Деодон
| Деодон | |
|---|---|

| |
| Череп Динохиуса Холланди в Музее естественной истории Карнеги. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Парнокопытные |
| Семья: | † Энтелодонтиды |
| Род: | † Деодон Коуп , 1878 г. |
| Типовой вид | |
| † Деодон шошонский Коуп , 1878 г.
| |
| Разновидность | |
| |
| Синонимы | |


Daeodon — вымерший род энтелодонтных , парнокопытных населявший Северную Америку примерно от 29 до 15,97 миллионов лет назад в период позднего олигоцена и раннего миоцена . Типовой вид — Daeodon shoshonensis , описанный Коупом по очень сомнительному голотипу. Некоторые авторы отождествляют его с Dinohyus hollandi и рядом других видов (см. ниже), но из-за отсутствия диагностического материала это в лучшем случае сомнительно.
Другой крупный представитель этого семейства, более крупный, чем Daeodon , — азиатский Paraentelodon , но он известен по весьма неполному материалу. [ 1 ] [ 2 ]
Таксономия
[ редактировать ]Род Daeodon был выделен американским анатомом и палеонтологом Эдвардом Дринкером Коупом в 1878 году. Он отнес его к непарнокопытным и считал, что он тесно связан с Menodus . [ 3 ] классификация сохранялась до описания Elotherium Эта Calkinsi в 1905 году. [ 4 ] сразу же отнес к виду Dynohyus . очень похожее и гораздо более полное животное из тех же пород, которое Петерсон (1909) [ 5 ] Это привело к Деодона реклассификации как члена семейства Entelodontidae . Точные отношения между Деодоном и другими энтелодонтами недостаточно изучены; некоторые авторы (Lucas et al., 1998) рассматривают большее морфологическое сходство Daeodon с Paraentelodon , а не с более ранними североамериканскими энтелодонтами, такими как Archaeotherium , как свидетельство того, что Daeodon является потомком позднеолигоценовой иммиграции крупных азиатских энтелодонтов в Северную Америку. [ 6 ] Однако существование отдельных экземпляров Archaeotherium , демонстрирующих признаки, напоминающие те, что присутствуют как у Paraentelodon , так и у Daeodon, повышает вероятность того, что оба рода действительно произошли от общего североамериканского предка. [ 1 ] [ 7 ] Хотя это и не указано в первоначальном описании Коупа, имя Деодон происходит от греческих слов daios , что означает «враждебный» или «ужасный», и odon , что означает «зубы». [ 8 ]
Разновидность
[ редактировать ]Типовым видом Daeodon является D. shohonensis, который основан на фрагменте нижней челюсти из формации Джон Дэй в Орегоне . В последующие десятилетия к этому роду были отнесены еще несколько видов, например, D.calkinsi , D.mento. [ 9 ] и Д. минор . [ 10 ] С 1945 года было высказано предположение, что два других на самом деле являются младшими синонимами Daeodon таксона . [ 11 ] но официальное оформление этого направления не произошло до работы Лукаса и др. (1998). [ 6 ] Ammodon leidyanum , названный соперником Коупа О. К. Маршем и Динохиусом Холланди, [ 12 ] полный скелет из карьера Агат-Спрингс в Небраске , [ 13 ] оказались неотличимыми друг от друга и, в свою очередь, оба были неотличимы от D. shoshonensis . [ 6 ] За исключением D.calkinsi , который был предварительно исключен из состава Daeodon , другие ранее признанные виды Daeodon также были синонимичны D. shoshonensis . [ 6 ] В том же году малоизвестный энтелодонт, Boochoerus humerosum , был также синонимом Daeodon Фоссом и Фремдом (1998), и, хотя его статус отдельного вида был сохранен, они отмечают, что различия все же можно отнести к индивидуальным или популяционным вариациям или половой диморфизм. [ 14 ]
Описание
[ редактировать ]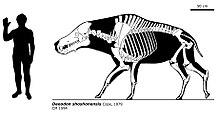

Daeodon shoshonensis — самый крупный из известных энтелодонтов ; [ 6 ] известные взрослые особи имели черепа около 90 см (3 фута) в длину и высоту в плечах около 1,77 м (5,8 фута). [ 5 ] Он отличается от других энтелодонтов набором уникальных особенностей зубов, формой и относительно небольшим размером скуловых выступов черепа по сравнению с таковыми у Archaeotherium , а также небольшим размером подбородочного бугорка , а также особенностями запястья и предплюсны . и срастание костей голени. [ 1 ] [ 5 ] [ 15 ] Как и у других энтелодонтов, его конечности были длинными и тонкими, кости передних ног срослись между собой. [ 6 ] [ 15 ] и только с двумя пальцами на каждой ноге. [ 5 ] [ 6 ] У него также была относительно легкая по размеру голова шея, вес которой в основном поддерживался мышцами и сухожилиями, прикрепленными к высоким отросткам грудных позвонков , как у современных бизонов и белых носорогов . [ 15 ]
Палеоэкология
[ редактировать ]среда обитания
[ редактировать ]Деодон имел широкий ареал в Северной Америке: многие окаменелости были найдены в агатовых ископаемых пластах , представляющих собой среду переходного периода между густыми лесами и обширными прериями, что, вероятно, стало основной причиной их исчезновения в раннем миоцене. [ 16 ] Он адаптировался к пастбищам с более беглым строением тела, чем у большинства базальных энтелодонтов, таких как Archaeotherium, полностью потеряв прибылые пальцы, проксимально сросшиеся пястные кости и мускулатуру плеч, аналогичную бизону. [ 15 ] [ 17 ]
Костное ложе Агат-Спрингс представляло собой пойму с влажными и засушливыми сезонами. Деодон делил этот ландшафт с маленькими газелеобразными верблюдами Stenomylus , большим халикотером Moropus размером от койота до волка , несколькими видами хищных амфиционид , которые жили стаями, наземными бобрами ( Paleocastor ), которые заполнили экологическую нишу современных луговых собачек, и тысячи мелких носорогов, живущих стадами. В засушливый сезон носороги периодически вымирали, но окаменелости даеодонов редки, что позволяет предположить, что они не были социальными животными и их не особенно привлекала падаль. [ 18 ]
Диета
[ редактировать ]Деодон был всеяден, как и все другие энтелодонты. Узоры на эмали предполагают употребление в пищу орехов, кореньев и виноградной лозы, а также мяса и костей. Внешнее сходство с пекариями, бегемотами и медведями предполагает широкий диапазон видов растений, деодон которыми питался . Засушливые сезоны в Северной Америке в то время могли быть очень суровыми, поэтому они, возможно, дополняли потребление воды, поедая виноградные лозы. Степень их плотоядности спорна, но износ зубов позволяет предположить, что они специализировались на дроблении костей и разрывании мяса, а следы укусов на костях халикотера позволяют предположить, что они либо охотились, либо добывали пищу на крупных травоядных животных. Фосс (2001) утверждает, что его голова была слишком тяжелой, чтобы эффективно уничтожать крупную добычу, поэтому он, должно быть, полагался исключительно на падальщиков, но его бизоньи приспособления к бегу, стереоскопическое зрение, характерное для хищников, и свидетельства хищничества энтелодонтов ставит эту интерпретацию под сомнение. [ 19 ] Неопределенность их рациона предполагает, что они, вероятно, были всеядными приспособленцами, похожими на медведей, которые ели все, что им нужно, в зависимости от обстоятельств. [ 20 ]
Поведение
[ редактировать ]Энтелодонты участвовали во внутривидовых кусаниях лица, о чем свидетельствуют следы зубов на их черепах. Самцы будут бороться за доминирование, возможно, используя бугорки нижней челюсти в качестве защиты в дополнение к их функции прикрепления мышц. [ 20 ] Половой диморфизм защиты скуловых костей существует у Archaeotherium, и при меньшем размере выборки Daeodon такой диморфизм нельзя исключить для Daeodon . В случае диморфизма функция расширенных скуловых костей, скорее всего, была демонстрационной, поддерживая большие предглазничные железы, подобные тем, которыми обладают лесные свиньи для химической коммуникации.
Ссылки
[ редактировать ]- ^ Jump up to: а б с Дональд Р. Протеро; Скотт. Э. Фосс (2007). Эволюция парнокопытных . Джу Пресс. ISBN 9780801887352 .
- ^ L. K. Gabunia (1964). Бернарская фауна олигоценовых позвоночных (The Benarskaya Fauna of Oligocene Vertebrates) . Metsniereba, Tbilisi. pp. 109–133 . Retrieved 2020-09-26 .
- ^ Коуп, ЭД (1878). «О некоторых признаках миоценовой фауны Орегона». Палеонтологический вестник . 30 : 1–16.
- ^ Синклер, WJ (1905). «Новые и недостаточно известные грызуны и копытные из серии Джона Дэя». Бык. Кафедра геологии, унив. Калифорния . 4 : 132–134.
- ^ Jump up to: а б с д Петерсон, ОА (1909). «Пересмотр энтелодонтид» . Мемуары музея Карнеги . 4 (3): 41–158. дои : 10.5962/стр.234831 . hdl : 2027/mdp.39015017493571 . S2CID 247000277 .
- ^ Jump up to: а б с д и ж г Лукас, СГ; Эмри, Р.Дж.; Фосс, SE (1998). «Таксономия и распространение Daeodon , энтелодонта олигоцен-миоцена (Mammalia: Artiodactyla) из Северной Америки» . Труды Биологического общества Вашингтона . 111 (2): 425–435.
- ^ Фосс, SE; Фремд, Ти Джей (2001). «Биостратиграфия Entelodontidae (Mammalia: Artiodactyla) из бассейна Джона Дэй, Орегон». ПалеоБиос . 21:53 .
- ^ Браун, RW (1954). Состав научных слов: Методическое пособие и словарь материалов для практики логотехники . Издательство Смитсоновского института. ISBN 978-0874740011 .
- ^ Аллен, генеральный директор (1926). «Ископаемые млекопитающие из Южной Каролины». Вестник Музея сравнительной зоологии . 67 : 447–467.
- ^ Лумис, ФБ (1932). «Два новых энтелодонта миоцена». Журнал маммологии . 13 (4): 358–362. дои : 10.2307/1374141 . JSTOR 1374141 .
- ^ Симпон, Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень Американского музея естественной истории . 85 : 1–350.
- ^ Петерсон, О.А. (1905b). «Исправление родового названия (Dinochoerus), данного некоторым ископаемым останкам из миоцена Луп-Форк в Небраске» . Наука . 22 (570): 719. Бибкод : 1905Sci....22..719P . дои : 10.1126/science.22.570.719 . ПМИД 17729479 .
- ^ Петерсон, О.А. (1905a). «Предварительная заметка о гигантском млекопитающем из пластов Лу-Форк в Небраске» . Наука . 22 (555): 211–212. Бибкод : 1905Sci....22..211P . дои : 10.1126/science.22.555.211 . ПМИД 17835750 .
- ^ Фосс, SE; Фремд, Т. (1998). «Обзор видов энтелодонтов (Mammalia, Artiodactyla) бассейна Джона Дэй, Орегон». Дакотерра . 5 : 63–72.
- ^ Jump up to: а б с д Эффингер, Дж. А. (1998). «Энтелодонтиды». В Янисе, CM; Скотт, КМ; Джейкобс, LL (ред.). Эволюция третичных млекопитающих Северной Америки. Том 1: Наземные хищники, копытные и копытные млекопитающие . Издательство Кембриджского университета. ISBN 9780521355193 .
- ^ Национального памятника «Окаменелости агата» Служба национальных парков . 2009 год
- ^ Эндрю Брант Клиффорд (май 2010 г.). Эволюция и механика унгулиградий парнокопытных (кандидатская диссертация). Университет Брауна .
- ^ «Окаменелости млекопитающих - Национальный памятник окаменелостей агата (Служба национальных парков США)» . www.nps.gov . Проверено 28 октября 2018 г.
- ^ «Реферат статей. Пятьдесят девятое ежегодное собрание Общества палеонтологии позвоночных» . Журнал палеонтологии позвоночных . 19 (3): А1–А93. 1999. ISSN 0272-4634 . JSTOR 4524027 .
- ^ Jump up to: а б Фосс, С.Е., 2001, Систематика и палеобиология энтелодонтид (Mammalia, Artiodactyla) [Ph.D. диссертация]: Декалб, Университет Северного Иллинойса.
- Энтелодонты
- Миоценовые парнокопытные
- Миоценовые млекопитающие Северной Америки
- Олигоценовые млекопитающие Северной Америки
- арикарийский
- Хемингфордианский
- Фауна Белой реки
- Вымирание рода Burdigalian
- Первые появления рода Chattian
- Ископаемые таксоны, описанные в 1878 г.
- Таксоны, названные Эдвардом Дринкером Коупом
- Род доисторических парнокопытных
- Олигоценовые парнокопытные