Энгидриодон
| Энгидриодон Временной диапазон: от позднего миоцена до раннего плейстоцена ,
| |
|---|---|
| |
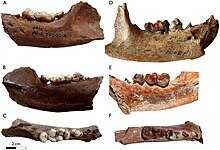
| Зубные ряды нижней челюсти Enhidriodon hendeyi (AC) и Enhidriodon africanus (DF) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Плотоядный |
| Семья: | Куньи |
| Подсемейство: | Лутрина |
| Племя: | † Энгидриодонтини |
| Род: | † Энгидриодон Фальконер , 1868 г. |
| Типовой вид | |
| † Энгидриодон сиваленский Фальконер, 1868 г.
| |
| Другие виды | |
| |
| Синонимы | |
|
Синонимия рода Синонимы E. sivalensis Синонимы E. ekecaman | |
Энгидриодон — вымерший род куньих , известный из Африки , Пакистана и Индии , живший с позднего миоцена до начала плейстоцена . Он содержит девять подтвержденных видов , два обсуждаемых вида и, по крайней мере, несколько других неописанных видов из Африки. Род принадлежит к трибе Enhidriodontini (в которую входят также Sivaonyx и Vishnuonyx выдр ) подсемейства Lutrinae . означает «зуб выдры» Энгидриодон в переводе с древнегреческого и является отсылкой к его зубному ряду, а не к роду Энгидра , который включает в себя современную калану и двух ее доисторических родственников.
Точные размеры и длина видов энгидриодонов неизвестны, учитывая отсутствие полных окаменелостей этого вида и большинства родственных ему ископаемых лютринов. По оценкам, индийские субконтинентальные виды имеют вес, аналогичный весу современных каланов , но африканские виды, по оценкам, тяжелее современных лютринов. В частности, несколько видов, таких как E. kamuhangirei , E. dikikae и E. omoensis , по оценкам, весили более 100 кг (220 фунтов). Учитывая эти оценки веса, эти три вида, вероятно, достигли размеров, сравнимых с нынешними медведями или львами , что делает их самыми крупными из известных куньих, хотя отсутствие полных экземпляров делает точные оценки невозможными.
Его развитый зубной ряд хорошо известен, а широкие бунодонты -хищники позволяют лютрину поедать добычу, раздавливая ее, а не стрижая, как современная калана, и в отличие от большинства других современных выдр. Таким образом, он сгруппирован среди бунодонтовых выдр - категорического термина, относящегося к ископаемым лютринам с нелезвийными хищниками в премолярах или коренных зубах от миоцена до плейстоцена, а также к каланам единственного сохранившегося рода Enhydra . Это я 3 зубы (или третьи резцы ) похожи на клыки и намного крупнее других резцов (хотя и короче клыков), черта, не встречающаяся у современных и вымерших родов лютринов. Предполагается, что индийские виды энгидриодонов вели полуводный образ жизни и питались двустворчатыми моллюсками , потому что их бунодонтные зубные ряды позволяли им поедать беспозвоночных с твердым панцирем. Неизвестно, были ли африканские виды водными, полуводными или наземными, но их потенциальная диета, подходящая для зубных рядов бунодонта, включает двустворчатых моллюсков, сомов , рептилий, яйца и падаль. E. omoensis из Эфиопии, в частности, мог быть наземным локомотором, который, по крайней мере, полурегулярно охотился или собирал наземную добычу с помощью растительной диеты C 4 , что, если это правда, делает его поведение непохожим на поведение любых современных выдр. Неизвестно, является ли этот вид выдающимся среди африканских видов бунодонтовых выдр, но было высказано предположение, что Enhydriodon dikikae и Sivaonyx beyi также были крупными наземными бунодонтными выдрами Африки.
Таксономический статус видов энгидриодонов осложнялся их сходством и сходством с другими родами бунодонтовых лютринов, такими как Sivaonyx и Paludolutra, вплоть до наших дней, хотя в настоящее время Paludolutra считается отдельным родом, не связанным тесно с энгидриодоном . В настоящее время племя Enhidriodontini считается эволюционно более близким к современному роду Enhydra , чем любой другой известный род бунодонтных выдр, которые, возможно, получили бунодонтный зубной ряд в результате параллельной эволюции , но степень их тесного родства остается нерешенной.
Таксономия
[ редактировать ]Ранняя история
[ редактировать ]
Энгидриодон был впервые создан в 1868 году Хью Фальконером на основе нескольких черепов с холмов Сивалик в Индии, которые он приписал E. sivalensis . [ 1 ] [ 2 ] Он пояснил, что научное название, означающее «зуб выдры», происходит от древнегреческих терминов ἐνυδρίς (выдра) и ὀδούς (зуб) и не является отсылкой к роду Enhydra , в который входит современная калан ( Enhydra lutris ). По словам Фальконера, окаменелости на холме Сивалик, принадлежащие E. sivalensis, ранее были классифицированы Фальконером и Проби Котли под названием рода и вида Amyxodon sivalensis в обзоре 1835 года ископаемых родов на холмах Сивалик, обнаруженных двумя палеонтологами, в котором ископаемый таксон считался хищником неизвестного семейства , хотя голотипа ему не было приписано ни , ни диагностических описаний. В результате переименования Amyxodon стал считаться « мертвым названием » или синонимом Enhydriodon, несмотря на то, что это старое название рода. Используя доступные экземпляры E. sivalensis было по четыре премоляра и коренных зуба , Фальконер подсчитал, что в верхней челюсти этого вида ( зубная формула была рассчитана как 3.1.4 2.1.5 ), что на один меньше, чем у современного рода. Лутра, но соответствует общему количеству Энгидры . Он описал верхнюю хищную особь E. sivalensis как наиболее уникальную особенность ее верхней челюсти: она почти квадратная, а корональные лопасти развиваются из конических мамелонов, в отличие от двух современных родов lutrine. [ 1 ] [ 3 ]
В течение 19 и 20 веков больше видов энгидриодонов, таких как E. Campanii было интродуцировано , и было описано больше родов лютринов с бунодонтом , таких как Sivaonyx и Vishnuonyx , что создало особенно сложную историю для самого раннего описанного доисторического рода выдр. В 1931 году Гай Пилигрим описал еще больше окаменелостей, обнаруженных на холмах Сивалик, включая новый вид под названием E. falconeri . Он также предположил, что энгидриодон и сиваоникс , несмотря на их сходство, различаются по строению 4-го премоляра верхней челюсти (P 4 ) и явное отсутствие переднего верхнего премоляра (P 1 ), что, как предполагается, отражается и на нижней челюсти (оба вопроса обсуждаются до сих пор). [ 4 ] В том же году, когда E. falconeri был описан , Эрнст Стромер описал E. africanus позднего плиоцена , его ископаемые зубы находились в Южной Африке , и это был первый описанный вид с африканского континента. [ 5 ]
Предполагаемые отношения с Палудолутрой и Энгидрой
[ редактировать ]В 1976 году Чарльз Репеннинг выдвинул идею о том, что энгидриодон связан с существующим родом энгидра, поскольку предполагаемый вид первого был эволюционной «ветвью» «выдр, питающихся крабоедами» в Италии, Испании и Калифорнии, что в конечном итоге привело к появлению современная калан. [ 6 ] Он правильно выдвинул идею о том, что Энгидра была родственна Энгидриодону , учитывая их бунодонтные зубные ряды, но предполагаемая европейская «ветвь» рода Энгидриодон была позже реклассифицирована Йоханнесом Хюрцелером и Буркартом Энгессером в новый род Палудолутра в 1976 году, хотя в 1976 году она оставалась относительно неясной. палеонтологические данные, пока более поздние исследования не пересмотрели его таксономическое состояние. [ 7 ] [ 8 ]
Таксономия отдельных видов и родов лютринов продолжала пересматриваться в 21 веке по мере того, как было описано все больше доисторических видов выдр, в то время как палеонтологи постоянно пересматривали ископаемые виды лютринов бунодонта, разделяя их на разные роды. Первоначально палудолутра была реклассифицирована подрод энгидриодона . как Джерардом Ф. Виллемсеном в 1992 году [ 5 ] Однако в январе 2005 года Мартин Пикфорд и его коллеги диагностировали Палудолутру как синоним Сиваоникса на основании диагноза Пилигрима последнего, отвергая синонимию Палудолутры и Энгидриодона Виллемсена . Кроме того, они выделили вид энгидриодона под названием E. hendeyi из типового местонахождения Лангебаанвег в Южной Африке, который датируется нижним плиоценом и был назван в честь палеонтолога Куинтона Б. Хенди, который, по их словам, описал первые известные экземпляры, которые были с тех пор отнесены к виду. [ 9 ] В декабре того же года Хорхе Моралес и Пикфорд вместо этого описали Paludolutra как отдельный род, который может быть связан с Sivaonyx на основании сближения зубных рядов. [ 8 ] В 2007 году два палеонтолога подтвердили, что морфология зубов Paludolutra достаточно различна, чтобы ее можно было реклассифицировать в род на основе полной родовой дифференциации, предполагая, что виды P. Campanii , P. lluecai и P. maremmana больше не будут классифицироваться как Энгидриодон в основе Paludolutra . подрода [ 10 ] [ 11 ]
Современные версии африканских видов
[ редактировать ]
В 2003 году Ларс Верделин выделил вид E. ekecaman из палеонтологического памятника Канапои в бассейне Туркана в Кении (ранний плиоцен, около 5,2–4,0 млн лет назад), описав его как одного из самых ранних представителей африканской линии энгидриодонов . Вид был назван в честь термина «экекаман» на языке туркана , что означает «рыбак», поскольку он предположил, что рыба могла быть пищей для этого вида. Он также объявил вид « E. pattersoni », описанный Р. Дж. Сэвиджем в 1978 году, nomen nudum E. ekecaman , поскольку ему не было присвоено ни типового образца, ни действительного диагноза. Эту точку зрения поддержали Моралес и Пикфорд в декабре 2005 года. [ 12 ] [ 8 ]
E. africanus , E. ekecaman и E. hendeyi были реклассифицированы в Sivaonyx Пикфордом и Моралесом в декабре 2005 года, где они дополнительно описали новый вид под названием Sivaonyx kamuhangirei . [ 8 ] Реклассификация африканских ископаемых выдр-бунодонтов в сиваоникс вызвала постоянные дебаты относительно практичности различий между энгидриодоном и сиваониксом , при этом некоторые исследователи заявляли о нейтральности из-за того, что предпочитали концентрироваться на изучении отдельных видов, а не на их родовом расположении. В 2022 году эти четыре вида были в конечном итоге реклассифицированы в энгидриодон в исследовательской работе Камиллы Гроэ и др. E. soriae также первоначально был отнесен к Sivaonyx, но в конечном итоге был отнесен к Enhydriodon , хотя его родовое отнесение остается спорным. [ 13 ] [ 14 ] В 2005 году Моралес и Пикфорд разделили Энгидриодона на недавно созданное племя Энгидриодонтини , которое они описали как принимающее роды вымерших выдр-бунодонтов с холмов Сивалик и Африки, включая Вишнуоникс , Сиваоникс и Палудолутра . В 2007 году Пикфорд сделал синонимом вид « E. aethiopicus », ранее описанный Денисом Гераадсом и соавт. в 2004 году — к Pseudocivetta ingens , вымершему члену семейства Viverridae . [ 10 ] В 2017 году Энгидра была явно исключена из трибы Энгидриодонтини, несмотря на ее сходство, а Палудолутра была реклассифицирована как родственный таксон этого племени. [ 8 ] [ 11 ]
В 2011 году Денис Гераадс и его коллеги описали E. dikikae на основе останков частичного черепа и бедренных костей в Нижнем Аваше Дикики , Эфиопия, местности, датируемой средним плиоценом. Было описано, что у него был значительно более тяжелый череп (хотя и сломанный), чем у других видов энгидриодонов или современных каланов. Названный вид базировался непосредственно на стоянке Дикика. [ 15 ] Он считался самым крупным видом энгидриодона , пока в 2022 году в долине Нижнего Омо не был описан другой вид, также из Эфиопии, E. omoensis , датируемый периодом от позднего плиоцена до границы плио- плейстоцена . Как и в случае с E. dikikae , название вида произошло непосредственно от места, где он был обнаружен. [ 14 ] Однако на конференции Альберто Валенсиано, Моралеса и Пикфорда в сентябре 2022 года (в том же месяце, что и исследовательская работа по E. omoensis ) они упомянули некоторые виды лютринов, ранее реклассифицированные в Enhydriodon , как Sivaonyx , а именно S. hendeyi и S. africana. . [ 16 ]
Классификация
[ редактировать ]
Энгидриодон принадлежит к трибе Enhidriodontini подсемейства Lutrinae , впервые появившейся в Евразии и Африке в эпоху позднего миоцена . [ 8 ] Это, пожалуй, самая известная доисторическая выдра, учитывая ее давнюю таксономическую историю, и она является основным источником сравнений с другими родами бунодонтовых выдр. Обычно считается, что энгидриодон возник в результате миоцен-плейстоценовой тенденции, которая дала доисторическим лютринам бунодонт зубы и большие размеры по сравнению с их нынешними родственниками. Он классифицируется как член группы бунодонтовых выдр, категорический термин, обычно используемый исследователями, который также включает в себя сиваоникс , палудолутру , вишнуоникс , торолутру , энгидритерий , джурабус , паралутру , тирренолутру , сиамогале и энгидру . [ 11 ] [ 17 ] [ 18 ] Бунодонтские выдры определяются как большие и очень крупные куньи Северной Америки, Евразии и Африки, у которых был крепкий зубной ряд по сравнению с большинством современных выдр, что обычно позволяло им охотиться на существ с твердой броней. [ а ] [ 13 ] [ 17 ] Несмотря на общие черты бунодонтного зубного ряда, существует по крайней мере несколько клад лютринов, принадлежащих к этой категории, а не одна, поэтому термин «бунодонтные выдры», следовательно, является категориальным, а не таксономическим, и охватывает лютрины в одни и те же периоды со схожим зубным рядом, а не с одним. что непосредственно определяет их таксономическое состояние. [ 11 ]
Следующая кладограмма Сяомин Вана и др. в 2018 году определены некоторые из следующих существующих и вымерших видов и родов выдр в подсемействе Lutrinae на основе консенсуса 50% большинства (роды бунодонтовых выдр выделены жирным шрифтом, начиная с « Paralutra jaegeri »): [ 11 ]
| Лутрина |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Как показано в приведенной выше филогении, энгидриодон имел более близкую морфологию со своими другими вымершими родственниками и энгидрой, чем другие существующие лютрины, у которых отсутствуют плотоядные зубы бунодонта ( Lutra aonychoides была описана как не связанная с Lutra ). Хотя консенсусное дерево большинства демонстрирует тесную морфологическую связь между энгидриодоном и энгидрой , авторы консенсусного дерева также создали байесовское дерево вывода, предполагающее, что энгидра как изолированная клада отделена от типичных представителей энгидриодонтини (« Паралютра » jaegeri была предложена как изолированная группа). клада из Сиамогале ). Тем не менее, они утверждали, что Энгидра ближе к кладе, состоящей из Энгидриодона , Сиваоникса и Вишнуоникса , чем любой другой род бунодонтовых выдр. Исследователи объяснили, что приобретение бунодонтного зубного ряда произошло как минимум трижды в эволюции лютринов, что отражено кладами филогенного дерева: у сиваоникса — энгидриодона — энгидры , у палудолутры — энгидритерия и у сиамогале. . [ 11 ] Не-бунодонтовые выдры, вероятно, развились отдельно от бунодонтовых выдр во время или до эпохи плиоцена, но их скудные летописи окаменелостей и ограниченность отложениями плио-плейстоцена по сравнению с ними оставляют мало понимания их эволюционной филогении. [ 20 ]
Описание
[ редактировать ]Череп
[ редактировать ]
В настоящее время известны только два части черепа, приписываемые Энгидриодону : один — E. sivalensis с холмов Сивалик, а другой — E. dikikae из долины Аваш. В настоящее время неизвестно, являются ли черты черепов обоих видов хорошо репрезентативными для других видов энгидриодонов , но известные черепа E. dikikae и E. sivalensis имеют несколько отличающиеся особенности друг от друга. [ 15 ]
Череп E. sivalensis , идентифицированный как принадлежащий взрослой особи, относительно хорошо сохранился с идентифицируемыми височными гребнями , лобными , верхнечелюстными , предчелюстными , носовыми , мордовыми и костей небными частями . Тем не менее, он также пострадал от износа и был слегка перекручен по часовой стрелке. В частности, зубная дуга цела, хотя левая M 1 и оставил я 1 оба отсутствуют, и большинство зубов отломаны от коронок. У него большая черепная коробка, широкая и короткая морда и большое носовое отверстие. Также можно идентифицировать очертания орбит вокруг лобных частей черепа. [ 2 ]
Сломанный череп, принадлежащий E. dikikae, содержит короткую непрогнатическую морду, части глазниц, почти полную верхнюю зубную дугу, в которой отсутствуют оба I 1 s и право я 2 и часть нижней челюсти. Морда черепа E. dikikae короткая, с небольшой передней границей глазницы, расположенной чуть выше задней стороны клыка . Передняя часть морды определяется как короткая, поэтому ее можно сравнить с мордой Энгидры . Хотя эволюция бунодонтных выдр, таких как Enhydriodon, неясна, предполагается, что короткая морда E. dikikae и очень большой размер клыков явно делают этот вид другим и более эволюционно произошедшим (или эволюционно недавним), чем E. sivalensis . [ 15 ]
Зубной ряд
[ редактировать ]Зубной ряд энгидриодона хорошо выражен благодаря чрезвычайно широким бунодонтным хищным зубам на коренных и премолярах, похожим на зубы современной калана. Видовые различия энгидриодона . и сиваоникса обычно объясняются зубным рядом, поэтому окаменелости премоляров или коренных зубов исследуются, чтобы различить два рода выдр-бунодонтов Общие различия (большие P 4 гипоконус, конические бугры постпротоконуса и явное отсутствие передних верхних премоляров у энгидриодона ) с помощью измерений зубов было трудно доказать из-за фрагментарного характера окаменелостей и относительной несогласованности измерений/размеров зубов по видам. [ 10 ] [ 15 ] Реклассификация всех видов «африканского сиваоникса », кроме S. beyi, в энгидриодон в 2022 году была связана с «[а] метаконидом, более высоким, чем протоконид на M 1 , наличием хищной выемки и одного или нескольких выступов между протоконидом и протоконидом. гипоконус на P 4 и/или дистолингвальное расширение M 1 ." [ 14 ]
Предполагается, что Энгидриодон, как последний появившийся род, имеет самый бунодонтный зубной ряд среди трибы Энгидриодонтини, которая включает самого раннего появившегося Вишнуоникса , а затем Сиваоникса . Зубной ряд энгидриодона предполагает почти полное подавление хищнических функций в пользу дробления как преобладающей функции. Я 3 (или третий верхний резец ) энгидриодона значительно крупнее его I 1 (самый маленький резец) и я 2 , выглядя крупнее и более похожим на собаку по сравнению с Палудолутрой и Энгидрой . По сравнению с другими родами бунодонта lutrine, у которых известен верхний резец, его третьи резцы лишь незначительно больше, чем их первый и второй резцы. [ 10 ] Прав я 1 Например, череп E. sivalensis имеет переднезадний диаметр (APD) 3 мм (0,12 дюйма) и поперечный диаметр (TD) 4,5 мм (0,18 дюйма). Череп прав я 2 имеет размеры 5,2 мм (0,20 дюйма) в APD и 5,5 мм (0,22 дюйма) в TD. Для сравнения, прав я 3 это самый крупный резец голотипа, его размеры составляют 10,5 мм (0,41 дюйма) при APD и 8 мм (0,31 дюйма) при TD (клыки крупнее резцов, их размеры 17,1 мм (0,67 дюйма) при APD и 13,8 мм ( 0,54 дюйма в диаметре). [ 2 ] Большой я 3 Этот признак также применим к E. dikikae , который был описан после общего описания Пикфордом рода Enhydriodon как имеющего гораздо больший I. 3 чем я 1 - Я 2 и иметь более коническую форму. ДИК-56 И 3 Размер зуба составляет 12,4 мм (0,49 дюйма) по мезиодистальной ширине (MD) и 11,6 мм (0,46 дюйма) по щечно-язычной ширине (BL) по сравнению с его I 2 размеры 5,5 мм (0,22 дюйма) в MD и 9,7 мм (0,38 дюйма) в BL. Как и E. sivalensis , I 3 короче клыков, с C 1 размером 16,9 мм (0,67 дюйма) в продольном направлении плюс 15 мм (0,59 дюйма) в BL и C 1 размером 19,5 мм (0,77 дюйма) в продольном направлении и 15,3 мм (0,60 дюйма) в BL. [ 15 ]
Конечности
[ редактировать ]
Посткраниальные остатки выдр-бунодонтов, включая энгидриодонов , немногочисленны, что оставляет слишком мало информации об общей анатомии многих родов. Единственные известные виды энгидриодонов с посткраниальными остатками — E. hendeyi , E. dikikae и E. omoensis . [ 13 ]
Ископаемые остатки E. hendeyi включают фрагментарную плечевую кость , локтевую кость , две бедренные кости и таранную кость (также известную как таранная кость). Бедренные кости E. hendeyi меньше, чем у S. beyi и E. dikikae , но также крупнее, чем у Enydritherium , Satherium и современной африканской бескогтевой выдры. Таранная кость похожа на E. omoensis, но отличается меньшей головкой и более тонкой шейкой, а также более крупным дистальным выступом костного бугорка . Трохлея таранной кости (рифленая поверхность, образующая суставы костей) неглубокая и широкая в медиально-латеральном направлении, а ее бугорок выступает примерно в центре, оба из которых образуют прочную и глубокую бороздку (или борозду) сухожилий мышц сгибания для подошвенного расширение стопы в лодыжке по сравнению с африканской бескогтевой выдрой. [ 13 ]
Из посткраниальных остатков E. dikikae известны проксимальная (верхняя часть) левого бедра, дистальная (нижняя часть) правого бедра и плечевая кость. Проксимальный отдел левого бедра известен по большому бугорку вдоль задней области шейки, расположенному посередине малому вертелу и аналогичному положению медиального мыщелка бедренной кости . Образцы бедренных костей указывают на большие общие размеры бедренных костей E. dikikae по сравнению даже с самыми крупными современными выдрами. Плечевая кость (целая, но состаренная и потрескавшаяся) намного длиннее и немного крепче по сравнению с плечевой костью Sivaonyx beyi , а ее дельтовидная бугристость хорошо сформирована. Латеральный надмыщелковый гребень длиннее, чем у S. beyi , тогда как медиальный надмыщелок не так заметен по размеру. Олекранонная ямка маленькая и более круглая по сравнению с S. beyi . [ 15 ]
E. omoensis представлен лишь единственным целым левым бедром, имеющим короткую шейку и округлую головку, ориентированную не в медиальном направлении (в центре), а в проксимальном направлении (близко к центру), причем первая 40° относительно долготы диафизарного отдела кости. Латеромедиальная ширина эпифиза узкая . Бедренная кость также имеет большую головку бедренной кости , расположенную на вентромедиальной головке (совпадая со средней нижней ее стороной), большой вертел , загибающийся назад и расположенный ниже головки бедренной кости, короткую и глубокую вертельную ямку и мощную меньшую кость. вертел, который сосредоточен больше посередине, чем на вентральной (или нижней стороне), и поэтому виден на виде сзади. Медиальный мыщелок бедренной кости больше латерального мыщелка той же кости. Межмыщелковая ямка бедренной кости прямоугольная, широкая. [ 14 ]
Масса тела
[ редактировать ]
Некоторые виды энгидриодонов , особенно те, которые обитали в Африке, являются крупнейшими из когда-либо существовавших куньих , если судить по оценкам веса, но их точные размеры и вес остаются неизвестными, учитывая отсутствие полных экземпляров в их летописях окаменелостей. Некоторые виды, такие как E. latipes(?), плохо изучены по сравнению с другими, и поэтому отсутствуют подтвержденные оценки размера и веса. [ 21 ] Обычно считается, что некоторые виды энгидриодонов по весу аналогичны современным крупным выдрам, в то время как другие считаются намного крупнее их (следует также отметить, что оценки веса чаще делаются для бунодонтных выдр, таких как энгидриодон, чем оценки размеров, хотя можно привести сравнение размеров с современными животными). [ 14 ]
Два вида энгидриодонов , обитающих на субконтиненте Индии, имели скромные оценки веса, сравнимые с большинством других родов бунодонтовых выдр, а также с существующими родами выдр. В мемуарах Фальконера 1868 года E. sivalensis описывался как туалет размером с пантеру . [ 1 ] В 1932 году Пилигрим диагностировал E. falconeri как меньший по размеру, чем E. sivalensis , хотя никаких оценок размера или веса он не предложил. [ 4 ] В 2007 году Пикфорд оценил E. sivalensis как самую крупную доисторическую выдру в Индии: масса тела варьировалась от 22 кг (49 фунтов) минимум до 25 кг (55 фунтов) максимум, а ее череп, возможно, был размером с волка. Он также оценил, что тело E. falconeri , исходя из размеров нижних зубов M 1 , похоже на тело африканской бескогтевой выдры ( A. capensis ), его средний вес составляет 16 кг (35 фунтов). [ 10 ]
По оценкам, африканские виды энгидриодонов являются одними из крупнейших видов выдр, когда-либо существовавших, что отражает тенденцию миоцена-плейстоцена, когда бунодонтовые выдры становятся крупнее, чем их собратья, не относящиеся к бунодонтам. Пикфорд описал E. kamuhangirei из Западной рифтовой долины в Уганде (в то время Sivaonyx kamuhangirei ), вес которой, возможно, превышал 100 кг (220 фунтов), что делало ее самой крупной известной доисторической выдрой того времени, хотя он упомянул, что неописанная окаменелость выдры в Эфиопии (вероятно, позже отнесенные к E. dikikae и/или E. omoensis ), возможно, были крупнее его. [ 10 ] E. hendeyi (тогда Sivaonyx hendeyi ), по оценкам, был размером с волка и весил около 40 кг (88 фунтов), в то время как E. africanus и E. ekecaman , как полагают, имели одинаковые размеры. [ 22 ] [ 14 ] По оценкам, E. dikikae из Эфиопии весил минимум 100 кг (220 фунтов) и максимум 200 кг (440 фунтов) (последнее упоминается как более вероятное), его голотип предполагает размер медведя. По сравнению с большинством других видов энгидриодонов или энгидр , его предполагаемая длина черепа составляла около 25 см (9,8 дюйма). [ 15 ] Позже было подсчитано, что E. omoensis весит более 200 кг (440 фунтов), что делает его тяжелее, чем E. dikikae и современные львы . По мнению Гроэ и др., E. omoensis потенциально был «размером со львом», что делало его самым крупным видом куньих, когда-либо существовавшим. [ 14 ]
Палеобиология
[ редактировать ]
Поскольку у ископаемых родов бунодонтовых выдр, включая Enhydriodon, обычно отсутствуют полные экземпляры и посткраниальные элементы, их передвижение и экологические ниши остаются неопределенными. Распространенная теория о индийских субконтинентальных видах энгидриодонов состоит в том, что из-за их крепких бунодонтных зубных рядов, подобных энгидре , E. falconeri и E. sivalensis специализировались на обычном поедании моллюсков . [ 5 ] Это утверждение было впервые сделано Виллемсеном на основе аналогий с рационом энгидры (уши и морские двустворчатые рыбы) и аоникса (пресноводные крабы), но палеонтологических доказательств, подтверждающих это утверждение, мало. Несмотря на это, предполагается, что толстая эмаль в заднем прикусе индийских видов энгидриодонов делает их более моллюскоядными, чем ракоядными (напротив, предполагается, что индийские виды Sivaonyx сочетают режущие функции хищных с общими коронками бунодонтов, чтобы больше охотиться на ракообразных ). хотя двустворчатые моллюски потенциально могли быть для него вторичной добычей). Возможность охоты энгидриодонов на двустворчатых моллюсков подтверждается наличием окаменелых пресноводных двустворчатых моллюсков родов Parreysia и Lamellidens в тех же местах, что и они, оба из которых распространены по всей осадочной колонке Сивалик, охватывающей период от 15 до 2 млн лет назад, в зависимости от присутствия племени энгидриодонтини на Индийском субконтиненте (Индия и Пакистан). [ 10 ]
Предполагается, что более крупные виды энгидриодонов на африканском континенте охотились на более широкий спектр продуктов питания в дополнение к своей основной добыче, включая более мягкую добычу, несмотря на их бунодонтные зубные ряды, что делает их потенциальный рацион отличным от рациона их индийских субконтинентальных собратьев. Одним из предполагаемых типов добычи была крупная рыба с твердым внешним покровом, например, сом . [ 23 ] Несколько родов сомов присутствовали в Африке, начиная с их первого появления в позднем миоцене, совпавшего с присутствием энгидриодона , включая современные роды Clarotes , Bagrus , Auchenoglanis и Chrysichthys и вымерший род Nkondobagrus . [ 24 ] В отличие от медлительных и многочисленных сомов, крабы в Африке были исключены как потенциальная добыча африканских видов энгидриодонов из-за отсутствия окаменевших крабов в Дикике, маловероятности биомассы крабов для поддержания популяций крупных выдр и очевидной несовместимости по эмали. зубной ряд. Маловероятно, что быстро плавающая рыба была постоянным источником пищи из-за специального зубного ряда, предназначенного для измельчения твердой пищи, а также из-за того, что крупные животные, вероятно, не имели возможности ловить быструю добычу. Другие панцирные жертвы, такие как молодые крокодилы, черепахи и страусиные яйца, также считались добычей E. dikikae . [ 15 ]

Бедра и останки зубов африканского энгидриодона, возможно, могут указывать на полуводный, а также наземный образ жизни, а это означает, что он мог питаться как водной, так и наземной добычей. Однако предположения об образе жизни энгидриодона противоречили друг другу, поэтому консенсуса большинства по этому поводу не существует. В 2008 году было высказано предположение, что более мелкие африканские виды энгидриодонов , исходя из их меньших размеров бедренных костей, были более универсальными локомоторными, похожими на большинство куньих, в то время как более крупные виды были полностью водными, поскольку их структуры бедренных костей имели сходство с энгидрой . Однако проксимальные концы бедренных костей Омо и Хадара указывали на более водную природу, чем у большинства лютрин, а их относительная длина напоминала длину наземных куньих-универсалов, включая полуводных выдр. [ 23 ] [ 14 ] Sivaonyx beyi из Чада , предположительно весивший от 56,4 кг (124 фунта) до 60,1 кг (132 фунта), имел неспециализированные пропорции конечностей, что подразумевало универсальное наземное передвижение и плохую водную адаптацию. [ 25 ] Из-за гипотезы о том, что S. beyi был наземным хищником, предполагается, что E. dikikae был в основном наземным, исходя из того, что его окаменелости были общими как с водной, так и с наземной фауной Дикики. [ 15 ] Палеобиология и разделение ниш E. ekecaman и E. cf. dikikae в Канапои , Кения, остаются неясными, поскольку их ископаемые материалы, обнаруженные в 1960-х годах, не были конкретно выражены за пределами «Канапои», которые должны были охватить будущие исследования. [ 26 ] Также отмечается, что африканские виды бунодонтовых выдр, такие как Enhydriodon и Sivaonyx, всегда обнаруживались в местах, связанных с постоянными водоемами, в отличие от слоев верхнего Laetolil в Лаэтоли , Танзания, где такая особенность отсутствовала, что ставит вопрос о том, насколько возможного наземного образа жизни африканских видов энгидриодонов и сиваониксов . [ 27 ]
E. hendeyi был проанализирован на основе индекса устойчивости бедренной кости (FRI) и надмыщелкового индекса бедренной кости (FEI), в которых его значение FRI сопоставимо с вымершими S. beyi , Enhidritherium и Satherium (последние два являются аналогами большого калан и гигантская речная выдра ( Pteronura brasiliensis ) соответственно и имеют большие значения бедренных индексов, чем у большинства других современных выдр), в то время как ее значение FEI аналогично современной африканской бескогтевой выдре и азиатской малокогтевой выдре ( Aonyx cinereus ). Поскольку и африканская бескогтистая выдра, и азиатская мелкокогтистая выдра, как правило, меньше связаны с водоемами по сравнению с другими современными выдрами, предполагается, что E. hendeyi и S. beyi были полуводными локомоторами, которые имели меньшую связь с водой, чем водные локомоторы Enguidritherium. и Satherium , хотя S. beyi считался более наземным, чем E. hendeyi . При этом самые низкие значения соответствуют E. dikikae , имеющему аналогичные значения с наземными полуфоссориальными. (адаптированных к рытью и жизни под землей) куньих, таких как американский барсук и полосатый скунс , тем самым подкрепляя гипотезу о том, что E. dikikae была более обобщенной наземной куньей, похожей на S. beyi . [ 13 ]
Учитывая общее отсутствие единого мнения относительно образа жизни африканских видов энгидриодонов , в исследовании 2022 года, посвященном E. omoensis, были измерены углерода и стабильные соотношения изотопов кислорода у видов энгидриодонов по сравнению с современными наземными млекопитающими, такими как кошачьи , гиениды и быки , а также полуводные виды. млекопитающие, такие как гиппопотамиды . Авторы объяснили, что использование изотопных отношений кислорода, или δ18O , можно использовать для понимания зависимости таксона от воды, при этом существующие водные и полуводные таксоны, включая речных и каланов, имеют значительно более низкие изотопные отклонения кислорода по сравнению с наземными хищниками. Исследователи, изучавшие E. omoensis, обнаружили, что значения δ18O его зубной эмали имели стандартное отклонение 2,7%, выходя за пределы стандартных отклонений δ18O калана и североамериканской речной выдры ( Lontra canadensis ), которые были зарегистрированы как 0,6. % и 0,3%-0,9% соответственно. Стандартное отклонение Омо Энгидриодона больше соответствует диапазону современных наземных хищников, таких как гиениды, что позволяет предположить, что E. omoensis не был таким полуводным, как первоначально предполагалось. Результаты исследования противоречат предположению 2008 года о том, что вид Омо Энгидриодон был водным. [ 14 ]
Гроэ и др. Первоначально считалось, что пищей энгидриодона могла быть устрица Etheria elliptica , которая присутствовала на континенте в тот же период времени. Согласно исследованиям с использованием стабильных изотопов углерода, рацион, состоящий из чистых устриц, приведет к значению δ13C эмали, равному -11,3%. рацион E. omoensis Однако не был основан исключительно на эфирии , поскольку его минимальные и максимальные значения углерода (от -9,7% до -4,7%) примерно на 2-7% более положительны, чем ожидаемая ценность диеты из чистых устриц. Значения δ13C его эмали попадают в диапазон смешанных питающихся C 3 - C 4 и лишь частично попадают в диапазон рационов водных растений, питающихся C 3 , таких как рыбы, черепахи или двустворчатые моллюски. Однако стандартное отклонение δ13C Omo Enhyriodon выходит за пределы изученных существующих популяций пресноводных выдр. Вместо этого считается, что E. omoensis потреблял наземную добычу с диетой C 4 , по крайней мере, полурегулярно, охотясь и/или собирая мусор. Большой бунодонтный зубной ряд этого вида предполагает дурофаговые способности, которые позволяли ему питаться падалью, включая кости, потенциально аналогично гиенам или куньим, сокрушающим кости. [ 14 ]
Палеоэкология
[ редактировать ]Пакистан и Индия
[ редактировать ]
E. falconeri и E. sivalensis , хотя оба вида Enhydriodon , которые присутствовали на холмах Сивалик в Индии и Пакистане в неогеновый период, не сосуществовали в одни и те же эпохи, судя по внешнему виду их отложений. Останки E. falconeri присутствовали в формации Нагри ( Дхок-Милан и Сетхи-Нагри , Пакистан ) и формации Дхок-Патан (Дхок-Патан и Хаснот , Пакистан), причем обе формации относятся к средним сиваликам, представляющим поздний миоцен. Вид также присутствовал в формации Татрот ( Татрот , Индия) Верхнего Сивалика из раннего или среднего плиоцена . Было показано, что в формациях Нагри и Дхок-Патан E. falconeri существовала вместе с несколькими архаичными семействами плотоядных млекопитающих, вымершими до плиоцена, такими как гиаиналоуриновые гиенодонты и амфициониды . Ранние виды выдр также существовали вместе с различными вымершими хищными представителями современных семейств в позднем миоцене, представляя других куньих , медведей, кошачьих ( кошачьих и кошачьих). mahairodontines ), гиениды ( percrocutinae hyaenids ictitheres и hyaenines), виверровые и герпестиды . Предполагается, что вымирание амфиционид и перкрокутид оставило пустые хищные ниши, которые быстро были заполнены другими родами гиенид, которые стали весьма разнообразными и сосуществовали с кошачьими на субконтиненте. [ 28 ]
Другие вымершие представители современных и вымерших семейств млекопитающих были обнаружены в формации Нагри и, таким образом, существовали вместе с E. falconeri, включая быков, жирафов , антракотериев , трагулид , свиней , гиппариониновых непарнокопытных , носорогов , халикотеридов , гомфотериев , гоминидов и спалацидов . [ 29 ] Вымерший вид рептилий гавиала , Gavialis lewisi (?), обнаружен из формации Дхок Патан в Пакистане и имеет плиоценовый возраст. [ 30 ] Роды млекопитающих, обнаруженные в формации Дхок Патан, в целом соответствуют родам млекопитающих, обнаруженных в формации Нагри, но также включают других быков, жирафов, оленьих оленей , антракотериев, суидов, гиппарионинов, носорогов, «тетралофодонтных гомфотериев», церкопитецидов и гистрицид. . [ 31 ] [ 32 ] [ 33 ] [ 34 ]

Переход от среднего миоцена к позднему миоцену отражал период, когда вечнозеленые и лиственные тропические леса, когда-то покрывавшие большую часть Индийского субконтинента, сократились и были заменены лугами из-за глобального похолодания, более засушливых условий и усиления азиатских муссонов. . [ 35 ] Переход от пойм Нагри к поймам Дхок Патана предполагает меньший сток в речной системе последнего по сравнению с первым, поскольку более мелкие реки Дхок Патана имеют больший сезонный сток, чем раньше. Это отражает общую тенденцию воздействия на климат позднего миоцена , приводящего к большей сезонности, вызывающей большие обороты фауны. Более сухой и сезонный климат наряду с речными изменениями постепенно привели к появлению более крупных открытых лесных массивов , преимущественно состоящих из растений C 4 вблизи миоценовых рек Потвара , в то время как сообщества, состоящие исключительно или преимущественно из растений C 3, значительно сократились и в конечном итоге исчезли на 7,0 млн лет назад вместе с C 3 . 3 кормушки, которые больше зависели от закрытой растительности. Эти изменения произошли вскоре после прибытия гиппарионинов и заметного сокращения групп млекопитающих на Индийском субконтиненте, таких как исчезновение гоминида Sivapithecus и дейнотерия Deinotherium в результате фрагментации закрытых местообитаний в пользу открытых местообитаний, которые лишили бы пищи. для С 3 браузера и плодоядных. [ 36 ] [ 37 ] [ 38 ]

Записи окаменелостей хищных животных формации Татрот в Индии скудны, но среди вымерших представителей, существовавших вместе с E. falconeri в плиоцене, были и другие лютрины, махайродонтины и гиениды. [ 28 ] Травоядные млекопитающие, обнаруженные в формации Татрот на плато Потвар, содержат очень разнообразные сообщества быков, но также включают оленьих, суидов, , слонов стегодонтид , гиппарионинов, антракотериев, бегемотов, жирафов и трагулид. [ 40 ] [ 41 ] [ 42 ] Крокодилы Crocodylus и Rhamphosurus , пеликан Pelecanus , черепахи ( Batagur , Geoclemys , Hardella и Pangshura ) и пресноводный краб Acanthopotamon также встречаются, по крайней мере, в формациях Татрот или Пинджор в Индии, что указывает на активную пресноводную среду обитания, которую E. falconeri , а позднее E. sivalensis присутствовали в. [ 30 ] [ 43 ] [ 44 ] [ 45 ]
Среди таксонов хищных животных Enhydriodon - самый долгоживущий род псовых , когда-либо существовавший среди сиваликцев Индийского субконтинента, идентифицированный из формаций Нагри-Пинджор. Однако вид, идентифицированный в формации Пинджор эпохи плио-плейстоцена, - это E. sivalensis , что позволяет предположить, что E. falconeri после длительного периода относительного успеха в конечном итоге мог пройти анагенез к плиоцену. Другие роды хищных животных, обнаруженные в формации Пинджор, включали вновь прибывших псовых , а также куньих, урсид, кошачьих (махайродонтины, пантерины и кошачьи), гиенид и виверровых. [ 28 ] Другие роды млекопитающих, обнаруженные в формации Пинджор, включают гоминид, церкопитецид, грызунов различных семейств, хоботных, лошадей трибы Equini , носорогов, свиней, оленей, жирафов и полорогих. [ 46 ]
Эфиопия
[ редактировать ]
E. dikikae и E. omoensis были крупными видами лютринов, обнаруженными в разных местах современной Эфиопии . Окаменелости E. dikikae были обнаружены в двух нижних толщах формации Хадар в долине Нижнего Аваша в Эфиопии, что указывает на то, что возраст их окаменелостей варьируется от 4 до 3,2 млн лет назад. Окаменелости E. omoensis расположены в формациях Усно и Шунгура в долине Нижнего Омо в Эфиопии. E. dikikae был назван в честь базальной пачки Дикикаэ формации Хадар, а E. omoensis произошло от долины Нижнего Омо. название [ 15 ] [ 14 ]
выделяются четыре пачки В составе плиоценовой хадарской свиты, от основания вверх, сложной толщи Дикика: пачки Базал, Сиди-Хакома, Денен-Дора и Када-Хадар. Все вместе они датируются ок. 3,5-2,9 млн лет назад и наиболее известны многочисленными останками Australopithecus afarensis . [ 47 ] Окаменелости E. dikikae известны в базальных членах формации и Сиги Хакома и неизвестны в двух других верхних членах. [ 15 ]
На основе методов определения палеообстановки, таких как экоморфологический анализ, микроизнос зубов бычьих, а также изотопов углерода и кислорода эмали, базальная пачка (БМ) имеет наибольшую численность быков и свиней в хадарской свите, что позволяет предположить, что среды, в которых они возможно, присутствовали на древесных лугах, а также в прибрежных лесах. Aepycerotini были обычным явлением в этом члене, что соответствовало племени предпочтению экотональных местообитаний между лугами и лесами. [ 48 ]
Подпачка Сиди-Хакома 1 (SH-1), возраст которой составляет от ~ 3,45 до 3,35 млн лет назад, имела аналогичную фауну и, следовательно, сходную среду обитания с другими пачками внутри формации Хадар, но также, вероятно, включала водно-болотные угодья в определенных регионах. У этого члена были обнаружены такие таксоны, как один из видов лесного племени Cephalophini и пять видов приматов, что еще раз указывает на наличие большого прибрежного леса с преимущественно лесными массивами в окрестностях. Эпицерос был самым многочисленным быком, а в SH-1 была самая низкая доля пасущихся быков среди всех подчленов формации Хадар. Гвинеи или Судана Растительность SH-1, возможно, очень напоминала растительность саванн Центральной Африки , которые переплетаются с тропическими лесами , что создает мозаику среды обитания из лугов, лесов и некоторых лесных полос. Комплексы остракод членов Basal и Sidi Hakoma указывают на источники поступления пресной воды, в которых их раковины также указывают только на трехмесячный засушливый сезон, характерный для саванн Центральной Африки. Единый засушливый сезон, обозначающий девятимесячный сезон дождей, является отличительным фактором члена Сиди Хакома от современного климата Восточной Африки, который имеет бимодальный формат засушливого сезона (два засушливых сезона), а не один. Подчлен Сиди-Хакома 2 (SH-2) похож на SH-1 и, как полагают, был связан с лесными массивами с некоторыми травянистыми равнинами, из которых Aepycerotini . Наиболее распространенным был [ 48 ]
Подчлен 3 Сиди-Хакома (SH-3) указывает на наличие лесных массивов и лугов с большим количеством водно- болотных угодий на берегах озер по сравнению с предыдущими подчленами, с повышенным присутствием бычьих быков и самой высокой численностью быков-трагелафинов , что указывает либо на более закрытые места обитания, либо на водно-болотные угодья. . Он также содержит крупнейшие комплексы микромлекопитающих современных родов мюридов , таких как современные Acomys , Golunda и Oenomys , а также вымерший Saidomys , из которых Golunda сейчас вымер в Африке. Подчлен 4 Сиди-Хакома указывает на водно-болотные угодья, окружающие озера в более засушливых условиях. Дальнейшее увеличение численности крупного рогатого скота Reduncinae и уменьшение численности коров-алселафинов указывает на указанную прибрежную среду и окружающие водно-болотные угодья. Данные о численности крупного рогатого скота предполагают одинаковую степень древесного покрова для SH-3 и SH-4 с той разницей, что последний был немного суше, чем первый. [ 48 ]

Формация Хадар представляет собой множество окаменелостей австралопитека афарского , в первую очередь частичный скелет, известный как « Люси ». Суммарный период существования вида составляет не менее 0,7 млн лет, от 3,7 до 3,0 млн лет. [ 49 ] Формация Хадар также известна своим огромным разнообразием видов быков, которые представляли большинство основных племен Африки. Трибы бычьих, которые были обнаружены в формации, включали Aepycerotini, Alcelaphini, Antilopini , Bovini , Caprini , Cephalophini, Hippotragini , Neotragini , Reduncini и Tragelaphini. Другие группы, представленные вымершими видами современных или вымерших родов, включают жирафов, гиппопотамов, свиней, псовых, кошачьих (наиболее распространенными были махайродонтины), гиенид, других куньих, виверровых, носорогов, непарнокопытных, катаров , дейнотериев и слонов. также встречается в пределах местности. Мелкие группы млекопитающих включают летучих мышей , лепорид, дикобразов Старого Света, мюридовых грызунов, спалацидов , белок и трубкозубов . [ 48 ] [ 47 ]
Другие формации плиоценового возраста в Эфиопии демонстрируют аналогичные тенденции большого разнообразия в семействе Bovidae из его многочисленных триб, а также свиней, гиппопатамидов, церкопитецидов, гоминид и непарнокопытных, принадлежащих, как правило, к тем же родам, что и формация Хадар. Большинство травоядных животных, присутствующих в формации Шунгура, либо демонстрируют постоянный рацион C 4 , либо в целом перешли от смешанного рациона C 3 -C 4 к обычному рациону C 4 , о чем свидетельствуют изменения в зубном ряду членов формации. Эти тенденции предполагают, что африканские травоядные животные в плиоцене все чаще переходили к травоядным животным C 4 , а не к бродячему и смешанному питанию, в результате растущего доминирования лугов C 4 в Африке. Однако было несколько исключений, поскольку Giraffidae и Deinotheriidae постоянно были браузерами C 3 внутри формации, в то время как трибы полорогих Aepycerotini и Tragelaphini питались преимущественно смешанным питанием с небольшими изменениями в рационе. [ 50 ] [ 51 ] [ 52 ] Из шунгурской свиты также известны остатки ископаемых рыб, а именно родов Polypterus , Sindacharax , Synodontis , Auchenoglanis и Lates . [ 53 ]
Примечания
[ редактировать ]Ссылки
[ редактировать ]- ^ Jump up to: а б с Хью Фальконер (1868). Палеонтологические мемуары и заметки покойного Хью Фальконера: Fauna Antiqua Sivalensis . Соединенное Королевство: Р. Хардвик. ISBN 112-0-015839 .
- ^ Jump up to: а б с Верма, Британская Колумбия; Гупта, СС (1992). «Enhidriodon sivalensis, гигантская ископаемая выдра из формации Сакети (верхний плиоцен), группа Сивалик, район Сирмур, Химачал-Прадеш». Журнал Палеонтологического общества Индии . 37 : 31–36.
- ^ Коутли, Проби Томас; Фальконер, Хью (1835). «Сводка ископаемых родов и видов из верхних отложений третичных толщ Сиваликской возвышенности, в сборнике авторов». Журнал Азиатского общества Бенгалии . 4 (12).
- ^ Jump up to: а б Гай Э. Пилигрим (1932). Ископаемое хищное животное Индии . Индия: Палеонтология Индики. стр. 80–88.
- ^ Jump up to: а б с Виллемсен, Джерард Ф. (1992). «Пересмотр плиоцена и четвертичного периода Lutrinae из Европы». Скрипта Геологика . 101 : 89–94.
- ^ Репеннинг, Чарльз А. (1976). «Энгидра и энгидриодон с тихоокеанского побережья Северной Америки». Журнал исследований Геологической службы США . 4 (3): 305–315.
- ^ Хурцелер, Йоханнес; Энгессер, Буркарт (1976). «Неогеновая фауна млекопитающих бассейна Баччинелло (Гроссето, Италия)». Доклады Парижской академии наук (на французском языке). 283 : 333–336.
- ^ Jump up to: а б с д и ж МОРАИС, Хорхе; Пикфорд, Мартин (2005). «Гигантский бунодонт Lutrinae из мио-плиоцена Кении и Уганды» . Геологические исследования (на испанском языке). 61 (3): 233–245. дои : 10.3989/egeol.05613-666 .
- ^ Пикфорд, Мартин; Сория, Боль; Моралес, Хорхе (2005). «Хищники позднего миоцена и базального плиоцена холмов Туген, Кения». Журнал Sociedad Geológica de España . 18 (1–2): 39–61.
- ^ Jump up to: а б с д и ж г час Пикфорд, Мартин (2007). «Ревизия мио-плиоценовых бунодонтообразных млекопитающих Индийского субконтинента». Estudios Geológicos (на испанском языке). 63 (2): 108–124. дои : 10.3989/egeol.07631192 .
- ^ Jump up to: а б с д и ж Ван, Сяомин; Гроэ, Камилла; Су, Дениз Ф.; Уайт, Стюарт К.; Цзи, Сюэпин; Келли, Джей; Яблонски, Нина Г.; Дэн, Тао; Ты, Юшан; Ян, Синь (2017). «Новая выдра гигантских размеров, Siamogale melilutra sp. nov. (Lutrinae: Mustelidae: Carnivora), из позднего миоценового стоянки Шуйтанба на северо-востоке Юньнани, юго-западного Китая, и полная доказательная филогения лютринов» . Журнал систематической палеонтологии . 16 (1): 1–24. Бибкод : 2018JSPal..16...39W . дои : 10.1080/14772019.2016.1267666 . S2CID 58892181 .
- ^ Верделин, Ларс (2003). «Хищники из стоянки гоминидов Канапои, бассейн Туркана, северная Кения » Вклад в науку 498 : 115–132. дои : 10.5962/стр.214388 . S2CID 82943977 .
- ^ Jump up to: а б с д и Валенсиано, Альберто; Гувендер, Ромала (2020). «Новые сведения о гигантских куньих (Mammalia, Carnivora, Mustelidae) из окаменелостей Лангебаанвег (Парк ископаемых Западного побережья, Южная Африка, ранний плиоцен)» . ПерДж . 8 : е9221. дои : 10.7717/peerj.9221 . ПМЦ 7271888 . ПМИД 32547866 .
- ^ Jump up to: а б с д и ж г час я дж к Гроэ, Камилла; Один, Кевин; Буассери, Жан-Рено (2022). «Lutrinae Bonaparte, 1838 (Carnivora, Mustelidae) из плио-плейстоцена долины Нижнего Омо, юго-западная Эфиопия: систематика и новые взгляды на палеоэкологию и палеобиогеографию выдр Туркана» . Comptes Rendus Palevol (на французском языке). 30 (30): 684–693. дои : 10.5852/cr-palevol2022v21a30 . S2CID 252106648 .
- ^ Jump up to: а б с д и ж г час я дж к Гераадс, Денис; Алемсегед, Зересенай; Бобе, Рене; Рид, Денне (2011). «Enhydriodon dikikae, sp. nov. (Carnivora: Mammalia), гигантская выдра из плиоцена Дикики, Нижний Аваш, Эфиопия». Журнал палеонтологии позвоночных . 31 (2): 447–453. Бибкод : 2011JVPal..31..447G . дои : 10.1080/02724634.2011.550356 . S2CID 84797296 .
- ^ Валенсиано, Альберто; Моралес, Хорхе; Пикфорд, Мартин (2022). «Lutra» libyca из мио/плиоцена Вади Натрун (Египет): новое предложение систематики . 2-й Международный симпозиум по палеонтологии позвоночных, 20–22 сентября 2022 г., стр. 31–32.
- ^ Jump up to: а б Каргопулос, Николаос; Валенсиано, Альберто; Кампуридис, Панайотис; Лехнер, Томас; Бёме, Мадлен (2021). «Новые раннепозднемиоценовые виды Vishnuonyx (Carnivora, Lutrinae) из местонахождения гоминидов Хаммершмиде, Бавария, Германия». Журнал палеонтологии позвоночных . 41 (3). Бибкод : 2021JVPal..41E8858K . дои : 10.1080/02724634.2021.1948858 . S2CID 240538139 .
- ^ «Энгидритерий терраеновае» . Флоридский музей естественной истории. 30 марта 2017 года . Проверено 10 октября 2022 г.
- ^ Константино, Пол Дж.; Ли, Джеймс Дж.-В.; Моррис, Дилан Дж.; Лукас, Питер В.; Хартстон-Роуз, Адам; Ли, Ва-Кит; Домини, Натаниэль Дж.; Каннингем, Эндрю; Вагнер, Марк; Лоун, Брайан Р. (2011). «Адаптация к питанию твердыми предметами у каланов и гомининов». Журнал эволюции человека . 61 (1): 89–96. Бибкод : 2011JHumE..61...89C . дои : 10.1016/j.jhevol.2011.02.009 . ПМИД 21474163 .
- ^ Рагхаван, П.; Пикфорд, М.; Патнаик, Р.; Гаятри, П. (2007). «Первая ископаемая мелкокогтистая выдра, Амблоникс, с примечанием о некоторых экземплярах Лутры из Верхнего Сивалика, Индия» . Estudios Geológicos (на испанском языке). 63 (2): 144–145. дои : 10.3989/egeol.07632200 .
- ^ Куфос, Джордж Д. (2011). «Миоценовая совокупность хищников Греции» . Геологические исследования (на испанском языке). 67 (2): 296–297. дои : 10.3989/egeol.40560.190 .
- ^ Альберто Валенсиано Вакеро; Ромала Говендер (21 июня 2020 г.). «Гигантские росомахи, выдры размером с волка: окаменелости открывают свежий взгляд на прошлое» . Разговор . Проверено 27 ноября 2022 г.
- ^ Jump up to: а б Льюис, Маргарет Э. (2008). «Бедренная кость вымерших бунодонтовых выдр в Африке (Carnivora, Mustelidae, Lutrinae)». Comptes Rendus Palevol (на французском языке). 7 (8): 607–627. Бибкод : 2008CRPal...7..607L . дои : 10.1016/j.crpv.2008.09.010 .
- ^ Стюарт, Кэтлин М. (2001). «Пресноводные рыбы неогеновой Африки (миоцен-плейстоцен): систематика и биогеография». Рыба и рыболовство . 2 (3): 177–230. Бибкод : 2001AqFF....2..177S . дои : 10.1046/j.1467-2960.2001.00052.x .
- ^ Пенье, Стефан; Де Бонис, Луи; Ликиус, Андосса; Маккей, Хасан Таиссо; Виньо, Патрик; Брюне, Мишель (2008). «Позднемиоценовые хищники из Чада: Lutrinae (Mustelidae)» . Зоологический журнал Линнеевского общества . 152 (4): 793–846. дои : 10.1111/j.1096-3642.2008.00377.x .
- ^ Верделин, Ларс; Манти, Фредрик Виллидж (2012). «Хищник из стоянки гоминидов Канапои, северная Кения» Журнал африканских наук о Земле . 64 : 1–8. Бибкод : 2012JAfES..64....1W . doi : 10.1016/j.jafrearsci.2011.11.003 .
- ^ Верделин, Ларс; Льюис, Маргарет Э. (2020). «Контекстуальный обзор хищников Канапои» . Журнал эволюции человека . 140 : 102334. Бибкод : 2020JHumE.14002334W . дои : 10.1016/j.jhevol.2017.05.001 . ПМИД 28625408 . S2CID 23285088 .
- ^ Jump up to: а б с Ясински, Стивен Э.; Аббас, Сайед Гьюр; Махмуд, Халид; Бабар, Мухаммад Адиб; Хан, Мухаммад Акбар (2022). «Новые экземпляры Carnivoran (Mammalia: Carnivora) от сиваликов Пакистана и Индии и их фаунистические и эволюционные последствия». Историческая биология: Международный журнал палеобиологии . 35 (11): 1–36. дои : 10.1080/08912963.2022.2138376 . S2CID 253346978 .
- ^ Хан, Мухаммад Акбар; Ахтар, Мухаммед; Икрам, Тасним. «Настоящие копытные из местонахождения типа Нагри (поздний миоцен), Северный Пакистан». Журнал наук о животных и растениях . 22 : 1–59.
- ^ Jump up to: а б Мартин, Жереми (2018). «Таксономическое содержание рода Gavialis с холмов Сивалик в Индии и Пакистане» (PDF) . Статьи по палеонтологии . 5 (3): 483–497. дои : 10.1002/spp2.1247 . S2CID 134966832 .
- ^ Хан, Мухаммад Акбар; Ахтар, Мухаммед; Илиопулос, Джордж (2012). «Трагулиды (Artiodactyla, Ruminantia, Tragulidae) из средних сиваликов Хаснота (поздний миоцен), Пакистан» Итальянский журнал палеонтологии и стратиграфии . 118 (2): 325–341. дои : 10.13130/2039-4942/6009 .
- ^ Гаффар, Абдул; Ахтар, Мухаммед; Бабар, Мухаммад Адиб; Рухи, Газала (2018). «Олени из районов Дхок-Патан и Хаснот (формация Дхок-Патан, Северный Пакистан) с комментариями о возрасте этих отложений» . Бразильский журнал палеонтологии . 21 (2): 132–140. дои : 10.4072/rbp.2018.2.04 . S2CID 59060740 .
- ^ Аббас, Дьёр; Бабар, Мухаммад Адиб; Хан, Мухаммад Акбар; Ницца, Бадар (2021). «Новый материал хоботных из группы Сивалик в Пакистане с замечаниями по некоторым видам» . ПалеоБиос 38 (1): 1–12. дои : 10.5070/P9381054113 . S2CID 237772570 .
- ^ Бабар, Мухаммад Адиб; Афтаб, Киран; Хан, Мухаммад Акбар; Аббас, Дюр (2021). «Tetraconodon и Sivachoerus (Suidae) из района Дхок-Патан и Хаснот на плато Потвар, северный Пакистан». Арабский журнал геонаук . 14 (2473): 1–6. Бибкод : 2021ArJG...14.2473B . дои : 10.1007/s12517-021-08684-x . S2CID 244120161 .
- ^ Ван, Сяомин; Флинн, Лоуренс; Фортелиус, Микаэль (2013). «Глава 17: Биостратиграфия млекопитающих неогенового периода Сивалик Индии: обзор». Ин Ван, Сяомин; Флинн, Лоуренс Дж.; Фортелиус, Микаэль (ред.). Ископаемые млекопитающие Азии: биостратиграфия и хронология неогена . Издательство Колумбийского университета. стр. 423–444. дои : 10.7312/wang15012-017 . ISBN 9780231520829 .
- ^ Барри, Джон К.; Морган, Мишель Э.; Флинн, Лоуренс Дж.; Пилбим, Дэвид; Беренсмейер, Анна К.; Раза, С. Махмуд; А. Хан, Имран; Бэджли, Кэтрин; Хикс, Джейсон; Келли, Джей (2002). «Изменения фауны и окружающей среды в позднемиоценовых сиваликах Северного Пакистана». Палеобиология . 28 (2): 1–71. doi : 10.1666/0094-8373(2002)28[1:FAECIT]2.0.CO;2 . S2CID 18408360 .
- ^ Нельсон, Шерри В. (2007). «Изотопные реконструкции изменения среды обитания, связанного с исчезновением сивапитека, миоценового гуманоида, в группе Сивалик в Пакистане» (PDF) . Палеогеография, Палеоклиматология, Палеоэкология . 243 (1–2): 204–222. Бибкод : 2007PPP...243..204N . дои : 10.1016/j.palaeo.2006.07.017 .
- ^ Патнаик, Раджив; Сингх, Нинтхуджам Премджит; Пол, Дебаджьоти; Сукумар, Раман (2019). «Изменения в питании и среде обитания в зависимости от климата неоген-четвертичных хоботных и связанных с ними млекопитающих Индийского субконтинента». Четвертичные научные обзоры . 224 : 105968. Бибкод : 2019QSRv..22405968P . doi : 10.1016/j.quascirev.2019.105968 . S2CID 210307849 .
- ^ Ма, Цзяо; Ван, Юань; Цзинь, Чанчжу; Ху, Яову; Бочеренс, Эрве (2019). «Экологическая гибкость и дифференциальная выживаемость плейстоценовых Stegodon orientalis и Elephas maximus на материковой части Юго-Восточной Азии, выявленная с помощью анализа стабильных изотопов (C, O)». Четвертичные научные обзоры . 212 : 33–44. Бибкод : 2019QSRv..212...33M . doi : 10.1016/j.quascirev.2019.03.021 . S2CID 135056116 .
- ^ Джукар, Адвайт Мухеш; Сунь, Боян; Нанда, Авинаш К.; Бернор, Раймонд Л. (2019). «Первое появление Eurygnathohippus Van Hoepen, 1930 (Mammalia, Perissodactyla, Equidae) за пределами Африки и его биогеографическое значение». Бюллетень Итальянского палеонтологического общества . 58 (2): 171–179. дои : 10.4435/BSPI.2019.13 .
- ^ Хан, Мухаммад Акбар; Джамиль, Мадиха; Махмуд, Халид; Бабар, Мухаммад Адиб (2014). «Некоторые новые остатки оленьих (Cervidae, Ruminantia) из формации Татрот в Северном Пакистане». Биология . 60 (2): 225–230.
- ^ Кумар, Суреш (2022). «Новые окаменелости Sivatherium Giganteum (Giraffidae, Mammalia) из Верхних Сиваликов Индийского субконтинента» . Acta Palaeontologica Romane . 18 (2): 85–92. doi : 10.35463/апреля 2022.02.04 . S2CID 248233024 .
- ^ Томас А., Стидхэм; Кришан, Кевал; Сингх, Бахадур; Гош, Абхик; Патнаик, Раджив (2014). «Пеликан Tarsometatarsus (Aves: Pelecanidae) из последних плиоценовых сиваликов Индии» . ПЛОС ОДИН . 9 (11): 1–8. Бибкод : 2014PLoSO...9k1210S . дои : 10.1371/journal.pone.0111210 . ПМЦ 4218845 . ПМИД 25365300 .
- ^ Гарбин, Рафаэлла Дж.; Бандиопадхьяй, Сасвати; Джойс, Уолтер Г. (2020). «Таксономический пересмотр геомидидных черепах эпохи Сивалик в Индии и Пакистане» . Европейский журнал таксономии (652): 1–67. дои : 10.5852/ejt.2020.724.1165 . S2CID 229387265 .
- ^ Клаус, Себастьян; Сингх, Бахадур; Хартманн, Лукас; Кришан, Кевал; Гош, Абхик; Патнаик, Раджив (2017). «Ископаемый пресноводный краб из плиоценовой формации Татрот (группа Сивалик) в Северной Индии (Crustacea, Brachyura, Potamidae)». Палеомир . 26 (3): 566–571. дои : 10.1016/j.palwor.2016.08.003 .
- ^ Нанда, Авинаш К.; Сегал, Рамеш К.; Чаухан, Парт Р. (2018). «Фауна возраста Сивалик из Гималайского предгорного бассейна Южной Азии». Журнал азиатских наук о Земле . 162 : 54–68. Бибкод : 2018JAESc.162...54N . дои : 10.1016/j.jseaes.2017.10.035 . S2CID 134929306 .
- ^ Jump up to: а б Герадс, Деннис; Алемсегед, Зересена; Зло, Рене; Рид, Денни (2015). «Плиоценовые хищники (млекопитающие) из формации Хадар в Дикике, нижняя долина Аваш, Эфиопия» Журнал африканских наук о Земле . 107 : 28–35. Бибкод : 2015JAfES.107...28G . doi : 10.1016/j.jafrearsci.2015.03.020 .
- ^ Jump up to: а б с д Кампизано, Кристофер Дж.; Роуэн, Джон; Рид, Кэй Э. (2022). «Глава 18: Формация Хадар, региональный штат Афар, Эфиопия: геология, фауна и реконструкции палеоэкологической среды». В Рейнольдсе, Салли К.; Бобе, Рене (ред.). Африканская палеоэкология и эволюция человека . Издательство Кембриджского университета. стр. 214–228. дои : 10.1017/9781139696470.018 .
- ^ Алемсегед, Зересенай; Винн, Джонатан Г.; Кимбел, Уильям Х.; Рид, Денне; Гераадс, Денис; Бобе, Рене (2005). «Новый человекообразный человек из базальной части формации Хадар, Дикика, Эфиопия, и его геологический контекст». Журнал эволюции человека . 49 (4): 499–514. Бибкод : 2005JHumE..49..499A . дои : 10.1016/j.jhevol.2005.06.001 . ПМИД 16026815 .
- ^ Бобе, Рене (2011). «Ископаемые млекопитающие и палеосреда в бассейне Омо-Туркана». Эволюционная антропология . 20 (6): 254–263. дои : 10.1002/evan.20330 . ПМИД 22170694 . S2CID 205826374 .
- ^ Негаш, Энкуй В.; Алемсегед, Зересена; Зло, Рене; Грин, Фредерик; Спонхаймер, Мэтт; Дж. Винн, Джонатан (2020). «Тенденции в питании травоядных животных формации Шунгура, юго-запад Эфиопии » Труды Национальной академии наук . 117 (36): 21921–21927. Бибкод : 2020PNAS..11721921N дои : 10.1073/pnas.2006982117 . ПМЦ 7486712 . ПМИД 32839326 .
- ^ Биби, Фейсал; Сурон, Энтони; Бочеренс, Эрве; Один, Кевин; Буассери, Эн-Рено (2013). «Экологические изменения в нижней части долины Омо около 2,8 млн лет назад» . по биологии Письма 9 (1): 1–4. дои : 10.1098/rsbl.2012.0890 . ПМЦ 3565503 . ПМИД 23234862 . S2CID 10791621 .
- ^ Стюарт, Кэтлин М.; Мюррей, Элисон (2008). «Рыбные останки из плио-плейстоценовой формации Шунгура, бассейн реки Омо, Эфиопия». Геобиос 41 (2): 283–295. Бибкод : 2008Geobi..41..283S . дои : 10.1016/j.geobios.2007.06.004 .







- Выдры
- Доисторические куньи
- Миоценовые куньи
- Ископаемые таксоны, описанные в 1868 году.
- Миоценовые млекопитающие Африки
- Плиоценовые млекопитающие Африки
- Плейстоценовые млекопитающие Африки
- Миоценовые млекопитающие Азии
- Плиоценовые млекопитающие Азии
- Таксоны, названные Хью Фалконером
- Роды доисторических хищников