наука . 274 (5295): 1993–1997. Бибкод : 1996Sci...274.1993M . дои : 10.1126/science.274.5295.1993f .
- ^ Авраамик, С.М. (19 ноября 1971 г.). «Разнообразие столбчатых строматолитов докембрия: отражение внешнего вида многоклеточных животных». Наука . 174 (4011): 825–827. Бибкод : 1971Sci...174..825A . дои : 10.1126/science.174.4011.825 . ПМИД 17759393 . S2CID 2302113 .
- ^ Бенгтсон, С. (2002). «Происхождение и ранняя эволюция хищничества» (PDF) . В Ковалевском, М.; Келли, PH (ред.). Ископаемая летопись хищничества . Документы Палеонтологического общества . Том. 8. Палеонтологическое общество. стр. 289–317. Архивировано (PDF) из оригинала 10 сентября 2008 г. Проверено 29 декабря 2014 г.
- ^ Шиэн, премьер-министр; Харрис, Монтана (2004). «Возрождение микробиалита после вымирания в позднем ордовике». Природа . 430 (6995): 75–78. Бибкод : 2004Natur.430...75S . дои : 10.1038/nature02654 . ПМИД 15229600 . S2CID 4423149 .
- ^ Riding R (март 2006 г.). «Обилие микробных карбонатов по сравнению с колебаниями разнообразия многоклеточных животных в течение геологического времени» (PDF) . Осадочная геология . 185 (3–4): 229–38. Бибкод : 2006SedG..185..229R . дои : 10.1016/j.sedgeo.2005.12.015 . Архивировано из оригинала (PDF) 26 апреля 2012 года . Проверено 9 декабря 2011 г.
- ^ Адамс, EW; Гротцингер, JP; Уоттерс, Вашингтон; Шредер, С.; Маккормик, Д.С.; Аль-Сияби, ХА (2005). «Цифровая характеристика распределения тромболит-строматолитовых рифов в системе карбонатных пандусов (конечный протерозой, группа Нама, Намибия)» (PDF) . Бюллетень AAPG . 89 (10): 1293–1318. Бибкод : 2005BAAPG..89.1293A . дои : 10.1306/06160505005 . Архивировано из оригинала (PDF) 7 марта 2016 года . Проверено 8 декабря 2011 г.
- ^ Руководящая группа по научной лаборатории астробиологии MEPAG (26 сентября 2006 г.). «Итоговый отчет Руководящей группы научной группы астробиологической полевой лаборатории MEPAG (AFL-SSG)» (.doc) . В Стиле, Эндрю; Бити, Дэвид (ред.). Полевая лаборатория астробиологии . США: Группа анализа программы исследования Марса (MEPAG) – НАСА. п. 72. Архивировано из оригинала 11 мая 2020 года . Проверено 29 декабря 2014 г.
- ^ Jump up to: а б Гротцингер, Джон П. (24 января 2014 г.). «Введение в специальный выпуск: обитаемость, тафономия и поиск органического углерода на Марсе» . Наука . 343 (6169): 386–387. Бибкод : 2014Sci...343..386G . дои : 10.1126/science.1249944 . ПМИД 24458635 .
- ^ Jump up to: а б Разное (24 января 2014 г.). «Специальный выпуск – Содержание – Исследование обитаемости Марса» . Наука . 343 (6169): 345–452. Архивировано из оригинала 29 января 2014 года . Проверено 24 января 2014 г.
- ^ Разное (24 января 2014 г.). «Специальная коллекция – Любопытство – Исследование обитаемости Марса» . Наука . Архивировано из оригинала 28 января 2014 года . Проверено 24 января 2014 г.
- ^ Гротцингер, JP; и др. (24 января 2014 г.). «Пригодная для жизни речная и озерная среда в заливе Йеллоунайф, кратер Гейла, Марс». Наука . 343 (6169): 1242777. Бибкод : 2014Sci...343A.386G . CiteSeerX 10.1.1.455.3973 . дои : 10.1126/science.1242777 . ПМИД 24324272 . S2CID 52836398 .
- ^ Мэр А. (2000). «Ваза «Троянский монстр»: самое раннее художественное свидетельство об открытии окаменелостей позвоночных?». Оксфордский журнал археологии . 19 (1): 57–63. дои : 10.1111/1468-0092.00099 .
- ^ Монге-Нахера, Хулиан (31 января 2020 г.). «Оценка гипотезы о вазе Троянского монстра как самой ранней художественной записи окаменелостей позвоночных» . Унисиенсия . 34 (1): 147–151. дои : 10.15359/ru.34-1.9 . ISSN 2215-3470 . S2CID 208591414 . Архивировано из оригинала 20 февраля 2023 года . Проверено 20 февраля 2023 г.
- ^ Мильмо, Кахал (25 ноября 2009 г.). «Кража ископаемых: пропал один из наших динозавров» . Независимый . Лондон. Архивировано из оригинала 28 ноября 2009 года . Проверено 2 мая 2010 г.
Саймонс, Льюис. «Ископаемые войны» . Нэшнл Географик . Национальное географическое общество . Архивировано из оригинала 27 февраля 2012 года . Проверено 29 февраля 2012 г.
Уиллис, Пол; Кларк, Тим; Деннис, Карина (18 апреля 2002 г.). «Торговля ископаемыми» . Катализатор . Архивировано из оригинала 24 мая 2012 года . Проверено 29 февраля 2012 г.
Фаррар, Стив (5 ноября 1999 г.). «Меловые преступления, которые подпитывают торговлю ископаемыми» . Высшее образование Таймс . Архивировано из оригинала 20 августа 2012 года . Проверено 2 ноября 2011 г. - ^ Уильямс, Пейдж. «Черный рынок динозавров» . Житель Нью-Йорка . Архивировано из оригинала 22 сентября 2020 года . Проверено 7 сентября 2020 г.
- ^ ван дер Гир, Александра; Дермицакис, Майкл (2010). «Ископаемые в аптеке: от «змеиных яиц» до «костей святого»; обзор» (PDF) . Греческий журнал геонаук . 45 : 323–332. Архивировано из оригинала (PDF) 19 июня 2013 года.
- ^ «Китайские жители ели «драконьи кости» динозавров » . MSNBC . Ассошиэйтед Пресс . 5 июля 2007 г. Архивировано из оригинала 22 января 2020 г. Проверено 7 марта 2020 г.
Дальнейшее чтение
- «Обрушение скалы Гранд-Каньона выявило ископаемые следы возрастом 313 миллионов лет» 21 августа 2020 г., CNN
- «Намеки на ископаемую ДНК, обнаруженные в черепе динозавра» , Майкл Грешко, 3 марта 2020 г., National Geographic
- «Ископаемые для детей | Узнайте все о том, как образуются окаменелости, их типах и многом другом!» Видео (2:23), 27 января 2020 г., Clarendon Learning
- «Ископаемые и их формирование» Видео (9:55), 15 ноября 2019 г., Академия Хана
- «Как образуются окаменелости динозавров?» Лиза Хендри, Музей естественной истории, Лондон.
- Видео "Окаменелости 101" (4:27), 22 августа 2019 г., National Geographic
- «Как обнаружить окаменелости, скрывающиеся на виду», Джессика Ли Хестер, 23 февраля 2018 г., Atlas Obscura
- «Чрезвычайно трудно стать ископаемым». Архивировано 4 сентября 2009 г. в Wayback Machine , Оливия Джадсон , 30 декабря 2008 г., The New York Times.
- «Кости — не единственные ископаемые». Архивировано 15 марта 2009 г. в Wayback Machine , Оливия Джадсон , 4 марта 2008 г., The New York Times.
Внешние ссылки
- Окаменелости в программе «В наше время » на BBC
- Виртуальный музей окаменелостей во времени и эволюции
- Палеопортал, геология и окаменелости США. Архивировано 30 сентября 2009 г. в Wayback Machine.
- Летопись окаменелостей, полный список семейств, отрядов, классов и типов, обнаруженных в летописи окаменелостей (архивировано 3 мая 2012 г.).
- Палеонтология в Керли
- Эрнест Ингерсолл (1920). . Американская энциклопедия .
- . Новая международная энциклопедия . 1905.