Креодонта
| Креодонта | |
|---|---|

| |
| Различные эоцена Колорадо . , США креодонты Сверху: Тритемнодон , Патриофелис , Махаероидес и Синопа. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Зеркальный порядок: | Дикий |
| Заказ: | † Креодонта Коуп , 1875 г. [ 1 ] |
| Семьи | |
| Синонимы | |
|
список синонимов: | |
Creodonta («мясные зубы») — бывший отряд вымерших плотоядных , плацентарных млекопитающих обитавших с раннего палеоцена до позднего миоцена в Северной Америке , Европе , Азии и Африке . Первоначально считалось, что это единая группа животных, являющаяся предком современных хищников , теперь этот отряд обычно считается полифилетической совокупностью двух разных групп, оксиенид и гиенодонтид , а не естественной группой. Оксиениды впервые известны из палеоцена Северной Америки, а гиенодонты родом из палеоцена Африки. [ 10 ]
Креодонты были доминирующими плотоядными млекопитающими 55-35 миллионов лет назад , достигнув пика разнообразия и распространенности в эоцене . [ 11 ] Первые крупные, очевидно хищные млекопитающие появились с радиацией оксиенид в позднем палеоцене. [ 12 ] В палеогене виды «креодонтов» были самыми многочисленными наземными хищниками в Старом Свете. [ 13 ] В олигоценовой Африке гиенодонты были доминирующей группой крупных плотоядных, сохранявшейся до середины миоцена .
Группы «креодонтов» имели обширный ареал как географически, так и во времени. Они известны от позднего палеоцена до позднего олигоцена в Северной Америке , от раннего эоцена до позднего олигоцена в Европе , от позднего палеоцена до позднего миоцена в Азии и от позднего палеоцена до позднего миоцена в Африке . [ 14 ] Хотя большинство из них были млекопитающими малого и среднего размера, среди них был саркастодон , один из крупнейших наземных хищников всех времен, весом около 800 кг. [ 15 ]
Хотя часто предполагается, что хищники вытеснили их, эмпирических подтверждений этому мало. Последний род, Dissopsalis , вымер около 11,1 миллиона лет назад .
Большинство современных палеонтологов согласны с тем, что оба семейства «креодонтов» связаны с Carnivora , но не являются их прямыми предками. До сих пор неясно, насколько тесно эти две семьи связаны друг с другом. В целом классификация осложняется тем фактом, что взаимоотношения между ископаемыми млекопитающими обычно определяются сходством зубов, но зубы гиперплотоядных видов могут приобретать сходную форму в результате конвергентной эволюции , чтобы справиться с механикой поедания мяса. [ 16 ]
«Креодонты», как и многие другие хищные млекопитающие, разделяют хищнические ножницы , похожую на ножницы модификацию верхних и нижних щечных зубов, которая использовалась для разрезания мышечной ткани. Эта адаптация наблюдается и у других клад хищных млекопитающих.
Систематика и история
[ редактировать ]«Креодонта» была придумана Эдвардом Дринкером Коупом в 1875 году. [ 1 ] Коуп включил оксиенид и виверравида Didymictis , но не включил Hyaenodontidae . В 1880 г. он расширил этот термин, включив в него семейства Miacidae (включая Viverravidae), Arctocyonidae , Leptictidae (ныне Pseudorhyncocyonidae ), Oxyaenidae , Ambloctonidae и Mesonychidae . [ 17 ] Первоначально Коуп поместил креодонтов в состав насекомоядных . Однако в 1884 году он рассматривал их как базальную группу, от которой произошли как хищные, так и насекомоядные животные. [ 18 ]
Hyaenodontidae не входили в число креодонтов до 1909 года. [ 19 ] Уильям Диллер-Мэтью рассматривал Creodonta как подотряд отряда Carnivora , разделенный на три группы:
- «Инадаптивные креодонты» (Creodonta inadaptiva), группа, в которую входят «псевдокреоди» (оксиениды и гиенодонтиды) и мезонихиды ,
- «Адаптивная креодонта» (Creodonta Adaptiva), состоящая из миацид и таксонов, входящих в корзину « Arctocyonidae »,
- и «Примитивная Креодонта» (Creodonta primitiva), состоящая из Oxyclaenidae .
Со временем из этого отряда были исключены различные группы и виды. В середине 20 века он стабилизировался и стал представлять собой оксиениды, гиенодонты, мезонихиды и арктоциониды. [ 20 ] под которыми понимались основные группы плотоядных плацентарных млекопитающих, не входящих в состав хищных. Становилось все более очевидным, что арктоциониды были таксоном мусорной корзины , а мезонихиды могли быть более тесно связаны с копытными . К 1969 году Creodonta содержала только оксиениды и гиенодонтиды. [ 21 ]
Совсем недавно «Creodonta» считалась недействительной полифилетической совокупностью плотоядных плацентарных млекопитающих (а не естественной группой), а члены Creodonta были сестринскими таксонами Carnivoramorpha (хищники и их стволовые родственники) в кладе Pan-Carnivora ( в mirorder Ferae ), разделенных на две группы: отряд Oxyaenodonta как одна группа и отряд Hyaenodonta плюс его стволовые родственники (семейство Wyolestidae и роды Altacreodus , Simidectes и Tinerhodon ) в другом. [ 22 ] [ 23 ] [ 24 ] [ 25 ] [ 26 ] Однако некоторые филогенетические анализы восстанавливают их как естественную группу, например, филогенетический анализ млекопитающих палеоцена, опубликованный в 2015 году, который подтвердил монофилию Creodonta и отнес эту группу к родственникам клады Pholidotamorpha ( панголины и их стволовые родственники). [ 27 ]
Полли утверждал, что единственная доступная синапоморфия между оксиенидами и гиенодонтидами - это большая метастилярная пластинка на первом моляре (M1), но он считает, что эта особенность характерна для всех базальных эутерий. [ 28 ] Отделение Oxyaenidae от Hyaenodontidae также согласуется с биогеографическими данными, поскольку первые оксиениды известны из раннего палеоцена Северной Америки, а первые гиенодонтиды происходят из очень позднего палеоцена Северной Африки. [ 12 ]
Эту договоренность усложняет предварительное одобрение Ганнелла. [ 14 ] возникновения третьего семейства Limnocyonidae . [ 29 ] В группу входят таксоны, которые когда-то считались оксиенидами, такие как Limnocyon , Thinocyon. [ 19 ] и Пролимноцион . [ 30 ] Вортман даже выделил подсемейство Limnocyoninae среди оксиенид. [ 31 ] Ван Вален гнездит то же подсемейство (включая Oxyaenodon ) в составе Hyaenodontidae. [ 21 ] Ганнелл не знает, являются ли Limnocyonidae группой внутри Hyaenodontidae (хотя и родственной группой остальным гиенодонтидам) или полностью отдельной группой. [ 14 ]
По мнению Ганнелла, отличительными чертами оксиенид являются: Небольшая черепная коробка в нижней части черепа. Затылок широкий у основания и сужается сверху (что придает ему треугольную форму). Слезная кость образует на лице полукруглое расширение. Нижние челюсти имеют тяжелый симфиз . М1 и м2 образуют хищных, а М3/м3 отсутствуют. Рука стопа и . стопоходящие или субстопоходящие сочленяется Малоберцовая кость с пяточной костью , а таранная кость - с кубовидной костью . Фаланги сжаты и растрескались на кончиках. [ 14 ]
Аналогичным образом, список определяющих особенностей гиенодонтид, составленный Ганнеллом, включает в себя: Длинный узкий череп с узким базикранием и высоким узким затылком. Лобные кости вогнуты между глазничными областями . М2 и м3 образуют хищных. M3 присутствует у большинства видов, тогда как m3 присутствует всегда. Рука и стопа варьируются от стопоходящих до пальцевидных. Малоберцовая кость сочленяется с пяточной костью, а таранно-кубовидное сочленение редуцировано или отсутствует. Концевые фаланги сжаты и трещиноваты на кончиках. [ 14 ]
Лимноциониды по Ганнелу имели следующие особенности: М3/м3 редуцированы или отсутствовали, остальные зубы не редуцированы. Трибуна была удлиненной. Сами животные были от маленького до среднего размера. [ 14 ]
Морфология
[ редактировать ]Зубной ряд
[ редактировать ]Среди примитивных креодонтов зубная формула — 3.1.4.3 3.1.4.3 , но более поздние формы часто имели уменьшенное количество резцов, премоляров и/или коренных зубов. [ 32 ] Клыки всегда большие и заостренные. Боковые резцы большие, а медиальные резцы обычно маленькие. [ 14 ] Премоляры примитивные, с одним первичным бугорком и различными вторичными бугорками. [ 13 ]
Креодонты имеют две или три пары хищных зубов . Одна пара выполняла наибольшую режущую функцию (либо М1/м2, либо М2/м3). [ 14 ] Такое расположение отличается от современных хищников, которые используют P4 и M1 для хищников. [ 33 ] Это различие предполагает конвергентную эволюцию мясоедов с отдельной эволюционной историей и различием на уровне отряда. [ 34 ] учитывая, что разные зубы развивались как хищные как между креодонтами и хищниками, так и между оксиенидами и гиенодонтами. Хищники также известны в других кладах плотоядных млекопитающих, таких как вымершая летучая мышь Necromantis , а также в совершенно неродственных таксонах, таких как плотоядное сумчатое Thylacoleo .
У двух основных групп креодонтов были задействованы разные коренные зубы. У Oxyaenidae M1 и m2 образуют хищных. Среди гиенодонтид это М2 и м3. В отличие от большинства современных хищных животных, у которых хищные являются единственными режущими зубами, другие коренные зубы креодонтов выполняют подчиненные режущие функции. [ 19 ] Разница в том, какие зубы образуют хищники, является основным аргументом в пользу полифилии Creodonta.

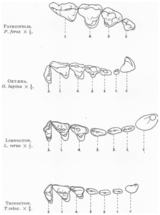
Череп
[ редактировать ]Креодонты имели длинные узкие черепа с маленьким мозгом. Череп значительно сузился за глазами, образуя отчетливые спланхнокраниума и нейрокраниума сегменты . У них были большие сагиттальные гребни и обычно широкие сосцевидные отростки (которые, вероятно, были производными чертами этой группы). [ 14 ] У многих креодонтов были пропорционально большие головы. [ 13 ] У примитивных форм слуховые буллы не были окостеневшими. В целом височные ямки были очень широкими. [ 14 ]





Посткраниальный скелет
[ редактировать ]Креодонты имели генерализованный посткраниальный скелет. Их конечности были меаксоническими (ось стопы проходила через середину пяти пальцев). Их метод передвижения варьировался от стопоходящего до пальцевого . Конечные фаланги представляли собой сросшиеся когти. [ 14 ]


Размер
[ редактировать ]Размеры креодонтов варьировались от размеров маленькой кошки до 800-килограммовых (1800 фунтов) саркастодонов . Однако более крупные животные были известны только в конце палеоцена, когда появились оксиениды. [ 12 ] размером с пуму такие как Dipsalidictis и, вероятно, сокрушающий кости падальщик Dipsalodon . [ 35 ]
Некоторые креодонты ( Arfia , Prolimnocyon и Palaeonictis ), по-видимому, испытали явление карликовости во время палеоцен-эоценового термического максимума, наблюдаемое у других родов млекопитающих. Предлагаемое объяснение этого явления заключается в том, что повышенный уровень углекислого газа в атмосфере напрямую повлиял на хищников из-за повышения температуры и засушливости, а также косвенно повлиял на них, уменьшив размер их травоядных жертв из-за того же давления отбора. [ 36 ]
Самый крупный североамериканский креодонт — Patriofelis . Экземпляр P.ferox, собранный в бассейне Бриджер на юге Вайоминга, был размером с взрослого черного медведя с головой, почти размером с голову взрослого льва-самца. [ 37 ]
Во время Среднеазиатской экспедиции 1930 года Американского музея естественной истории был собран самый крупный когда-либо обнаруженный креодонт: Sarkastodon mongoliensis . Его размеры были описаны как на 50% больше, чем у Patriofelis, с которым он имел много общего. [ 38 ] Было подсчитано, что саркастодон достиг массы тела, вдвое превышающей самый крупный американский лев. [ 15 ]
Биология
[ редактировать ]Диета и кормление
[ редактировать ]Ранние креодонты (как оксиениды, так и гиенодонтиды) имели трибосфенические коренные зубы, общие для базальных териев . Мелкие формы имели несколько сильные постметакриста-метазвездные гребни. [ 39 ] предполагая, что они, вероятно, были оппортунистическими кормушками, поедая такие вещи, как яйца, птицы, мелкие млекопитающие, насекомые и, возможно, также растительные вещества, [ 14 ] возможно, как современные виверровые . [ 32 ] Более крупные формы имели большую режущую способность, и со временем эта способность увеличивалась. Арфия , одно из наиболее распространенных хищных млекопитающих в раннем эоцене Северной Америки, в течение раннего эоцена развило более открытый тригонид на M3, что увеличило режущую способность хищных животных. [ 39 ] Аналогичное развитие можно увидеть, сравнив Oxyaena , Prototomus и Limnocyon с более мелкими и более распространенными питателями среди креодонтов. [ 14 ]
Вымирание
[ редактировать ]Некоторые теории предполагают, что гиенодонты и оксиениды вымерли, потому что их вытеснили недавно появившиеся хищники . Однако прямых доказательств того, что существование крупных хищных животных стало причиной исчезновения этих таксонов, нет, и во многих случаях (в Африке на протяжении раннего и среднего миоцена, а также в Северной Америке и Евразии на протяжении большей части олигоцена) гиенодонты процветали в среда, в которой в качестве конкурентов также присутствовали крупные хищники, такие как нимравиды и (позже) более крупные амфициониды. Теории, которые предполагают, что хищные животные вытеснили их, включают в себя то, что их меньший мозг ограничивал их интеллект, но размеры мозга хищных животных не всегда были постоянно большими на протяжении всей их эволюции, а важность размера мозга как фактора интеллекта в прошлом сильно переоценивалась. когда эти идеи были опубликованы. Другие предположения сосредоточены на строении их конечностей, которое ограничивало движение ног вертикальной плоскостью, как у копытных; они не могли повернуть запястья и предплечья внутрь, чтобы споткнуться, порезать или схватить добычу, как это могут сделать некоторые современные хищники. Креодонтам приходилось полностью полагаться на свои челюсти, чтобы поймать добычу, и, возможно, поэтому у креодонтов обычно был больший размер головы по сравнению с телом, чем у хищников аналогичного роста. Однако многие хищники, например крупные псовые также зависят только от своих челюстей при захвате добычи, но делают это эффективно даже в ситуациях, когда им приходится ловить крупную добычу в одиночку, поэтому это также не дает удовлетворительного объяснения.
У хищных животных последний верхний премоляр и первый нижний коренной зуб являются хищными , что позволяет самым задним коренным зубам развить приспособления для питания немясной пищей. У креодонтов первичными хищными были либо первые верхние и вторые нижние коренные зубы, либо вторые верхние и третьи нижние коренные зубы, а задние зубы образовывали хищнический ряд. Эта структура обязывала их есть почти исключительно мясо, что, возможно, ограничивало их способность использовать экологические ниши мезохищных и всеядных животных . Эти различия, возможно, привели к тому, что изменения окружающей среды повлияли на гиенодонтов и оксиенид иначе, чем на многих хищных животных, поскольку первые были бы ограничены в основном или полностью фауноядными диетами, в то время как многие (хотя и не все) линии хищных животных могли/способны существовать за счет растений. тоже имеет значение.
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Коуп, Эдвард Дринкер (1875). О предполагаемых хищниках эоцена Скалистых гор . Том. 27. С. 444–449.
{{cite book}}:|work=игнорируется ( помогите ) - ^ Т. К. Винклер (1893 г.) «Позвоночные животные прошлого». Палеонтологические исследования в музее Тельера 1-291
- ^ Кеннет Э. Кинман (1994.) «Система Кинмана: на пути к стабильной кладисто-эклектической классификации организмов: живые и вымершие, 48 типов, 269 классов, 1719 отрядов», Хейс, Канзас (почтовый ящик 1377, Hays 67601) , 88 страниц
- ^ Артур Сперри Пирс, (1936) «Зоологические названия. Список типов, классов и отрядов, подготовленный для раздела F, Американская ассоциация содействия развитию науки» Американская ассоциация развития науки
- ^ Миклош Крецой (1945) «Замечания о системе Хищника». Annales Historico-Naturales Musei Nationalis Hungarici, Будапешт, том. 38, стр. 59-83.
- ^ Ромер, А.С. (1966) «Палеонтология позвоночных». Издательство Чикагского университета, Чикаго; 3-е издание ISBN 0-7167-1822-7
- ^ Крецой, Н. (1929.) «Материалы для филогенетической классификации элуроидов. X Международный зоологический конгресс, Будапешт, 1927., 2, 1293–1355.
- ^ WD Мэтью (1909.) «Хищники и насекомоядные животные бассейна Бриджер, средний эоцен». Мемуары Американского музея естественной истории 9: 289-567.
- ^ Труессар, Э.Л. (1879 г.) «Каталог живых и ископаемых млекопитающих. III. Насекомоядные». Преподобный. Маг. Зоол. 3-й вечер. 7: 219 – 285.
- ^ Соле, Ф.; Люлье, Дж.; Адачи, М.; Бенсала, М.; Махбуби, М.; Табусе, Р. (2013). «Hyaenodontidans из района Гоур-Лазиб (? Ранний эоцен, Алжир): значения, касающиеся систематики и происхождения Hyainailourinae и Teratodontinae». Журнал систематической палеонтологии . 12 (3): 303–322. дои : 10.1080/14772019.2013.795196 . S2CID 84475034 .
- ^ Ламберт, Дэвид; и др. (Группа Диаграмм) (1985). Полевое руководство по доисторической жизни . Нью-Йорк: факты о файловых публикациях. ISBN 978-0-8160-1125-4 .
- ^ Перейти обратно: а б с Дженис, Кристин М.; Баскин, Джон А.; Берта, Анналиса; Флинн, Джон Дж.; Ганнелл, Грегг Ф.; Хант, Роберт М. младший; Мартин, Ларри Д.; Мунте, Кэтлин (1998). «Плотоядные млекопитающие» . В Дженисе, Кристина М.; Скотт, Кэтлин М.; Джейкобс, Луи Л. (ред.). Наземные хищники, копытные и копытные млекопитающие . Эволюция третичных млекопитающих Северной Америки. Том. 1. Кембридж: Издательство Кембриджского университета. стр. 73–90. ISBN 978-0-521-35519-3 .
- ^ Перейти обратно: а б с Роуз, Кеннет Дэвид; Арчибальд, Дж. Дэвид (2005). Возникновение плацентарных млекопитающих: происхождение и взаимоотношения основных существующих клад . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса.
- ^ Перейти обратно: а б с д и ж г час я дж к л м Ганнелл, Грегг Ф. (1998). «Креодонта» . В Дженисе, Кристин М.; Скотт, Кэтлин М.; Джейкобс, Луи Л. (ред.). Наземные хищники, копытные и копытные млекопитающие . Эволюция третичных млекопитающих Северной Америки. Кембридж, Великобритания: Издательство Кембриджского университета. стр. 91–109. ISBN 978-0-521-35519-3 .
- ^ Перейти обратно: а б Соркин, Борис (декабрь 2008 г.) [10 апреля 2008 г.]. «Биомеханическое ограничение массы тела у наземных хищников-млекопитающих» . Летайя . 41 (4): 333–347. дои : 10.1111/j.1502-3931.2007.00091.x .
- ^ Муизон, Кристиан; Ланге-Бадре, Бриджит (29 марта 2007 г.). «Плотоядные зубные адаптации трибосфенических млекопитающих и филогенетическая реконструкция» . Летайя . 30 (4): 353–366. дои : 10.1111/j.1502-3931.1997.tb00481.x .
- ^ Коуп, ЭД (март – декабрь 1880 г.). «О родах Креодонтов». Труды Американского философского общества . 19 (107): 76–82. JSTOR 982610 .
- ^ Коуп, ЭД (1884). Позвоночные третичных формаций Запада . Вашингтон, округ Колумбия: Типография правительства США. дои : 10.3133/70038954 .
- ^ Перейти обратно: а б с Мэтью, Уильям Диллер (август 1909 г.). «Хищники и насекомоядные животных бассейна Бриджер, средний эоцен» . Мемуары Американского музея естественной истории . 9 : 289–576. hdl : 2246/5744 .
- ^ Рассел, Лорис С. (1954). «Свидетельства строения зубов о взаимоотношениях ранних групп хищных животных». Эволюция . 8 (2): 166–171. дои : 10.2307/2405640 . ISSN 0014-3820 . JSTOR 2405640 .
- ^ Перейти обратно: а б Ван Вален, Ли М. (1966). «Дельтатеридии, новый отряд млекопитающих» . Бюллетень Американского музея естественной истории . 132 . hdl : 2246/1126 .
- ^ Бортс, Мэтью Р.; Стивенс, Нэнси Дж (2017). «Личный прикус и прорезывание зубов Hyainailouroidea (Hyaenodonta, Creodonta, Placentalia, Mammalia)» . Электронная палеонтология . 20 (3):55А. дои : 10.26879/776 .
- ^ Соле, Флореаль; Ладевез, Сандрин (2017). «Эволюция гиперплотоядных зубных рядов млекопитающих (Metatheria, Eutheria) и ее влияние на развитие трибосфенических коренных зубов». Эволюция и развитие . 19 (2): 56–68. дои : 10.1111/ede.12219 . ПМИД 28181377 . S2CID 46774007 .
- ^ Превости, Ф.Дж., и Форасиепи, AM (2018). «Введение. Эволюция южноамериканских млекопитающих-хищников в кайнозое: палеобиогеографические и палеоэкологические факторы»
- ^ Мэтью Р. Бортс; Нэнси Дж. Стивенс (2019). « Африканские хищники , род и виды ноябрь (Hyainailourinae, Hyaenodonta, Creodonta, Mammalia), гигантские хищники из раннего миоцена Кении » Журнал палеонтологии позвоночных . 39 (1): e1570222. Бибкод : 2019JVPal..39E0222B . дои : 10.1080/02724634.2019.1570222 . S2CID 145972918 .
- ^ Флореаль Соле; Бернар Марандат; Фабрис Лиоро (2020). «Гиенодонты (Mammalia) из французского местонахождения Амелас (Эро) с возможными новыми представителями из позднего Ипра» . Геодиверситас . 42 (13): 185–214. doi : 10.5252/geodiversitas2020v42a13 . S2CID 219585388 .
- ^ Холлидей, Томас, доктор медицинских наук; Апчерч, Пол; Госвами, Анджали (2015). «Разрешение взаимоотношений плацентарных млекопитающих палеоцена» (PDF) . Биологические обзоры . 92 (1): 521–550. дои : 10.1111/brv.12242 . ISSN 1464-7931 . ПМК 6849585 . ПМИД 28075073 .
- ^ Полли, П.Д. (1994). «Что такое креодонт?». Журнал палеонтологии позвоночных . 14 :42А. дои : 10.1080/02724634.1994.10011592 .
- ^ Газин, Чарльз Льюис (17 января 1962 г.). «Дальнейшее исследование фауны млекопитающих нижнего эоцена юго-западного Вайоминга» (PDF) . Разные коллекции Смитсоновского института . Вашингтон, округ Колумбия: Смитсоновский музей. стр. 1–98.
- ^ Мэтью, Уильям Диллер; Грейнджер, Уолтер (1915). «Пересмотр фауны нижнего эоцена Уосатч и Уинд-Ривер» . Бюллетень Американского музея естественной истории . 34 : 4–103. hdl : 2246/1373 . В этой статье авторы переименовывают Limnocyon protenus Марша в Didymictis protenus и включают его в число миацидов.
- ^ Вортман, Дж. Льюис (июль 1902 г.). «Исследования эоценовых млекопитающих в коллекции Марша, Музей Пибоди» . Американский научный журнал . стр. 17–23.
- ^ Перейти обратно: а б Денисон, Роберт Хауленд (октябрь 1937 г.). «Широкочерепные псевдокреоды». Анналы Нью-Йоркской академии наук . 37 (1): 163–255. Бибкод : 1937NYASA..37..163D . дои : 10.1111/j.1749-6632.1937.tb55483.x . S2CID 129936019 . (Требуется подписка или оплата.)
- ^ Фельдхамер, Джордж А.; Дрикамер, Ли С.; Весси, Стивен Х.; Мерритт, Джозеф Ф.; Краевски, Кэри (2015). Маммология: адаптация, разнообразие, экология . Балтимор: Издательство Университета Джонса Хопкинса. п. 356 . ISBN 978-0-8018-8695-9 .
- ^ Тернер, Алан; Энтони, Морис (2004). Развивающийся Эдем: иллюстрированное руководство по эволюции фауны крупных млекопитающих Африки . Нью-Йорк: Издательство Колумбийского университета. п. 77. ИСБН 978-0-231-11944-3 .
- ^ Ганнелл, Грегг Ф.; Джинджерич, Филип Д. (30 сентября 1991 г.). «Систематика и эволюция позднепалеоценовых и раннеэоценовых Oxyaenidae (Mammalia, Creodonta) в бассейне Кларкс-Форк, Вайоминг» (PDF) . Материалы Музея палеонтологии Мичиганского университета . Анн-Арбор: Музей палеонтологии Мичиганского университета. стр. 141–180.
- ^ Честер, Стивен ГБ; Блох, Джонатан И.; Секорд, Росс; Бойер, Дуг М. (2010). «Новый мелкотелый вид Palaeonictis (Creodonta, Oxyaenidae) из палеоцен-эоценового термического максимума» . Журнал эволюции млекопитающих . стр. 227–243.
- ^ Вортман, Джейкоб Л. (1894). «Остеология Patriofelis, креодонта среднего эоцена» (PDF) . Бюллетень Американского музея естественной истории . стр. 129–164.
- ^ Грейнджер, Уолтер (21 апреля 1938 г.). «Гигантская оксиенида из верхнего эоцена Монголии» (PDF) . Американский музей Новитатов . стр. 1–5.
- ^ Перейти обратно: а б Джинджерич, Филип Д. (1989). «Новая фауна древнейших млекопитающих васатчи из эоцена северо-западного Вайоминга: состав и разнообразие в редко отобранном комплексе высокой поймы» . Статьи по палеонтологии (28). Анн-Арбор: Музей палеонтологии Мичиганского университета. hdl : 2027.42/48628 .
Дальнейшее чтение
[ редактировать ]- Макдональд, Дэвид (январь 1992 г.). Бархатный коготь: естественная история хищников . Книги Би-би-си . ISBN 978-0-563-20844-0 .