Leconontosaurus
| Leconontosaurus Временный диапазон: поздний меховой ( сеноманский ),
| |
|---|---|

| |
| Реконструированный Череп С. Сахарикуса , Музей науки Миннесоты | |
| Научная классификация | |
| Домен: | Эукариота |
| Королевство: | Животное |
| Филум: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Carcharodontosauridae |
| Subfamily: | †Carcharodontosaurinae |
| Genus: | †Carcharodontosaurus Stromer, 1931 |
| Type species | |
| †Carcharodontosaurus saharicus | |
| Other species | |
| Synonyms | |
|
List | |
Carcharodontosaurus ( / ˌ K ːr k ər oʊ ˌ d ɒ t n oʊ ˈ s ə s / ; ) из «Карачародон ( lit. ' jagged зубчатая ящерица ' или акул-зубца) Кархародон, «зубчатый зубчик») Карчарадон, великая белая акула. Названный немецким палеонтологом Эрнстом Стромером фон Рейхенбахом в 1931 году. Это род хищного динозавра , который жил в Северной Африке примерно с 100 до 94 миллионов лет назад в сеноманский век позднего мелового . Два зуба рода, теперь потерянные, были впервые описаны из Алжира французскими палеонтологами Чарльзом Депером и Джастином Саворнином в роли Megalosaurus Saharicus . Частичный скелет был собран экипажами немецкого палеонтолога Эрнста Стромера во время экспедиции 1914 года в Египет . Стромер не сообщал о египетской находке до 1931 года, в которой он назвал новый род Carcharodontosaurus , делая типовые виды C. saharicus . К сожалению, этот скелет был разрушен во время Второй мировой войны . В 1995 году был обнаружен почти полный череп C. saharicus, первого хорошо сохранившегося образца, который был обнаружен за столетие Кем Кем Марокко кровати ; Он был обозначен неотипом в 1996 году. Окаменелости, обнаруженные из формации Эхкара Северного Нигера, были описаны и названы в качестве другого вида, C. iguidensis , в 2007 году.
Carcharodontosaurus is one of the largest theropod dinosaurs known, reaching 10–12.5 m (33–41 ft) in length and approximately 4–7 metric tons (4.4–7.7 short tons) in body mass. It had a large, lightly built skull with a triangular rostrum. Its jaws were lined with sharp, recurved, serrated teeth that bear striking resemblances to those of the great white shark (genus Carcharodon), the inspiration for the name. Though giant, its cranium was made lighter by greatly expanded fossae and fenestra, but also making it more fragile than tyrannosaurids'. The forelimbs were tiny whereas the hindlimbs were robust and muscular. Like most other theropods, it had an elongated tail for balance.
Many gigantic theropods are known from North Africa during this period, including both species of Carcharodontosaurus as well as the spinosaurid Spinosaurus, the possible ceratosaur Deltadromeus, the large, dubious theropod Bahariasaurus, and an unnamed large abelisaurid. Studies of the bite force and tooth anatomy of carcharodontosaurids have found them to possess strong, but relatively weak bites compared to other theropod families. North Africa at the time was blanketed in mangrove forests and wetlands, creating a hotspot of fish, crocodyliforms, and pterosaur diversity.
Discovery and species
[edit]Initial finds
[edit]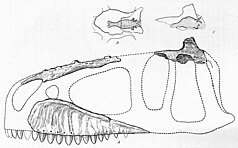
In 1924, two teeth of Carcharodontosaurus were unearthed from wall cuts in different foggaras near Timimoun, French Algeria. These sedimens came from the Cretaceous-aged[1] Continental intercalaire Formation.[2] The fossils were taken to the governor of Timimoun, Captain Burté, who gave them to French geologist Charles Depéret later that year. In 1925, Depéret and his colleague Justin Savornin described the teeth as syntypes (name-bearing specimens) of a new species of theropod dinosaur, Megalosaurus saharicus. These were the first fossils of theropods to be described from the region.[3] The name saharicus refers to the Sahara Desert where the teeth had been found.[4] The genus Megalosaurus was a wastebasket taxon, with many new species referred to it without justification, including M. saharicus.[5] It was later considered to be a species of Dryptosaurus in 1927,[2] though this is unjustified.[6][7] By accident, another species of Megalosaurus, M. africanus, was named by German paleontologist Friedrich von Huene based on the teeth.[6] It is therefore considered a junior synonym of M. saharicus.[8] Both syntypic teeth of M. saharicus have since been lost, possibly being kept in a collection in Algeria, Paris, or Lyon, and lack distinguishing characteristics from other carcharodontosaurids.[9] In 1960, French paleontologist Albert-Félix de Lapparent reported the discovery of more teeth and several caudal vertebrae from sites in Algeria belonging to Carcharodontosaurus,[10] though some of these fossils might belong to other genera.[8] Later authors have mentioned finds of teeth and isolated fossils from other provinces of Algeria.[11][1]
However, fossils of C. saharicus were first found in marls near Ain Gedid, Egypt in early April 1914 by Austro-Hungarian paleontologist Richard Markgraf. Marls from this region derive from the Cenomanian-aged Bahariya Formation, one of many Cretaceous-aged sites of North Africa.[7][12][13] In Bahariya, Markgraf did extensive collecting of dinosaur skeletons for his employer, German paleontologist Ernst Stromer of the Paläontologisches Museum München (Bavarian State Collection of Paleontology). The skeleton of Carcharodontosaurus (IPHG 1922 X46) consisted of: a partial skull, including much of the braincase, teeth, three cervical and a caudal vertebra, incomplete pelvis, a manual ungual, femora, and the left fibula. An isolated ilium was also considered to be from C. saharicus,[14] but it is likely from a ceratosaur.[8]
Due to political tensions between the German Empire and then British-owned Egypt, the Carcharodontosaurus skeleton took years to get to Germany. It was not until 1922 that they were transported overseas to Munich where they were described by Stromer in 1931.[15] Stromer recognized that IPHG 1922 X46’s teeth matched the characteristic dentition of those described by Depéret and Savornin, which led to Stromer conserving the species name saharicus. However, he found it necessary to erect a new genus for this species, Carcharodontosaurus, for their similarities, in sharpness and serrations, to the teeth of the great white shark (Carcharodon carcharias).[13] World War II would break out in 1939, leading IPHG 1922 X46 and other material from Bahariya to be destroyed during a British bombing raid on Munich during the night of April 24/25, 1944.[16][17] An endocast was made and survived the war, being the only remaining relic of the specimen.[18]
Resurgent discoveries, C. iguidensis, and confusion with spinosaurids
[edit]
Few discoveries of Carcharodontosaurus material were made until 1995, when American paleontologist Paul Sereno found an incomplete skull during an expedition embarked on by the University of Chicago. This skull (UCRC PV12) was found in the Cenomanian-aged rocks of the Lower Douira Formation, Kem Kem Beds in Errachidia, southeastern Morocco. The specimen was taken to the University of Chicago and described in 1996 by Sereno and colleagues. In a later paper, UCRC PV12 was designated as the neotype of C. saharicus due to the loss of other specimens and the similar age and geographic location to previously noted material.[7] The taxonomy of Carcharodontosaurus was discussed by Chiarenza and Cau (2016),[19] who suggested that the neotype of C. saharicus was similar but distinct from the holotype in the morphology of the maxillary interdental plates. However, paleontologist Mickey Mortimer put forward that the suggested difference between the C. saharicus neotype and holotype was actually due to damage to the neotype.[8]
Several other fossils of C. saharicus have been unearthed from the Kem Kem Beds, such as dentary fragments, a cervical vertebra, and many teeth.[20][21][22] Sereno et al. also referred a multitude of cervical vertebrae described as the spinosaurids Sigilmassasaurus and "Spinosaurus B" to C. saharicus reasoning that stout cervicals would be needed to carry the skulls of carcharodontosaurids.[9][7] Later research proved otherwise, with the vertebrae being placed in Spinosaurus aegyptiacus by Ibrahim et al. (2020).[23] French paleontologist René Lavocat was the first to note the possible presence of Carcharodontosaurus in Morocco as early as 1954.[24]
В 2007 году новый вид Carcharodontosaurus, C. iguidensis, был назван палеонтологами Стивом Брусатте и Полом Серено. Окаменелости C. iguidensis были обнаружены во время экспедиции в формирование Echkar Iguidi, Niger , частичную верхнюю версию (MNN Igu2), обозначаемую голотипом. Название вида iguidensis после Iguidi, где ископаемые были обнаружены. Несколько останков, таких как мозговой, слезное, зубьев, шейный позвонок и набор зубов, были переданы C. iguidensis в зависимости от размера и предполагаемого сходства с другими костями Carcharodontosaurus . [9] Chiarenza and Cau (2016) identified the referred material of C. iguidensis as belonging to Sigilmassasaurus (later referred to Spinosaurus sp.)[25] and a non-carcharodontosaurine, and therefore chose to limit C. iguidensis to the holotype pending future research.[19] Another carcharodontosaurid from the Kem Kem Beds, Sauroniops pachytholus, was dubbed in 2012 based on a single frontal.[26] This species has been proposed to be synonymous with C. saharicus,[ 27 ] Хотя это наблюдало сопротивление и достоверность, поддерживаемую в большинстве литературы. [ 26 ] [ 28 ] [ 19 ] [ 29 ] Южноамериканский род Giganotosaurus был синонимизирован Carcharodontosaurus Figueiredo (1998) и Пол (2010), [ 30 ] [ 31 ] Однако с тех пор ни один авторы не следовали этой оценке. [ 19 ]
Другие упомянутые образцы
[ редактировать ]
- Lapparent (1951, 1960) описал несколько зубов Carcharodontosaurus из континентального интеркалера Гермерса, Тунис . [ 10 ] [ 3 ]
- Посторбитальная кость и несколько посткраниальных останков, назначенных Carcharodontosaurus, были обнаружены в формировании Элрхаза Северного Нигера. Таке (1976) отметил, что посторбитал был похож на то, что у Acrocanthosaurus, родственника Carcharodontosaurus , [ 32 ] в то время как посткраниальные окаменелости могут принадлежать к другим тероподам. [ 8 ]
- Два фрагмента мозга, 137 зубов, два каудальных позвонка и ручная фаланга из формации Эшкар были названы Carcharodontosaurus Lapparent (1960). [ 10 ] Фаланса педали также была описана как Carcharodontosaurus, но вместо этого она, вероятно, происходит от спинозаурида. [ 33 ]
- Многие позвонки, в том числе два связанных дорсала, были обнаружены в ранних меловых слоях ирхазерской , Нигер группы Агадеса . Лаппарент упомянул эти окаменелости как C. saharicus в 1960 году, [ 10 ] хотя они могут принадлежать к другим родам теропода. [ 8 ]
- Каудальные позвонки из Tefidet и зубов из сайтов Akarazeras континентального интеркалера Агадеса, Нигер, были зарегистрированы Lapparent (1960) [ 10 ] и Таке (1976) соответственно. [ 32 ] Позвонки могут быть из других теропод. [ 8 ]
- Из неизвестного местного интеркалера пустыни Сахара Лапарент (1960) задокументировал избранные позвонки, плечевую кость и ручную фалангу как из С. Сахарируса. [ 10 ] Эти элементы могут быть от других теропод. [ 8 ]
- Две документы, первые в 1978 году, описали зубы и каудальный позвонок Carcharodontosaurus из формирования Ченини Южного Туниса . [ 34 ] Тем не менее, каудальный позвонок теперь помечен Carcharodontosauridae Indet. [ 35 ]
- Окаменелости из формирования QUSEIR западного Египта были назначены Carcharodontosaurus в 1999 году. [ 36 ]
- В статье 1999 года описано несколько позвонков и элементов конечностей карчамодонтозаврида, которые были обнаружены из образования молока Вади в Судане . Они были размещены только в качестве Carcharodontosaurid, но исследователь Микки Мортимер предположил, что они принадлежат к самому Carcharodontosaurus на основе присутствия плеврокоэлей (мелкие отверстия) в хвостовых позвонках. [ 8 ]
- В 2015 году большая нейронная арка из спинного позвонка русло Kem Kem была неофициально описана как принадлежащая к новому роду и видам Мегараптора, дублированного «остеопорозия Gigantea». Этот образец принадлежит главе польской цепочки тематических парков, которая описала его как принадлежащую к 15 -метровому (49 футам) карнозавру, аналогичному Mapusaurus и Carcharodontosaurus. [ 37 ] Тем не менее, он был намного меньше, чем предлагается, и может принадлежать C. saharicus или Saurooniops на основе его качеств и происхождения Carcharodontosaurid. [ 38 ]
Ошибочно назначенные образцы
[ редактировать ]- Верхнечелюстный зуб, извлеченный из формации Вильяр -дель -Арзобиспо в Испании, был передан Carcharodontosaurus в 1966 году. [ 39 ] Тем не менее, в нем отсутствуют черты зубов карчадонтозавридов и вместо этого больше похожи на другие аллозавра . [ 40 ] [ 41 ]
- Бонд и Бромли (1970) описали зубы, вытекающие из формирования Gokwe Zimbabwe , как аналогичные Carcharodontosaurus , причем Микки Мортимер назначал их роду, предварительно предназначенным. [ 42 ] Однако более поздние исследования показали, что они неопределенны. [ 43 ]
- Зубы из формации Алькантары в Бразилии были помещены в Carcharodontosaurus в 2002 году, [ 44 ] Но это было оспорено на основе его географического происхождения. [ 8 ]
Описание
[ редактировать ]

Размер
[ редактировать ]Стромер предположил, что C. saharicus был примерно таким же размером с тиранозаврида Gorgosaurus , который поместил бы его на высоте около 8–9 метров (26–30 футов), на основе его образца IPHG 1922 X46. Этот человек был примерно на 15% меньше, чем неотип, [ 45 ] Последний, по оценкам, составляет 12–12,5 метра (39–41 фута) по длине и приблизительно 5–7 метрических тонн (5,5–7,7 коротких тонн) в массе тела. [ 46 ] [ 31 ] [ 47 ] [ 48 ] Это делает Carcharodontosaurus saharicus одним из крупнейших известных динозавров и наземных плотоядных животных, являющихся 2 -м по величине кархадонтозавридам и 3 -м по величине теропод в целом, согласно большинству оценок. [ 49 ] [ 31 ] C. Iguidensis был намного меньше, достигая всего 10 метров (33 фута) по длине и 4 метрических тонн (4,4 тонны) в массе тела. [ 31 ]
Череп
[ редактировать ]Череп
[ редактировать ]
Самый большой и самый полный череп C. saharicus будет измерять 1,6 метра (5,2 фута) при завершении, примерно тот же размер, что и самые большие черепа тиранозавра . Никаких черепов рода, заповедника , предварительные оксиллы , полные задние области черепа или нижние челюсти . Черепа автомобилей, как правило, являются более стройными и слегка построенными, чем у более поздних тиранозавридов, которые имеют надежные сборки и адаптацию для дробления. Нетипический череп сужается в сторону передней части в боковом виде, создавая треугольный контур. Это похоже на то, что у других кархародонтозавридов, таких как Mapusaurus и Giganotosaurus . Его череп был легче, чем у тиранозавридов, при этом анторбитальный фенестра сочинял более 30% от общей длины черепа, а также окружено ямками в верхних количествах (верхняя челюстная кость), носовые кости), Jugals (скула) и и и и Локрималы (передняя орбита кость). Сродни другим родам, его носа удлиняется и имеет переднюю поверхность, покрытую поверхностью с морщинами. Эти удары, вероятно, были расширены кератиновыми оболочками, создавая рогоподобную структуру, как в Ceratosaurus . Аналогичная побойщика обнаруживается на слезной части, которая также будет удлинена кератином, образуя аналогичный элемент. [ 50 ] [ 7 ] Наиболее характерной чертой Carcharodontosaurus черепа является скульптурная внешность верхних челюстей, которая уникальна для рода. Тем не менее, C. iguidensis имеет анторбитальные ямки, ограниченную близостью анторбитального фенестра, гребень, проходящего вдоль медиальной поверхности верхней челюсти, и процесс вдоль его средней линии. Эти черты отсутствуют в C. saharicus , дифференцируя два вида. [ 9 ]




Верхтка IPHG 1922 X46 составила бы длину 70 сантиметров (28 дюймов), когда завершена, тогда как полная верхняя челюсть неотипа намного больше. 14 зубьев присутствуют в каждой верхней челюсти. Части мозгового мозга известны, хотя большая часть их морфологии такая же, как Гиганотозавра » . Тем не менее, C. saharicus имеет гораздо более заметный Nuchal Crest , который наступает на крыше черепа. Фронтальные кости твердо слиты, характерно, очевидно в большинстве теропод. [ 51 ] Джугалы широкие и в форме треугольника. была Артикуляция нижней челюсти помещена дальше назад за затылочным мыщелком (где шея прикреплена к черепу) по сравнению с другими тероподами. [ 7 ] Два зубных (нижняя челюстная кость), которые были переданы C. saharicus Ibrahim et al. (2020) имеют глубокие и расширенные альвеолы (зубные розетки), признаки, обнаруженные в других крупных тероподах. [ 20 ] [ 22 ] Если вроде тиранотитана и гиганотозавра , у зубца было бы 16 альвеол (зубные розетки). [ 52 ]


Оценки количества зубов Carcharodontosaurus различаются, но недавняя оценка 30 зубных зубных, 8 предвзящих пластин и 24 верхнечелюстных зубов для 62 зубов была сделана. [ 53 ] Зубы Carcharodontosaurid являются одними из крупнейших из всех групп динозавров, а верхнечелюстный зуб от IPHG 1922 X46 - 6,8 сантиметра (2,7 дюйма) высотой и 3,5 сантиметра (1,4 дюйма). [ 13 ] [ 54 ] Тем не менее, они чрезвычайно тонкие, большинство из которых находятся меньше сантиметра. Смыслы многочисленны на передней и задней краях, с более чем 18-20 зубцами на сантиметр края у C. saharicus и до 32 на сантиметр у C. iguidensis . [ 9 ] [ 13 ] Его зубы прямые, сглажены в боковом направлении и веретена в форме веретена в поперечном сечении. Тем не менее, зубные зубные узоры к задней части рта стали более повторяющимися, чем в верхней челюсти. Задний край этих коронок повторяется и выпуклый при его прекращении. поддержанных Свева для эмали присутствует на обеих дорсовентральных сторонах коронок. Эти морщины изгибаются по направлению к маргинальным зубцам, составляя полосу группы вдоль концов. [ 7 ] [ 55 ] [ 52 ]
Мозг и внутреннее ухо
[ редактировать ]В 2001 году Hans CE Larsson опубликовал описание внутреннего уха и эндокрания Carcharodontosaurus Saharicus . Начиная с части мозга, ближайшей к кончику морды животного, - это передний мозг, за которым следует средний мозг. Средний мозг наклонен вниз под углом 45 градусов и к задней части животного. За этим следует задним изделиям , который примерно параллельно передней части и образует примерно 40-градусный угол с средним мозгом. В целом, мозг C. saharicus был бы похож на мозг связанного динозавра, Allosaurus fragilis . Ларссон обнаружил, что отношение головного мозга к объему мозга в целом у Carcharodontosaurus было типичным для неавийской рептилии. Carcharodontosaurus также имел большой зрительный нерв . [ 56 ]
Три полукруглых канала внутреннего уха Carcharodontosaurus Saharicus - когда смотрели со стороны - имели субтраулярный контур. Эта субтрианговая конфигурация внутреннего уха присутствует в аллозавре , ящерицах и черепах , но не у птиц . Сами полу- "циркулярные" каналы были очень линейными, что объясняет заостренный силуэт. В жизни флокковая доля мозга проецировалась бы в область, окруженную полукруглыми каналами, как и в других неавийских тероподах, птицах и птерозаврах. [ 56 ]
Посткрания
[ редактировать ]
Немногие посткраниальные элементы уверенно известны из Carcharodontosaurus, хотя многие изолированные кости из Сахары были направлены на род без подробного исследования. [ 9 ] [ 23 ] [ 32 ] [ 10 ] Как и другие Carcharodontosaurids, он был надежным с небольшими передниками , удлиненным хвостом и короткой шеей. Самым полным образцом был IPHG 1922 X46, но он был уничтожен. Этот образец сохранил 3 шейных позвонков , которые подвергались выветрину. Одна из них - ось , а две другие - это сочленение передних шейных матков, которые длиннее и шире, чем ось. Цервикальные позвонки Carcharodontosaurus - это полные и опишкокулюсы (вогнутые задние концы). [ 57 ] [ 52 ] Шейные позвонки в этом роде, как и в Giganotosaurus , увенчаны низкими нейронными шипами, соединенными с прочными поперечными процессами , которые висели над плеорокоэлем (мелкие депрессии на сторонах центра), которые будут содержать пневматические воздушные мешки, чтобы осветлить позвонки. Центра этих позвонков украшены килями вдоль их вентральных сторон. Был также известен передний каудальный позвонок , который был уплачиваемым (плоские передние и задние концы) и короткие. Этот каудаль был неполным, пропустив большую часть нейронного позвоночника, но имел диапофизы, которые конъюгировали бы с шевронами . Стороны его центра были также плеврокоэлюс. Два лезвия, похожие на лезвия, также были сохранены и у этого человека. [ 13 ]
Таз был неполным, содержащий как лобки , так и левый исий , хотя полные осадки известны в родственных родах. Иший направился назад, тогда как лобки указывали вперед, диагностическую черту сауришцев . Лобки, вероятно, были почти на 1 метр (3,3 фута), когда они были полностью сохранены, с тонкими валами, но были поперечно расширены на передних концах, где они соединялись, создавая V-образную форму в переднем виде. Оба бедра в дополнение к левой малоберцовой кости были извлечены, первый элемент является одним из крупнейших зарегистрированных из теропода на 1,26 метра (4,1 фута) в длину. Его бедренке не хватало сильной кривизны и в основном прямой, за исключением передних и задних концов. Большой вертел невелик, но имеет заметное выступление, которое будет прикрепляться к M. Caudofemoralis Longus мышца хвоста. Его малоберцовая кость составляла всего 88 сантиметров (35 дюймов) длиной, около 1/3 длины бедра. Передний конец был треугольным в боковом виде с выпуклыми мыщелками , тогда как задний конец округлен. [ 13 ]
Классификация
[ редактировать ]Систематика
[ редактировать ]
Carcharodontosaurus является типовым родом семейства Carcharodontosauridae и подсемейства Carcharodontosaurinae . Эта подсемейство содержит сам Carcharodontosaurus , а также другие Carcharodontosaurines Giganotosaurus, Mapusaurus Meraxes и , Tyrannotitan ; Тем не менее, эти роды составляют независимое племя: гиганотозаурини. Carcharodontosauridae была кладкой, созданной Стромером для Carcharodontosaurus и Bahariasaurus , хотя название оставалось неиспользованным до признания других членов группы в конце 20 -го века. Он отметил подобие костей Carcharodontosaurus с американским аллозавром Theropods и Tyrannosaurus , что заставило его рассмотреть семейную часть Theropoda. [ 58 ] [ 13 ]
Описание Пола Серено о окаменелостях Carcharodontosaurus в 1996 году привело к реализации трансконтинентальной клады карчадонтозавридов. По мере того, как было обнаружено больше кархадонтозавридов, их взаимосвязи стали еще более ясными. Группа была определена как все аллозавроиды ближе к Carcharodontosaurus, чем аллозавр или синраптор от палеонтолога Томаса Р. Хольца и коллег в 2004 году. [ 59 ] Carcharodontosaurus более плохо известен, чем большинство других кархародонтозавридов, при этом Meraxes и Giganotosaurus представлены почти полными скелетами. [ 60 ] [ 61 ] Члены семьи были признаны от поздней юры до среднего мела на каждом континенте, кроме Океании и Антарктиды. [ 49 ] [ 62 ] [ 7 ]
Canale et al. (2022) восстановил Carcharodontosaurus как самый ранний отклоняющийся член Carcharodontosaurinae. Результаты кладограммы их филогенетического анализа отображаются в кладограмме ниже: [ 60 ]


В своем обзоре тероподных отношений в 2024 году Кау не восстановил все испытанные образцы Carcharodontosaurus в монофилетической кладе. Результаты его филогенетического анализа показаны в кладограмме ниже: [ 63 ]
| Funceodontosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюция
[ редактировать ]Родольфо Кория и Леонардо Сальгадо предположили, что сходящаяся эволюция гигантизма в тероподах могла быть связана с общими условиями в их среде или экосистемах . [ 61 ] Серено и его коллеги обнаружили, что присутствие карчадонтозавридов в Африке ( Carcharodontosaurus ), Северной Америки ( Acrocanthosaurus ) и Южной Америки ( Giganotosaurus ), показали, что у группы была трансконтинентальная дистрибуция в раннем меловом периоде . Маршруты рассеивания между северным и южным континентами, по -видимому, были разорваны океаническими барьерами в позднем меловом периоде, что привело к более четким, провинциальным фаунам, предотвращая обмен. [ 64 ] [ 7 ] Раньше считалось, что в мелах -чуде был биогеографически отделен, а на северных континентах преобладают тиранозавриды, Южная Америка от абелизавридов и Африки со стороны карчадонтозавридов. [ 65 ] Подсемейство Carcharodontosaurinae, в которой принадлежит Carcharodontosaurus , по -видимому, был ограничен южным континентом Гондваны (сформированной Южной Америкой и Африкой), где они, вероятно, были хищниками Apex . [ 59 ] Южноамериканское племя Гиганотозаурини, возможно, были отделены от их африканских родственников через , когда Гондвана рассталась во время аптиан -альбианских викарианс веков раннего мела. [ 66 ]
Палеобиология
[ редактировать ]Подъемные возможности
[ редактировать ]Биомеханический анализ возможностей для подъема Carcharodontosaurus был проведен палеонтологом Дональдом Хендерсоном и палеоартистом Робертом Николсом в 2015 году. Авторы использовали 3D -модели животного, а также субадульт -сауропод , которые, хотя и не были найдены вместе с Carcharodontosaurus , сходны с рефбахизаврами , которые не были найдены вместе с Carcharodontosaur Кровати Кем Кем. Модели включали размер легких и других пневматических структур двух, способствующих точным моделированию веса сценария. Исследование Henderson & Nicholls показало, что взрослый C. Saharicus может иметь максимум 424 кг (935 фунтов), половину веса взрослого Лимайсаура . Однако два взрослых C. saharicus могли бы вместе поднять до 850 килограммов (1870 фунтов). [ 46 ]
Кормление и диета
[ редактировать ]
Расточие зубные зубы аллозавроидов отличаются, с зубами кархадонтозавридов явно тонкие и лезвия зубы. Тем не менее, эти зубы тонкие и, вероятно, не могут поддерживать воздействие на твердые поверхности, такие как кость, без потенциального изгиба и схватки. Эта опасность усугубляется прямыми краями, слегка повторяющимися кончиками и синусоидальными формами, наблюдаемыми в их зубном зубе. Несмотря на эти черты, зубы все еще гораздо более надежны, чем у меньших теропод, и из -за их общего размера может потребоваться большее давление. Carcharodontosaurus также имел высокую скорость замены зубов, что означает, что поврежденные зубы можно было легко заменить в отличие от существующих костей млекопитающих, которые тратят большую часть своей энергии, поддерживая свои зубы. [ 67 ] [ 68 ] Свидетельство о укусах кости наблюдается в аллозавре , который будет участвовать в ритуальном обращении с другими людьми и кусается в упор стегозавр , как показано следованиями укусов. [ 69 ] [ 70 ] [ 71 ]
силы укуса Carcharodontosaurus, а также другие гигантские тероподы, включая AcroCanthosaurus и Tyrannosaurus Были проанализированы . Исследования сообщают, что у карчадонтозавридов были гораздо более низкие силы укуса, чем у тиранозавра, несмотря на то, что они находятся в классе одного размера. Передняя сила укуса C. saharicus была оценена в статье 2022 года, которая составляет 11 312 ньютонов, в то время как задняя сила укуса составляла 25 449 ньютонов. Это намного ниже, чем у тиранозавра, подразумевая, что он не ел кости. [ 72 ] [ 73 ] Также были взяты конечные элементы, которые также были взяты, что также подтвердило идею, что Carcharodontosaurus ел более мягкую пищу, чем тираннозавриды. Большое количество стресса было извлечено в задней части черепа возле квадрата в Carcharodontosaurus, спинозавра и акрокантозавра . Черепа этих теропод имели более высокие относительные количества стресса в противодействии меньшим родам. Это указывает на то, что череп гигантских таксонов (например, Carcharodontosaurus ) была нестабильна из -за наличия больших пневматических структур для экономии веса вместо создания твердой сборки. Тем не менее, спинозавр и сухомус испытали еще большие ценности стресса, что означает, что они могли потреблять только светлую, небольшую добычу, а не более крупные предметы, которые более сильный череп Carcharodontosaurus мог кусать при поддержании стресса. [ 74 ]
Изотопные анализы зубов C. saharicus обнаружили значения Δ18O, которые выше, чем у современного спинозавра , что предполагает, что последний преследул полуаватные привычки, тогда как Carcharodontosaurus был более земным. [ 75 ] Это дополнительно подтверждается тафономией зубов C. saharicus , которые чаще встречаются на территории суши, чем водные. [ 53 ] Carcharodontosaurus также была гомеотермой с эндотермической , термофизиологией которая была предполагалась этими изотопами, что означает, что большая часть его кислорода была накоплена питьевой водой, а не в ней. [ 76 ] [ 77 ]
Функция гребня
[ редактировать ]Тероподы, такие как Carcharodontosaurus, Allosaurus и Acrocanthosaurus , увеличили ласковые гребни, цель которых неизвестна. Палеонтолог Даниэль Чуре предположил, что эти гребни использовались для «разбивания головы» между людьми, но насколько они долговечны, не изучались. [ 78 ]
Зрение
[ редактировать ]
Исследование, проведенное в 2006 году биолога Кента Стивенса, проанализировало возможности бинокулярного зрения аллозавра Carcharodontosaurus и Allosaurus, а также несколько колурозавров, включая Tyrannosaurus и Stenonychosaurus . Применяя модифицированную периметрию к моделям головок этих динозавров, Стивенс пришел к выводу, что бинокль Carcharodontosaurus было ограничено, побочным эффектом его большой удлиненной рострум. Его наибольшая степень бинокулярного зрения была на более высоких высотах, что позволяет предположить, что Carcharodontosaurus , возможно, обычно держал голову под углом вниз на 40 °, когда его глаза соответственно обращались к максимальному бинокулярному зрению. Диапазон видения, наблюдаемый в этих аллозавроидах, сопоставим с видами крокодилов, что позволяет предположить, что они были засадными хищниками . Скорее всего, они почувствовали добычу через параллакс движения между добычей и фоном, с узким бинокулярным полем зрения, помогая хищникам судить о добыче расстояния и временные атаки. [ 79 ]
Патология
[ редактировать ]Неотипный череп C. saharicus является одним из многих аллозавроидных лиц, которые сохраняют патологии , с признаками укуса, инфекции и разрывов, наблюдаемых в аллозавре и акрокантозавре среди других. [ 80 ] [ 81 ] Этот череп несет круглую пункцию, растущую в носе, и «аномальная проекция кости на анторбитальном ободе». [ 81 ] Позднее исследование предположило, что это было результатом черепно -лицевых укусов. [ 82 ]
Палеоэкола
[ редактировать ]
Окаменелости Carcharodontosaurus известны из нескольких мела-эпох по всей Северной Африке, аналогично диапазонам спинозавра и deltadromeus . [ 20 ] [ 7 ] Северная Африка в течение этого периода граничал с морем Тетис , которое превратило регион в прибрежную среду, в которой домят, наполненные мангро , заполненную обширными приливными квартирами и водными путями . [ 83 ] [ 84 ] [ 85 ] Изотопы из окаменелостей Carcharodontosaurus и Spinosaurus предполагают, что кемки Kem Kem были свидетелями временного сезона муссонов , а не постоянного количества осадков, аналогичных современным условиям, присутствующим в субтропических и тропических средах в Юго-Восточной Азии и Африке к юго-кара . [ 75 ] [ 86 ] Эти речные месторождения носили больших рыб, в том числе « Sawskate Onchopristis» , Coelacanth Mawsonia и Bichir Bawitius . [ 87 ] Это привело к обилию писци иродных крокодилиформных, в ответ, таких как гигантский стоматосухид -стоматосухус в Египте и роды Elosuchus , Laganosuchus и развивающихся Aegisuchus из Марокко. [ 88 ] [ 89 ] Марокко также носила изобилие птерозавров, таких как Siroccopteryx и Nicorhynchus . [ 90 ] [ 91 ]
Композиция фауны динозавров этих участков является аномалией, поскольку меньше видов динозавров меньше по сравнению с плотоядными динозаврами, чем обычно. Это указывает на то, что между различными тероподскими кладами было нишевое распределение, а спинозавриды потребляли рыбу, в то время как другие группы охотились на травоядных динозавров. [ 92 ] Изотопные данные подтверждают это, которые обнаружили большее количество значительных наземных животных в рационах карчадонтозавридов и цератозавров как из пластов Kem Kem, так и в формации Elrhaz. [ 93 ] [ 10 ] Некоторые сауроподы известны из бахарии, такой как паралититан и айгиптозавр , [ 94 ] в то время как Ребебахисавр находится в кроватях Kem Kem. [ 95 ] Карчадонтозавриды представлены C. saharicus и Saurooniops в слоях Kem Kem, Eocarchia и потенциально Carcharodontosaurus в формации Elrhaz и C. iguidensis в формации Echkar. [ 9 ]
Ссылки
[ редактировать ]- ^ Jump up to: а беременный Beyyoucef, предательство; Pérez-García, Adn; Бендель, Мохамед; Ортега, Франциско; Вулло, римляне; Bouchemla, Imad; Ферре, Бруно (2022). Криторус . Границы в октябре науки 10 : 927059. Bibcode : 2022freeaas . doi : 10.3389/Feart . ISSN 2296-6463 .
- ^ Jump up to: а беременный Депер, Чарльз; Саворнин, Джастин (1927). «Фауна рептилий и ядов Альбиана из Тмимуна (Алжир Сахара)». Бюллетень Геологического общества Франции . 27 : 257–265.
- ^ Jump up to: а беременный Buffetaut, Eric; Ouaja, Mohamed (2002). «Новый образец спинозавра (Dinosauria, Theropoda) из нижнего мела Туниса, с замечаниями об эволюционной истории Spinosauridae» (PDF) . Бюллетень de la Société Géologique de France . 173 (5): 415–421. doi : 10.2113/173.5.415 . S2CID 53519187 .
- ^ Депер, Чарльз; Саворнин, Джастин (1925). «Об обнаружении фауны позвоночных Альбиана в Тмимуне (Западный Сахара)» . Отчеты Академии наук . 181 : 1108–1111.
- ^ Бенсон, Роджер Б.Дж.; Барретт, Пол М.; Пауэлл, Х. Филип; Норман, Дэвид Б. (2008). «Таксономический статус Megalosaurus Bucklandii (Dinosauria, Theropoda) из среднего юрского периода в Оксфордшире, Великобритания» . Палеонтология . 51 (2): 419–424. Bibcode : 2008Palgy..51..419b . doi : 10.1111/j.1475-4983.2008.00751.x . S2CID 83324840 .
- ^ Jump up to: а беременный Хуин, Фридрих (1956). Палеонтология и филогения нижних тетрапод . Том. 1. Джена : просьба Veb Gustav Fischer. п. 716. OCLC 489883421 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Серено, Пол С.; Dutheil, Didier B.; Iarochene, M.; Ларссон, Ганс Се; Лион, Габриель Х.; Magwene, Paul M.; Сидор, Кристиан А.; Варриккио, Дэвид Дж.; Уилсон, Джеффри А. (1996). «Хищные динозавры из Сахары и поздней меловой дифференцировки фауны» (PDF) . Наука . 272 (5264): 986–991. Bibcode : 1996sci ... 272..986s . doi : 10.1126/science.272.5264.986 . ISSN 0036-8075 . PMID 8662584 . S2CID 39658297 .
- ^ Jump up to: а беременный в дюймовый и фон глин час я Дж Мортимер, Микки (2023). "Carnosauria" . База данных Theropod . Архивировано из оригинала 28 мая 2023 года . Получено 6 июня 2023 года .
- ^ Jump up to: а беременный в дюймовый и фон глин Брусатт, Стивен Л.; Серено, Пол С. (12 декабря 2007 г.). «Новый вид Carcharodontosaurus (Dinosauria: Theropoda) из сеномана Нигера и пересмотр рода» . Журнал палеонтологии позвоночных . 27 (4): 902–916. doi : 10.1671/0272-4634 (2007) 27 [902: ansocd] 2.0.co; 2 . ISSN 0272-4634 . S2CID 86202969 .
- ^ Jump up to: а беременный в дюймовый и фон глин час De Lapparent, Albert (1960). "Les Dinosauriens du" Continental Intercalaire "Du Sahara Central" (PDF) . Мемуары Геологического общества Франции . 88a : 1–57.
- ^ Габани, А.; Маммери, С.; Adaci, M.; Бенсала, М.; Mahboubi, M. (2016). «Континентальный меловой меловой фоне на южной границе плато Тинхерта: стратиграфические соображения и палеонтологическая оценка». Память о геологической службе Алжира . 19 : 39–61.
- ^ Ibrahim et al. 2020 , с. 162.
- ^ Jump up to: а беременный в дюймовый и фон глин Стромер, Эрнст (1931). «Результаты исследовательских поездок Профессор Э. Стромерс в пустынях Египта II. Остаток позвоночных Бахари -джестуфов (Нижний Сеноман). 10. Остаток скелета остаточным остатками Carcharodontosaurus Nov.» (PDF) . Трактаты Баварской академии наук математического и научного факультета . Новый эпизод (на немецком языке). 9 : 1–23.
- ^ Стромер 1934 , с. 46
- ^ «Скалистая дорога: Эрнст Стромер» . www.strangescience.net . Получено 8 июля 2023 года .
- ^ Смит, Джошуа Б.; Ламанна, Мэтью С.; Майр, Гельмут; Лаковара, Кеннет Дж. (2006). «Новая информация о голотипе спинозавра Aegyptiacus Stromer, 1915» . Журнал палеонтологии . 80 (2): 400–406. doi : 10.1666/0022-3360 (2006) 080 [0400: nirtho] 2.0.co; 2 . ISSN 0022-3360 . S2CID 130989487 .
- ^ Nothdurft & Smith 2002 .
- ^ Ibrahim et al. 2020 , с. 162, 164.
- ^ Jump up to: а беременный в дюймовый Чиаренца, Альфио Алессандро; Кау, Андреа (29 февраля 2016 г.). «Большой абелисаврид (Dinosauria, Theropoda) из Марокко и комментирует тероподы сеномана из Северной Африки» . ПЕРЕЙ . 4 : E1754. doi : 10.7717/peerj.1754 . ISSN 2167-8359 . PMC 4782726 . PMID 26966675 .
- ^ Jump up to: а беременный в Рассел, Дейл (1996). «Изолированные кости динозавров от среднего мела тафилальта, Марокко» . Бюллетень Национального музея естественной истории, 4 -я серия - Раздел C - Земные науки, палеонтология, геология, минералогия (на французском языке). 18 (2–3).
- ^ Ibrahim, N., Sereno, PC, Zouhri, S. & Zouhri, S. (2017). Марокко динозавры - исторический аукцион и недавние работы. Мемуары Геологического общества Франции , 180 , 249-284.
- ^ Jump up to: а беременный Ibrahim et al. 2020 , с. 167
- ^ Jump up to: а беременный Ibrahim et al. 2020 , с. 169
- ^ Лавокат, Рене (1954). «На динозаврах межполосного континентального континентального кэма дауры». Отчеты 19 -й Международный геологический конгресс . 1952 : 65–68.
- ^ Серено, Пол С.; Myhrvold, Nathan; Хендерсон, Дональд М.; Рыба, Фрэнк Э.; Видал, Даниэль; Baumgart, Stephanie L.; Кейлор, Тайлер М.; Formoso, Kiersten K.; Конрой, Лорен Л. (30 ноября 2022 г.). Чжу, мин; Руц, Кристиан; Чжу, мин; Хольц, Томас Р.; Хон, Дэвид (ред.). « Спинозавр - это не водный динозавр» . элиф . 11 : E80092. doi : 10.7554/elife.80092 . ISSN 2050-084X . PMC 9711522 . PMID 36448670 .
- ^ Jump up to: а беременный Кау, Андреа; Далла Веккия, Фабио М.; Фаббри, Маттео (1 марта 2013 г.). «Толсто-кокулярный теропод (Dinosauria, Saurischia) из верхнего мела Марокко с последствиями для эволюции черепа Carcharodontosaurid» . Мерашные исследования . 40 : 251–260. Bibcode : 2013crres..40..251c . doi : 10.1016/j.cretres.2012.09.002 . ISSN 0195-6671 .
- ^ Ibrahim et al. 2020 , с. 171.
- ^ Кандейро, Карлос Роберто Дос Анжос; Брусатт, Стивен Луи; Видал, Лучано; Перейра, Пауло Виктор Луис Гомес да Коста (26 июля 2018 г.). «Палеобиогиогеография эволюция и распределение Carcharodontosauridae (Dinosauria, Theropoda) во время среднего мела Северной Африки » . Одиночные роли зоологии . 58 : 20185829. Doi : 10.11606/1807-0205/2018.58.29 . HDL : 20.500.11820/C4CA0A5C-4F8E-4136-8355-8BD32D6EA544 . ISSN 0031-1049 . S2CID 53353652 .
- ^ Патерна, Алессандро; Кау, Андреа (11 октября 2022 г.). «Новый гигантский материал Theropod из комплексной комплексной компоненты Kem Kem (Марокко) с последствиями на разнообразие среднеклезных кархародонтозавридов из Северной Африки» . Историческая биология . 35 (11): 2036–2044. doi : 10.1080/08912963.2022.2131406 . ISSN 0891-2963 . S2CID 252856791 .
- ^ Figueiredo (1998). «Плотноядные динородологии: их описание и образ жизни». Португальский центр геоистории и доистории . стр. 1–4.
- ^ Jump up to: а беременный в дюймовый Пол, Грегори С. (2016). Полевой гид Принстона по динозаврам . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. С. 103–104. ISBN 978-1-78684-190-2 Полем OCLC 985402380 .
- ^ Jump up to: а беременный в Таке, Филипп (1976). Геология и палеонтология депозита Gadoufaoua (Niger Aptian) . Париж: палеонтологические записные книжки, Национальный центр научных исследований. п. 53. ISBN 978-2-222-02018-9 .
- ^ Ибрагим, Низар; Серено, Пол С.; Дал Сассо, Кристиан; мусор, Симона; Фаббри, Маттео; Martill, David M.; Зухри, Самир; Myhrvod, Nathan; Iurino, Dgawid A. (26 сентября 2014 г.). «Полуадация в гигантском хищническом динозавре » Наука 345 (6204): 1613–1 Bibcode : 2014sci ... 345.1613i Doi : 10.1126/science.1258750 . ISSN 0036-8 PMID 25213375 S2CID 34421257
- ^ Ошибка; Schwarzhans (1978). «Места для костей от Wealden Sud-Tunesia (окружение KSAR CRERACHFA)». Берлинская Геоссаука трактивы а . 8 : 53–65.
- ^ Фанти, Федерико; Кау, Андреа; Мартинелли, Агнезе; Contressi, Michela (15 сентября 2014 г.). «Интеграция палеэкологии и морфологии в оценку разнообразия теропод: случай от аптиан-албцев из Туниса» . Палеогеография, палеоклиматология, палеоэкология . 410 : 39–57. Bibcode : 2014ppp ... 410 ... 39f . doi : 10.1016/j.palaeo.2014.05.033 . ISSN 0031-0182 . S2CID 129225979 .
- ^ Churcher, CS (1999). «Примечание на покойной меловой позвоночной фауне оазиса Дахле». Монография Oxbow : 55-68.
- ^ Певица (2015). «Юрапарк на тропе новых динозавров из Марокко» . Jurapark . Архивировано из оригинала на Delta 6, 2015 . Получено 27 июня 2023 года .
- ^ Молина-Пресес, Рубен; Larramendi, Asier; Коннолли, Дэвид; Круз, Гонсало Ангел Рамирес (25 июня 2019 г.). Факты и цифры динозавров: Тнуподы и другие динозавриформные . ПРИЗНАЯ УНИВЕРСИТЕТА ПРИСЕТА. ISBN 978-0-691-19059-4 .
- ^ Crusafont-Pairó, M.; Адровер Р. (1966). «Первый представитель класса мамматического класса, найденный в мезозойском Испании». Теруэль (на испанском). 35 : 139–143.
- ^ Kuhne, WG; Crusafont-Pairo, M. (1968). Dec. "Mamíferos The Wealdinse Denmark Actta Geológica латиноамериканцы . 3 (5): 133–134.
- ^ Гасон, Франциско; Кобос, Альберто; Royo-Towers, Рафаэль; Мампель, Луис; Алькала, Луис (1 июня 2012 г.). Riodeva (Teruel, Испания) " Палайский 92 (2): 273–285. Bibcode 2012PdPe...92..273G: два 10.1007/s12549-012-0079-3: ISSN 1867-1 129930988S2CID
- ^ Бонд, Джеффри; Бромли К. (1 декабря 1970 г.). «Отложения с останками динозавров возле Гокве, Родезия». Палеогеография, палеоклиматология, палеоэкология . 8 (4): 313–327. Bibcode : 1970ppp ..... 8..313b . doi : 10.1016/0031-0182 (70) 90104-5 . ISSN 0031-0182 . S2CID 128716797 .
- ^ Munyikwa, D.; Сэмпсон, SD; Роджерс, RR; Форстер, Калифорния; Карри, Ка; Curtice, BD (1998). «Палеонтология позвоночных и геология формирования Гокве, Зимбабве». Журнал африканских наук о Земле . 27 (1): 142.
- ^ Медейрос, Мануэль; Schultz, Cesar (2002). «Динозаврская фауна" Koring Laje ", Creatace Medio из северо -восточной Бразилии» . Архив музея националь, Рио -де -Жанейро . 60 (3): 155–162.
- ^ Nothdurft & Smith 2002 , p. 109
- ^ Jump up to: а беременный Хендерсон, DM; Николс Р. (2015). «Баланс и сила-оценивая максимальный потенциал добычи больших хищных динозавров Carcharodontosaurus saharicus » . Анатомическая запись . 298 (8): 1367–1375. doi : 10.1002/ar.23164 . PMID 25884664 . S2CID 19465614 .
- ^ Seebacher, F. (2001). «Новый метод для расчета аллометрических отношений динозавров с длиной массы» (PDF) . Журнал палеонтологии позвоночных . 21 (1): 51–60. Citeseerx 10.1.1.462.255 . doi : 10.1671/0272-4634 (2001) 021 [0051: anmtca] 2.0.co; 2 . ISSN 0272-4634 . S2CID 53446536 .
- ^ Hurlburt, GS; Ridgely, RC; Witmer, LM (5 июля 2013 г.). «Относительный размер мозга и головного мозга у динозавров тиранозавридов: анализ с использованием количественных взаимосвязи и эндокастов мозга у существующих аллигаторов» . В Parrish, MJ; Молнар, Re; Керри, PJ; Koppelhus, EB (Eds.). Тиранозаврид палеобиология . Издательство Университета Индианы. С. 134–154. ISBN 978-0-253-00947-0 Полем Получено 20 октября 2013 года .
- ^ Jump up to: а беременный Кория, Родольфо А.; Керри, Филипп Дж.; Ортега, Франциско; Байано, Маттия А. (1 июля 2020 г.). «Ранний меловой теропод среднего размера среднего размера (Dinosauria, Saurischia) из формирования Mulichinco (верхний валангинский), провинция Нойкен, Патагония, Аргентина» . Мерашные исследования . 111 : 104319. BIBCODE : 2020CRRES.11104319C . doi : 10.1016/j.cretres.2019.104319 . HDL : 11336/122794 . ISSN 0195-6671 . S2CID 214475057 .
- ^ Ibrahim et al. 2020 , с. 165.
- ^ Кория, Родольфо А.; Керри, Филипп Дж. (14 января 2003 г.). «Мозговой мозг Giganotosaurus carolinii (Dinosauria: Theropoda) из верхнего мела Аргентины» . Журнал палеонтологии позвоночных . 22 (4): 802–811. doi : 10.1671/0272-4634 (2002) 022 [0802: tbogcd] 2.0.co; 2 . ISSN 0272-4634 . S2CID 85678725 .
- ^ Jump up to: а беременный в Novas, Fernando E.; де Вале, Сильвина; Виккерс-богатый, Пэт; Рич, Том (1 мая 2005 г.). «Большой меловой теропод из Патагонии, Аргентины, и эволюция карчадонтозавридов» . Naturwissenschaften . 92 (5): 226–230. Bibcode : 2005nw ..... 92..226n . doi : 10.1007/s00114-005-0623-3 . HDL : 11336/103474 . ISSN 1432-1904 . PMID 15834691 . S2CID 24015414 .
- ^ Jump up to: а беременный Бивор, Томас; Куигли, Аарон; Смит, Рой Э.; Смит, Роберт Ш; Ибрагим, Низар; Зухри, Самир; Мартилл, Дэвид М. (1 января 2021 г.). «Тафономические данные подтверждают водный образ жизни для спинозавра» . Мерашные исследования . 117 : 104627. Bibcode : 2021crres.11704627b . doi : 10.1016/j.cretres.2020.104627 . ISSN 0195-6671 . S2CID 224888268 .
- ^ «Открытия | Пол Серено - Палеонтолог | Чикагский университет» . Paulsereno.uchicago.edu . Получено 30 июня 2023 года .
- ^ Брусатт, Стивен Л.; Бенсон, Роджер Б.Дж.; Карр, Томас Д.; Уильямсон, Томас Э.; Серено, Пол С. (12 декабря 2007 г.). «Систематическая полезность эмалевых морщин теропод» . Журнал палеонтологии позвоночных . 27 (4): 1052–1056. doi : 10.1671/0272-4634 (2007) 27 [1052: tsuote] 2.0.co; 2 . ISSN 0272-4634 . S2CID 85615205 .
- ^ Jump up to: а беременный Larsson, HCE 2001. Эндокраниальная анатомия Carcharodontosaurus saharicus (Theropoda: Allosauroidea) и его последствия для эволюции мозга теропод. С. 19–33. В: Жизнь мезозойских позвоночных . Ed.S Tanke, DH, Carpenter, K., Skrepnick, MW Indiana University Press.
- ^ Харрис, Джеральд Д. (1998). «Реанализ AcroCanthosaurus atokensis , его филогенетический статус и палеобиологические последствия, основанные на новом образе из Техаса». Музей естественной истории и науки Нью -Мексико . 13 : 1–75.
- ^ Стромер 1934 , с. 60-61.
- ^ Jump up to: а беременный Novas, Fernando E.; Agnolín, Federico L.; Ezure, Martín D.; Порппхири, Джон; Каналель, Джон I. (1 октября 2013 г.). «Эволюция плотоядных динозавров во время меловых: доказательства из Патагонии » Мерашные исследования 45 : 174–2 Bibcode : 2013rres..45..174n Doi : 10.1016/ j.cretres.2013.04.0 HDL : 11336/1 ISSN 0195-6 S2CID 129504278
- ^ Jump up to: а беременный Canale, Juan I.; Apeesteguía, Sebastián; Галлина, Пабло А.; Митчелл, Джонатан; Смит, Натан Д.; Каллен, Томас М.; Шинья, Акико; Галуза, Алехандро; Gianechini, Federico A.; Маковицки, Питер Дж. (7 июля 2022 г.). «Новый гигантский плотоядный динозавр выявляет сходящиеся эволюционные тенденции в уменьшении теропода» . Текущая биология . 32 (14): 3195–3202.e5. Bibcode : 2022cbio ... 32E3195C . doi : 10.1016/j.cub.2022.05.057 . ISSN 0960-9822 . PMID 35803271 . S2CID 250343124 .
- ^ Jump up to: а беременный Кория, Родольфо А.; Сальгадо, Леонардо (1995). «Новый гигантский плотоядный динозавр из мелового периода Патагонии» . Природа . 377 (6546): 224–226. Bibcode : 1995natur.377..224c . doi : 10.1038/3772224A0 . ISSN 1476-4687 . S2CID 30701725 .
- ^ Брусатт, Стивен Л.; Бенсон, Роджер Б.Дж.; Сюй, Син (2012). «Переоценка Kelmayisaurus Petrolicus , крупного тероподного динозавра из раннего мела Китая» . Acta Palaeontologica Polonica . 57 (1): 65–72. doi : 10.4202/app.2010.0125 . HDL : 20.500.11820/95DE36FB-46B2-4ACC-BD2E-1D5A496FC78C . ISSN 0567-7920 . S2CID 53387460 .
- ^ Кау, Андреа (2024). «Унифицированная структура для макроэволюции хищных динозавров» (PDF) . Bollettino della società paleontologica italiana . 63 (1): 1-19. doi : 10.4435/bspi.2024.08 . Архивировано из оригинала (PDF) 27 апреля 2024 года . Получено 1 мая 2024 года .
- ^ Керри, Филипп Дж. (17 мая 1996 г.). «Из Африки: мясные динозавры, которые бросают вызов Tyrannosaurus rex ». Наука . 272 (5264): 971–972. Bibcode : 1996sci ... 272..971c . doi : 10.1126/science.272.5264.971 . ISSN 0036-8075 . S2CID 85110425 .
- ^ Кория, Родольфо А.; Сальгадо, Леонардо (июнь 1996 г.). «Плотоядные динозавры Южной Америки» . Исследования и наука (по -испански) (237): 39–40.
- ^ Canale, Ji; Novas, Fe; Пол Д. (2014). «Остеология и филогенетические отношения тиранотитана Chubutensis novas, de Valais, Vickers-Rich and Rich, 2005 (Theropoda: Carcharodontosauridae) из нижнего мела Патагонии, Аргентина» . Историческая биология . 27 (1): 1–32. doi : 10.1080/08912963.2013.861830 . HDL : 11336/17607 . S2CID 84583928 .
- ^ Ван Валкенбург, Блэр (1988). «Частота распада зубов среди крупных хищных млекопитающих» . Американский натуралист . 131 (2): 291–302. doi : 10.1086/284790 . ISSN 0003-0147 . JSTOR 2461849 . S2CID 2223330098 .
- ^ Ван Валкенбург, Блэр (2008). «Затраты на плотоявку: перелом зубов у плейстоцена и недавних карниворанов» . Биологический журнал Линневого общества . 96 (1): 68–81. doi : 10.1111/j.1095-8312.2008.01108.x . S2CID 85623961 .
- ^ Хон, Дэвид, мы; Раухут, Оливер В.М. (2010). «Поведение кормления и использование костей динозаврами Theropod» (PDF) . Летая . 43 (2): 232–244. Bibcode : 2010Letha..43..232H . doi : 10.1111/j.1502-3931.2009.00187.x . ISSN 0024-1164 . S2CID 86037076 .
- ^ Танк, Даррен Х.; Керри, Филипп Дж. (1998). «Поведение с кусочком у динозавров в Тероподах: палеопатологические данные» . Гайя (15): 167–184. doi : 10.7939/r34t6fj1p . S2CID 90552600 .
- ^ Драмхеллер, Стефани К.; MCHUGH, Джулия Б.; Кейн, Мириам; Ридель, Анджа; D'Amore, Domenic C. (27 мая 2020 г.). «Высокие частоты отметок укуса теропода предоставляют доказательства кормления, утилизации и возможного каннибализма в подчеркнутой поздней юрской экосистеме» . Plos один . 15 (5): E0233115. Bibcode : 2020ploso..1533115d . doi : 10.1371/journal.pone.0233115 . ISSN 1932-6203 . PMC 7252595 . PMID 32459808 .
- ^ Сакамото, Манабу (12 июля 2022 г.). «Оценка силы укуса в вымерших динозаврах с использованием филогенетических прогнозируемых физиологических площадей поперечного сечения мышц аддуктора челюсти» . ПЕРЕЙ . 10 : E13731. doi : 10.7717/peerj.13731 . ISSN 2167-8359 . PMC 9285543 . PMID 35846881 .
- ^ Гиньак, Пол М.; Эриксон, Грегори М. (17 мая 2017 г.). «Биомеханика, стоящая за экстремальной остеофагией в Tyrannosaurus rex » . Научные отчеты . 7 (1): 2012. BIBCODE : 2017NATSR ... 7.2012G . doi : 10.1038/s41598-017-02161-w . ISSN 2045-2322 . PMC 5435714 . PMID 28515439 .
- ^ Рэйфилд, Эмили Дж. (2011). «Структурные характеристики тетануранских черепах, с акцентом на мегалозауриды, спинозауриды и carcharodontosauridae» (PDF) . Специальные статьи в палеонтологии . 86 : 241–253. [ Постоянная мертвая ссылка ]
- ^ Jump up to: а беременный Goedert, J.; Амиот, Р.; Будад, Л.; Баффеттаут, Э .; Fourel, F.; Godefroit, P.; Sharp, N.; Suteetthor, v.; Тонг, H.; Watabe, M.; Lecuyer, C. (2016). Это композиции кислорода. Палуиос 31 (1): 10–1 Bibcode : 2016palai . doi : 10.2110/palo . S2CID 130878403 .
- ^ Амиот, Роман; Ван, Сюй; Лекуйер, Кристоф; Buffetaut, Eric; Будад, Ларби; Кавин, Лайонел; Дин, Чжунли; Fluteau, Frédéric; Келлнер, Александр Ва; Тонг, Хайян; Zhang, Fusong (2010). «Состав из кислорода и углерода средних меловых позвоночных из Северной Африки и Бразилии: экологическое и экологическое значение» . Палеогеография, палеоклиматология, палеоэкология . 297 (2): 439–451. Bibcode : 2010ppp ... 297..439a . doi : 10.1016/j.palaeo.2010.08.027 . S2CID 131607253 .
- ^ Амиот, Роман; Лекуйер, Кристоф; Buffetaut, Eric; Эскаргуэль, Жиль; Fluteau, Frédéric; Мартино, Франсуа (15 июня 2006 г.). «Изотопы кислорода из биогенных апатитов предполагают широкую эндотермию в меловых динозаврах» (PDF) . Земля и планетарные научные письма . 246 (1): 41–54. Bibcode : 2006e & psl.246 ... 41a . doi : 10.1016/j.epsl.2006.04.018 . ISSN 0012-821X . JSTOR 41125672 . S2CID 55100956 .
- ^ Чуре, Даниэль (2000). «На орбите динозавров Теропод» . Гайя . 15 : 233–240.
- ^ Стивенс, Кент А. (12 июня 2006 г.). «Бинокулярное зрение у динозавров с теропод» . Журнал палеонтологии позвоночных . 26 (2): 321–330. doi : 10.1671/0272-4634 (2006) 26 [321: bvitd] 2.0.co; 2 . ISSN 0272-4634 . JSTOR 4524572 . S2CID 85694979 .
- ^ Чинсами, Анусуя; Tumarkin-Deratzian, Allison (2009). «Патологические костные ткани в стервятнике индейки и неавийский динозавр: последствия для интерпретации эндостаальной кости и радиальной фиброламеллярной кости у ископаемых динозавров» . Анатомическая запись: достижения в области интегративной анатомии и эволюционной биологии . 292 (9): 1478–1484. doi : 10.1002/ar.20991 . PMID 19711479 . S2CID 41596233 .
- ^ Jump up to: а беременный Molnar, Re (2001). «Теропод палеопатология: литературное обследование». В Карпентере, Кеннет; СКРЕПНИК, Майкл Уильям; Танк, Даррен Х. (ред.). Мезозойская жизнь позвоночных . Индиана Univ. Нажимать. С. 337–363. ISBN 978-0-253-33907-2 Полем OCLC 248649755 .
- ^ Ротшильд, Брюс; Танк, Даррен (2005). «Теропод палеопатология: современный обзор». Плотоядные динозавры (1 -е изд.). Индианаполис: издательство Университета Индианы. п. 351. ISBN 9780253345394 .
- ^ Wanas, Hamdalla A.; Ассал, Эхаб М. (1 марта 2021 г.). «Происхождение, тектоническая обстановка и исходная зона, проведенное в области песчаников бахарии в Оазисе Бахарии, Египет: значение палеоклимама и палеогеографии южного региона Нео-Тетис во время раннего сеноманиана». Осадочная геология . 413 : 105822. Bibcode : 2021sedg..41305822W . doi : 10.1016/j.sedge.2020.105822 . ISSN 0037-0738 . S2CID 230598476 .
- ^ Хамед, Юнес; Аль-Гамал, Самир Анвар; Али, Вайсим; Нахид, Абедерацзак; DHIA, Хамед Бен (1 марта 2014 г.). «Палеоэрадома ископаемого континентального интеркалера из позднего мела (Барремано-албийца) в Северной Африке: тематическое исследование южного Туниса» . Аравийский журнал «Геоссауки» . 7 (3): 1165–1177. Bibcode : 2014Arjg .... 7.1165H . doi : 10.1007/s12517-012-0804-2 . ISSN 1866-7538 . S2CID 128755145 .
- ^ Ibrahim et al. 2020 , с. 185.
- ^ Амиот, Р.; Buffetaut, E.; Lécuyer, C.; Ван, х.; Будад, Л.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Мо, Дж.; Simon, L.; Suteethorn, v.; Sweetman, S.; Тонг, H.; Чжан, Ф.; Zhou, Z. (2010). «Изотопные доказательства кислорода для полуаватических привычек среди спинозавридов» (PDF) . Геология . 38 (2): 139–142. Bibcode : 2010geo .... 38..139a . doi : 10.1130/g30402.1 .
- ^ Ibrahim et al. 2020 , с. 184.
- ^ Холлидей, Кейси М.; Гарднер, Николас М. (31 января 2012 г.). «Новая эусухинская крокодилиформ с новым черепно -полным покровом и его значение для происхождения и эволюции крокодилии» . Plos один . 7 (1): E30471. Bibcode : 2012ploso ... 730471H . doi : 10.1371/journal.pone.0030471 . ISSN 1932-6203 . PMC 3269432 . PMID 22303441 .
- ^ Ibrahim et al. 2020 , с. 180, 189.
- ^ Ибрагим, Низар; Unwin, David M.; Martill, David M.; Байдер, Лахссен; Зухри, Самир (26 мая 2010 г.). «Новый птерозавр (Pterodactyloidea: Azhdarchidae) из верхнего мела Марокко» . Plos один . 5 (5): E10875. Bibcode : 2010ploso ... 510875i . doi : 10.1371/journal.pone.0010875 . ISSN 1932-6203 . PMC 2877115 . PMID 20520782 .
- ^ Борха Холгадо; Родриго В. Пегас (2020). «Таксономический и филогенетический обзор группы Anhanguerid Pterosaur Coloborhynchinae и New Clade Tropeognathinae» . Acta Poolonological Польша 65 (4): 743–761. doi : 10.4202/ app.00751.2 222075296S2CID
- ^ Ибрагим, н; Дал Сассо, C; Maganuco, S; Фаббри, м; Martill, D; Gorscak, E; Ламанна, М. (2016). «Свидетельство полученного титанозавриана (Dinosauria, Sauropoda) в« Кеем -кем -кемках »Марокко, с комментариями о палеоэкологии Sauropod в меловом фоне Африки» . Меловой период: биотическое разнообразие и биогеография. Музей естественной истории и науки Нью -Мексико . 71 : 149–159.
- ^ Hassler, A.; Мартин, JE; Амиот, Р.; Tacail, T.; Годо, Ф. Арно; Allain, R.; Balter, V. (11 апреля 2018 г.). «Изотопы кальция предлагают подсказки о разделении ресурсов среди меловых хищных динозавров» . Труды Королевского общества B: Биологические науки . 285 (1876): 20180197. DOI : 10.1098/rspb.2018.0197 . ISSN 0962-8452 . PMC 5904318 . PMID 29643213 .
- ^ Смит, Джошуа Б.; Ламанна, Мэтью С.; Лаковара, Кеннет Дж.; Додсон, Питер; Смит, Дженнифер Р.; Пул, Джейсон С.; Гигенгак, Роберт; Attia, Yousry (2001). «Гигантский динозавр Sauropod из верхнего мелового депозита в Египте» . Наука . 292 (5522): 1704–1706. Bibcode : 2001sci ... 292.1704S . doi : 10.1126/science.1060561 . ISSN 0036-8075 . PMID 11387472 . S2CID 33454060 .
- ^ Уилсон, Джеффри А.; Аллейн, Ронан (4 июля 2015 г.). «Остеология Ребебахисавра Гарасбай Лавокат, 1954, диплодокоид (Dinosauria, Sauropoda) из ранних поздних мелово -волновых кемских слоев юго -восточного Марокко» . Журнал палеонтологии позвоночных . 35 (4): E1000701. BIBCODE : 2015JVPAL..35E0701W . doi : 10.1080/02724634.2014.1000701 . ISSN 0272-4634 . S2CID 129846042 .
Библиография
[ редактировать ]- Стромер, Эрнст (1934). «Результаты исследовательской поездки профессора Э. Стромеров в пустынях Египта II. Остаток позвоночных на уровне Бахаридже (самый низкий сеноман). 13. Динозаурия» (PDF) . Трактаты Баварской академии наук математического и научного факультета . Новый эпизод (на немецком языке). 22 : 1–79.
- Nothdurft, Уильям; Смит, Джош (2002). Потерянные динозавры Египта . Нью -Йорк: Random House Publishing Group . ISBN 978-1-58836-117-2 .
- Ибрагим, Низар; Серено, Пол С.; Варриккио, Дэвид Дж.; Martill, David M.; Dutheil, Didier B.; Unwin, David M.; Байдер, Лахссен; Ларссон, Ганс Се; Зухри, Самир; Каукая, Абдельхади (2020). «Геология и палеонтология верхней меловодной группы Kem Kem of Eastern Morocco» . Зоуки (928): 1–216. Bibcode : 2020zook..928 .... 1i . doi : 10.3897/Zookeys.928.47517 . ISSN 1313-2970 . PMC 7188693 . PMID 32362741 .














