Параптоз
Параптоз (от греческого παρά para , «связанный» и апоптоз ) — тип запрограммированной гибели клеток , морфологически отличный от апоптоза и некроза . Определяющими особенностями параптоза являются цитоплазматическая вакуолизация , независимая от активации и ингибирования каспаз , а также отсутствие апоптотической морфологии. В параптозе отсутствуют некоторые характерные характеристики апоптоза, такие как образование пузырьков на мембране , конденсация хроматина и фрагментация ядра . Подобно апоптозу и другим типам запрограммированной гибели клеток, клетка участвует в возникновении собственной смерти, и для этого экспрессия генов требуется . В этом отличие от некроза, который представляет собой незапрограммированную гибель клеток, возникающую в результате повреждения клетки.
Параптоз был обнаружен при гибели некоторых клеток в процессе развития и при нейродегенеративных заболеваниях , а также при воздействии некоторых противораковых препаратов.
Параптоз не был признан Номенклатурным комитетом по клеточной смерти формой гибели клеток в их обзорной статье 2018 года. Комитет явно не одобрял использование этого термина в своей редакции 2012 года. [1]
История
[ редактировать ]Первое сообщение об использовании термина «параптоз» было сделано Сабиной Сперандио и др. в 2000 году. Группа использовала рецептор инсулиноподобного фактора роста 1 человека (IGF-1R) для стимуляции гибели клеток в клетках 293T и эмбриональных фибробластах мыши , наблюдая явные отличия от других форм гибели клеток. Они ввели термин «параптоз», происходящий от греческого предлога para , означающего «рядом» или «связанный с», и «апоптоз» . [2] [3]
Хотя Сперандио был первым, кто опубликовал термин «параптоз», это был не первый случай, когда наблюдалась гибель клеток со свойствами параптоза. Такие термины, как «цитоплазматическая» и «гибель клеток 3-го типа», ранее использовались для описания этих форм гибели клеток. Эти формы морфологически очень похожи на параптоз, и возможно, что некоторые случаи гибели клеток, первоначально описанные как одна из этих форм, являются проявлениями параптоза. [2] [4]
Морфология
[ редактировать ]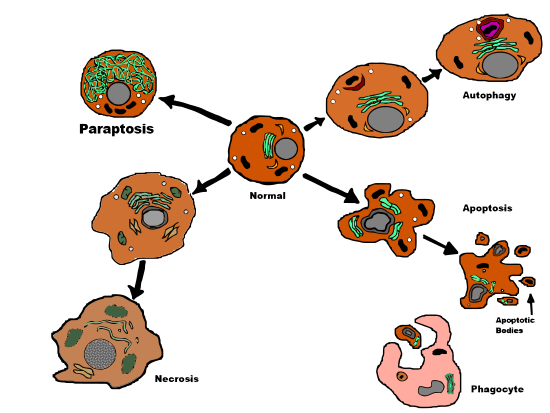
Параптоз — это форма запрограммированной гибели клеток III типа с уникальным сочетанием определенных апоптотических и некротических характеристик. Параптоз не демонстрирует фрагментации ядра , образования апоптотических телец или окончательной демонстрации конденсации хроматина – все это наблюдается при апоптозе. Вместо этого параптоз демонстрирует несколько примитивный путь гибели клеток. [5] сравнимо с некрозом, включая характерное образование цитоплазматических вакуолей и позднее набухание митохондрий. [2] и слипаться. [6] Количество и размер вакуолей со временем увеличиваются. В конце концов, размеры вакуоли достигают точки невозврата, и клетка не может восстановиться. [6]
Подобно апоптозу, методы окрашивания можно использовать для идентификации параптотических клеток путем выявления транслокации фосфатидилсерина из цитоплазматического (внутреннего) листка плазматической мембраны на поверхность клетки или внешний листок. [7]
Морфологические изменения параптоза аналогичны морфологическим изменениям, претерпеваемым в процессе развития нервной системы . [2]
Основная структурная перестройка
[ редактировать ]Почти сразу же с помощью световой микроскопии можно увидеть основные структурные перестройки, такие как округлые клетки, цитоплазматическая реорганизация и вакуолизация клеток, подвергшихся параптозу. [8] Наблюдается физическое увеличение митохондрий и эндоплазматической сети. Такое опухание можно объяснить внутриклеточным ионным дисбалансом и возможным осмотическим лизисом . После разрушения высвобождаются частицы и вещества, в том числе: (1) группа высокой подвижности B-1 ( HMGB1 ) [9] (2) белки теплового шока [10] и (3) различные другие протеазы . Эти вещества являются «сигналами опасности» и приводят к воспалению. [11]
Путь
[ редактировать ]Хотя известно, что некоторые шаблоны запрограммированной гибели клеток зависят от синтеза белка de novo , параптотическая гибель клеток, индуцированная IGFIR-IC в клетках 293T, сдерживается актиномицином D и циклогексимидом , что демонстрирует зависимость от транскрипции и трансляции. [2]
Было установлено, что индукция параптоза опосредована двумя путями передачи положительного сигнала, MAPK и JNK , с использованием IGF-IR на уровне рецептора. Таким образом, параптоз можно предотвратить путем ингибирования специфических протеинкиназ этих путей. [4]
Взаимодействие AIP1 (через его карбоксильный конец ) с эндофилинами может индуцировать образование внутриклеточных вакуолей. [12] Было установлено, что AIP1/Alix является «первым специфическим ингибитором» параптоза. [4]
Параптозоподобный фенотип также был описан в клетках колоректального рака человека после сверхактивации нерецепторной тирозинкиназы c-Src, что позволяет предположить потенциальное участие передачи сигналов Src в параптозе. [13]
Отличия от других путей гибели клеток
[ редактировать ]Гибель клеток, индуцированная IGFIR-IC в клетках 293T, продемонстрировала гибель клеток без связанной с ней каспазной активности. Это по сравнению с апоптозом, при котором проапоптотический белок Bax индуцирует активацию каспазы и гибель клеток. Кроме того, исследования показали, что ингибиторы каспаз (zVAD.fmk, p53 , BAF), ингибитор, связанный с Х-хромосомой ( xiap ), и Bcl-xL (из семейства Bcl-2) не предотвращают гибель клеток 293T при индуцировании ИГФИР-ИК. Таким образом, был сделан вывод, что параптоз отличается от апоптоза (клеточной смерти типа 1) тем, что на него не влияют ингибиторы апоптоза. [2]
При апоптозе HMGB1 , белок хроматина, удерживается внутри ядра, что приводит к образованию апоптотических телец, тогда как при параптозе HMGB1 высвобождается. [14]
Наиболее определяющее различие, наблюдаемое (по состоянию на апрель 2014 г.) между параптозом и аутофагической гибелью клеток (тип гибели клеток 2), заключается в отсутствии у параптоза характерных аутофагических вакуолей, наблюдаемых при аутофагической гибели клеток. [2] Как и ожидалось, ингибиторы гибели аутофагических клеток (например, 3-метиладенин) неэффективны в ингибировании параптоза. [8]
Сравнение типов гибели клеток
[ редактировать ]| Параптоз | Апоптоз | Некроз | |
|---|---|---|---|
| Морфология | |||
| Цитоплазматическая вакуолизация | Да | Нет | Да |
| Конденсация хроматина | Нет | Да | Нет |
| Ядерная фрагментация | Нет | Да | Нет |
| Апоптотические тельца | Нет | Да | Нет |
| Митохондриальный отек | Да | Иногда | Да |
| Мембранное блеббинг | Нет | Да | Да, поздно |
| Запрограммированная гибель клеток | Да | Да | Нет |
| Каспазная активность | Нет | Да | Иногда |
| Ингибиторы | |||
| Актиномицин D, циклогексимид | Да | Иногда | Нет |
| АИП1/Аликс | Да | Нет | Нет |
| Ингибиторы каспаз (zVAD.fmk, p53, BAF) | Нет | Да | Нет |
| Сяп | Нет | Да | Нет |
| Bcl-XL | Нет | Да | Обычно нет |
Протеомный профиль
[ редактировать ]клетки претерпевают как морфологические, так и протеомные При параптозе изменения. Во время параптоза наблюдались изменения в структурных белках, белках сигнальной трансдукции и митохондриальных белках. [8]
Структурные белки
[ редактировать ]В клетках, подвергающихся параптозу:
- α-Тубулин более сконцентрирован в эндосомах и аппарате Гольджи (светлая мембрана) и менее распространен в цитозоле и темной мембране (состоящей из митохондрий и лизосом ). [8]
- Уровень β-тубулина в целом снижается во фракциях параптотических клеток. [8]
- Тропомиозин , подобно α-тубулину, демонстрирует более высокое присутствие в эндосомах и аппарате Гольджи, но имеет уменьшенное содержание в цитозоле и темной мембране. [8]
Белки сигнальной трансдукции
[ редактировать ]PEBP, или киназы Raf белок-ингибитор (RKIP), снижается в параптотических клетках, поэтому результирующая понижающая регуляция PEBP и/или других ингибиторов киназы, по-видимому, указывает на участие в путях MAPK и JNK , поскольку снижение PEBP позволит повысить уровни MAPK и JNK накапливается в достаточном количестве, чтобы вызвать гибель клеток. [8]
Белки митохондрий
[ редактировать ]АТФ-синтаза состоит из множества субъединиц и находится в митохондриях. При параптозе более высокие количества β-субъединицы АТФ-синтазы были продемонстрированы в P20. [8]
Окрашивание митохондрий показывает, что округлые параптотические клетки с повышенным уровнем запретина , по-видимому, подвергаются реорганизации митохондриальной сети. [8]
В параптотических клетках наблюдалось увеличение содержания запретина в 3,4 раза. Повышенные уровни запретина при конъюгации с параптотическим стимулом могут привести к гибели клеток, которую невозможно ингибировать ингибиторами каспаз. [8]
Потенциальное медицинское значение
[ редактировать ]Рак
[ редактировать ]Было показано, что многие противораковые вещества вызывают параптоз в большом количестве раковых клеток человека. Сюда входят несколько соединений, полученных из природных источников. [15] [16] а также металлокомплексы. [17] [18] Параптоз также представляет интерес для исследований рака как способ лечения рака, устойчивого к апоптозу. [15] [19]
Паклитаксел , обычно распространяемый под торговым названием Таксол, представляет собой противораковый препарат, используемый для лечения рака молочной железы и яичников. Одно исследование показало, что при высоких концентрациях (70 мкМ) он вызывает параптозоподобную гибель клеток и может быть важным механизмом лечения рака, устойчивого к апоптозу. [19]
Исследователи сообщили о том, что γ-токотриенол , форма витамина Е, полученная из пальмового масла, вызывает гибель клеток, подобную параптозу, в клетках рака толстой кишки . Наряду с индукцией параптоза γ-токотриенол также подавлял сигнальный путь Wnt , который играет роль в развитии опухоли. Сочетание этих двух особенностей может обеспечить новый механизм лечения рака толстой кишки. [16]
приготовленный на пару экстракт американского женьшеня «мощно убивает клетки колоректального рака». Сообщается, что [20] В частности, производные протопанаксадиола Rg3 и Rh2 являются ключевыми гинсенозидами, обнаруженными в экстракте. [20] В клеточных линиях колоректального рака HCT116 цитозольная вакуолизация индуцируется Rh2. [21] Кроме того, Rh2-индуцированная вакуолизация ингибировалась специфическим ингибитором MEK1/2 U0126 , циклогексимидом , [21] таким образом подтверждая два характерных свойства параптоза: передачу сигнала через MAP-киназу и необходимую трансляцию белка. [4] Rh2 также индуцирует повышение уровней АФК , которые активируют сигнальный путь NF-κB , в то время как блокирование АФК с помощью NAC или каталазы предотвращает активацию передачи сигналов NF-κB и еще больше усиливает гибель клеток, индуцированную Rh2. Это предполагает усиление антиоксидантного противоракового эффекта Rh2. [21]
Хонокиол , соединение, полученное из магнолии лекарственной , может вызывать параптоз в лейкемии клетках человека. В клеточной линии NB4 параптоз был основным методом гибели клеток. В клетках K562 первичным механизмом был апоптоз, иногда обнаруживался параптоз. [15] Исследователи заявили, что это предполагает, что гибель клеток лейкемии может быть вызвана несколькими путями. [15]
В одном эксперименте комплекс фосфина меди(I) вызвал параптоз в клетках рака толстой кишки, вызывая стресс эндоплазматического ретикулума . [17] Другой комплекс меди, комплекс тиоксотриазола меди (II) А0, также вызывал параптоз в клетках фибросаркомы HT1080 посредством стресса эндоплазматического ретикулума и вакуолизации цитоплазмы. Наряду с цитотоксическими эффектами, такими как увеличение окисленного глутатиона и предотвращение активности протеасом , А0 предотвращает активность каспазы-3, которая может ингибировать апоптоз и вызывать гибель клеток в результате параптоза. [18]
Нейродегенеративная гибель клеток
[ редактировать ]Активность супрессора опухолей р53 млекопитающих зависит от уровней изоформ р53 , р44. В эксперименте с трансгенными мышами, у которых была сверхэкспрессия р44, произошла гиперактивация IGF-1R , что, в свою очередь, привело к ускоренному старению и смерти. У мышей также наблюдалась гибель нейронов в областях мозга, связанных с формированием и воспроизведением памяти. Эта нейродегенерация, индуцированная IGF-1R, была вызвана как параптозом, так и аутофагической гибелью клеток. [22] IGF-1R является важной областью исследований нейродегенеративных заболеваний, поскольку в мозге с болезнью Альцгеймера были обнаружены дефекты передачи сигналов IGF-1R, включая повышенные уровни IGF-1R. пациентов [23]
Другие примеры
[ редактировать ]Запрограммированная гибель клеток, подобная параптозу, наблюдалась как у растений, так и у простейших . Апоптотическая смерть, аналогичная той, что наблюдается у животных, не происходит у растений из-за того, что клеточная стенка растительных клеток предотвращает фагоцитоз . В эксперименте с табаком блеомицин использовался для внесения двухцепочечных разрывов в ДНК клеток. Затем это привело к тому, что клетки подверглись запрограммированной гибели клеток со значительной вакуолизацией и отсутствием фрагментации ДНК и ингибирования каспаз, аналогично параптозу. [24] Исследование с водорослями Dunaliella viridis продемонстрировало способность протистов подвергаться запрограммированной клеточной гибели посредством нескольких типов, включая параптоз и апоптоз, в зависимости от различных стимулов окружающей среды. [25] Сочетание этих факторов привело к предположению, что параптоз может быть наследственной формой запрограммированной гибели клеток, сохраняющейся у разных форм жизни. [24]
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Галуцци и др. (июль 2011 г.). «Молекулярные определения подпрограмм клеточной гибели: рекомендации Номенклатурного комитета по клеточной смерти 2012» . Смерть клеток и дифференцировка . 19 (1): 107–120. дои : 10.1038/cdd.2011.96 . ПМЦ 3252826 . ПМИД 21760595 .
- ^ Jump up to: а б с д и ж г час Сперандио, С; де Белль, я; Бредесен, Делавэр (19 декабря 2000 г.). «Альтернативная, неапоптотическая форма запрограммированной гибели клеток» . Труды Национальной академии наук Соединенных Штатов Америки . 97 (26): 14376–81. Бибкод : 2000PNAS...9714376S . дои : 10.1073/pnas.97.26.14376 . ЧВК 18926 . ПМИД 11121041 .
- ^ «Параптоз — Оксфордские словари» . Архивировано из оригинала 29 декабря 2012 года.
- ^ Jump up to: а б с д и Сперандио С., Поксай К., де Белль И., Лафуэнте М.Дж., Лю Б., Насир Дж., Бредесен Д.Е. (октябрь 2004 г.). «Параптоз: опосредование MAP-киназ и ингибирование AIP-1/Alix» . Смерть клеток и дифференцировка . 11 (10): 1066–75. дои : 10.1038/sj.cdd.4401465 . ПМИД 15195070 .
- ^ «Параптоз — новый путь к запрограммированной гибели клеток» . Bioresearch Online (новость). 19 декабря 2000 года. Архивировано из оригинала 26 марта 2014 года . Проверено 25 марта 2014 г.
- ^ Jump up to: а б Кейгл, Филип Т.; Аллен, Тимоти Крейг (10 июня 2009 г.). Основные понятия молекулярной патологии . Спрингер. п. 30. ISBN 9780387896267 . Проверено 19 апреля 2014 г.
- ^ Ван, Ю; Ли, Х; Ван, Л; Дин, П; Чжан, Ю; Хан, В; Ма, Д. (15 марта 2004 г.). «Альтернативная форма гибели клеток, подобная параптозу, вызванная TAJ/TROY и усиленная сверхэкспрессией PDCD5» . Журнал клеточной науки . 117 (Часть 8): 1525–32. дои : 10.1242/jcs.00994 . ПМИД 15020679 .
- ^ Jump up to: а б с д и ж г час я дж Сперандио, С; Поксай, Канзас; Шиллинг, Б; Криппен, Д; Гибсон, BW; Бредесен, Делавэр (15 декабря 2010 г.). «Идентификация новых модуляторов и изменений белков при неапоптотической запрограммированной гибели клеток» . Журнал клеточной биохимии . 111 (6): 1401–12. дои : 10.1002/jcb.22870 . ПМЦ 5668132 . ПМИД 20830744 .
- ^ Ян, Х; Ван, Х; Чура, CJ; Трейси, KJ (июль 2005 г.). «Цитокиновая активность HMGB1» . Журнал биологии лейкоцитов . 78 (1): 1–8. дои : 10.1189/jlb.1104648 . ПМИД 15734795 .
- ^ Мельчер, А; Тодрик, С; Хардвик, Н.; Форд, М; Джейкобсон, М; Вайле, Р.Г. (май 1998 г.). «Имуногенность опухоли определяется механизмом гибели клеток посредством индукции экспрессии белка теплового шока». Природная медицина . 4 (5): 581–7. дои : 10.1038/nm0598-581 . ПМИД 9585232 . S2CID 21603158 .
- ^ Галлуччи, С; Матцингер, П. (февраль 2001 г.). «Сигналы опасности: SOS иммунной системе». Современное мнение в иммунологии . 13 (1): 114–9. дои : 10.1016/s0952-7915(00)00191-6 . ПМИД 11154927 .
- ^ Шательяр-Косс, К; Блот, Б; Кристина, Н; Факел, С; Миссоттен, М; Садул, Р. (9 августа 2002 г.). «Аликс (белок X, взаимодействующий с ALG-2), белок, участвующий в апоптозе, связывается с эндофилинами и индуцирует вакуолизацию цитоплазмы» . Журнал биологической химии . 277 (32): 29108–15. дои : 10.1074/jbc.M204019200 . ПМИД 12034747 .
- ^ Уэлман А., Коуторн С., Понсе-Перес Л. и др. (2006). «Повышение уровня и активности экспрессии c-Src не способствует росту клеток колоректальной карциномы человека in vitro и in vivo» . Неоплазия . 8 (11): 905–16. дои : 10.1593/neo.06475 . ПМК 1716016 . ПМИД 17132222 .
- ^ Коэн, Джей-Джей (март 1993 г.). «Апоптоз». Иммунология сегодня . 14 (3): 126–30. дои : 10.1016/0167-5699(93)90214-6 . ПМИД 8466628 .
- ^ Jump up to: а б с д Ван, Ю; Чжу, Х; Ян, З; Чжао, X (18 января 2013 г.). «Хонокиол индуцирует каспазо-независимый параптоз посредством производства активных форм кислорода, что сопровождается апоптозом в лейкозных клетках». Связь с биохимическими и биофизическими исследованиями . 430 (3): 876–82. дои : 10.1016/j.bbrc.2012.12.063 . ПМИД 23262230 .
- ^ Jump up to: а б Чжан, Дж. С.; Ли, ДМ; Может; Он, Н; Гу, Кью; Ван, Ф.С.; Цзян, SQ; Чен, BQ; Лю, младший (2013). «γ-Токотриенол индуцирует параптозоподобную гибель клеток в клетках карциномы толстой кишки человека SW620» . ПЛОС ОДИН . 8 (2): e57779. Бибкод : 2013PLoSO...857779Z . дои : 10.1371/journal.pone.0057779 . ПМЦ 3585143 . ПМИД 23469066 .
- ^ Jump up to: а б Гандин, В; Пелли, М; Тисато, Ф; Порчиа, М; Сантини, К; Марцано, К. (январь 2012 г.). «Новый комплекс меди вызывает параптоз в клетках рака толстой кишки посредством активации передачи сигналов стресса ER» . Журнал клеточной и молекулярной медицины . 16 (1): 142–51. дои : 10.1111/j.1582-4934.2011.01292.x . ПМК 3823100 . ПМИД 21388518 .
- ^ Jump up to: а б Тардито, С; Изелла, К; Медико, Э; Маркио, Л; Бевилаква, Э; Хацоглу, М; Буссолати, О; Франки-Газзола, Р. (4 сентября 2009 г.). «Комплекс тиоксотриазола меди(II) А0 вызывает стресс эндоплазматического ретикулума и параптотическую гибель раковых клеток человека» . Журнал биологической химии . 284 (36): 24306–19. дои : 10.1074/jbc.M109.026583 . ПМК 2782024 . ПМИД 19561079 .
- ^ Jump up to: а б Го, WJ; Чен, ТС; Ван, XP; Чен, Р. (2010). «Таксол индуцирует зависимую от концентрации апоптозную и параптозоподобную гибель клеток в клетках аденокарциномы легких человека (ASTC-a-1)». Журнал рентгеновской науки и техники . 18 (3): 293–308. дои : 10.3233/XST-2010-0261 . ПМИД 20714087 .
- ^ Jump up to: а б Ли, Б; Ван, Чехия; Он, ТК; Юань, CS; Ду, В. (1 марта 2010 г.). «Антиоксиданты усиливают вызванное американским женьшенем уничтожение клеток колоректального рака» . Письма о раке . 289 (1): 62–70. дои : 10.1016/j.canlet.2009.08.002 . ПМК 2824022 . ПМИД 19716228 .
- ^ Jump up to: а б с Ли, Б; Чжао, Дж; Ван, Чехия; Сирл, Дж; Он, ТК; Юань, CS; Ду, В. (28 февраля 2011 г.). «Гинсенозид Rh2 индуцирует апоптоз и параптозоподобную гибель клеток в клетках колоректального рака посредством активации р53» . Письма о раке . 301 (2): 185–92. дои : 10.1016/j.canlet.2010.11.015 . ПМК 3022099 . ПМИД 21194832 .
- ^ Пехар М., О'Риордан К.Дж., Бернс-Кусато М., Анджеевски М.Е., дель Алькасар К.Г., Бургер С., Скрэбл Х., Пульелли Л. (апрель 2010 г.). «Измененная активность р53:р44, гарантирующая долголетие, у мышей вызывает потерю памяти, нейродегенерацию и преждевременную смерть» . Стареющая клетка . 9 (2): 174–90. дои : 10.1111/j.1474-9726.2010.00547.x . ПМЦ 2848983 . ПМИД 20409077 .
- ^ Молони, AM; Гриффин, Р.Дж.; Тиммонс, С; О'Коннор, Р.; Равид, Р; О'Нил, C (февраль 2010 г.). «Дефекты рецептора IGF-1, рецептора инсулина и IRS-1/2 при болезни Альцгеймера указывают на возможную устойчивость к IGF-1 и передаче сигналов инсулина». Нейробиология старения . 31 (2): 224–43. doi : 10.1016/j.neurobiolaging.2008.04.002 . ПМИД 18479783 . S2CID 14265087 .
- ^ Jump up to: а б Сметана, О; Широкий, Ю; Ульне, Дж; Опатрный, З; Чабуте, Мэн (апрель 2012 г.). «Неапоптотическая запрограммированная гибель клеток с параптотическими характеристиками в обработанных блеомицином растительных клетках подавляется путем ингибирования путей ATM/ATR или сверхэкспрессии NtE2F» . Журнал экспериментальной ботаники . 63 (7): 2631–44. дои : 10.1093/jxb/err439 . ПМИД 22268149 .
- ^ Хименес, К; Капассо, Дж. М.; Эдельштейн, CL; Ривард, CJ; Люсия, С; Бреузегем, С; Берл, Т; Сеговия, М (2009). «Различные способы смерти: способы гибели клеток одноклеточного хлорофита Dunaliella viridis, подвергающегося различным стрессам окружающей среды, опосредуются каспазоподобной активностью DEVDазы» . Журнал экспериментальной ботаники . 60 (3): 815–28. дои : 10.1093/jxb/ern330 . ПМК 2652065 . ПМИД 19251986 .