Мезонихия
| Мезонихия Временной диапазон:
| |
|---|---|

| |
| Гигантский череп гарпаголестеса | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Клэйд : | Пан-Эунгулата |
| Зеркальный порядок: | Эунгулата |
| Клэйд : | Параксония |
| Заказ: | † Мезонихия Ван Вален , 1966 год. |
| Семьи | |
Мезонихия («средние когти мелких и крупных размеров, ») — вымерший таксон плотоядных копытных родственный парнокопытным . Мезонихи впервые появились в раннем палеоцене , пошли на резкий упадок в конце эоцена и полностью вымерли, когда последний род, Mongolestes вымер в раннем олигоцене . Судя по их истории, в Азии они постепенно становились все крупнее и хищнее, а затем перешли к образу жизни, связанному с падальщиками и дроблением костей, прежде чем группа вымерла. [ 2 ]
наиболее примитивный мезонихий, Yantanglestes Мезонихи, вероятно, возникли в Китае, где с раннего палеоцена известен . Наиболее разнообразны они были также в Азии, где встречаются во всех основных фаунах палеоцена . Поскольку другие хищники, такие как креодонты и хищные животные , были либо редки, либо отсутствовали в этих сообществах животных, мезонихии, скорее всего, доминировали крупных хищников в нише в палеоцене Восточной Азии.
Один род , Dissacus , успешно распространился в Европе и Северной Америке к началу палеоцена. Диссакус был хищником размером с шакала , обитавшим по всему Северному полушарию. [ 3 ] но виды близкородственного или идентичного рода Ankalagon из раннего и среднего палеоцена Нью-Мексико были намного крупнее и вырастали до размеров медведя . [ 4 ] Более поздний род, Pachyaena , проник в Северную Америку в самом раннем эоцене, где развился в виды, которые были по крайней мере такими же крупными. Мезонихи Северной Америки были крупнейшими хищными млекопитающими в период от раннего палеоцена до среднего эоцена.
Характеристики
[ редактировать ]
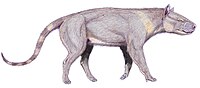
Мезонихий часто реконструировали как похожих на волков, хотя и внешне, но в жизни они выглядели совсем иначе. Имея короткий нижний отдел позвоночника, усиленный вращающимися суставами, они могли бы бегать с жесткой спиной, как современные копытные, а не прыгать или прыгать с гибкими позвоночниками, как современные хищники . Хотя позже у мезонихов развился набор приспособлений конечностей для бега, аналогичных тем, что есть у волков и оленей, их ноги оставались сравнительно толстыми. [ 5 ] Они не походили бы ни на одну группу живых животных. Ранние мезонихи, вероятно, ходили на ступнях ( стопоходные ), а более поздние — на носках ( пальцеходящие ). У этих более поздних мезонихий были копыта, по одному на каждом пальце, и по четыре пальца на каждой ноге. Стопа была сжата для эффективного бега с осью между третьим и четвертым пальцами стопы (параксоническая); это было бы похоже на копытную лапу. [ 6 ]
Мезонихи различались по размеру; некоторые виды были размером с лису, другие — с лошадь. Некоторые представители группы известны только по черепам и челюстям или имеют фрагментарные посткраниальные останки. Но там, где известны скелеты, они указывают на то, что у мезонихий были большие головы с сильными челюстными мышцами, относительно длинные шеи и крепкие тела с крепкими конечностями, которые могли эффективно бегать, но не могли вращать руками или тянуться в стороны. Неродственная ранняя группа хищников-млекопитающих, креодонты , также имела необычно большие головы и конечности, в которых гибкость менялась на эффективность бега; большой размер головы может быть связан с неспособностью использовать ноги и когти для ловли и переработки пищи, как это делают многие современные хищники. Некоторые мезонихи реконструируются как хищные (сравнимы с псовыми ), другие - как падальщики или хищники-падальщики с приспособлениями для дробления костей зубов (сравнимы с большими гиенами), а некоторые - как всеядны (сравнимы со свиньями, людьми или черными медведями). . Они могли не включать гиперплотоядные животные (сравнимы с кошачьими ); их зубы не были столь эффективны при разделке мяса, как у более поздних групп крупных хищников-млекопитающих. В некоторых местах в разных экологических нишах сосуществовали несколько видов или родов. Есть данные, позволяющие предположить, что некоторые роды имели половой диморфизм. [ 7 ] Некоторые роды могут нуждаться в пересмотре, чтобы уточнить фактическое количество видов или устранить двусмысленность в отношении родов (например, Dissacus и Ankalagon ). [ 5 ]
Эти «волки на копытах», вероятно, были одной из наиболее важных групп хищников в экосистемах позднего палеоцена и эоцена Европы (которая в то время была архипелагом), Азии (которая была островным континентом) и Северной Америки. Мезонихийский зубной ряд состоял из коренных зубов, модифицированных для создания вертикального сдвига, тонких лезвий нижних коренных зубов и хищных выемок, но не настоящих хищных зубов. Коренные зубы были сжаты с боков и часто тупые и, вероятно, использовались для разделки мяса или дробления костей. Считается, что самые крупные виды были падальщиками . Многие виды подозреваются в том, что они питаются рыбой, хотя на некоторые из этих реконструкций могут влиять более ранние теории о том, что эта группа была предком китообразных .
Филогения и эволюционные отношения
[ редактировать ]
Мезонихи долгое время считались креодонтами , но теперь были исключены из этого отряда и помещены в три семейства (Mesonychidae, Hapalodectidae и Triisodontidae), либо в пределах своего собственного отряда Mesonychia, либо в отряде Condylarthra как часть когорты или надотряда. Лавразиатерия . Почти все мезонихи в среднем крупнее большинства палеоценовых и эоценовых креодонтов и миакоидных хищников.
Орден иногда называют своим старым названием Акреоди .
Недавнее исследование показало, что мезонихии являются базальными копытными животными, наиболее близкими родственниками « арктоционид » Mimotricentes , Deuterogonodon и Chriacus . « Triisodontidae » могут быть парафилетическими. [ 1 ]
Отношения с китами
[ редактировать ]Мезонихи обладают необычными треугольными коренными зубами, которые похожи на зубы китообразных (китов и дельфинов), особенно археоцетов , а также имеют аналогичную анатомию черепа и другие морфологические особенности. По этой причине ученые долгое время считали, что мезонихи были прямыми предками китообразных, но открытие хорошо сохранившихся задних конечностей архаичных китообразных, а также более поздние филогенетические анализы [ 8 ] [ 9 ] [ 10 ] теперь показывают, что китообразные более тесно связаны с гиппопотамидами и другими парнокопытными , чем с мезонихиями, и этот результат согласуется со многими молекулярными исследованиями. [ 11 ] Сходство зубных рядов и черепа может быть результатом того, что примитивные структуры копытных в родственных группах независимо развивались для удовлетворения тех же потребностей, что и хищники; некоторые исследователи предположили, что отсутствие первого пальца ноги и редуцированная плюсневая кость являются базальными особенностями ( синапоморфиями ), указывающими на то, что мезонихи, непарнокопытные и парнокопытные являются сестринскими группами. [ 5 ]
Большинство палеонтологов в настоящее время сомневаются в том, что киты произошли от мезонихий, и вместо этого предполагают, что мезонихи произошли от базальных копытных , а китообразные произошли от продвинутых копытных ( парнокопытных , либо имеющих общего с ними предка ), либо происходящих от антракотериев (полуводных предков копытных ). бегемоты). [ 12 ] Однако тесная группировка китов с бегемотами в кладистическом анализе появляется только после удаления Эндрюсарха , который часто включался в мезонихии. [ 13 ] [ 14 ] Один из возможных выводов состоит в том, что Эндрюсарх был классифицирован неправильно. Нынешняя неопределенность может частично отражать фрагментарный характер останков некоторых важнейших ископаемых таксонов, таких как Эндрюсарх . [ 13 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б Сара Л. Шелли, Томас Э. Уильямсон, Стивен Л. Брусатте, Разрешение филогенетических отношений более высокого уровня «Triisodontidae» («Condylarthra») внутри плаценталии, октябрь 2015 г., Общество палеонтологии позвоночных (аннотация)
- ^ «Новые млекопитающие-мезонихиды, обнаруженные из нижнего палеогена Эрляньской котловины, Ней-Монгол» . физ.орг . Проверено 16 мая 2019 г.
- ^ Йеле, Мартин (2006). «Плотоядные, креодонты и хищные копытные: млекопитающие становятся хищниками» . Палеоценовые млекопитающие мира (онлайн) .
- ^ О'Лири, Морин А.; Лукас, Спенсер Г.; Уильямсон, Томас Э. (2000). «Новый экземпляр Анкалагона (Mammalia, Mesonychia) и свидетельства полового диморфизма у мезонихий». Журнал палеонтологии позвоночных . 20 (2): 387–93. doi : 10.1671/0272-4634(2000)020[0387:ANSOAM]2.0.CO;2 . JSTOR 4524103 .
- ^ Jump up to: а б с «Мезоникс и другие мезонихиды-мезонихии (мезонихии, часть IV) | ScienceBlogs» . scienceblogs.com . Проверено 16 мая 2019 г.
- ^ О'Лири, Морин А.; Роуз, Кеннет Д. (1995). «Посткраниальный скелет раннеэоценовой мезонихиды Pachyaena (млекопитающие: Mesonychia)». Журнал палеонтологии позвоночных . 15 (2): 401–430. дои : 10.1080/02724634.1995.10011238 . ISSN 0272-4634 . JSTOR 4523639 .
- ^ О'Лири, Морин А.; Лукас, Спенсер Г.; Уильямсон, Томас Э. (27 июня 2000 г.). «Новый экземпляр Анкалагона (Mammalia, Mesonychia) и свидетельства полового диморфизма у мезонихий». Журнал палеонтологии позвоночных . 20 (2): 387–393. doi : 10.1671/0272-4634(2000)020[0387:ANSOAM]2.0.CO;2 . ISSN 0272-4634 .
- ^ Гейслер, Джонатан Х.; Уэн, Марк Д. (2003). «Морфологическое подтверждение тесной связи бегемотов и китов». Журнал палеонтологии позвоночных . 23 (4): 991–6. дои : 10.1671/32 . JSTOR 4524409 . S2CID 59143599 .
- ^ Гейслер, Джонатан Х.; Уэн, Марк Д. (2005). «Филогенетические взаимоотношения вымерших цетартиодактилей: результаты одновременного анализа молекулярных, морфологических и стратиграфических данных». Журнал эволюции млекопитающих . 12 (1–2): 145–60. дои : 10.1007/s10914-005-4963-8 . S2CID 34683201 .
- ^ Буассери, младший; Лихоро, Ф.; Брюне, М. (2005). «Положение бегемотов внутри Cetartiodactyla» . Труды Национальной академии наук . 102 (5): 1537–41. Бибкод : 2005PNAS..102.1537B . дои : 10.1073/pnas.0409518102 . JSTOR 3374466 . ПМК 547867 . ПМИД 15677331 .
- ^ Гейтси, Дж.; Хаяси, К.; Кронин, Массачусетс; Арктандер, П. (1996). «На основании генов молочного казеина получены доказательства того, что китообразные являются близкими родственниками гиппопотамидных парнокопытных» . Молекулярная биология и эволюция . 13 (7): 954–63. doi : 10.1093/oxfordjournals.molbev.a025663 . ПМИД 8752004 .
- ^ Гейслер, Джонатан Х.; Теодор, Джессика М. (2009). «Филогения бегемотов и китов». Природа . 458 (7236): E1–4, обсуждение E5. Бибкод : 2009Natur.458....1G . дои : 10.1038/nature07776 . ПМИД 19295550 . S2CID 4320261 .
- ^ Jump up to: а б Тьювиссен, JGM ; Купер, Лиза Ноэль; Клементц, Марк Т.; Баджпай, Сунил; Тивари, Б.Н. (2009). «Тевиссен и др. Ответ» . Природа . 458 (7236): Е5. Бибкод : 2009Natur.458....5T . дои : 10.1038/nature07775 . S2CID 4431497 .
- ^ О'Лири, Морин А.; Гейтси, Джон (2008). «Влияние увеличения выборки признаков на филогению Cetartiodactyla (Mammalia): комбинированный анализ, включая окаменелости» . Кладистика . 24 (4): 397–442. дои : 10.1111/j.1096-0031.2007.00187.x . ПМИД 34879630 . S2CID 85141801 .