Аноплотерий
| Аноплотерий | |
|---|---|
 | |
| Неполный скелет коммуны Anoplotherium из коммуны Пантен , Национальный музей естественной истории, Франция. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Семья: | † Аноплотерииды |
| Подсемейство: | † Аноплотериины |
| Род: | † Аноплотерий Кювье , 1804 г. |
| Типовой вид | |
| † Аноплотерийная коммуна Кювье, 1804 г. | |
| Другие виды | |
| |
| Синонимы | |
Синонимия рода Синонимы А. коммуны Синонимы A. latipes Синонимы A. laurillardi | |
Anoplotherium — типовой род вымершего палеогенового парнокопытных семейства Anoplotheriidae , эндемичного для Западной Европы. Он жил с позднего эоцена до самого раннего олигоцена . Это был пятый род ископаемых млекопитающих, описанный официальными таксономическими авторитетами, история которого восходит к 1804 году, когда его окаменелости с Монмартра в Париже , Франция, были впервые описаны французским натуралистом Жоржем Кювье . Открытия неполных скелетов A. commune в 1807 г. побудили Кювье тщательно описать необычные особенности, не имеющие современных аналогов. Его нарисованные реконструкции скелета и мышц A. commune в 1812 году были одними из первых примеров анатомических реконструкций, основанных на ископаемых свидетельствах. Вклад Кювье в палеонтологию, основанный на его работах по этому роду, был революционным для этой области, не только доказав развивающиеся идеи вымирания и экологической преемственности, но и проложив путь для таких областей, как палеоневрология . Сегодня известно четыре вида.
Аноплотерий был одним из крупнейших парнокопытных, не являющихся виппоморфами , палеогенового периода: он весил в среднем от 115 кг (254 фунта) до 271 кг (597 фунтов), имел длину головы и тела не менее 2,5 м (8 футов 2 дюйма) и длину тела 1,25 м. (4 фута 1 дюйм) на высоте плеч. , имел трехпалые лапы Это был эволюционно развитый и необычный парнокопытный, у некоторых видов, таких как A. latipes , длинный и крепкий хвост и высокоразвитый мозг с сильной поддержкой как обоняния, так и сенсорного восприятия . Его общее крепкое телосложение, возможно, позволяло ему стоять на двух ногах и щипать растения на большей высоте, достигая примерно 3 м (9,8 футов) в высоту, что эффективно конкурировало с немногими другими травоядными животными среднего и крупного размера, с которыми он жил. Однако полная степень его двуногости требует подтверждения дополнительными исследованиями. Более крупный двупалый A. commune и немного меньший трехпалый A. latipes могут быть половыми диморфами, поскольку первый - женский, а второй - мужской, но эта идея остается спекулятивной. Его ближайшим родственником был Diplobune , который, как предполагается, также имел особое поведение.
Парнокопытные жили в Западной Европе еще тогда, когда это был архипелаг , изолированный от остальной части Евразии, а это означает, что они жили в среде с различными другими фаунами, которые также развивались с сильным уровнем эндемизма. Его точное происхождение неизвестно, но оно возникло спустя много времени после перехода к более сухим, но все еще субгумидным условиям, что привело к появлению абразивных растений и вымиранию крупных Lophiodontidae , достигнув гигантизма и утвердившись в качестве доминирующего травоядного животного на всей территории Западной Европы. регион, учитывая его многочисленные ископаемые свидетельства.
Его успех был внезапно остановлен вымиранием Гранд-Купюр и сменой фауны в самом раннем олигоцене Западной Европы, что было вызвано сдвигом в сторону дальнейшего оледенения и сезонности. Тропические и субтропические леса были быстро заменены более умеренной средой, а большинство океанских барьеров, ранее отделявших Западную Европу от Восточной Евразии, закрылись, что привело к значительному расселению фауны из Азии. Хотя конкретные причины неизвестны, Anoplotherium, вероятно, не смог адаптироваться к этим серьезным изменениям и вымер.
История исследований
[ редактировать ]Идентификации
[ редактировать ]

Хотя Жорж Кювье знал об ископаемых костях из гипсовых карьеров на окраине Парижа (известных как Парижский бассейн ) по крайней мере еще в 1800 году, он описал их только в 1804 году. После описания Palaeotherium он написал о следующем наборе окаменелостей, которые он смог отличить от Palaeotherium на основании формы зубных рядов, включая очевидное отсутствие клыков , оставляющее большой зазор между резцами и премолярами . Он заметил, что получелюсть (половина нижней челюсти ) имела три нижних резца вместо четырех или вообще не имела ни одного, что, по его словам, характерно для других « толстокожих ». Кювье, основываясь на очевидном отсутствии у него подходящих рук и собак для наступательных атак, придумал название Anoplotherium . [1] [2]
Название рода Anoplotherium означает «безоружный зверь» и представляет собой соединение греческих слов αν- ( ан , «не»), ὅπλον ( хоплон , «доспехи, большой щит») и θήρ ( thēr , «зверь, дикое животное»). ). [3]
Кювье назвал три вида Anoplotherium В том же году , первый из которых был A. commune «размером с овцу» , а остальные три были «более мелкими видами», которые он назвал A. medium , A. minus и A. minus . минимум . Этимология названия вида A. commune указывает на то, насколько «общими» были окаменелости этого вида, в то время как этимология двух других видов основывалась на размерах по сравнению с A. commune . [а] Он также приписал раздвоенное копыто (или дидактильное копыто), A. commune поскольку экземпляр оказался большого размера. Он думал, что у Anoplotherium были дидактильные копыта вместо трехпалых, что отличало бы его от Palaeotherium . Основываясь на копытах и зубных рядах, он пришел к выводу, что Anoplotherium был похож на жвачных животных или верблюдов . [4] [5] Однако в 1807 году Кювье обнаружил, что у Anoplotherium commune на задних конечностях было три пальца, хотя третьи указательные пальцы были меньших размеров по сравнению с двумя другими. [6]
Скелеты
[ редактировать ]

В 1807 году Кювье писал о двух неполных скелетах, которые были недавно обнаружены, хотя первый был частично поврежден, поскольку его не собирали тщательно (что, по его словам, изначально расстроило его понимание анатомии скелета Anoplotherium ). Первый скелет, найденный в каменоломнях Монмартра в коммуне Пантен , помог подтвердить ранее поставленного Кювье диагноза Anoplotherium правильность . Встроенный скелет был размером с небольшую лошадь и помог подтвердить большие дидактильные ступни и 44 зуба (по 11 на каждой стороне челюсти). У него также было 11 целых ребер и фрагмент 12-го, соответствующий количеству ребер верблюдовых. Однако самым удивительным элементом для Кювье стал огромный хвост с 22 позвонками в скелете - особенность, о которой, по его словам, он не знал раньше, поскольку не существует современных аналогов удлиненного и толстого хвоста ни у одного крупного четвероногого млекопитающего. . [7]
Второй неполный скелет принадлежал Антонию , на этот раз удаленный более тщательно под наблюдением экспертов, чем первый скелет. В нем ему удалось подтвердить шесть поясничных позвонков и три крестцовых позвонка , все из которых были чрезвычайно прочными и, вероятно, поддерживали длинный хвост. Наиболее примечательным для Кювье было подтверждение того, что у Anoplotherium было два больших пальца и один мизинец на передних ногах, что было необычно для родственных ему млекопитающих. [7]
Значение в палеонтологической истории
[ редактировать ]
Хотя Palaeotherium и Anoplotherium не так хорошо известны по сравнению с ископаемыми животными других периодов (например, динозаврами мезозоя и млекопитающими неоген - четвертичного периода ), их ископаемые открытия на Монмартре и формальные описания Кювье признаны критическими моментами, которые положили начало палеонтологии в современную эпоху. В отличие от ископаемых родов плейстоцена в Америке в ранней палеонтологической истории, таких как Megatherium и Mammut , Palaeotherium и Anoplotherium не были обнаружены в отложениях на уровне поверхности, а были заключены в более глубокие и твердые отложения пород, датируемые эоценом . Жители Парижа ранее на протяжении веков были знакомы со скелетами животных, находившимися в их районе, некоторые из которых позже были сохранены и официально описаны. Однако именно Кювье формально выделил два ископаемых рода, происходящих из более древних отложений и с его родины на европейском континенте, а не в Америке, где были обнаружены Megatherium и Mammut . [8] Окаменелости палеогенового возраста не оставили никаких свидетельств существования каких-либо более поздних потомков, вымерших или сохранившихся до наших дней, хотя сходство палеотериев с тапирами затрудняло доказательство этой теории. Он заметил, что под гипсом находились более древние отложения морских ракушек и рептилий, подобных тому, кого Кювье описал как гигантского «крокодила», который позже будет известен как Мозазавр . Тогда Кювье знал, что мир, из которого произошли Anoplotherium и Palaeotherium, находился в другом промежутке времени, предшествующем предшествующему времени морских рептилий и последующим временам Megatherium и Mammut , тем самым доказывая концепцию естественного вымирания. [9]
Описания Кювье эндокаста ( окаменевшего черепа) полушария головного мозга, принадлежащего сломанному черепу A. commune с Монмартра, начиная с 1804 по 1822 год, признаны первым настоящим примером палеоневрологии , исследования эволюции мозга. Самое первое определение «эндокаста» датируется 1822 годом, когда Кювье описал слепок мозга A. commune , заметив, что он намекает на истинную форму мозга ныне вымершего млекопитающего (хотя позже было обнаружено, что быть частью мозга, а не целым его). Со времени первого исследования эндокаста на протяжении второй половины XIX века было проведено множество других исследований мозга других ископаемых млекопитающих. [10] [11] [12] Описание Кювье в 1822 году зажившего перелома бедренной кости A. commune упоминается как ранний пример палеопатологии , изучения древних болезней и травм доисторических организмов. [13] [14] [15]
Ранние изображения
[ редактировать ]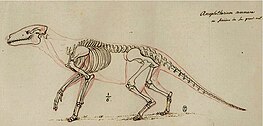
В 1812 году Кювье опубликовал рисунок реконструкции скелета A. commune на основе известных ископаемых останков этого вида, включая вышеупомянутые неполные скелеты. Основываясь на крепком телосложении этого вида млекопитающих, он выдвинул гипотезу, что его строение тела было похоже на строение тела выдры, за исключением ног, что оно было приспособлено к полуводной жизни, плавая для поедания водных растений, и у него не было длинных ушей, как у полуводных животных. млекопитающие, живущие в болотистой среде. Кювье предположил, что его образ жизни был похож на образ жизни полуводных четвероногих млекопитающих, таких как гиппопотамы и муроидные грызуны. Он считал, что для сравнения другие виды Anoplotherium, такие как A. medium и A. minus, были адаптированы к наземному поведению и смешанному питанию (просматриванию и выпасу). [16] [17] Сегодня реконструкция анатомии скелета хорошо устарела и в основном выдержала испытание временем с 1812 года. [18] Аноплотерий и палеотерий также были изображены на рисунках 1822 года французского палеонтолога Шарля Леопольда Лорийяра под руководством Кювье, хотя реставрации не были такими подробными, как у Кювье. [19]
Реконструкция Anoplotherium как водного пловца была поддержана многими европейскими палеонтологами XIX века и сохранялась более века. [20] [21] до 1938 года, когда М. Дор отверг теорию этого рода как адаптированного к водной среде, основанную на анатомических отличиях от выдр и гиппопотамов, которые противоречат полуводному поведению и более соответствуют наземной жизни. [22] Этот отказ был поддержан Джерри Дж. Хукером в 2007 году и Свитозаром Давыденко и др. в 2023 году на основании анатомических особенностей, хотя первый не соглашался с наблюдениями Дора по поводу хвоста. Хукер утверждал, что, хотя дистальные хвостовые позвонки аноплотера менее заметны, чем у кенгуру ( Macropus ), структура позвонков Anoplotherium больше похожа на Macropus, чем на копытных, таких как Bos или Equus . Сегодня Anoplotherium считается наземным браузером со специальным поведением. [23] [24]

А. коммуна была особенно изображена на аттракционе «Динозавры Хрустального дворца» в парке Хрустального дворца в Соединенном Королевстве , открытом для публики с 1854 года и построенном английским скульптором Бенджамином Уотерхаусом Хокинсом . Точнее, были изготовлены три статуи А. Коммуны , две из которых стоят, а третья находится в покое. Эти статуи напоминают гибриды оленей и больших кошек и имеют длину 3,6 м (12 футов). Его включение в парк Хрустального дворца отражает популярность и общественный интерес к аноплотерию в 19 веке, поскольку он был символом палеонтологии, геологии и естествознания и регулярно включался в палеонтологические тексты и учебные классы (его популярность снизилась с 20 века). века). [25] [26]
Скульптуры A. commune в целом были основаны на описании рода Хокинсом, основанном на известных останках, включая неопубликованные предположения Кювье о мускулах, которые по современным стандартам считаются точными. Однако Хокинс также отклонился от описаний Кювье, вероятно, основываясь на дизайне лица и неточном наличии тетрадактильных конечностей (четыре пальца на каждой ноге) вместо дидактильных или трехпалых конечностей у современных верблюдовых. Помимо этих ошибок, статуи во многом соответствуют современным изображениям аноплотерия . [26]
Путаница с другими группами млекопитающих.
[ редактировать ]
На протяжении большей части XIX века палеонтологи путали млекопитающих других семейств с Anoplotherium во многом из-за того, что палеонтология находилась на ранних стадиях своего развития. Одним из более ранних примеров является 1822 год, когда Кювье ввел названия A. gracile , A. murinus , A. obliquum , A. leporinum и A. secundaria , заменив более ранние названия видов в пределах Anoplotherium за пределами A. commune . У A. gracile он заметил различия в коренных зубах , которые возвел в подрод Xiphodon . Для A. leporinum подрод Dichobune исходя из ее небольших размеров. им был создан [11] В 1852 году Поль Жерве повысил два подрода до родовых рангов и выделил дополнительный род Amphimeryx для A. murinus . Таким образом, виды больше не относят к Anoplotherium , а к отдаленным родам. [27] [28]
Другие млекопитающие, первоначально спутанные с родом Anoplotherium , но в конечном итоге реклассифицированные в XIX веке, представляли собой эндемичное европейское семейство парнокопытных Cainotheriidae ( Cainotherium [29] [30] [28] [31] ), европейские и индийские субконтинентальные представители семейства непарнокопытных Chalicotheriidae ( Anisodon [32] [33] [34] и несторитий [35] [36] [37] ), и даже эндемичные южноамериканские представители отряда Litopterna ( Scalabrinitherium и Proterotherium [38] ).
Изменения в Anoplotheriidae
[ редактировать ]
В 1851 году Помель заметил, что виды Anoplotherium могут быть определены как имеющие либо дидактильные копыта (уменьшенный третий индекс), либо трехпалые копыта (более развитый третий индекс), и что единственными ранее установленными видами, которые являются действительными, являются A. commune и A. secundaria . Кроме того, он вывел три новых вида на основе дополнительных останков: A. duvmoyi , сделанных Кювье (на основе ископаемых иллюстраций A. commune ), A. platypus , A. laurillardi (выпуклые резцы на передней поверхности) и A. cuvieri . A. laurillardi происходит от имени Чарльза Лорилларда. Видовое название [30]
Жерве в 1852 году назвал род Eurytherium на основании наличия у него трехпалых копыт вместо дидактильных, поскольку он сделал новый вид E. latipis типовым видом, а A. platypus - синонимом первого. [20] Анри Филхол последовал примеру Жерве, установив E. quercyi и E. minus на основе размеров зубов и переклассифицировав A. secundarium (или A. secundaria ) в Eurytherium . [39]
В 1862 году Людвиг Рютимейер выделил подрод Diplobune для рода Dichobune на том основании, что это был эволюционный переход между Anoplotherium secundarium и дихобунидом. [40] выделил его в отдельный род, и один вид D. bavaricum был помещен в этот род Однако к 1870 году Оскар Фраас . [41]
В 1883 году Макс Шлоссер сделал Eurytherium синонимом Anoplotherium , поскольку утверждал, что анатомия конечностей и зубные ряды представляют собой конкретные различия в характеристиках, а не основные различия, которые определяют весь род. Слоссер указал, что все виды Anoplotherium в той или иной форме имели три индекса, несмотря на то, что у A. commune третьи индексы были менее развиты, чем у A. latipes . Он также подтвердил идею о том, что « A. platypus » является синонимом A. latipes . Название A. latipes имеет приоритет над A. platypus, до сих пор поскольку Помель в 1851 году не перечислил ни одного экземпляра этого вида, что фактически сделало его nomen dubium . Он также упомянул, что статус A. duvmoyi не был стабильным из-за того, что он основан на иллюстрациях, что он считал «безнадежной попыткой». Он также поддержал признание рода Diplobune действительным, утверждая, что A. secundaria следует переименовать в D. secundaria на основании зубного ряда и меньших размеров. Шлоссер также сказал, что A. cuvieri является недействительным видом, поскольку диагноз основан на изолированных данных. плюсневых костей было достаточно. [21] [28] [23]
Ричард Лидеккер вывел вид A. cayluxense в 1885 году на основании его меньшего размера и уникальных вариаций бугорков коренных зубов. Он также понизил род Diplobune как синоним Anoplotherium , а это означает, что виды первого были добавлены/добавлены к Anoplotherium как A. secundarium , A. quercyi , A. modicum , A. bavaricum и A. minus (= A.minor , Филхол 1877). [28] Синонимия Diplobune с Anoplotherium не была поддержана Гансом Георгом Штелином в 1910 году, поскольку он утверждал, что первый в целом отличается от второго, несмотря на их близкое родство, тем самым восстанавливая предыдущий вид в Diplobune (за исключением D. modicum , который он сделал его синонимом D. bavarica ) и добавил « A. secundarium » в Diplobune как D. secundaria . Он также писал, что A. cayluxense — синоним D. secundaria . Штелин также предварительно отнес « A. » obliquum к роду Haplomeryx как H? косой . В результате доработок единственными действительными видами Anoplotherium были A. commune , A. latipes и A. laurillardi . [42]
В 1922 году Вильгельм Отто Дитрих вывел четвертый вид A. pompeckji из местонахождения Меринген в Германии, названный в честь немецкого палеонтолога Йозефа Феликса Помпека . Этот вид был описан как трехпалый вид среднего размера с 4-палыми передними конечностями и 3-палыми задними конечностями, с более тонкими пропорциями костей рук и меньшими таранными костями . [43] A. pompeckji — наименее изученный вид, и его зубные ряды аналогичны A. laurillardi , что делает его статус менее определенным по сравнению с тремя другими видами. [23] [44]
В 1964 году палеонтолог Луи де Бонис кратко рассмотрел таксономические синонимы Anoplotherium , посчитав, что A. duvernoyi произошел от молодой особи с характеристиками резцов, которые Помель не уточнил, и что A. cuvieri не отличается по размерам пястных костей от A. laurillardi . Он последовал примеру Стелина в признании трех основных видов Anoplotherium , хотя и не упомянул в своем обзоре A. pompeckji . [45]
Классификация
[ редактировать ]
Anoplotherium — это типовой род Anoplotheriidae, палеогенового семейства парнокопытных, эндемичного для Западной Европы, жившего от среднего эоцена до раннего олигоцена (от ~ 44 до 30 млн лет назад, возможно, самая ранняя запись около 48 млн лет назад). Точное эволюционное происхождение и распространение аноплотериид неясны, но они проживали исключительно в пределах континента, когда он был архипелагом , изолированным морскими барьерами от других регионов, таких как Балканатолия и остальная часть восточной Евразии. Отношения Anoplotheriidae с другими членами парнокопытных не совсем изучены: некоторые определяют, что это либо тилопод (включая верблюдовых и мерикододонтов палеогена), либо близкий родственник инфраотряда, а некоторые другие полагают, что это мог быть ближе к Ruminantia (к которым относятся трагулиды и другие близкие палеогеновые родственники). [46] [44]
Anoplotheriidae состоит из двух подсемейств: Dacrytheriinae и Anoplotheriinae , последнее из которых является младшим подсемейством, Anoplotherium к которому принадлежит . Dacrytheriinae — более древнее из двух подсемейств, впервые появившихся в среднем эоцене (начиная с подразделения MP13 Палеозоны млекопитающих , возможно, до MP10), хотя некоторые авторы считают их отдельным семейством в виде Dacrytheriidae. [47] [48] Anoplotheriines впервые появились в позднем эоцене (MP15-MP16), или ~41-40 млн лет назад, в Западной Европе вместе с Duerotherium и Robiatherium . Однако к MP17a-MP17b в летописи окаменелостей аноплотиинов в целом наблюдается заметный пробел, поскольку первые два рода, по-видимому, в последний раз появлялись на предыдущем уровне MP MP16. [49]
К MP18 Anoplotherium и Diplobune впервые появились в Западной Европе, но их точное происхождение неизвестно. Эти два рода были широко распространены по всей Западной Европе, что подтверждается многочисленными ископаемыми свидетельствами из Португалии, Испании, Великобритании, Франции, Германии и Швейцарии на большей части Европы до Grande Coupure (до MP21), а это означает, что они были типичными элементами позднего эоцена вплоть до самого раннего олигоцена. [50] [49] [44] Более ранние аноплотериины считаются более мелкими видами, тогда как более поздние аноплотериины были более крупными. Anoplotherium и Diplobune считаются наиболее производными (или эволюционно недавними) аноплотериидами, основанными на морфологии зубов, и достигли гигантизма среди парнокопытных, не относящихся к виппоморфам , что делает их одними из крупнейших парнокопытных, не являющихся виппоморфами, палеогена, а также одними из крупнейших млекопитающих, бродивших по западу. Европа того времени (все виды Anoplotherium были от больших до очень крупных, тогда как не все виды Diplobune были крупными). [44] [51] [52] [12]

Проведение исследований, посвященных филогенетическим связям внутри Anoplotheriidae, оказалось затруднительным из-за общей нехватки ископаемых образцов большинства родов. [49] Филогенетические отношения Anoplotheriidae, а также Xiphodontidae , Mixtotheriidae и Cainotheriidae также были неуловимыми из-за селенодонтной морфологии коренных зубов, которые сходились с тилоподами или жвачными животными. [52] Некоторые исследователи считали, что семейства селенодонтов Anoplotheriidae, Xiphodontidae и Cainotheriidae относятся к Tylopoda из-за посткраниальных особенностей, сходных с тилоподами из Северной Америки в палеогене. [23] Другие исследователи на основании морфологии зубов связывают их с более близкими родственниками жвачных животных, чем с тилоподами. Различные филогенетические анализы дали разные результаты для «производных» селенодонтных европейских семейств парнокопытных эоцена, что делает неясным, были ли они ближе к Tylopoda или Ruminantia. [53] [54]
В статье, опубликованной в 2019 году, Ромен Веппе и др. провел филогенетический анализ Cainotherioidea среди парнокопытных на основе характеристик нижней челюсти и зубов, в частности, с точки зрения взаимоотношений с парнокопытными палеогена. Результаты показали, что это надсемейство было тесно связано с Mixtotheriidae и Anoplotheriidae. Они определили, что Cainotheriidae, Robiacinidae , Anoplotheriidae и Mixtotheriidae образовали кладу, которая была сестринской группой для Ruminantia, в то время как Tylopoda вместе с Amphimerycidae и Xiphodontidae разделились на дереве раньше. [54] Филогенетическое древо, использованное для журнала и другой опубликованной работы о каинотериоидах, представлено ниже: [55]
| |||||||||||||||||||||||||||||||||||||
провел филогенетический анализ В 2022 году Веппе в своей академической диссертации линий палеогеновых парнокопытных, уделив особое внимание эндемичным европейским семействам. Филогенетическое древо, по мнению Веппе, является первым, которое выявило филогенетическое сходство всех родов аноплотериид, хотя не все отдельные виды были включены. Он обнаружил, что Anoplotheriidae, Mixtotheriidae и Cainotherioidea образуют кладу, основанную на синапоморфных зубных чертах (признаки, которые, как полагают, произошли от их самого недавнего общего предка). Результат, как упомянул Веппе, совпадает с предыдущими филогенетическими анализами Cainotherioidea и других эндемичных европейских палеогеновых парнокопытных, которые поддерживают эти семейства как кладу. В результате он утверждал, что предложенное надсемейство Anoplotherioidea, состоящее из Anoplotheriidae и Xiphodontidae, предложенное Аланом В. Джентри и Хукером в 1988 году, недействительно из-за полифилии линий в филогенетическом анализе. Тем не менее, было обнаружено, что Xiphodontidae составляют часть более широкой клады вместе с тремя другими группами. Anoplotherium и Diplobune составляют кладу Anoplotheriidae из-за производных от них стоматологических признаков, что подтверждается тем, что они являются последними появившимися аноплотериидами. [52] [56]
Описание
[ редактировать ]Череп
[ редактировать ]
Anoplotheriidae частично характеризуются низкими пропорциями черепа с удлиненной мордой (морда совпадает с верхней частью черепа в случае Anoplotherium ) и широко открытой орбитой черепа. [57] [47] [44] У аноплотерия отсутствуют костные отростки и слезные ямки . Имеет крупные околозатылочные отростки и более короткие заглазничных отростков выступы слезной кости . [58] [59]
Череп Anoplotherium узкий и удлиненный, с суженной заглазничной костью, что указывает на плохое развитие мозга. Он имеет мощные сагиттальный и затылочный гребни , первый из которых имеет высокую высоту и выходит из низких заглазничных гребней, а второй имеет сложные смещения высоты. На спине имеется круглое большое затылочное отверстие и большие затылочные мыщелки. На нижней стороне имеется удлиненное небо с суставными поверхностями и сильными постгленоидными отростками чешуйчатой кости . [60]
Кости черепа крепкие, губчатая кость диплоэ сильно развита. Сила черепа объясняется массивными височными мышцами , которые являются частью общего сильного телосложения. Череп имеет неглубокое турецкое седло грушевидной формы , черепную ямку , обширные теменные кости , большую чешуйчатую кость, узкую затылочную кость и две небольшие затылочные бугорки для прикрепления мышц. Многие черепные черты, наблюдаемые у Anoplotherium, также встречаются у близкородственного Diplobune . [60] [61]
В слуховой области (включая височные кости) околоушная кость внутреннего уха обширна, внутренний слуховой проход и отверстия лицевого канала в нижней треугольной области околоушной кости видны височной кости. Барабанная часть височной кости частично соединяется с чешуйчатой костью, остается отдельной от околоушной кости и состоит из небольшой, но толстой слуховой буллы (полой костной структуры слуховой области), выступающей под каменистой частью височной кости . кость . [60]
Во фрагменте черепа A. laurillardi с резцами и альвеолами клыков известная длина носовой области велика и составляет 38,1 мм (1,50 дюйма). [44] Признак больших носовых костей аналогичен тому, что наблюдалось у черепа Diplobune secundaria , который, как зарегистрировано, был массивным, удлиненным и соединенным друг с другом и верхней челюстью . Сирил Ганьесон и Жан-Жак Леру предположили в случае D. secundaria , что удлиненная носовая область подтверждает наличие очень заостренного языка, который, как и у жирафов, мог позволить ему тянуть ветки растений. [62]
Анатомия эндокаста
[ редактировать ]

В 1913 году Р. У. Палмер провел исследования слепка мозга из черепа Anoplotherium commune , происходящего от фосфоритов Керси в коллекциях Британского музея (эндокаст сейчас находится в Национальном музее естественной истории, Франция, как образец BMNH 3753). По оценкам, на момент смерти рассматриваемая особь весила 80 кг (180 фунтов), как и современные ламы , и весила значительно меньше, чем типичные оценки взрослых Anoplotherium . Общая длина мозга составляет менее 10 см (3,9 дюйма), его объем составляет около 230 мл (8,1 имп. жидких унций; 7,8 жидких унций США). [60] [63] [44]
Форма мозга от природы узкая и вытянутая. [60] [12] Мозжечок и большой мозг занимают более высокое положение по сравнению с современными копытными, у которых полушария мозга расположены над мозжечком. Палмер заметил, что мозг похож на мозг современного трубкозубого ( Orycteropus afer ). Высокоразвитый головной мозг, обеспечивающий сильное обоняние у Anoplotherium , делает его макросматическим (происходящим от обоняния), на что также указывают увеличенные обонятельные луковицы и небольшой размер неокортекса . [60] И у Anoplotherium и у Diplobune , ринальная щель делит полушарие мозга горизонтально и поровну пополам. Червь мозжечка почти поровну разделен первичной щелью мозжечка (или «fissura prima»). [64]
Кроме того, обонятельные луковицы толстые, а обонятельные бугорки имеют форму гладких круглых возвышений, изогнутых больше назад, чем у трубкозубов, и легко заметных. [60] В другом эндокасте Anoplotherium обонятельные луковицы составляют 7,5% от общего объема мозга, что выше среднего как для вымерших, так и для современных парнокопытных. [12]
Область неокортекса головного мозга, отвечающая за сенсорное восприятие среднего размера . A. commune и другие сенсорные функции мозга, покрывает 28% площади поверхности эндокаста [63] Другой эндокаст, принадлежащий Anoplotherium sp., имеет размер 7173,92 мм (282,438 дюйма). 2 на поверхности головного мозга - 4419,56 мм (173,998 дюйма) 2 на поверхности неопалия и 416,09 см (163,81 дюйма) 3 в эндокраниальном объеме. Первые два данных при совместном расчете (поверхность неопаллия/поверхность головного мозга) составляют 61,6% от общей площади неокортикальной поверхности мозга, а это означает, что взрослый Anoplotherium имеет большие размеры площади мозга и неокортикальной поверхности по сравнению с большинством палеогеновых парнокопытных, причем последнее измерение находится на на одном уровне или меньше, чем у современных парнокопытных. [12]
Anoplotherium и другие аноплотерииды имеют общие черты: обычно удлиненные и параллельные борозды (мелкие бороздки) в коре головного мозга , а также вертикальная (сердечная) борозда, соответствующая латеральной (боковой) борозде. Трещины (глубокие борозды) на поверхности центральной области мозга демонстрируют четкие образования сложной латеральной борозды (также известной как сильвиева трещина) в процессе, известном как оперкулизация. [12] Оперкулизация мозга аноплотериид сходна с Anthracotheriidae , но не указывает на какое-либо близкое филогенетическое родство, а это означает, что сходство является примером параллельной эволюции . Согласно исследованию Гислена Тьери и Стефана Дюкрока, проведенного в 2015 году, размеры эндокастов Anoplotherium больше, чем у других палеогеновых парнокопытных. [65]
Зубной ряд
[ редактировать ]
В отличие от большинства ископаемых родов млекопитающих, диагноз Anoplotherium ставится в основном на основании посткраниальной морфологии, а не морфологии зубов, но у него есть диагнозы, основанные на последней. [44] Зубная формула Anoplotherium всего и других аноплотериидов равна 3.1.4.3 3.1.4.3, 44 зуба, что соответствует примитивной зубной формуле плацентарных млекопитающих раннего среднего палеогена. [57] [66] У аноплотериидов есть селенодонтные или буселенодонтные премоляры и коренные зубы, предназначенные для фоливорной/просматривающей диеты. Клыки Anoplotheriidae имеют премолярную форму, а это означает, что клыки в целом не отличаются от других зубов, таких как резцы. Нижние премоляры семейства пронзительные и удлиненные. Верхние коренные зубы имеют форму буноселенодонта, а нижние коренные зубы имеют селенодонтные губные клыки и бунодонтальные лингвальные клыки. Подсемейство Anoplotheriinae, типовым родом которого является Anoplotherium , отличается от Dacrytheriinae отсутствием третьего бугорка между метаконидом и энтоконидом на нижних коренных зубах, а также молярообразными премолярами с серповидными параконулами. [44]
Верхние коренные зубы Anoplotherium характеризуются трапециевидными очертаниями на окклюзионных проекциях (или видах зубной эмали сверху), W-образными эктолофами (гребнями или гребнями верхних коренных зубов) и специфическими различиями бугров. Более конкретно, верхние коренные зубы этого рода содержат почти центральные и конические бугорки протоконуса, тесно совмещенные с бугорками мезостиля, конические параконулы, которые соединены с парастилем задними гребнями, а также сжатые парастили и мезостили. Нижние коренные зубы аноплотериид содержат бугорки параконида и метаконида, которые имеют выраженные разделения впадиной между ними. [57] [44]
Позвонки и ребра
[ редактировать ]
Всего у Anoplotherium 7 шейных позвонков серии C1-C7, типичной для большинства млекопитающих. Атлант Lama (C1) подобен атласу верблюдовых, таких как , по форме, а также положению «крыльевых отверстий» в сочетании с соединениями фасеточных суставов, включающими ось (C2). [11] [23] Ось, приписываемая A. commune (но, возможно, также принадлежащая его близкому родственнику Diplobune secundaria ), удлинена в длину и имеет уменьшенный остистый отросток. Позвонки С3-С7 аналогичны Cainotherium . Позвонок C4 выглядит наклоненным, что указывает на изменение ориентации шеи с позвонка C3 на C4 как потенциальный изгиб в передней части шеи, как у современных медведей. Из-за морфологии шейных позвонков у Anoplotherium , вероятно, было наклонное положение шеи вверх. [23]
Аноплотерий также имел 12 грудных позвонков , 6 поясничных позвонков и 3 крестцовых позвонка. Поясничные позвонки, особенно L4–L6, содержат поперечные отростки, широкие, длинные и слегка направленные вперед. Три крестцовых позвонка крепкие и содержат апофизы для прочного прикрепления к длинному хвосту. Позвонки представителей рода аноплотериид устроены так, чтобы совершать типичные движения копытных. [11] [23]
Самым необычным посткраниальным аспектом Anoplotherium по сравнению с другими парнокопытными является длинный и толстый хвост, состоящий из 22 хвостовых позвонков, обеспечивающих сильную мышечную поддержку. Лобные позвонки имели хорошо выраженный отросток, на всех позвонках, кроме самых дальних дистальных, имеются гемальные дуги . [23]
Как и у халикотера Chalicotherium и в отличие от других млекопитающих, таких как козы из рода Ovis и Cainotherium , ребра изгибаются на более широких участках, а их бугорки не так сильно выступают в дорсальном направлении. Ребра Anoplotherium образуют бочкообразный туловище, а это означает, что грудная клетка намного шире, чем у современных жвачных животных. Ребра вообще выступают вбок из-за сильно изогнутого их положения, положения бугорка и выступающих на верхних сторонах грудных позвонков. [23]
Конечности
[ редактировать ]
У Anoplotherium короткие конечности, и считается, что они были негулиградными в положении конечностей, при этом у большинства видов по три пальца как на передних, так и на задних конечностях. A. commune отличается от аналогичных A. latipes своими дидактильными («двупалыми») пальцами, а не трехпалыми («трехпалыми») пальцами. [23] [44] [67]
Передние конечности
[ редактировать ]Лопатка кайму и (или лопатка) имеет выпуклую клювовидную аналогична таковой у Diplobune . Подобно верблюдам ( Camelus ), надостная ямка шире подостной , но у верблюдов лопатки уже, особенно в дистальных (задних) концах надостной ямки. Лопаточная ость крепкая, толстая и постепенно поднимается в высоту в дистальном направлении, пока не достигнет края суставной впадины, как у верблюдов, но в отличие от большинства других современных парнокопытных. Коракоидный отросток (обычно напоминающий небольшую крючковидную структуру) редуцируется до тупого выступа, лишь слегка выступающего. Широкая надостная ямка и широко изогнутый клювовидный край лопаток Anoplotherium не похожи на Cainotherium и Merycoidodon , поскольку у Anoplotherium нет ни треугольной формы лопаток, ни узких надостных ямок. [23]
Морфология локтя Anoplotherium , основанная на структурах и сочленениях локтевых костей, таких как плечевая , лучевая и локтевая кости , демонстрирует свидетельства адаптации к перемещению локтя вверх и вниз при вращениях супинации-пронации максимум на 13°. Полностью выпрямленный локоть может образовывать угол между локтевой костью и плечевой костью, составляющий примерно 135°, что указывает на высокую гибкость по сравнению с другими парнокопытными, включая уже полугибкие локти Cainotherium . [23]
Подобно морфологии запястий свиней рода Sus , копыта Anoplotherium раздвигаются примерно на 16° при движении вниз, что поддерживается морфологией следа. Запястье, возможно, было способно вращаться вверх и вниз, но лишь в ограниченной степени и далеко от гибкой морфологии запястья приматов, что позволяет предположить, что адаптация была не основной особенностью рода парнокопытных, а результатом восстановления примитивного признака. [23]
Запястье состоит из ладьевидной , полулунной , трехгранной и гороховидной костей в первом ряду и трапеции , трапециевидной , головчатой и крючковидной кости во втором ряду. Anoplotherium имеет четырехпалые кости, но кости V пальца, а у A. commune II пальца развиты слабо. [68] Второй палец (пальец II) Anoplotherium не имеет возможности вращения или гибких движений, что означает, что он не играет никакой роли, подобной большому пальцу, как у приматов или гигантской панды . [23]
Задние конечности
[ редактировать ]
, Подвздошная кость часть бедренной кости большого таза , широкая и имеет твердо закругленный гребень подвздошной кости , который под острым углом соединяется с вогнутым нижним краем. Подвздошную кость Anoplotherium можно отличить от Palaeotherium по более короткому телу подвздошной кости, более длинной седалищной кости (нижняя и задняя часть бедренной кости) и более прямому заднему краю таза, что приводит к более длинному лобковому симфизу . Область вертлужной ямки поверхности вертлужной таза большая, ее вертлужная вырезка находится в заднем (или заднем) положении, аналогично таковому у Chalicotherium . [23]
Бедренная кость крупнее голени , имеет только два вертела, как и у других базальных парнокопытных, имеет узкую щель между головкой бедренной кости и большим вертелом и длинную шейку бедренной кости . Вертельная ямка — впадина на поверхности большого вертела, широкая по глубине и узкая по форме, углубляющаяся по бокам. Большеберцовая кость прочная и надежно поддерживает мышечные соединения, основанные на ее гребнях и отростках. Дистальный конец малоберцовой кости плюс выступ медиальной лодыжки большеберцовой кости окружают центральную область таранной кости, чтобы предотвратить ее смещение в сторону. [23]
Аноплотерииды с известными посткраниальными окаменелостями имеют пропорционально широкие, коренастые и скошенные астрагалы (или таранную кость, или лодыжку), что сильно отличается от других парнокопытных. A. latipes частично отличается от A. commune морфологией фасеток и ямок таранной кости, а также более короткой и прочной пяточной костью (пяточной костью). [23] [51] Астрагалы аноплотеров имеют одинаковые уровни возвышения и положения определенных граней с мерикоидодонтами, которых нет у современных парнокопытных, что, возможно, является примером конвергентной эволюции . [69] [70] Медиальная (поддерживающая) грань Anoplotherium и Diplobune вогнутая, контрастирующая с плоской или слегка выпуклой фасеткой Dacrytherium .
Предплюсна состоит из ладьевидной, трех клиновидных костей и кубовидной кости. Стопа A. commune состояла из двух пальцев, о чем свидетельствуют сравнительно небольшие наружная и средняя клиновидные кости. [68]
Следы
[ редактировать ]Крупногабаритные следы с юга Франции и севера Испании, датируемые поздним эоценом. [71] возможно, был из Anoplotherium . Ихнород во Франции в 1980 году . получил название Anoplotheriipus и был впервые описан Полом Элленбергером в департаменте Гар Название рода произошло от того, что ихнотаксон наиболее близок к Anoplotheriidae. Ихногенус диагностирован как принадлежащий к очень крупному парнокопытному, площадь аутопода превышает таковую у A. commune примерно на 33%, расположение двух копыт субпараллельное, а задняя часть педальной подошвы такая же поперечно широкая, как и передняя. подошвы педали. [72] Anoplotheriipus имеет форму от круглой до прямоугольной, с широкими и хорошо выраженными отпечатками раздвоенных пальцев, напоминающими плохо сохранившиеся верблюжьи следы. [73] Подобный парнокопытный ихногенус Diplartiopus отличается от него параллельностью двух пальцев, более удлиненных. [74]
Типовой вид — Anoplotheriipus lavocati , который Элленбергер назвал в честь палеонтолога Рене Лавоката и считал «самым величественным» из трёх ихновидов из-за проявленной специфической подвижности плюсневых костей. Его размеры составляют от 170 мм (6,7 дюйма) до 180 мм (7,1 дюйма) в длину и 120 мм (4,7 дюйма) в ширину, коренастые по форме и расхождение пальцев ног на 12°. Два пальца почти одинаковой длины и имеют размер как минимум 115 мм (4,5 дюйма) без учета плюсневых костей и 225 мм (8,9 дюйма) с плюсневыми костями. Размеры значительно превышают типичные размеры пальцев ног A. commune , которые составляют 85 мм (3,3 дюйма) без плюсневых костей и 170 мм (6,7 дюйма) с ними. [72]
Anoplotheriipus similicommunis , происходящий в этимологии вида от «similis» (похожий на латыни) и A. commune , похож на типовой ихновид, но меньше по размеру и более точно соответствует типичным размерам стопы A. commune по его длине 140 мм (5,5 мм). дюйма) и шириной 105 мм (4,1 дюйма). Угол расхождения между двумя основными пальцами стопы составляет 10°, а минимальная длина пальцев составляет 90 мм (3,5 дюйма) без плюсневых костей и 180 мм (7,1 дюйма) с плюсневыми костями. [72]
Anoplotheriipus Compactus - третий ихновид, который в этимологии вида происходит от латинского слова «compactus», что на английском языке означает «компактный» из-за короткого и закругленного автопода. У него менее точный диагноз по сравнению с двумя другими ихнотаксонами, но он похож по размеру на A. similicommunis и имеет почти круглую подошву педали для поддержки немного более коротких пальцев. Его длина составляет 120 мм (4,7 дюйма), ширина - 100 мм (3,9 дюйма), а длина пальцев составляет 70 мм (2,8 дюйма) - 80 мм (3,1 дюйма) без плюсневых костей и 140 мм (5,5 дюйма) - 150. мм (5,9 дюйма) с. Следы могли быть оставлены A. latipes, хотя ответ пока неясен. [72]
Размер
[ редактировать ]
Виды Anoplotherium были особенно крупными в позднем эоцене, достигая размеров, необычных для большинства групп парнокопытных в палеогене. Оценки крупных размеров начались в 1995 году, когда Мартинес и Судре оценили вес палеогеновых парнокопытных на основе размеров их астрагалов и M 1 зубов . Астрагалы являются обычными костями в комплексах ископаемых из-за их меньшей уязвимости к фрагментации из-за их коренастой формы и компактной структуры, что объясняет их выбор для их использования. Два измерения A. commune дали разные результаты: M 1 дал массу тела 312,075 кг (688,01 фунта), а астрагал - 265,967 кг (586,36 фунта). Эти оценки намного превышают оценки большинства других палеогеновых парнокопытных, участвовавших в исследовании, хотя исследователи отметили, что размеры M 1 могут быть завышены по сравнению с оценкой астрагала. [51]
В 2014 году Такехиса Цубамото пересмотрел взаимосвязь между размером астрагала и предполагаемой массой тела на основе обширных исследований современных наземных млекопитающих, повторно применив методы к палеогеновым парнокопытным, ранее проверенным Судре и Мартинесом. Исследователь использовал линейные измерения и их продукты с скорректированными поправочными коэффициентами. Перерасчеты привели к несколько более низким оценкам по сравнению с результатами 1995 года (за исключением Diplobuneminor , у которого пропорция астрагала короче, чем у большинства других парнокопытных), что показано на графике ниже: [75]

В 2022 году Веппе рассчитал массу тела A. commune , которая составила 360 кг (790 фунтов). [52] В 2023 году Айнара Бадиола и др. По оценкам, вес Anoplotherium колеблется от 115 кг (254 фунта) до 271 кг (597 фунтов). По их расчетам, A. laurillardi был меньшим аноплотериидом, который весил в среднем 157 кг (346 фунтов). A. latipes был крупнее и имел средний вес 229 кг (505 фунтов), а A. commune имеет самый тяжелый вес - 271 кг (597 фунтов). [44]
В 2007 году Хукер оценил размеры A. latipes на основе неполного скелета неполовозрелой особи из пачки Хэмстед формации Боулднор на острове Уайт , Соединенное Королевство. Реконструированный человек уровня 3 в Хэмстеде имел размеры головы и тела 2 м (6 футов 7 дюймов). Плечевая кость незрелой особи Anoplotherium имеет длину 330 мм (13 дюймов), поэтому длина плечевой кости зрелых особей могла составлять около 410 мм (16 дюймов). В результате взрослые A. latipes могли иметь длину головы и тела 2,5 м (8 футов 2 дюйма) и высоту в плечах 1,25 м (4 фута 1 дюйм). Стоя на двух ногах на задних конечностях со спиной, шеей и головой под углом около 15°, особь Хэмстеда уровня 3 могла достигать 2,5 м (8 футов 2 дюйма) при просмотре, в то время как более зрелые особи A. latipes могли стоял чуть более 3 м (9,8 футов). [23]
Палеобиология
[ редактировать ]
С 2007 года Anoplotherium считается четвероногим животным, которое могло стоять на задних лапах как двуногий браузер благодаря сильному тазу, длинному и крепкому хвосту для равновесия и растопыренным задним лапам. Двуногие адаптации демонстрируют некоторые примеры конвергенции с другими животными, такими как халикотерии, различные роды наземных ленивцев , гигантские панды ( Ailuropoda melanoleuca ), гориллы ( Gorilla ) и геренуки ( Litocranius walleri ). В остальном общая форма тела напоминает таковую у Canidae . В результате изгиба шейных позвонков С3-С4 шея и голова могли сохранять горизонтальную ориентацию, стоя на двух ногах. Передние конечности могли выходить за пределы морды горизонтально, пока особь стояла на двух ногах, хотя она не могла дотягиваться вверх и не имела когтей или хватательных органов на кисти, в отличие от Chalicotherium . Следовательно, предплечья могли использоваться не для вырывания и вырывания растений, а в качестве двуногой опоры. Возможно, он бродил, стоя под крутым углом, больше напоминающим геренука, чем Халикотерий . [23]
Его большой размер и способность передвигаться на двух ногах, возможно, дали Anoplotherium несколько источников наземной конкуренции, кроме Palaeotherium magnum большого размера , палеотера с длинной шеей, масса тела которого могла достигать 240,3 кг (530 фунтов). [23] [76] Подвид P. magnum magnum достигал чуть более 2 м (6 футов 7 дюймов) в высоту просмотра в четвероногом положении, и нет никаких доказательств какой-либо адаптации к прямохождению у палеотериев. [23] Anoplotherium , вероятно, участвовал в разделении ниш с палеотериями позднего эоцена и Diplobune . Хотя все они были листоядными браузерами, palaeotheres Plagiolophus и Palaeotherium, возможно, имели небольшую степень плодоядности, в то время как Diplobune , вероятно, был адаптирован к древесному образу жизни . [77] [78] [67] Насколько хорошо Anoplotherium был адаптирован к абразивным листьям и более сухим, но все еще субвлажным условиям в позднем эоцене, малоизвестно, и для ответа требуются будущие исследования в области зубочелюстной системы. [23]

Хукер предположил возможность того, что дидактильные A. commune и A. latipes могли быть половыми диморфами одного и того же вида (в котором A. latipes будет синонимом A. commune ). Между этими двумя видами существует мало постоянных различий в морфологии зубов, причем любые небольшие различия потенциально могут объяснить индивидуальные различия. Различия в количестве пальцев между видами могли отражать A. latipes трехпалость A. commune и двупалость . Палеонтолог объяснил, что, хотя нет никаких доказательств того, что дополнительный палец касался земли во время ходьбы, дополнительный палец A. latipes мог служить дополнительным балансом при движении на двух ногах. [23]
Третий палец мог также служить частью спарринга во внутривидовой конкуренции между особями мужского пола. Однако он отметил, что, несмотря на очевидное «преимущество» A. latipes в прямохождении, нет никаких свидетельств половых различий в пищевом поведении или предпочтениях. Кроме того, оба вида встречаются в одних и тех же местах в Боулдноре в Соединенном Королевстве, а также в Ла-Дебрюж и Монмартре во Франции. Хотя A. latipes более распространен в Ла-Дебрюж, чем на Монмартре, это может быть результатом поведенческих и/или тафономических факторы. [23] [44] Грегуар Метэ выразил неубежденность в том, что третий палец A. latipes является сексуально диморфной адаптацией к прямохождению, вместо этого предположив, что они использовались в спаррингах между мужчинами, если A. latipes и A. commune были половыми диморфами. [67]
Некоторые свидетельства морфологии Anoplotherium подверглись критике со стороны некоторых источников. В своем исследовании морфологии геренука, допускающей прямохождение, исследователи Мэтт Картмилл и Кэй Браун утверждали, что некоторые посткраниальные особенности, которые предположительно были адаптацией Litocranius и других двуногих родов, не отличают геренука от других коров. [79] Киаран Кларк и др. (включая Дж. Дж. Хукера) на основании микроКТ -сканирования обнаружили , что Anoplotherium , являющийся факультативным двуногим браузером, не поддерживается трабекулярной архитектурой проксимальной области бедренной кости. Это могло быть результатом плохих результатов данных микроКТ и меньшего размера выборки, на что данные микроКТ с более высокой контрастностью могут лучше ответить в информации о положении тела. [80]
Следы следов Anoplotheriipus позволяют предположить, что Anoplotherium ходили с очень похожей скоростью передвижения. Судя по группировкам ихнотаксона по следам в местности Фондота в муниципалитете Абьего в Испании, Anoplotherium , возможно, обычно ходил небольшими группами, что может указывать на некоторое стадное (или общительное) поведение. [81]
Палеоэкология
[ редактировать ]Ранняя Европа до Grande Coupure
[ редактировать ]
На протяжении большей части эоцена преобладал тепличный климат с влажной тропической средой и постоянно большим количеством осадков. Современные отряды млекопитающих, включая Perissodactyla, Artiodactyla и Primates (или подотряд Euprimates), появились уже в раннем эоцене, быстро диверсифицируясь и развивая зубные ряды, специализирующиеся на фоливории. Всеядные формы в основном либо перешли на лиственное питание, либо вымерли к среднему эоцену (47–37 млн лет назад) вместе с архаичными « кондилартами ». К позднему эоцену (приблизительно 37–33 млн лет назад) у большинства копытных форм зубные ряды сместились с бунодонтовых бугорков на режущие гребни (т.е. лофы) для фоливорной диеты. [82] [83]
Наземные связи с севером развивающегося Атлантического океана были прерваны около 53 млн лет назад, а это означает, что Северная Америка и Гренландия больше не были хорошо связаны с Западной Европой. С раннего эоцена и до вымирания Гранд-Купюр (56–33,9 млн лет назад) западная часть Евразийского континента была разделена на три массива суши, первые два из которых были изолированы морскими путями: Западная Европа (архипелаг), Балканатолия и восточная часть суши. Евразия (Балканатолия находилась между морем Паратетис на севере и океаном Неотетис на юге). [46] фауна Таким образом , голарктическая млекопитающих Западной Европы была в основном изолирована от других континентов, включая Гренландию, Африку и восточную Евразию, что позволяло эндемизму встречаться в Западной Европе. [83] В результате европейские млекопитающие позднего эоцена (от MP17 до MP20) были в основном потомками эндемичных групп среднего эоцена. [84]
Появление производных аноплотериинов с помощью MP18 произошло спустя много времени после вымирания эндемичного европейского семейства непарнокопытных Lophiodontidae в MP16, включая крупнейшего лофиодонта Lophiodon lautricense , вероятно, в результате перехода от влажной и высокотропической среды к более сухим и умеренным лесам с открытыми территориями. и более абразивная растительность. Выжившие травоядные представители фауны соответствующим образом изменили свои зубные ряды и диетические стратегии, чтобы адаптироваться. [85] [86] Однако окружающая среда все еще была влажной и полна субтропических вечнозеленых лесов. Palaeotheriidae были единственной оставшейся европейской группой непарнокопытных, а плодоядно-листоядные или чисто листоядные парнокопытные стали доминирующей группой в Западной Европе. [87] [88] MP16 также ознаменовал последнее появление большинства европейских крокодиломорфов , из которых аллигатороид Diplocynodon был единственным выжившим из-за, по-видимому, адаптации к общему упадку тропического климата позднего эоцена. [89] [90] [91]
Поздний эоцен
[ редактировать ]
После значительного разрыва в окаменелостях аноплотериинов в MP17a и MP17b, производные аноплотериины Anoplotherium и Diplobune впервые появились в отряде MP18. [49] Они обитали исключительно на западноевропейском архипелаге, но их точное происхождение и пути распространения неизвестны. К тому времени Anoplotherium и Diplobune обитали в Центральной Европе (тогда это был остров) и на Пиренейском полуострове, только первый род из которых позже распространился в южную Англию благодаря MP19 из-за очевидного отсутствия океанских барьеров. [44] [23]
К MP18 Anoplotherium сосуществовал с широким разнообразием парнокопытных в Западной Европе, начиная от более распространенных Dichobunidae , Tapirulidae и Anthracotheriidae и заканчивая многими другими эндемичными семействами, состоящими из Xiphodontidae, Choeropotamidae (однако недавно было установлено, что они полифилетичны), Cebochoeridae , Amphimerycidae , и Каинотерииды. [47] [53] [92] [93] Anoplotherium также сосуществовал с Palaeotheriidae, оставшимся семейством непарнокопытных в Западной Европе. [84] Позднеэоценовые европейские группы клады Ferae представляли преимущественно Hyaenodonta ( Hyaenodontinae , Hyainailourinae и Proviverrinae ), но содержали также Carnivoramorpha ( Miacidae ) и Carnivora (небольшие по размеру Amphicyonidae ). [87] Другие группы млекопитающих, присутствовавшие в позднем эоцене Западной Европы, представляли лепптитидов ( Pseudorhyncocyonidae ), [94] приматы ( Adapoidea и Omomyoidea ), [95] эвлипотифланы ( Nyctitheriidae ), [96] рукокрылые , [83] герпетотерииды , [97] апаторианцы , [98] и эндемичные грызуны ( Pseudosciuridae , Theridomyidae и Gliridae ). [99] Аллигатороид Diplocynodon , присутствующий только в Европе с верхнего палеоцена, также сосуществовал с фаунами до Grande Coupure. [100] Помимо змей, лягушек и саламандр , в Западной Европе известны также богатые комплексы ящериц из MP16-MP20, представляющие Iguanidae , Lacertidae , Gekkonidae , Agamidae , Scincidae , Helodermatidae и Varanoidea . [101]
В местонахождении MP18 в Самбране в Испании A. laurillardi и A. sp. останки были обнаружены у неопределенных групп лягушек и чешуйчатых, аллигатороида Diplocynodon , герпетотериида Peratherium , грызунов ( Theridomys , Elfomys , Pseudoltinomys , Remys ), омомид Microchoerus , carnivoraformes Quercygale и Paramiacis , дихобунида Dichobune , мечедонтов Xiphodon и Haplomeryx . и палеотериев ( Palaeotherium , Leptolophus , иберолоф , пахинолоф , паранчилоф ). [102]
Будучи в то время частью отдельного массива суши, Ла Дебрюж во Франции, датируемый MP18, содержал несколько иные фауны, сосуществовавшие с A. commune , A. latipes и A. laurillardi , а именно герпетотериид Peratherium , грызунов ( Blainvillimys , Theridomys , Плезиарктомис , Гламис ), гиенодонты ( Hyaenodon и Pterodon ), амфициониды Cynodictis , палеотерии ( Plagiolophus , Anchilophus , Palaeotherium ), дихобуниды Dichobune , херопотамиды Choeropotamus , цебохоериды Cebochoerus и Acotherulum , аноплотерииды Dacrytherium и Diplobune . , тапирулид Tapirulus , ксифодонты Xiphodon и Dichodon , каинотер Oxacron , амфимерицида Амфимерикс и антракотер Эломерикс . [103]
Вымирание
[ редактировать ]
Вымирание Гранд-Купюр и смена фауны в Западной Европе, относящееся к самому раннему олигоцену (MP20-MP21), является одним из крупнейших и наиболее резких фаунистических событий в кайнозойской летописи, которое совпадает с событиями, вызывающими изменение климата , более прохладными и более холодными. сезонный климат. [104] Результатом этого события стало 60-процентное вымирание западноевропейских линий млекопитающих, в то время как их заменили иммигранты из азиатской фауны. [105] [106] [107] Палеонтологи часто отмечают Гранд-Купюр как часть границы эоцена и олигоцена в результате 33,9 млн лет назад, хотя по некоторым оценкам, это событие началось 33,6–33,4 млн лет назад. [108] [109] Это событие напрямую коррелирует с переходом эоцен-олигоцен или после него , резким переходом от парникового мира, характеризующего большую часть палеогена, к миру холодных/ледяных камер раннего олигоцена и далее. Массовое падение температур связано с первым крупным расширением антарктических ледяных щитов , которое привело к резкому снижению pCO 2 и, по оценкам, к падению уровня моря примерно на 70 м (230 футов). [110]
Динамика морских путей, в значительной степени отделяющих Западную Европу от других массивов суши, но допускающих некоторые уровни расселения до Гранд-Купюр, сложна и спорна, но многие палеонтологи согласились с тем, что оледенение и вызванное этим падение уровня моря сыграли важную роль в высыхании морские пути ранее выступали в качестве основных препятствий для восточных мигрантов из Балканатолии и Западной Европы. Тургайский пролив часто предлагается в качестве главного европейского морского барьера до Гранд-Купюра, но некоторые исследователи недавно оспорили это мнение, утверждая, что он полностью отступил уже 37 млн лет назад, задолго до перехода от эоцена к олигоцену. Алексис Лихт и др. предположил, что Гранд-Купюр, возможно, мог быть синхронным с оледенением Ои-1 (33,5 млн лет назад), которое фиксирует снижение содержания CO 2 в атмосфере , усиливая оледенение Антарктики, которое уже началось с переходом эоцена-олигоцена. [46] [111]
Событие Гранд-Купюр также ознаменовало большой оборот фауны, ознаменовавший прибытие более поздних антракотериев, энтелодонтов , жвачных животных ( Gelocidae , Lophiomerycidae ), носорогов ( Rinocerotidae , Amynodontidae , Eggysodontidae ), хищных животных (позже Amphicyonidae, Amphicynodontidae , Nimravidae и Ursidae ), восточно-евразийских грызуны ( Eomyidae , Cricetidae и Castoridae ) и эвлипотифланы ( Erinaceidae ). [112] [113] [105] [114]
Эоцен-олигоценовый переход Западной Европы в результате глобальных климатических условий отмечен переходом от тропических и субтропических лесов к более открытым, умеренным или смешанным лиственным местообитаниям с адаптацией к повышенной сезонности. Хотя Anoplotherium просуществовал недолго в раннем олигоцене, существуют разногласия относительно того, пережил ли он Grande Coupure или вымер в результате этого события. [88] [47] Хотя данные указывают на то, что Anoplotherium был истреблен в таких регионах, как Франция и Соединенное Королевство, в результате Великого купюра (последние случаи MP20), [103] [105] [52] восприятие осложняется очевидным последним выживанием A. commune в местонахождении MP21 Мёрен 19 на юге Германии (край западной Европы) вместе с Palaeotherium medium и Diplobune quercyi (немного более молодые местоположения указывают на их вымирание и замену иммигрантами Grande Coupure такие как антракотерий Anthracotherium и носорог Epiaceratherium ). [115]
Хукер отметил, что такие места, как Мёрен 19, относятся к более ранним временам, когда выжившие эндемичные фауны сопровождались некоторыми иммигрантами из Гранд-Купюр, но в остальном к ним еще не присоединились некоторые иммигранты, такие как Anthracotherium . Кроме того, сохранившиеся эндемики этой местности отсутствуют в других районах, датируемых MP21. Таким образом, он утверждал, что некоторые более старые местоположения MP21 с сохранившимися эндемичными фаунами заполняют длинный разрыв между самой молодой пачкой Лоуэр-Хэмстед до Гранд-Купюр и более молодой пачкой Верхний Хэмстед после Гранд-Купюр в формации Боулднор. Эта интерпретация, как объяснил Хукер, означает, что эти места представляли собой очень краткие моменты выживания эндемичных фаун во время Гранд-Купюр, что подтверждает идею крупного и быстрого вымирания фауны и события иммиграции, включая исчезновение Anoplotherium в этом событии. [105] [116]
Вымирание большинства эндемичных парнокопытных, включая Anoplotherium , объясняют конкуренцией с иммигрантской фауной, изменениями окружающей среды из-за похолодания климата или некоторой комбинацией этих двух факторов. [108] Сара К. Джумун и др. определили, что некоторые фауны могли появиться позже и, следовательно, не сыграли роли в вымирании. Они пришли к выводу, что изменение климата, которое привело к усилению сезонности и изменению доступности растительной пищи, привело к тому, что парнокопытные стали неспособными адаптироваться к серьезным изменениям и вымерли. [117] Веппе приводил аналогичные аргументы в пользу того, что изменение климата является основной причиной вымирания вида Гранд-Купюр, утверждая, что похолодание климата вытеснило ранее стабильную субтропическую среду Западной Европы и вызвало коллапс сообщества парнокопытных, которое после своего исчезновения оставило пустые экологические ниши, которые были пассивно заполнены пришлыми фаунами. [52]
Примечания
[ редактировать ]- ^ Французское прилагательное « коммуна» переводится на английский как «общий».
Ссылки
[ редактировать ]- ^ Кювье, Жорж (1804). «Продолжение исследований: О видах животных, от которых происходят ископаемые кости, найденные в гипсовом камне окрестностей Парижа» . Анналы Национального музея естественной истории, Париж (на французском языке). 3 : 364–387. Архивировано из оригинала 27 июля 2023 г. Проверено 30 августа 2023 г.
- ^ Рудвик, Мартин Дж.С. (2022). «Призыв Жоржа Кювье к международному сотрудничеству, 1800 год» . История геологии . 46 (1): 117–125. дои : 10.18814/epiiugs/2022/022002 . S2CID 246893918 .
- ^ Робертс, Джордж (1839). Этимологический и толковый словарь терминов и языка геологии . Лондон: Лонгман, Орм, Браун, Грин и Лонгманс. п. 8 . Проверено 29 декабря 2021 г.
- ^ Кювье, Жорж (1804). «Продолжение исследований: Продолжение исследований ископаемых костей гипсового камня из окрестностей Парижа. Третье воспоминание. Реституция стоп. Первый раздел. Реституция различных задних лап» . Анналы Национального музея естественной истории, Париж (на французском языке). 3 : 442–472. Архивировано из оригинала 27 июля 2023 г. Проверено 30 августа 2023 г.
- ^ Кювье, Жорж (1805). «Третье воспоминание. Второй раздел. Восстановление разных передних лап» . Анналы Национального музея естественной истории, Париж (на французском языке). 6 :253–283. Архивировано из оригинала 10 ноября 2012 г. Проверено 30 августа 2023 г.
- ^ Кювье, Жорж (1807). «Продолжение исследований ископаемых костей окрестностей Парижа. Третья память, третий отдел, фаланги. Четвертая память, на костях конечностей, первый отдел, длинные кости задних конечностей» . Летопись Музея естественной истории . 9 :10–44. Архивировано из оригинала 02 сентября 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б Кювье, Жорж (1807). «Продолжение исследований ископаемых костей в окрестностях Парижа. В. и память, II и раздел, описание двух почти полных скелетов Anoplotherium commune» . Annales du Muséum d'Histoire Naturelle (на французском языке). 9 : 272–282. Архивировано из оригинала 02 сентября 2023 г. Проверено 30 августа 2023 г.
- ^ Белхосте, Бруно (2017). «Глава 10: От карьера к бумаге. Три эпистемологические культуры Кювье». В Чемле, Карин; Келлер, Эвелин Фокс (ред.). Культуры без культурализма: создание научных знаний . Издательство Университета Дьюка. стр. 250–277.
- ^ Уоллес, Дэвид Рейнс (2004). «Глава 1: Толстокожие в катакомбах». Звери Эдема: ходячие киты, утренние лошади и другие загадки эволюции млекопитающих . Издательство Калифорнийского университета. стр. 1–13.
- ^ Аллеманд, Реми (2017). Эндокраниальное микротомографическое исследование морских рептилий (Plesiosauria и Mosasauroidea) турона (поздний мел) Марокко: палеобиологические и поведенческие последствия (доктор философии). Национальный музей естественной истории, Франция.
- ^ Jump up to: а б с д Кювье, Жорж (1822). Исследования ископаемых костей, где восстановлены характеристики нескольких животных, виды которых были уничтожены революциями земного шара . Полет. 3. Ж. Дюфур и Э. д’Окань. Архивировано из оригинала 19 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б с д и ж Орлиак, Маева Дж.; Могуст, Джейкоб; Балькарсель, Ана; Гилиссен, Эммануэль (2023). «Палеоневрология парнокопытных, обзор эволюции мозга парнокопытных» (PDF) . В Дозо; Паулина-Карабахал, Ариана; Макрини, Томас Э.; Уолш, Стиг (ред.). Палеоневрология амниотов . Спрингер Чам. стр. 507–555. дои : 10.1007/978-3-031-13983-3_13 . ISBN 978-3-031-13982-6 . Архивировано (PDF) из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Муди, Рой Ли (1917). «Исследования по палеопатологии. I. Общее рассмотрение доказательств патологических состояний, обнаруженных среди ископаемых животных» . Анналы истории медицины . 1 (4): 374–393. ПМЦ 7927727 . ПМИД 33943144 .
- ^ Дьегес, Кармен; Исидро, Альберт; Мальгоса, Ассумпсьо (1996). «Введение в зоопалеопатологию и обновленную информацию об ископаемых фитопалеопатологиях из Испании» . Журнал палеопатологии . 8 (3): 133–142.
- ^ Томас, Ричард (2012). «Глава 66: Нечеловеческая палеопатология». В Буйкстре, Джейн; Робертс, Шарлотта (ред.). Глобальная история палеопатологии: пионеры и перспективы . Издательство Оксфордского университета. стр. 652–664. doi : 10.1093/acprof:osobl/9780195389807.003.0066 .
- ^ Кювье, Жож (1812). «Общий обзор и обнаружение скелетов различных видов». Исследование ископаемых костей четвероногих: мы восстанавливаем характеристики нескольких видов животных, которые, по-видимому, были уничтожены мировыми революциями (на французском языке). Полет. 3. В Детервилле. Архивировано из оригинала 31 июля 2023 г. Проверено 30 августа 2023 г.
- ^ Рудвик, Мартин Дж.С. (1997). «Глава 6: Животные из гипсовых пластов Парижа». Жорж Кювье, Ископаемые кости и геологические катастрофы: новые переводы и интерпретации первичных текстов . Издательство Чикагского университета.
- ^ Мануччи, Фабио; Романо, Марко (2022). «Обзор иконографии и центральной роли «палеоискусства»: четыре столетия геопалеонтологического искусства». Историческая биология . 35 (1): 1–48. дои : 10.1080/08912963.2021.2017919 . S2CID 246054069 .
- ^ Рудвик, Мартин Дж.С. (1992). «Глава 2: Замочные скважины в прошлое». Сцены из глубин времени: ранние графические изображения доисторического мира . стр. 27–58.
- ^ Jump up to: а б Жерве, Поль (1848–1852). «Примечание о роде Eurytherium, за которым следует сравнительный список млекопитающих, наблюдавшихся в бассейнах Парижа и Апта, и замечания о копытных, наблюдавшихся во Франции». Французская зоология и палеонтология (позвоночные животные): или новые исследования живых и ископаемых животных Франции . Полет. 2. Артюс Бертран. Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б Шлоссер, Макс (1883). «Обзор известных Anoplotheria и Diplobunena, а также объяснение взаимоотношений между Anoplotherium и другими семействами млекопитающих» . Новогодний ежегодник по минералогии, геологии и палеонтологии, трактаты . 2 . Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Дор, М. (1938). «О биологии аноплотерия (был ли аноплотерий водным?)». Млекопитающие . 2 :43–48.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д р с т в v В х и Хукер, Джерри Дж. (2007). «Двуногие адаптации необычного тилопода Anoplotherium позднего эоцена – раннего олигоцена (Artiodactyla, Mammalia)» . Зоологический журнал Линнеевского общества . 151 (3): 609–659. дои : 10.1111/j.1096-3642.2007.00352.x .
- ^ Давыденко, Свитозар; Гольдин, Павел; Босселерс, Марк; Вальдик, Бернд; Влит, Хенк Ян ван (2023). «Массовая и микроскопическая анатомия большеберцовой кости, предположительно приписываемая китообразным из среднего эоцена Европы, с примечанием о парнокопытном Anoplotherium и о непарнокопытном Lophiodon». Палеонтологическая газета . 97 (3): 627–652. Бибкод : 2023PalZ...97..627D . дои : 10.1007/s12542-023-00653-x . S2CID 259897461 .
- ^ Филлипс, Сэмюэл; Шентон, Фрэнсис Кингстон Джон (1860). Путеводитель по Хрустальному дворцу и парку . Архивировано из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б Виттон, Марк П.; Мишель, Эллинор (2022). «Глава 4: Скульптуры: млекопитающие». Искусство и наука динозавров Хрустального дворца . Кровуд Пресс. стр. 68–91.
- ^ Жерве, Поль (1848–1852). «Различные виды ископаемых копытных». Французская зоология и палеонтология (позвоночные животные): или новые исследования живых и ископаемых животных Франции . Полет. 2. Артюс Бертран. Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б с д Лидеккер, Ричард (1885). Каталог ископаемых млекопитающих в Британском музее (Естественная история): Часть II. Включает отряд Ungulata, подотряд Artiodactyla . Орден Попечителей, Лондон. Архивировано из оригинала 02 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Сен-Илер, Этьен Жоффруа (1833). «Рассуждения об ископаемых костях, наиболее неизвестных, найденных и наблюдаемых в Оверни». Энциклопедический обзор, или аргументированный анализ наиболее замечательных произведений . Полет. 59. Центральное бюро Энциклопедического обозрения. Архивировано из оригинала 25 августа 2023 г. Проверено 19 сентября 2023 г.
- ^ Jump up to: а б Помель, Огюст (1851). «Новые наблюдения над строением стоп у животных семейства Anoplotherium и рода Hyaemoschus» . Еженедельные отчеты сессий Академии наук . 33 :16–17. Архивировано из оригинала 03 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Берте, Дидье (2003). «Род Cainotherium (Mammalia, Artiodactyla): морфометрическое изучение, систематический пересмотр, эволюционные и палеогеографические последствия, вымирание» . Работы и документы геологических лабораторий Лиона . 159 (159): 3–205. Архивировано из оригинала 06 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Ларте, Эдуард (1837). «Обратите внимание на ископаемые кости из третичных стоянок Симорре, Сансана и т. д. в департаменте Жер, а также на недавнее открытие ископаемой челюсти обезьяны» . Доклады Академии наук . 4 :85–93. Архивировано из оригинала 13 июня 2020 г. Проверено 30 августа 2023 г.
- ^ Ларте, Эдуард (1839). «Геологическое извещение». Выписка из Директории департамента Жер .
- ^ Анкетен, Жереми; Антуан, Пьер-Оливье; Тасси, Паскаль (2007). «Средний миоцен Chalicotheriinae (Mammalia, Perissodactyla) из Франции, с обсуждением филогении халикотериинов» . Зоологический журнал Линнеевского общества . 151 (3): 577–608. дои : 10.1111/j.1096-3642.2007.00327.x .
- ^ Коутли, Проби Т.; Фальконер, Хью (1837). «Извещение об останках ископаемой обезьяны из третичных слоев холмов Севалик на севере Индостана» . Труды Геологического общества : 499–504. Архивировано из оригинала 02 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Лидеккер, Ричард (1886). Каталог ископаемых млекопитающих в Британском музее (Естественная история): Часть III. Включает отряд Ungulata, подотряды Perissodactyla, Toxondontia, Condylarthra и Amblypoda . Орден Попечителей, Лондон. Архивировано из оригинала 02 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Кольбер, Эдвин Х. (1935). «Правильное использование общего названия Nestoritherium». Журнал маммологии . 16 (3): 233–234. дои : 10.1093/jmammal/16.3.233 .
- ^ Баффето, Эрик (2016). «От комментариев Чарльза Дарвина к первому упоминанию гигантских ископаемых птиц Южной Америки: каталог ископаемых видов из Аргентины Огюста Бравара (1860 г.) и его значение». Бюллетень геологического общества Франции . 187 (1): 41–53. дои : 10.2113/gssgfbull.187.1.41 .
- ^ Филхол, Анри (1877). «Исследование фосфоритов Керси. Изучение найденных там окаменелостей, особенно млекопитающих» . Анналы геологических наук Парижа . Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Рютимейер, Людвиг (1862). «Эокеновые млекопитающие из региона швейцарской Юры» . Новые меморандумы Швейцарского общества естественных исследований . 19 :1–98. Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ фон Фраас, Оскар Фридрих (1870). «Диплобуне баварикум» . Палеонтографика . 17 : 177–184. Архивировано из оригинала 4 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Штелин, Ганс Георг (1910). "Млекопитающие Швейцарского Эокена. Шестая часть: Катодонтерий - Дакритерий - Лептотерий - Аноплотерий - Диплобуна - Ксифодон - Псевдамфимерикс - Амфимерикс - Диходон - Гапломерикс - Тапирулус - Гелокус. Дополнения, Artiodactyla incertae sedis, окончательные наблюдения над парнокопытными, дополнения к Периссодактилен» . Трактаты Швейцарского палеонтологического общества . 36 . Архивировано из оригинала 05 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Дитрих, Вильгельм Отто (1922). «Вклад в знания о формации бобовых руд, содержащих млекопитающих, в Швабии. 1. Забытое, недавно разработанное пещерное месторождение земного эоцена на Ульм-Альбе». Центральный журнал минералогии, геологии и палеонтологии . 19 :209-224.
- ^ Jump up to: а б с д и ж г час я дж к л м н Бадиола, Айнара; Де Викунья, Наья Хименес; Пералес-Гогенола, Лейре; Гомес-Оливенсия, Асье (2023). «Первые явные свидетельства существования Anoplotherium (Mammalia, Artiodactyla) на Пиренейском полуострове: обновленная информация об иберийских аноплотериинах» . Анатомические записи: достижения в интегративной анатомии и эволюционной биологии . дои : 10.1002/ar.25238 . ПМИД 37221992 . S2CID 258864256 .
- ^ де Бонис, Луи (1964). «Изучение некоторых млекопитающих Лудиана Ла Дебрюж (Воклюз)». Анналы палеонтологии : 121–154.
- ^ Jump up to: а б с Лихт, Алексис; Метэ, Грегуар; Костер, Полина; Ибилиоглу, Дениз; Оджакоглу, Фарук; Вестервил, Ян; Мюллер, Меган; Кэмпбелл, Клей; Мэттингли, Спенсер; Вуд, Мелисса С.; Борода, К. Кристофер (2022). «Балканатолия: островная биогеографическая провинция млекопитающих, которая частично проложила путь к Гранд-Купюру» . Обзоры наук о Земле . 226 : 103929. Бибкод : 2022ESRv..22603929L . doi : 10.1016/j.earscirev.2022.103929 .
- ^ Jump up to: а б с д Эрфурт, Йорг; Метэ, Грегуар (2007). «Эндемичные европейские палеогеновые парнокопытные». В Протеро, Дональд Р.; Фосс, Скотт Э. (ред.). Эволюция парнокопытных . Издательство Университета Джонса Хопкинса. стр. 59–84.
- ^ Орлиак, Маева; Гилиссен, Эммануэль (2012). «Виртуальный эндокраниальный слепок ранних эоценовых диакодексов (парнокопытные, млекопитающие) и морфологическое разнообразие мозга ранних парнокопытных» . Труды Королевского общества Б. 279 (1743): 3670–3677. дои : 10.1098/rspb.2012.1156 . ПМЦ 3415922 . ПМИД 22764165 .
- ^ Jump up to: а б с д Куэста, Мигель-Анхель; Бадиола, Айнара (2009). «Duerotherium sudrei gen. et sp. nov., новый аноплотиевый парнокопытный из среднего эоцена Пиренейского полуострова» . Журнал палеонтологии позвоночных . 29 (1): 303–308. Бибкод : 2009JVPal..29..303C . дои : 10.1671/039.029.0110 . JSTOR 20491092 . S2CID 55546022 . Архивировано из оригинала 10 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Шмидт-Киттлер, Норберт; Годино, Марк; Францен, Йенс Л.; Хукер, Джереми Дж. (1987). «Европейские контрольные уровни и таблицы корреляции». Мюнхенские геонаучные трактаты А10 . Пфайль Верлаг, Мюнхен. стр. 13–31.
- ^ Jump up to: а б с Судре, Жан; Мартинес, Жан-Ноэль (1995). «Астрагал палеогеновых парнокопытных: сравнительная морфология, изменчивость и прогноз массы тела». Летайя . 28 (3): 197–209. Бибкод : 1995Лета..28..197М . дои : 10.1111/j.1502-3931.1995.tb01423.x .
- ^ Jump up to: а б с д и ж Веппе, Ромен (2022). Упадок эндемичных европейских парнокопытных, вскрытие вымирания (Диссертация) (на французском языке). Университет Монпелье. Архивировано из оригинала 11 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Jump up to: а б Луччизано, Винсент; Судре, Жан; Лихоро, Фабрис (2020). «Ревизия эоценовых парнокопытных (Mammalia, Placentalia) из Аумеласа и Сен-Мартен-де-Лондра (известняки Монпелье, Эро, Франция) ставит под сомнение раннюю европейскую радиацию парнокопытных». Журнал систематической палеонтологии . 18 (19): 1631–1656. Бибкод : 2020JSPal..18.1631L . дои : 10.1080/14772019.2020.1799253 . S2CID 221468663 .
- ^ Jump up to: а б Веппе, Ромен; Блондель, Сесиль; Виани-Лио, Моник; Эскаргюэль, Жиль; Пелисье, Тьерри; Антуан, Пьер-Оливье; Орлиак, Маэва Юдит (2020). «Cainotheriidae (Mammalia, Artiodactyla) из Дамса (Кверси, юго-запад Франции): филогенетические взаимоотношения и эволюция в период перехода эоцена в олигоцен (MP19 – MP21)» (PDF) . Журнал систематической палеонтологии . 18 (7): 541–572. Бибкод : 2020JSPal..18..541W . дои : 10.1080/14772019.2019.1645754 . S2CID 202026238 . Архивировано (PDF) из оригинала 07 марта 2022 г. Проверено 19 сентября 2023 г.
- ^ Веппе, Ромен; Блондель, Сесиль; Виани-Лио, Моник; Пелисье, Тьерри; Орлиак, Маэва Юдит (2020). «Новая Cainotherioidea (Mammalia, Artiodactyla) из Паламбера (Кверси, юго-запад Франции): филогенетические взаимоотношения и история эволюции зубного ряда Cainotheriidae» . Электронная палеонтология (23(3):a54). дои : 10.26879/1081 . S2CID 229490410 .
- ^ Джентри, Алан В.; Хукер, Джерри Дж. (1988). «Филогения парнокопытных». Филогения и классификация четвероногих: Том 2: Млекопитающие (специальный том Ассоциации систематики, № 35B) . Издательство Оксфордского университета. стр. 235–272.
- ^ Jump up to: а б с фон Циттель, Карл Альфред (1925). Шлоссер, Макс (ред.). Учебник палеонтологии. Том III. Млекопитающие . Макмиллан и Ко. Лимитед. стр. 179–180. Архивировано из оригинала 14 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Беддард, Фрэнк Эверс (1902). Хармер, Сидни Фредерик; Шипли, Артур Эверетт (ред.). Кембриджская естественная история: млекопитающие . Макмиллан и Ко. Лимитед. стр. 332–333. Архивировано из оригинала 17 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Скотт, Уильям Б. (1945). «Млекопитающие олигоцена реки Дюшен». Труды Американского философского общества . 34 (3): 209–253. дои : 10.2307/1005542 . JSTOR 1005542 .
- ^ Jump up to: а б с д и ж г Палмер, RW (1913). «Мозг и черепная коробка ископаемого копытного рода Anoplotherium» . Труды Лондонского зоологического общества . 83 (4): 878–893. дои : 10.1111/j.1096-3642.1913.tb01994.x . Архивировано из оригинала 13 октября 2023 г. Проверено 19 сентября 2023 г.
- ^ Пирсон, Хельга Шарп (1927). «О черепах ранних третичных свиней вместе с описанием слуховой области некоторых других примитивных парнокопытных» . Философские труды Лондонского королевского общества. Серия Б, содержащая статьи биологического характера . 215 (421–430): 440–445. дои : 10.1098/rstb.1927.0009 .
- ^ Гагнейсон, Сирил; Леру, Жан-Жак (2013). «Череп Diplobune secundaria Cuvier, 1822 год из Сен-Капрез-д'Эйме (Дордонь)» . Симбиозы (на французском языке). 29 : 43–46.
- ^ Jump up to: а б Джерисон, Гарри Дж. (2009). «Глава 10: Как окаменелости могут рассказать нам об эволюции неокортекса?». В Кассе, Джон Х.; Стридтер, Георг Ф.; Рубинштейн, Джон Л.Р.; Буллок, Теодор Х.; Крубицер, Лия; Пройсс, Тодд (ред.). Эволюционная нейронаука . Академическая пресса. стр. 497–508.
- ^ Эдингер, Тилли (1948). «Эволюция лошадиного мозга» . 25: Эволюция мозга лошади . Мемуары Геологического общества Америки. Том. 25. Геологическое общество Америки. стр. 1–178. дои : 10.1130/MEM25-p1 . Архивировано из оригинала 13 октября 2023 г. Проверено 19 сентября 2023 г.
- ^ Тьери, Гислен; Дюкрок, Стефан (2015). «Эндокасты и эволюция мозга у Anthracotheriidae (Artiodactyla, Hippopotamoidea)» . Журнал анатомии . 227 (3): 277–285. дои : 10.1111/joa.12348 . ПМК 4560562 . ПМИД 26278931 .
- ^ Лихоро, Фабрис; Буассери, Жан-Рено; Вирио, Лоран; Брюне, Мишель (2006). «Антракотерийная дентальная анатомия показывает чадо-ливийскую биопровинцию позднего миоцена» . Труды Национальной академии наук . 103 (23): 8763–8767. Бибкод : 2006PNAS..103.8763L . дои : 10.1073/pnas.0603126103 . ПМЦ 1482652 . ПМИД 16723392 .
- ^ Jump up to: а б с Метэ, Грегуар (2014). На «большом пальце» аноплотериинов: сравнительное 3D-исследование руки Anoplotherium и Diplobune . Швейцарская встреча по геонаукам, 2014 г.
- ^ Jump up to: а б Абуш-Зиверт, Сюзанна (1989). «Заметки об аноплотериях (парнокопытных, млекопитающих) парижских гипсов». Мюнхенские геонаучные трактаты (А) . 15 :55–78.
- ^ Хейссиг, Курт (1993). «Астрагал у аноплотеров и ореодонтов, филогенетические и палеогеографические последствия». Каупия . 3 : 173–178.
- ^ Протеро, Дональд Р.; Фосс, Скотт Э., ред. (2007). "Краткое содержание". Эволюция парнокопытных . Издательство Университета Джонса Хопкинса. п. 307.
- ^ де Карвальо, Карлос Нето; Муньис, Фернандо; Касерес, Луис М.; Белаустеги, Зейн; Родригес-Видаль, Хоакин; Прекрасно, Жуан; Морейра, Ноэль; Качао, Марио; Кунья, Педро П.; Фигейредо, Сильверио; и др. (2022). «В позднем плейстоцене зубры кочевали вдоль юго-западного побережья Андалусии (Испания)» . Научные отчеты . 12 (9911): 9911. Бибкод : 2022NatSR..12.9911D . дои : 10.1038/s41598-022-14137-6 . ПМЦ 9198092 . ПМИД 35701579 .
- ^ Jump up to: а б с д Элленбергер, Пол (1980). «О следах крупных млекопитающих из верхнего эоцена Гарриг-ст-Эулали (Гар)» . Палеовертебрата . 9 :37–78. Архивировано из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Лукас, Спенсер Г.; Хант, Адриан П. (2007). «Ихнотаксономия верблюжьих следов». В Лукасе, Спенсер Г.; Спилманн, Джастин А.; Локли, Мартин Г. (ред.). Следы и следы кайнозойских позвоночных: Бюллетень 42 . Музей естественной истории и науки Нью-Мексико.
- ^ Монтес, Мартин Линарес (2020). Палеонтология и экологическая интерпретация ихнитов млекопитающих из стоянки Фондота (Палеоген, Абьего, Уэска) (PDF) (Диссертация). Университет Сарагосы. Архивировано (PDF) из оригинала 19 октября 2023 г. Проверено 30 августа 2023 г.
- ^ Цубамото, Такехиса (2014). «Оценка массы тела по таранной кости у млекопитающих» . Acta Palaeontologica Polonica . 59 (2): 259–265. дои : 10.4202/app.2011.0067 . S2CID 54686160 .
- ^ Макларен, Джейми Александр; Наувелартс, Сандра (2020). «Современные тапиры как морфофункциональные аналоги передвижения эндемичных эоценовых европейских периссодактилей». Журнал эволюции млекопитающих . 27 (2): 245–263. дои : 10.1007/s10914-019-09460-1 . hdl : 10067/1580640151162165141 . S2CID 254703475 .
- ^ Джумун, Сара С.; Хукер, Джерри Дж.; Коллинсон, Маргарет Э. (2009). Различия в диетических реакциях Perissodactyl Plagiolophus и парнокопытных Diplobune на переходные события эоцена/олигоцена в Европе (PDF) . 69-е ежегодное собрание Общества палеонтологии позвоночных и 57-й симпозиум палеонтологии и сравнительной анатомии позвоночных (SVPCA). Том. 29. Архивировано (PDF) из оригинала 31 мая 2023 г. Проверено 30 августа 2023 г.
- ^ Джумун, Сара С.; Хукер, Джерри Дж.; Коллинсон, Маргарет Э. (2010). «Изменения в износе зубов Plagiolophus Minor (Mammalia: Perissodactyla) в период эоцен-олигоценового перехода» . Журнал палеонтологии позвоночных . 30 (2): 563–576. Бибкод : 2010JVPal..30..563J . дои : 10.1080/02724631003618124 . JSTOR 40666176 . S2CID 86429890 . Архивировано из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Картмилл, Мэтт; Браун, Кэй (2017). «Осанка, передвижение и двуногость: случай геренука (Litocranius walleri)». В Мароме – Асаф; Ховерс, Эрелла (ред.). Палеонтология и предыстория человека . Палеобиология и палеоантропология позвоночных. Спрингер, Чам. стр. 53–70. дои : 10.1007/978-3-319-46646-0_6 . ISBN 978-3-319-46644-6 .
- ^ Кларк, Кьяран; Дженис, Кристин М.; Хукер, Джерри Дж.; Рэйфилд, Эмили Дж. (2018). Положение в летописи окаменелостей: можно ли использовать микроструктуру кости для более точной реконструкции поведения в прошлом? (PDF) . Палеонтологическая ассоциация: 62-е ежегодное собрание; 14–17 декабря 2018 г.; Бристольский университет. п. 72. Архивировано (PDF) из оригинала 18 июня 2022 г. Проверено 30 августа 2023 г.
- ^ Линарес Монтес, Мартин; Канудо, Хосе Игнасио; Лусон, Мария Арансасу; Кастанера, Диего (2021). «Исключительная палеоихнологическая запись парнокопытных в нижнем олигоцене Абьего (Уэска, Испания)» (PDF) . Геологические коммуникации (на испанском языке). 108 : 103–107. дои : 10.34637/6f8b-cj24 . Архивировано (PDF) из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Эронен, Юсси Т.; Дженис, Кристин М.; Чемберлен, Чарльз Пейдж; Мульч, Андреас (2015). «Поднятие гор объясняет различия в палеогеновых закономерностях эволюции и вымирания млекопитающих между Северной Америкой и Европой» . Труды Королевского общества Б. 282 (1809). дои : 10.1098/rspb.2015.0136 . ПМК 4590438 . ПМИД 26041349 .
- ^ Jump up to: а б с Мэтр, Элоди (2014). «Западноевропейские рукокрылые среднего эоцена-раннего олигоцена: систематика, филогения и палеоэкология на основе нового материала из Керси (Франция)» . Швейцарский журнал палеонтологии . 133 (2): 141–242. Бибкод : 2014SwJP..133..141M . дои : 10.1007/s13358-014-0069-3 . S2CID 84066785 .
- ^ Jump up to: а б Бадиола, Айнара; Пералес-Гогенола, Лейре; Астибия, Умберто; Субербиола, Хавьер Переда (2022). «Синтез эоценовых экоидов (Perissodactyla, Mammalia) с Пиренейского полуострова: новые признаки эндемизма». Историческая биология . 34 (8): 1623–1631. Бибкод : 2022HBio...34.1623B . дои : 10.1080/08912963.2022.2060098 . S2CID 248164842 .
- ^ Робине, Селин; Реми, Жан Альбер; Лоран, Ив; Данило, Лора; Лихоро, Фабрис (2015). «Новый род Lophiodontidae (Perissodactyla, Mammalia) из раннего эоцена Ла Бори (Южная Франция) и происхождение рода Lophiodon Cuvier, 1822». Геобиос . 48 (1): 25–38. Бибкод : 2015Geobi..48...25R . дои : 10.1016/j.geobios.2014.11.003 .
- ^ Пералес-Гогенола, Лейре; Бадиола, Айнара; Гомес-Оливенсия, Азиер; Переда-Субербиола, Хавьер (2022). «Замечательный новый палеотериид (Mammalia) в эндемичной непарнокопытной фауне Иберийского эоцена». Журнал палеонтологии позвоночных . 42 (4). Бибкод : 2022JVPal..42E9447P . дои : 10.1080/02724634.2023.2189447 . S2CID 258663753 .
- ^ Jump up to: а б Соле, Флореаль; Фишер, Валентин; Ле Верже, Кевин; Меннекар, Бастьен; Спейер, Роберт П.; Пенье, Стефан; Смит, Тьерри (2022). «Эволюция европейских хищных млекопитающих в палеогене». Биологический журнал Линнеевского общества . 135 (4): 734–753. doi : 10.1093/biolinnean/blac002 .
- ^ Jump up to: а б Блондель, Сесиль (2001). «Эоцен-олигоценовые копытные Западной Европы и их среда обитания» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 168 (1–2): 125–139. Бибкод : 2001PPP...168..125B . дои : 10.1016/S0031-0182(00)00252-2 . Архивировано (PDF) из оригинала 22 августа 2017 г. Проверено 30 августа 2023 г.
- ^ Мартин, Джереми Э.; Поша-Котийу, Йохан; Лоран, Ив; Перье, Винсент; Роберт, Эммануэль; Антуан, Пьер-Оливье (2022). «Анатомия и филогения исключительно крупного себецида (Crocodylomorpha) из среднего эоцена юга Франции». Журнал палеонтологии позвоночных . 42 (4). Бибкод : 2022JVPal..42E3828M . дои : 10.1080/02724634.2023.2193828 . S2CID 258361595 .
- ^ Мартин, Джереми Э. (2015). «Себекозухий в карсте среднего эоцена с комментариями на спинном щите Crocodylomorpha» . Acta Palaeontologica Polonica . 60 (3): 673–680. дои : 10.4202/app.00072.2014 . S2CID 54002673 .
- ^ Антунес, Мигель Теллес (2003). «Нижнепалеогеновые крокодилы из Силвейринья, Португалия» . Палеовенебрата . 32 : 1–26. Архивировано из оригинала 29 августа 2023 г. Проверено 30 августа 2023 г.
- ^ Бай, Бин; Ван, Юань-Цин; Теодор, Джессика М.; Мэн, Джин (2023). «Маленькие парнокопытные с тапироподобными зубами из среднего эоцена Эрляньской котловины, Внутренняя Монголия, Китай» . Границы в науках о Земле . 11 :1–20. Бибкод : 2023FrEaS..1117911B . дои : 10.3389/feart.2023.1117911 .
- ^ Костопулос, Димитрис С.; Куфос, Джордж Д.; Кристанис, Кимон (2012). «Останки некоторых антракотериид (Artiodactyla, Mammalia) из северной Греции: комментарии к палеозоогеографии и филогении Elomeryx». Швейцарский журнал палеонтологии . 131 (2): 303–315. Бибкод : 2012SwJP..131..303K . дои : 10.1007/s13358-012-0041-z . S2CID 195363034 .
- ^ Хукер, Джерри Дж. (2013). «Происхождение и эволюция Pseudorhyncocyonidae, европейского палеогенового семейства насекомоядных плацентарных млекопитающих» . Палеонтология . 56 (4): 807–835. Бибкод : 2013Palgy..56..807H . дои : 10.1111/пала.12018 . S2CID 84322086 .
- ^ Мариго, Юдит; Сюзанна, Иветт; Минвер-Баракат, Раф; Малапейра, Джоан Мадурелл; Мойя-Сола, Сальвадор; Казановас-Вилар, Исаак; Хименес, Хосе Мария Роблес; Альба, Дэвид М. (2014). «Летопись окаменелостей приматов на Пиренейском полуострове» . Журнал иберийской геологии . 40 (1): 179–211. дои : 10.5209/rev_JIGE.2014.v40.n1.44094 .
- ^ Манц, Карли; Блох, Джонатан Иван (2014). «Систематика и филогения палеоцен-эоцена Nyctitheriidae (Mammalia, Eulipotyphla?) с описанием нового вида из позднего палеоцена бассейна Кларкс-Форк, Вайоминг, США». Журнал эволюции млекопитающих . 22 (3): 307–342. дои : 10.1007/s10914-014-9284-3 . S2CID 254704409 .
- ^ Бадиола, Айнара; Куэста, Мигель-Анхель (2006). «Сумчатые из верхнеэоценовой стоянки Замбрана (Алава, Баскско-Кантабрийский регион)» . Геологические исследования (на испанском языке). 62 (1): 349–358. дои : 10.3989/egeol.0662130 .
- ^ Сиже, Бернар (1997). «Насекомоядные млекопитающие из новых коллекций Соссиса и связанных с ним памятников (верхний эоцен, Испания)». Геобиос . 30 (1): 91–113. Бибкод : 1997Geobi..30...91S . дои : 10.1016/S0016-6995(97)80260-4 .
- ^ Доусон, Мэри Р. (2003). «Палеогеновые грызуны Евразии». Распространение и миграция третичных млекопитающих Евразии . Том. 10. С. 97–127.
- ^ Хроуст, Милан; Мазух, Мартин; Лухан, Анхель Эрнандес (2019). «Новый крокодиловый материал из эоцен-олигоценового перехода Северо-Западной Богемии (Чехия): обновленная летопись окаменелостей в Центральной Европе во время Гранд-Купюр». Новогодний ежегодник геологии и палеонтологии - Трактаты . 293 (1): 73–82. дои : 10.1127/njgpa/2019/0832 . S2CID 199104151 .
- ^ Ярость, Жан-Клод (2012). «Земноводные и чешуйчатые в эоцене Европы: о чем они нам говорят?». Палеобиоразнообразие и палеосреда . 92 (4): 445–457. Бибкод : 2012ПдПе...92..445Р . дои : 10.1007/s12549-012-0087-3 . S2CID 128651937 .
- ^ Минвер-Баракат, Раф; Бадиола, Айнара; Мариго, Юдит; Моя-Сола, Сальвадор (2013). «Первая запись рода Microchoerus (Omomyidae, Primates) на западе Пиренейского полуострова и ее палеобиогеографическое значение». Журнал эволюции человека . 65 (3): 313–321. Бибкод : 2013JHumE..65..313M . дои : 10.1016/j.jhevol.2013.07.002 . ПМИД 23916791 .
- ^ Jump up to: а б Агилар, Жан-Пьер; Лежандр, Серж; Мишо, Жак (1997). «Таблицы синтеза и корреляции». Материалы конгресса Био-хром'97. Мемуары и работы Института EPHE в Монпелье 21 (на французском языке). Практическая школа перспективных исследований в области наук о жизни и Земле, Монпелье. стр. 769–850.
- ^ Сун, Чимин; Ни, Сицзюнь; Би, Шуньдун; Ву, Вэньюй; Да, Цзе; Мэн, Джин; Виндли, Брайан Ф. (2014). «Синхронный круговорот флоры, фауны и климата на рубеже эоцена и олигоцена в Азии» . Научные отчеты . 4 (7463): 7463. Бибкод : 2014NatSR...4E7463S . дои : 10.1038/srep07463 . ПМК 4264005 . ПМИД 25501388 .
- ^ Jump up to: а б с д Хукер, Джерри Дж.; Коллинсон, Маргарет Э.; Силле, Николас П. (2004). «Обмен фауны млекопитающих эоцена-олигоцена в бассейне Хэмпшира, Великобритания: калибровка по глобальному временному масштабу и крупное похолодание» (PDF) . Журнал Геологического общества . 161 (2): 161–172. Бибкод : 2004JGSoc.161..161H . дои : 10.1144/0016-764903-091 . S2CID 140576090 . Архивировано (PDF) из оригинала 8 августа 2023 г. Проверено 31 августа 2023 г.
- ^ Лежандр, Серж; Мурер-Шовире, Сесиль; Хьюгени, Маргарита; Мастер, Элоди; Сиже, Бернар; Эскаргюэль, Жиль (2006). «Динамика разнообразия палеогеновых млекопитающих и птиц Центрального массива (Керси и Лимань, Франция)» . СТРАТА . 1 (на французском языке). 13 : 275–282.
- ^ Эскаргюэль, Жиль; Лежандр, Серж; Сиге, Бернар (2008). «Обнаружение глубоких изменений биоразнообразия: палеогеновое метасообщество млекопитающих в районе Керси и Лимань (Центральный массив, Франция)» . Comptes Rendus Geoscience . 340 (9–10): 602–614. Бибкод : 2008CRGeo.340..602E . дои : 10.1016/j.crte.2007.11.005 . Архивировано из оригинала 13 октября 2023 г. Проверено 19 сентября 2023 г.
- ^ Jump up to: а б Коста, Элисенда; Гарсес, Мигель; Саес, Альберто; Кабрера, Луис; Лопес-Бланко, Мигель (2011). «Возраст оборота млекопитающих «Grande Coupure»: новые ограничения из эоцен-олигоценовых записей Восточного бассейна Эбро (северо-восток Испании)». Палеогеография, Палеоклиматология, Палеоэкология . 301 (1–4): 97–107. Бибкод : 2011PPP...301...97C . дои : 10.1016/j.palaeo.2011.01.005 . hdl : 2445/34510 .
- ^ Хатчинсон, Дэвид К.; Коксалл, Хелен К.; Лант, Дэниел Дж.; Стейнторсдоттир, Маргрет; Де Бур, Агата М.; Баатсен, Мишель Л.Дж.; Фон дер Хейдт, Анна С.; Хубер, Мэтью; Кеннеди-Ассер, Алан Т.; Кунцманн, Лутц; Ладан, Жан-Батист; Лир, Кэролайн; Моравек, Каролин; Пирсон, Пол; Пига, Эмануэла; Паунд, Мэтью Дж.; Зальцманн, Ульрих; Шер, Хоуи Д.; Сейп, Виллем П.; Сливиньска, Касия К; Уилсон, Пол А.; Чжан, Чжунши (2021). «Переход от эоцена к олигоцену: обзор морских и наземных прокси-данных, моделей и сравнения модельных данных» . Климат прошлого . 17 (1): 269–315. Бибкод : 2021CliPa..17..269H . дои : 10.5194/cp-17-269-2021 . S2CID 234099337 .
- ^ Тумулен, Агата; Тардиф, Дельфина; Доннадье, Янник; Лихт, Алексис; Ладан, Жан-Батист; Кунцманн, Лутц; Дюпон-Ниве, Гийом (2022). «Эволюция сезонности континентальной температуры от эоценовой теплицы до олигоценового ледника – сравнение модели и данных» . Климат прошлого . 18 (2): 341–362. Бибкод : 2022CliPa..18..341T . дои : 10.5194/cp-18-341-2022 .
- ^ Булила, Слах; Дюпон-Ниве, Гийом; Гальбрун, Бруно; Бауэр, Хьюз; Шатонеф, Жан-Жак (2021). «Возраст и движущие механизмы перехода эоцена-олигоцена на основе астрономической настройки озерных летописей (бассейн Ренна, Франция)» . Климат прошлого . 17 (6): 2343–2360. Бибкод : 2021CliPa..17.2343B . дои : 10.5194/cp-17-2343-2021 . S2CID 244097729 .
- ^ Соперники, Флоран; Беляев Руслан И.; Басова Вера Борисовна; Прилепская, Наталья Евгеньевна (2023). «Свиньи, бегемоты или медведи? Палеодиета европейских олигоценовых антракотериев и энтелодонтов» . Палеогеография, Палеоклиматология, Палеоэкология . 611 : 111363. Бибкод : 2023PPP...61111363R . дои : 10.1016/j.palaeo.2022.111363 . S2CID 254801829 .
- ^ Беккер, Дэмиен (2009). «Самые ранние находки носорогов (Mammalia: Perissodactyla) из Швейцарии: систематика и биостратиграфия» . Швейцарский журнал геонаук . 102 (3): 489–504. дои : 10.1007/s00015-009-1330-4 . S2CID 67817430 .
- ^ Соле, Флореаль; Фишер, Фишер; Денайер, Жюльен; Спейер, Роберт П.; Фурнье, Морган; Ле Верже, Кевин; Ладевез, Сандрин; Безумие, Аннелиза; Смит, Тьерри (2020). «Хитоядные млекопитающие верхнего эоцена-олигоцена из Керси Фосфоритов (Франция), находящиеся в бельгийских коллекциях» . Геологика Бельгика . 24 (1–2): 1–16. дои : 10.20341/gb.2020.006 . S2CID 224860287 .
- ^ Кочиш, Ласло; Ожварт, Питер; Беккер, Дэмиен; Зиглер, Рейнхард; Шерлер, Лорелин; Влад, Кодря А. (2014). «Орогения вызвала изменение земного климата в период позднего эоцена – начала олигоцена в Европе» (PDF) . Геология . 42 (8): 727–730. Бибкод : 2014Geo....42..727K . дои : 10.1130/G35673.1 . Архивировано (PDF) из оригинала 21 сентября 2017 г. Проверено 19 сентября 2023 г.
- ^ Хукер, Джерри Дж. (2010). «Grande Coupure» в бассейне Хэмпшира, Великобритания: таксономия и стратиграфия млекопитающих по обе стороны от этого крупного палеогенового круговорота фауны». В Уиттакере, Джон Э.; Харт, Малкольм Б. (ред.). Микропалеонтология, осадочная среда и стратиграфия: дань уважения Деннису Карри (1912–2001) . Том. 4. Лондонское геологическое общество. стр. 147–215. дои : 10.1144/TMS004.8 . ISBN 9781862396227 .
- ^ Джумун, Сара С.; Хукер, Джерри Дж.; Коллинсон, Маргарет Э. (2010). Изменение климата и конкуренция как причина вымирания копытных в Гранд-Купюре (ранний олигоцен, Европа) (PDF) . Международный палеонтологический конгресс: Лондон, 2010. с. 221. Архивировано (PDF) из оригинала 11 июня 2020 г. Проверено 31 августа 2023 г.
Внешние ссылки
[ редактировать ]![]() СМИ, связанные с аноплотерием, на Викискладе?
СМИ, связанные с аноплотерием, на Викискладе?