Келенкен
| Келенкен Временной диапазон: средний миоцен ( Коллонкуран ). ~
| |
|---|---|

| |
| Реконструированный скелет в Музее динозавров префектуры Фукуи. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Птицы |
| Заказ: | Кариамиобразные |
| Семья: | † Форусрациды |
| Подсемейство: | † Форусрацины |
| Род: | † Келенкен Бертелли и др. 2007 год |
| Разновидность: | † К. Гильермои
|
| Биномиальное имя | |
| † Келенкен гильермои Бертелли и др. 2007 год
| |
Келенкен — род phorusrhacid ( « птица ужаса»), вымершей группы крупных хищных птиц , обитавших на территории нынешней Аргентины в среднем миоцене около 15 миллионов лет назад. Единственный известный экземпляр был обнаружен старшеклассником Гильермо Агирре-Сабала в Комальо , в регионе Патагонии , и стал голотипом нового рода и вида Kelenken guillermoi в 2007 году. Название рода отсылает к духу из Теуэльче мифологии , и конкретное название чтит первооткрывателя. Голотип состоит из одного из наиболее полных известных черепов крупного форусрацида, а также кости предплюсне-плюсневого сустава голени и фаланги пальца стопы . Открытие Келенкена прояснило анатомию крупных форусрацидов, поскольку ранее они были гораздо менее известны. Ближайшими живыми родственниками форусрацидов являются сериемы . келенкен Было обнаружено, что принадлежит к подсемейству Phorusrhacinae, наряду, например, с Devincenzia .
Форусрациды представляли собой крупные нелетающие птицы с длинными задними конечностями, узким тазом, пропорционально маленькими крыльями и огромными черепами, с высоким, длинным, сжатым вбок крючковатым клювом. Келенкен — самый крупный из известных форусрацидов, он на 10% крупнее своих крупнейших родственников, известных ранее. Череп голотипа длиной 716 мм (2,3 фута) является самым крупным из известных птиц и сравним по размеру с черепом лошади . Кость предплюсне-плюсневой кости ноги имеет длину 437 мм (17 дюймов). Считается, что Келенкен имел рост около 3 м (9,8 футов) и вес более 100 кг (220 фунтов). Келенкен отличался от других форусрацидов такими особенностями, как длина клюва, наличие супраорбитального окостенения (закругленный край над глазницей ), которое вписывается в лунку заглазничного отростка , а также наличие почти треугольного затылочного отростка (большого заглазничного отростка). отверстие в основании черепа, через которое входит спинной мозг ).
Считается, что форусрациды были наземными хищниками или падальщиками и часто считались высшими хищниками , которые доминировали в кайнозойской Южной Америке в отсутствие плацентарных хищников-млекопитающих, хотя они действительно сосуществовали с некоторыми крупными хищными млекопитающими боргиенид . Длинные и тонкие предплюсны келенкена позволяют предположить, что он мог бегать быстрее, чем предполагалось ранее для крупных форусрацидов, и мог преследовать мелких животных. Исследования родственных Andalgalornis показывают, что крупные форусрациды имели очень жесткий и жесткий череп; это указывает на то, что они, возможно, проглотили мелкую добычу целиком или нацелились на более крупную добычу повторяющимися ударами клюва. Келенкен известен из формации Коллон Кура и жил в эпоху Коллонкура в Южной Америке, когда преобладала открытая среда, что позволяло более беглым встречаться (приспособленным к бегу) и крупным животным. В формации обнаружены окаменелости широкого спектра млекопитающих , а также несколько окаменелостей птиц, рептилий, земноводные и рыбы.
Таксономия
[ редактировать ]

Примерно в 2004 году окаменелости форусрацида ( или «птицы ужаса», группы крупных хищных птиц) были обнаружены аргентинским старшеклассником Гильермо Агирре-Сабала между двумя домами, примерно в 100 м (330 футов) от железной дороги Комалло . небольшая деревня на северо-западе провинции Рио-Негро в регионе Патагония в Аргентине (координаты: 41 ° 01'59,4 дюйма ю.ш., 70 ° 15'29,7 дюйма з.д. / 41,033167 ° ю.ш., 70,258250 ° з.д. ). Обнажения, где был обнаружен образец, относятся к формации Коллон Кура . Агирре-Сабала сам подготовил образец , и это открытие заставило его переключиться с изучения психологии на изучение палеонтологии и наук о Земле. [ 1 ] [ 2 ] [ 3 ]

Образец стал частью коллекции Museo Asociación Paleontológica Bariloche в Рио-Негро, где он был внесен в каталог как образец BAR 3877-11. [ 3 ] До того, как животное получило научное название , об этом образце сообщили и обсудили аргентинские палеонтологи Луис М. Чиаппе и Сара Бертелли в короткой статье 2006 года. [ 3 ] [ 4 ] В 2007 году Бертелли, Кьяппе и Клаудия Тамбусси сделали этот экземпляр голотипом Kelenken guillermoi ; Название рода относится к духу в мифологии народа теуэльче в Патагонии, который представлен в виде гигантской хищной птицы , а видовое название дано в честь его первооткрывателя. [ 3 ]
Голотип и единственный известный экземпляр состоит из почти целого черепа, несколько раздавленного сверху вниз, при этом большая часть глазниц , крыша черепа, черепная коробка и левая квадратная кость сохранились, в то время как большая часть небных костей за глазницами сохранилась . отсутствующий. Образец также включает связанную левую предплюсну (кость голени птиц), небольшую верхнюю часть кости фаланги стопы (кость пальца ноги) и несколько неопределенных фрагментов. Описатели пришли к выводу, что эти кости принадлежали одному экземпляру, поскольку они были собраны вместе (и при отсутствии других окаменелостей), потому что их общая сохранность (например, цвет и текстура) была одинаковой, а также потому, что они морфологически соответствовали принадлежности к большому форусрацид. У этого экземпляра был самый полный череп крупного форусрацида, известного на тот момент. Ранее такие черепа были известны только из фрагментарных Девинченции и Форусракоса . Череп последнего при сборе распался (остался только кончик клюва), что затруднило сравнение форусрацида таксоны разных размеров, вплоть до открытия келенкена . [ 3 ]
Эволюция
[ редактировать ]

В своем описании 2007 года Бертелли и его коллеги классифицировали келенкен как члена семейства Phorusrhacidae , основываясь на его огромном размере в сочетании со сжатым вбок, сильно крючковатым клювом (или рострумом , частью челюстей, образующих клюв) и выпуклой кульмен (верхушка верхнего клюва). форусракид В то время было признано пять подсемейств (Brontornithinae, Phorusrhacinae, Patagornithinae, Mesembriornithinae и Psilopterinae), хотя их достоверность тогда не была подтверждена кладистическим анализом , и описатели обнаружили, что келенкен наиболее похож на таксоны, которые традиционно считались форусракинами. Общие черты с форусракинами включают низкую и сжатую сверху вниз заднюю часть черепа, широкий затылочный столик , тупой заглазничный отросток и предплюсне-плюсневую кость, сходную с таковой у Титаниса в том, что супратрохлеарная поверхность нижнего конца плоский. Дальнейшее сравнение было затруднено отсутствием анатомической информации о форусракинах. [ 3 ] [ 5 ]
Бразильский палеонтолог Эркулано Альваренга и его коллеги опубликовали в 2011 году филогенетический анализ Phorusrhacidae, который показал, что Kelenken и Devincenzia являются сестринскими таксонами , ближайшими родственниками друг друга. Хотя анализ подтвердил существование пяти подсемейств, полученная кладограмма не разделила Brontornithinae, Phorusrhacinae и Patagornithinae. [ 6 ] В своем описании Llallawavis в 2015 году аргентинский палеонтолог Федерико Дж. Дегранж и его коллеги провели филогенетический анализ Phorusrhacidae, в ходе которого они обнаружили, что Phorusrhacinae являются полифилетическими (неестественная группировка). Следующая кладограмма показывает положение Келенкена по итогам анализа 2015 года: [ 7 ]
В раннем кайнозое , после вымирания динозавров, не являющихся птицами , млекопитающие подверглись эволюционному разнообразию , а у некоторых групп птиц по всему миру развилась тенденция к гигантизму ; сюда входили Gastornithidae , Dromornithidae , Palaeognathae и Phorusrhacidae. [ 6 ] [ 8 ] Форусрациды — вымершая группа Cariamiformes , единственными живыми представителями которой являются два вида серием семейства Cariamidae. Хотя это наиболее специфическая группа среди Cariamiformes, взаимоотношения между форусрацидами неясны из-за неполноты их остатков. [ 9 ]
Форусрациды присутствовали в Южной Америке с палеоцена (когда континент был изолированным островом) и дожили до плейстоцена . Они также появились в Северной Америке в конце плиоцена , во время Великого американского биотического обмена , и хотя к этой группе относят окаменелости из Европы, их классификация оспаривается. Неясно, откуда возникла группа; и кариамиды, и форусрациды могли возникнуть в Южной Америке или прибыли из других мест, когда южные континенты были ближе друг к другу или когда уровень моря был ниже. [ 5 ] [ 6 ] Келенкен Сам жил в среднем миоцене , около 15 миллионов лет назад. [ 3 ] Поскольку форусрациды дожили до плейстоцена, они, по-видимому, были более успешными, чем, например, южноамериканские метатерийные тилакосмилиды- хищники (которые исчезли в плиоцене), и возможно, что они экологически конкурировали с плацентарными хищниками, пришедшими из Северной Америки в плейстоцене. . [ 10 ]
Описание
[ редактировать ]
Форусрациды представляли собой крупные нелетающие птицы с длинными задними конечностями, узким тазом, пропорционально маленькими крыльями и огромными черепами, с высоким, длинным, сжатым вбок крючковатым клювом. [ 9 ] Келенкен — самый крупный из известных форусракидов, примерно на 10% крупнее самых крупных ранее известных форусрацидов, таких как Форусракос . Череп голотипа имеет длину около 716 мм (2,3 фута) от кончика клюва до центра сагиттального затылочного гребня в верхней части задней части головы (размер сравним с размером его черепа лошади), что делает самый большой череп среди всех известных птиц. Задний конец черепа имеет ширину 312 мм (12 дюймов). Кость предплюсне-плюсневой кости ноги имеет длину 437 мм (17 дюймов). [ 1 ] [ 3 ] [ 8 ] Высота головы достигала 3 м (9,8 фута), а современные серии достигают 90 см (3,0 фута) в высоту. Хотя вес Келенкена точно не подсчитывался, предполагается, что он превышал 100 кг (220 фунтов). [ 8 ] [ 11 ]
Череп
[ редактировать ]До открытия Келенкена черепа не полностью известных крупных форусрацидов были реконструированы как увеличенные версии черепов более мелких и полных родственников, таких как Psilopterus и Patagornis , о чем свидетельствует часто воспроизводимый в 1895 году эскиз разрушенного черепа большого форусракоса . который сам был основан на Патагорнисе . Эти реконструкции подчеркнули их предполагаемые очень высокие клювы, круглые, высокие глазницы и сводчатые черепные коробки, но Келенкен продемонстрировал значительную разницу между черепами крупных и мелких членов группы. [ 3 ] [ 4 ] Череп голотипа очень массивный, при взгляде сверху треугольный, со сжатой сверху вниз задней частью. Верхний клюв очень длинный, превышает половину общей длины черепа, в отличие от Mesembriornis и Patagornis , и длиннее, чем у Phorusrhacos . Соотношение между верхним клювом и черепом келенкена составляет 0,56, исходя из расстояния между костной ноздрей и передним кончиком. Несмотря на раздавливание сверху вниз, верхний клюв высокий и очень крепкий, хотя, по-видимому, не такой высокий, как у патагорнисов, таких как Patagornis , Andrewsornis и Andalgalornis . [ 3 ]

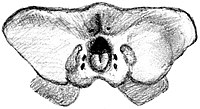
Передний конец предчелюстной кости (самая передняя кость верхней челюсти) заметно выступает в виде острого, загнутого вниз крючка. Такая сильная проекция вниз больше всего напоминает состояние, наблюдаемое у форусрацидов большого и среднего размера, таких как Phorusrhacos , Patagornis , Andrewsornis и Andalgalornis , а не более слабые выступы более мелких псилоптеринов. Нижняя часть передней части верхнего клюва образует пару выступающих гребней, каждый из которых отделен бороздкой от томиума или острого края клюва. Эти гребни также отделены от более широкой центральной части предчелюстной кости продольной бороздкой (ростральный предчелюстной канал). Патагорнис имел аналогичную морфологию передней части неба. Большая часть верхней части клюва покрыта небольшими ямками неправильной формы, которые служили выходами нервов. В задних двух третях верхнего клюва имеется выступающая бороздка, идущая параллельно краю томиума. [ 3 ]
Ноздри маленькие, прямоугольной формы, расположены в верхнем заднем углу верхнего клюва, как у патагорнитинов (у более крупных форусракинов и бронторнитинов размер и расположение ноздрей неизвестны). Ноздри кажутся длиннее спереди назад, чем высота, хотя это может быть преувеличено из-за раздавливания, а их задний край образован верхнечелюстным отростком носовой кости (выступом от носовой кости к верхней челюсти , основной кости носовой кости). верхняя челюсть). соединены ли ноздри друг с другом посередине (без перегородки Неизвестно, , как у других форусрацидов). Четырехугольная форма анторбитального окна (отверстия перед глазницей) четкая, несмотря на то, что оно несколько раздавлено с обеих сторон. Передняя граница этого отверстия находится примерно на одном уровне с задним краем ноздри, а нижний край его прямой, если смотреть с левой стороны. Прочные слезные кости образуют задние края предглазничных окон, и эти кости были утоплены по отношению к скуловая перемычка (образовавшая нижний край глазницы) и наружный боковой край каждой лобной кости (основные кости лба). Анторбитальное окно пропорционально меньше, чем у Патагорниса . [ 3 ] [ 4 ]

Хотя форма глазниц могла слегка измениться при сжатии сверху вниз, вполне вероятно, что они были низкими, почти прямоугольной формы, с вогнутым верхним краем и слегка выпуклым нижним краем. Верхняя часть глазницы очерчена толстым закругленным краем (надглазничное окостенение ), задняя часть которого при взгляде сбоку кажется нависающей вниз. У Патагорниса подобная структура, как предполагается, является отростком слезной кости, и хотя связь между ними не ясна у Келенкена , эта структура, вероятно, также была продолжением слезной кости. Супраорбитальное окостенение помещается в лунку, образованную частью лобной кости, образующей заглазничный отросток, конфигурация, неизвестная у других форусрацидов. Нижний край глазницы образован мощной скуловой перемычкой, очень высокой (больше, чем у Devincenzia ) и плоской из стороны в сторону. Скуловая кость примерно в четыре раза выше толщины у нижнего центра глазницы, а ее высота больше, чем у других форусрацид. [ 3 ]
Лобные кости, по-видимому, были плоскими с верхней стороны. Область соприкосновения лобных костей с предчелюстными костями повреждена настолько, что их швы (стыки между ними) не могут быть идентифицированы, но швы между лобными костями и носовыми и теменными костями полностью сращены. Из-за этого слияния трудно определить, каким образом эти кости были частью крыши черепа, но тупые и прочные заглазничные отростки, вероятно, в основном образовались из лобных костей. На нижних сторонах каждая лобная часть образует большое углубление, к которому прикрепляется челюстная мышца. Заглазничный отросток узко отделен от мощного скулового отростка , и эти два выступа заключают в себе узкую височную ямку (отверстие на виске ). Заглазничный отросток содержит рубцы, оставленные массивными челюстными мышцами , части которых прорастали большую часть крыши черепа на уровне теменных костей. Позади скулового отростка, со стороны чешуйчатой кости , имеется хорошо развитое углубление, соответствующее мышце, закрывающей челюсть. Подвисочная ямка сзади широкая, ее задняя часть определяется тупым, направленным в сторону расширением затылочного гребня. [ 3 ]
Максиллы образуют обширное небо, боковые края которого почти параллельны на большей части длины верхнего клюва, а небо расширяется от передней части назад к области глазниц. Как и у Patagornis , эти кости разделены по средней линии отчетливой продольной впадиной, проходящей на большей части их длины, а вдоль задней половины неба эта впадина окружена частями верхних челюстей. Боковой край задней части верхней челюсти имеет хорошо выраженный шовный контакт со скуловой костью, как у Patagornis . Часть крыши черепа за глазницами плоская и покрыта рубцами из-за развития височной мускулатуры . Затылочный столик очень широкий, как у Devicenzia , и низкий, что придает ему прямоугольный вид, если смотреть сзади. ( Затылочный мыщелок округленный выступ на затылке, соприкасающийся с первым шейным позвонком ) круглый, с вертикальной бороздкой, берущей начало на его верхней поверхности и доходящей почти до центра мыщелка. foramen magnum (большое отверстие в основании черепа, через которое входит спинной мозг ) имеет почти треугольную форму, что уникально для этого рода, имеет тупую верхнюю вершину и немного меньше мыщелка. Над большим затылочным отверстием имеется гребневидный выступ, вертикально идущий от края отверстия до поперечного затылочного гребня. Ямка (неглубокое углубление) под мыщелком не видна, в отличие от Patagornis и Devicenzia , у которых ямки отчетливые. [ 3 ] [ 4 ]
Кость ноги
[ редактировать ]
стержень предплюсны несколько тонкий, с почти прямоугольной средней частью, как у Phorusrhacos . Верхние две трети его верхней поверхности вогнутые, а нижняя треть более плоская. Предплюсневидные кости имеют котилы (две чашеобразные полости на верхнем конце стержня), почти овальные и глубоко вогнутые. Латеральная котила на внешней стороне меньше медиальной котилии на внутренней стороне и находится немного ниже ее. Междольковые возвышения между семядолями хорошо развиты и крепкие, как и у других форусрацидов. Уникальным для этого рода является наличие круглого бугорка в медиоподошвенном углу латеральной котилии, ниже по высоте, чем междольковое возвышение. Середина стержня предплюсны неправильно-четырехугольная, в отличие от таковой у бронторнитинов, которые прямоугольные и очень широкие. Блок третьей плюсневой кости («костяшки» предплюсне-плюсневой кости, сочленяющиеся с верхней частью пальцевых фаланг) намного больше двух других блоков (второго и четвертого) и выступает гораздо дальше вниз, а четвертый блок находится ниже. больше второго. Четвертый блок неправильно-четырехугольной формы, что контрастирует с прямоугольным блоком Девеценция . Дистальное сосудистое отверстие — отверстие на нижней передней стороне предплюсне-плюсневого сустава — имеет центральное положение, над верхними концами третьего и четвертого блоков. [ 3 ]
Палеобиология
[ редактировать ]Кормление и диета
[ редактировать ]
Считается, что форусрациды были наземными хищниками или падальщиками и часто считались высшими хищниками , которые доминировали в кайнозойской Южной Америке в отсутствие плацентарных хищников-млекопитающих, хотя они действительно сосуществовали с некоторыми крупными хищными млекопитающими боргиенид . Более ранние гипотезы об экологии питания форусрацидов основывались главным образом на их больших черепах с крючковатыми клювами, а не на основе детальных гипотез и биомеханических исследований, а такие исследования их беговых и хищнических адаптаций проводились только с начала XXI века. [ 9 ] [ 12 ]
Альваренга и Элизабет Хёфлинг сделали несколько общих замечаний о привычках к форусрациду в статье 2003 года. Они не летали, о чем свидетельствует пропорциональный размер их крыльев и масса тела, а размер крыльев был более уменьшен у более крупных членов группы. Эти исследователи отметили, что сужение таза, верхней части верхней челюсти и грудной клетки могло быть адаптацией, позволяющей птицам искать и ловить более мелких животных в высоких растениях или на пересеченной местности. Большие расширения над глазами, образованные слезными костями (похожие на то, что наблюдается у современных ястребов ), защищали глаза от солнца и обеспечивали острое зрение, что указывает на то, что они охотились зрением на открытых, освещенных солнцем местах, а не в тени. леса. [ 5 ]
Функция ноги
[ редактировать ]
В 2005 году Рудемар Эрнесто Бланко и Вашингтон В. Джонс исследовали прочность большеберцовой кости (голени) форусрацидов, чтобы определить их скорость, но признали, что такие оценки могут быть ненадежными даже для современных животных. В то время как сила большеберцовой кости Патагорниса и неопределенный большой форусрацин предполагали скорость 14 м / с (50 км / ч; 31 миль в час), а у Мезембриорниса - 27 м / с (97 км / ч; 60 миль в час), последний больше, чем у современного страуса , приближаясь к скорости гепарда , 29 м/с (100 км/ч; 65 миль в час). [ 13 ] [ 14 ] Они сочли эти оценки маловероятными из-за большого размера тела этих птиц и вместо этого предположили, что эта сила могла быть использована для того, чтобы сломать длинные кости млекопитающих среднего размера, размером, например, сайгака или газели Томсона . Эту силу можно было использовать для доступа к костному мозгу внутри костей или для использования ног в качестве ударного оружия (как это делают некоторые современные наземные птицы), что соответствует большим, изогнутым и сжатым в стороны когтям, известным у некоторых форусрацидов. Они также предположили, что будущие исследования позволят выяснить, могли ли они использовать свои клювы и когти против хорошо бронированных млекопитающих, таких как броненосцы и глиптодонты . [ 13 ]
По мнению Чьяппе и Бертелли в 2006 году, открытие келенкена поставило под сомнение традиционную идею о том, что размер и ловкость форусрацидов коррелируют, причем более крупные члены группы более громоздки и менее приспособлены к бегу. Вместо этого длинные и тонкие предплюсны Келенкена показывают, что эта птица, возможно, была намного быстрее, чем меньший, более тяжелый и медленный Brontornis . [ 4 ] В новостной статье 2006 года об открытии Чьяппе заявил, что, хотя келенкен , возможно, и не был таким быстрым, как страус, он явно мог бегать быстрее, чем предполагалось ранее для крупных форусрацидов, основываясь на длинных, тонких костях ног, внешне похожих на них. до современных нелетающих реев . В статье предполагалось, что Келенкен мог бы преследовать мелких млекопитающих и рептилий. [ 1 ] В другой новостной статье 2006 года Чьяппе заявил, что келенкен был бы таким же быстрым, как борзая , и что, хотя в то время в Южной Америке были и другие крупные хищники, их численность была ограничена, и они не были такими быстрыми и проворными, как форусракиды, и многочисленные пасущиеся млекопитающие могли бы обеспечить обильную добычу. Чьяппе заявил, что форусрациды грубо напоминали более ранних хищных динозавров , таких как тираннозавр , имея гигантские головы, очень маленькие передние конечности и очень длинные ноги, и, следовательно, имели такие же приспособления к поеданию мяса. [ 15 ]
Функция черепа и шеи
[ редактировать ]

среднего размера Исследование форусрацида Andalgalornis , проведенное Дегранжем и его коллегами в 2010 году и основанное на анализе методом конечных элементов с использованием компьютерной томографии , позволило оценить силу укуса и распределение напряжения в черепе. Они обнаружили, что сила укуса составляет 133 ньютона на кончике клюва, и показали, что он в значительной степени утратил внутричерепную неподвижность (подвижность костей черепа по отношению друг к другу), как и в случае с другими крупными форусрацидами, такими как келенкен . Эти исследователи интерпретировали эту потерю как адаптацию к повышенной жесткости черепа; По сравнению с современными красноногими серимами и орланами-белохвостами , череп форусрацида демонстрировал относительно высокую нагрузку при боковых нагрузках, но низкую нагрузку при приложении силы вверх и вниз, а также при моделировании «отката». Из-за относительной слабости черепа по бокам и средней линии эти исследователи сочли маловероятным, что Andalgalornis вел потенциально рискованное поведение, заключающееся в использовании клюва для подчинения крупной борющейся добычи. Вместо этого они предположили, что он либо питался более мелкой добычей, которую можно было убить и съесть более безопасно, например, заглатывая ее целиком, либо, нацеливаясь на крупную добычу, он использовал серию точно нацеленных повторяющихся ударов клювом, Стратегия «нападение-отступление». Борющуюся добычу также можно было удержать ногами, несмотря на отсутствие острых когтей. [ 12 ]
Последующее исследование 2012 года, проведенное Тамбусси и его коллегами, проанализировало гибкость шеи Andalgalornis , основываясь на морфологии его шейных позвонков , и обнаружило, что шея разделена на три части. Манипулируя позвонками вручную, они пришли к выводу, что мускулатура и скелет шеи Andalgalornis приспособлены для того, чтобы нести большую голову и помогать ей подниматься из максимального разгибания после удара вниз. Исследователи предположили, что то же самое можно сказать и о других крупных , большеголовые форусрациды. [ 16 ] Исследование морфологии черепа форусрацидов, проведенное Дегранжем в 2020 году, показало, что внутри группы существует два основных морфотипа, происходящих от сериемоподобного предка. Это был «Тип черепа псилоптерина», который был плезиоморфным (более похожим на предковый тип), и «Тип черепа ужасной птицы», который включал келенкена и других крупных представителей, который был более специализированным, с более жесткими и жесткими черепами. Несмотря на различия, исследования показали, что эти два типа обращались с добычей одинаково, в то время как более твердые черепа и, как следствие, большая сила укуса типа «Птица ужаса» были бы адаптацией к обращению с более крупной добычей. [ 9 ]
Палеосреда
[ редактировать ]
Келенкен был обнаружен в пирокластических (камнях, выброшенных в результате извержений вулканов ) обнажениях, принадлежащих формации Коллон Кура в юго-восточном углу Комальо, Патагония, области, покрытой беловатыми туфами . этого района Стратиграфия в то время была только предварительно изучена, и возраст отложений не был должным образом определен, но по сравнению с другими ископаемыми пластами возраста наземных млекопитающих Южной Америки и радиоизотопными датировками из разных областей формации Коллон Кура, он По оценкам, датируется коллонкуранским возрастом среднего миоцена, около 15 миллионов лет назад. [ 3 ] [ 8 ] [ 17 ] Образования аккумулировались в разорванной системе форландов , характеризующейся несколькими бассейнами , отсоединенными друг от друга. Формация сложена в основном вулканокластическими известняками и песчаниками , накопившимися в континентальных условиях от аллювиальных (отложившихся проточной водой) до озерных (отложившихся озерами). [ 18 ]
Формация Коллон Кура и эпоха Коллонкура в Южной Америке представляют собой время, когда преобладали более открытые среды с уменьшенным растительным покровом, похожие на полузасушливые и умеренные или теплые, сухие леса или кустарники . Открытая среда позволяла обитать более беглым (приспособленным к бегу) и крупным животным, в отличие от более ранних условий позднего раннего миоцена с хорошо развитыми лесами с древесными животными. В то время леса были бы ограничены долинами горных хребтов Кордильер с небольшим количеством древесных видов. Это изменение происходило постепенно на раннем фризском этапе. [ 19 ] [ 20 ] Переход к более засушливым ландшафтам должен был произойти одновременно с изменениями климата , которые соответствовали климатическому переходу в среднем миоцене , глобальному похолоданию , которое оказало эффект высыхания континентов. [ 20 ]
The Collón Curá Formation of Argentina has provided a wide assemblage of mammals, including at least 24 taxa such as the xenarthrans Megathericulus , Prepotherium , Prozaedyus , and Paraeucinepeltus , the notoungulate Protypotherium , the astrapothere Astrapotherium , the sparassodonts Patagosmilus and Cladosictis , the marsupial Abderites , примат , Proteropithecia и грызуны такие как Maruchito , Protacaremys , Neoreomys и Prolagostomus . [ 17 ] [ 18 ] [ 21 ] [ 22 ] Помимо млекопитающих, характерных для отложений этого возраста, есть также несколько окаменелостей птиц, рептилий, амфибий и рыб. [ 19 ]
Ссылки
[ редактировать ]- ^ Jump up to: а б с Уилфорд, Джон Ноубл (31 октября 2006 г.). «Найдены окаменелости большой птицы, которые Кермиту не понравились бы» . Нью-Йорк Таймс . Проверено 26 мая 2022 г.
- ^ Хотц, Роберт Ли (26 октября 2006 г.). «Подросток нашел ископаемый череп самой большой известной птицы» . Питтсбург Пост-Газетт . п. 13 . Проверено 26 мая 2022 г.
- ^ Jump up to: а б с д и ж г час я дж к л м н тот п д Бертелли, Сара; Кьяппе, Луис М.; Тамбусси, Клаудия (2007). «Новый форусрацид (Aves: Cariamae) из среднего миоцена Патагонии, Аргентина» . Журнал палеонтологии позвоночных . 27 (2): 409–419. doi : 10.1671/0272-4634(2007)27[409:ANPACF]2.0.CO;2 . S2CID 85693135 .
- ^ Jump up to: а б с д и Кьяппе, Луис М.; Бертелли, Сара (2006). «Морфология черепа гигантских птиц-террористов» . Природа . 443 (7114): 929. дои : 10.1038/443929a . ПМИД 17066027 . S2CID 4381103 .
- ^ Jump up to: а б с Альваренга, Эркулано МФ; Хёфлинг, Элизабет (2003). «Синтетический обзор Phorusrhacidae (Aves: Ralliformes)» . Отдельные статьи по зоологии . 43 (4). дои : 10.1590/S0031-10492003000400001 .
- ^ Jump up to: а б с Альваренга, Геркулано; Кьяппе, Луис; Бертелли, Сара (2011). «Форусрациды: птицы ужаса». В Дайке, Гарет; Кайзер, Гэри (ред.). Живые динозавры . стр. 187–208. дои : 10.1002/9781119990475.ch7 . ISBN 978-1-119-99047-5 .
- ^ Дегранж, Федерико Дж.; Тамбусси, Клаудия П.; Тальоретти, Матиас Л.; Дондас, Алехандро; Скалья, Фернандо (2015). «Новый вид Mesembriornithinae (Aves, Phorusrhacidae) дает новое представление о филогении и сенсорных возможностях птиц-страшителей» . Журнал палеонтологии позвоночных . 35 (2): e912656. Бибкод : 2015JVPal..35E2656D . дои : 10.1080/02724634.2014.912656 . hdl : 11336/38650 . S2CID 85212917 .
- ^ Jump up to: а б с д Ксепка, Дэниел Т. (2014). «Полёты фантазии в эволюции птиц». Американский учёный . 102 (1): 39. дои : 10.1511/2014.106.36 . ISSN 0003-0996 . JSTOR 43707746 .
- ^ Jump up to: а б с д Дегранж, Федерико Дж. (2020). «Пересмотр морфологии черепа Phorusrhacidae (Aves, Cariamiformes)» . Журнал палеонтологии позвоночных . 40 (6): e1848855. Бибкод : 2020JVPal..40E8855D . дои : 10.1080/02724634.2020.1848855 . S2CID 234119602 .
- ^ Тамбусси, Клаудия; Убилла, Мартин; Переа, Дэниел (1999). «Самая молодая крупная хищная птица (Phorusrhacidae, Phorusrhacinae) Южной Америки (плиоцен-ранний плейстоцен Уругвая)» . Журнал палеонтологии позвоночных . 19 (2): 404–406. Бибкод : 1999JVPal..19..404T . дои : 10.1080/02724634.1999.10011154 . ISSN 0272-4634 . JSTOR 4524003 .
- ^ Тамбусси, Клаудия П.; Дегранж, Федерико Дж. (2013). «Неогеновые птицы Южной Америки». Южноамериканские и антарктические континентальные кайнозойские птицы . Springer Briefs по наукам о системе Земли. Спрингер. стр. 59–86. дои : 10.1007/978-94-007-5467-6_7 . ISBN 978-94-007-5466-9 .
- ^ Jump up to: а б Дегранж, Федерико Дж.; Тамбусси, Клаудия П.; Морено, Карен; Уитмер, Лоуренс М.; Роу, Стивен (2010). «Механический анализ пищевого поведения вымершей «террористической птицы» Andalgalornis steulleti (Gruiformes: Phorusrhacidae)» . ПЛОС ОДИН . 5 (8): е11856. Бибкод : 2010PLoSO...511856D . дои : 10.1371/journal.pone.0011856 . ПМЦ 2923598 . ПМИД 20805872 .
- ^ Jump up to: а б Бланко, Рудемар Эрнесто; Джонс, Вашингтон В. (2005). «Ужасающие птицы в бегах: механическая модель для оценки максимальной скорости бега» . Труды Королевского общества B: Биологические науки . 272 (1574): 1769–1773. дои : 10.1098/rspb.2005.3133 . ПМЦ 1559870 . ПМИД 16096087 .
- ^ Хадсон, Пенни Э.; Корр, Сандра А.; Уилсон, Алан М. (2012). «Скорый галоп у гепарда ( Acinonyx jubatus ) и беговой борзой ( Canis familiris ): пространственно-временные и кинетические характеристики» . Журнал экспериментальной биологии . 215 (14): 2425–2434. дои : 10.1242/jeb.066720 . ПМИД 22723482 . S2CID 13543638 .
- ^ Джойс, Кристофер (25 октября 2006 г.). «Огромная окаменелость «птицы ужаса» обнаружена в Патагонии» . NPR.org . Проверено 26 мая 2022 г.
- ^ Тамбусси, Клаудия П.; из Мендосы, Ричард; Дегранж, Фредерик Дж.; Пикассо, Мариана Б. (2012). «Гибкость шеи неогеновой ужасной птицы Andalgalornis steulleti (Aves Phorusrhacidae)» . ПЛОС ОДИН 7 (5): e37701. Бибкод : 2012PLoSO... 737701T дои : 10.1371/journal.pone.0037701 . ПМК 3360764 . ПМИД 22662194 .
- ^ Jump up to: а б Крамарц, Алехандро; Гарридо, Альберто; Бонд, Мариано (2019). « Астрапотерий из формации Коллон-Кура среднего миоцена и упадок астрапотериев на юге Южной Америки». Амегиниана . 56 (4): 290. doi : 10.5710/AMGH.15.07.2019.3258 . S2CID 199099778 .
- ^ Jump up to: а б Брандони, Диего; Руис, Лорен Гонсалес; Бучер, Хоакин (2020). «Эволюционные последствия Megathericulus patagonicus (Xenarthra, Megatheriinae) из миоцена Патагонии, Аргентина» Журнал эволюции млекопитающих . 27 (3): 445–460. дои : 10.1007/ s10914-019-09469-6 S2CID 163164163 .
- ^ Jump up to: а б Тонни, Эдуардо П.; Карлини, Альфредо А. (2008). «Неогеновые позвоночные из аргентинской Патагонии: их связь с наиболее значительными климатическими изменениями». В Рабассе, Хорхе (ред.). Поздний кайнозой Патагонии и Огненной Земли . Эльзевир Наука. стр. 269–278. ISBN 978-0-08-055889-9 .
- ^ Jump up to: а б Генисе, Хорхе Ф.; Беллози, Эдуардо С.; Кантиль, Лилиана Ф.; Гонсалес, Мирта Г.; Пуэрта, Пол (2022). «Переход климата в среднем миоцене, отраженный изменениями в ихнофациях и палеопочвах из Патагонии, Аргентина». Палеогеография, Палеоклиматология, Палеоэкология . 594 : 110932. Бибкод : 2022PPP...59410932G . дои : 10.1016/j.palaeo.2022.110932 . S2CID 247423841 .
- ^ Кей, Ричард Фредерик; Джонсон, Дерек; Мелдрам, Дон Джеффри (1998). «Новый примат питеций из среднего миоцена Аргентины» . Американский журнал приматологии . 45 (4): 317–336. doi : 10.1002/(SICI)1098-2345(1998)45:4<317::AID-AJP1>3.0.CO;2-Z . ПМИД 9702279 . S2CID 22214720 .
- ^ Эчарри, Себастьян; Уллоа-Гуайкин, Карен С.; Агирресабала, Гильермо; Форасиепи, Аналия М. (2021). « Cladosictis patagonica (Metatheria, Sparassodonta) из формации Коллон Кура (средний миоцен), Рио-Негро, Аргентина». Амегиниана . 58 (6). дои : 10.5710/AMGH.06.08.2021.3439 . hdl : 11336/171288 . S2CID 240529252 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Келенкеном, на Викискладе?
СМИ, связанные с Келенкеном, на Викискладе?
Роды Серимас и их вымершие союзники |
|---|


