неандерталец
| неандерталец Временной диапазон: от среднего до позднего плейстоцена, | |
|---|---|
 | |
| Примерная реконструкция скелета неандертальца. Центральная грудная клетка (включая грудину) и части таза принадлежат современному человеку. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Приматы |
| Подотряд: | Хаплорини |
| Инфрапорядок: | Симииформы |
| Семья: | Гоминиды |
| Подсемейство: | Люди |
| Племя: | Люди |
| Род: | гомо |
| Разновидность: | † H. neanderthalensis |
| Биномиальное имя | |
| † Человек неандертальский Кинг , 1864 г. | |
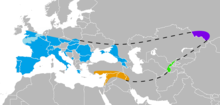 | |
| Известный ареал неандертальцев в Европе (синий), Юго-Западной Азии (оранжевый), Узбекистане (зеленый) и Горном Алтае (фиолетовый). | |
| Синонимы [6] | |
гомо Палеоантроп Протантроп Акантроп | |
Неандертальцы ( / n i ˈ æ n d ər ˌ t ɑː l , n eɪ -, - ˌ θ ɑː l / nee- AN -də(r)- TAHL , nay-, - THAHL ; [7] Homo neanderthalensis или H. sapiens neanderthalensis ) — вымершая группа архаичных людей (обычно рассматриваемых как отдельный вид, хотя некоторые считают ее подвидом Homo sapiens ), живших в Евразии примерно 40 000 лет назад. [8] [9] [10] [11] Типовой экземпляр , Neanderthal 1 , был найден в 1856 году в долине Неандер на территории современной Германии .
Неясно, когда линия неандертальцев отделилась от линии современных людей ; исследования дали в разное время от 315 000 [12] до более чем 800 000 лет назад. [13] дата отделения неандертальцев от их предка H. heidelbergensis Неясна и . Самые старые потенциальные кости неандертальцев датируются 430 000 лет назад, но классификация остается неопределенной. [14] Неандертальцы известны по многочисленным окаменелостям, особенно датированным 130 000 лет назад. [15]
Причины вымирания неандертальцев спорны. [16] [17] Теории их исчезновения включают демографические факторы, такие как небольшой размер популяции и инбридинг, конкурентное замещение, [18] скрещивание и ассимиляция с современным человеком, [19] изменение климата, [20] [21] [22] болезнь, [23] [24] или сочетание этих факторов. [22]
На протяжении большей части начала 20 века европейские исследователи изображали неандертальцев примитивными, неразумными и жестокими. Хотя с тех пор знания и восприятие их в научном сообществе заметно изменились, образ неразвитого пещерного человека архетипа по-прежнему преобладает в массовой культуре. [25] [26] По правде говоря, технология неандертальцев была весьма сложной. Включает мустьерскую каменно-орудийную промышленность. [27] [28] а также способности создавать огонь , [29] [30] строить пещерные очаги [31] [32] (готовить еду, согреваться, защищаться от животных, размещая ее в центре своего жилища), [33] сделать клей из бересты , [34] смастерите хотя бы простую одежду, похожую на одеяла и пончо, [35] ткать, [36] отправиться в мореплавание по Средиземному морю, [37] [38] использовать лекарственные растения , [39] [40] [41] лечить тяжелые травмы, [42] хранить еду, [43] и использовать различные методы приготовления, такие как запекание , варка , [44] и курение . [45] Неандертальцы потребляли разнообразную пищу, в основном копытных млекопитающих . [46] но и мегафауна , [25] [47] растения, [48] [49] [50] мелкие млекопитающие, птицы, а также водные и морские ресурсы. [51] Хотя они, вероятно, были высшими хищниками , они все же конкурировали с пещерными львами , пещерными гиенами и другими крупными хищниками. [52] Ряд примеров символической мысли и палеолитического искусства были безрезультатно изучены. [53] приписывают неандертальцам, а именно возможные украшения из птичьих когтей и перьев, [54] [55] ракушки, [56] коллекции необычных предметов, включая кристаллы и окаменелости, [57] гравюры, [58] музыкальное производство (возможно, обозначено флейтой Дивье Бабе ), [59] и испанские наскальные рисунки вызывают споры [60] датируется примерно 65 000 лет назад. [61] [62] Были сделаны некоторые заявления о религиозных убеждениях. [63] Неандертальцы, вероятно, были способны к речи, возможно, членораздельной, хотя сложность их языка неизвестна. [64] [65]
По сравнению с современными людьми неандертальцы имели более крепкое телосложение и пропорционально более короткие конечности. Исследователи часто объясняют эти особенности адаптацией к сохранению тепла в холодном климате, но они также могли быть адаптацией к бегу на короткие дистанции в более теплом лесном ландшафте, который часто населяли неандертальцы. [66] У них были приспособления, специфичные для холода, такие как специализированное накопление жира в организме. [67] и увеличенный нос для теплого воздуха [68] (хотя нос мог быть вызван генетическим дрейфом [69] ). Средний рост мужчин-неандертальцев составлял около 165 см (5 футов 5 дюймов), а женщин — 153 см (5 футов 0 дюймов), что соответствовало доиндустриальным современным европейцам. [70] Размер черепной коробки неандертальцев мужчин и женщин составлял в среднем около 1600 см3. 3 (98 куб. Дюймов) и 1300 см 3 (79 у.е. дюймов) соответственно [71] [72] [73] что значительно больше, чем в среднем у современного человека (1260 см). 3 (77 куб. Дюймов) и 1130 см. 3 (69 у.е. дюймов) соответственно). [74] Череп неандертальца был более вытянутым, а теменные доли мозга были меньше. [75] [76] [77] и мозжечок, [78] [79] но более крупные височные, затылочные и орбитофронтальные области. [80] [81]
Общая популяция неандертальцев оставалась низкой, распространяя слабовредные варианты генов. [82] и препятствуют созданию эффективных сетей дальней связи. Несмотря на это, есть свидетельства существования региональных культур и регулярного общения между сообществами. [83] [84] Возможно, они часто посещали пещеры и перемещались между ними сезонно. [85] Неандертальцы жили в условиях сильного стресса и высокого уровня травматизма, и около 80% из них умерли в возрасте до 40 лет. [86]
2010 года В проекте отчета проекта генома неандертальцев представлены доказательства скрещивания неандертальцев и современных людей . [87] [88] [89] Возможно, это произошло от 316 000 до 219 000 лет назад. [90] но более вероятно 100 000 лет назад и снова 65 000 лет назад. [91] Неандертальцы также, по-видимому, скрещивались с денисовцами , другой группой архаичных людей, в Сибири. [92] [93] Около 1–4% геномов евразийцев , коренных австралийцев , меланезийцев , коренных американцев и жителей Северной Африки имеют неандертальское происхождение, в то время как большинство жителей стран Африки к югу от Сахары имеют около 0,3% генов неандертальцев, за исключением возможных следов от ранних сапиенсов до Поток генов неандертальцев и/или недавняя обратная миграция евразийцев в Африку. В целом около 20% явно неандертальских вариантов генов выживают у современных людей. [94] Хотя многие из вариантов генов, унаследованных от неандертальцев, возможно, были вредными и были исключены, [82] неандертальцев, Интрогрессия по-видимому, повлияла на иммунную систему современного человека . [95] [96] [97] [98] а также участвует в ряде других биологических функций и структур, [99] но большая часть, по-видимому, представляет собой некодирующую ДНК . [100]
Таксономия
[ редактировать ]Этимология
[ редактировать ]
Неандертальцы названы в честь долины Неандер , в которой был найден первый идентифицированный экземпляр. Долина называлась Neanderthal назывался Neanderthaler , а вид на немецком языке до реформы правописания 1901 года . [б] Написание этого вида «неандерталец» иногда встречается в английском языке, даже в научных публикациях, но научное название H. neanderthalensis всегда пишется через th в соответствии с принципом приоритета . Народное название вида на немецком языке всегда Neandertaler («обитатель долины Неандер»), тогда как неандерталец всегда относится к долине. [с] Сама долина была названа в честь немецкого богослова и автора гимнов конца 17 века Иоахима Неандера , который часто посещал этот район. [101] Его имя, в свою очередь, означает «новый человек», являясь учёной грецизацией немецкой фамилии Нейман .
Неандерталец можно произносить с помощью /t/ (как в / n i ˈ æ n d ər t ɑː l / ) [104] или стандартное английское произношение th с фрикативным звуком / θ / (as / n i ˈ æ n d ər θ ɔː l / ). [105] [106]
Неандерталец 1 , типовой экземпляр , был известен в антропологической литературе как «череп неандертальца» или «череп неандертальца», а человека, реконструированного на основе черепа, иногда называли «неандертальцем». [107] Биномиальное название Homo neanderthalensis , расширяющее название «неандерталец» от отдельного экземпляра до всего вида и формально признающее его отличие от человека, было впервые предложено ирландским геологом Уильямом Кингом в статье, прочитанной на 33-й сессии Британской научной ассоциации в 1863. [108] [109] [110] Однако в 1864 году он рекомендовал отнести неандертальцев и современных людей к разным родам, сравнивая черепную коробку неандертальцев с черепной коробкой шимпанзе и утверждая, что они «неспособны к моральным и [ теистическим [д] ] концепции». [111]

История исследований
[ редактировать ]
Первые останки неандертальца — Энжис 2 (череп) — были обнаружены в 1829 году голландско-бельгийским доисториком Филиппом-Шарлем Шмерлингом в Грот-д'Энгис , Бельгия. Он пришел к выводу, что эти «слабо развитые» человеческие останки, должно быть, были захоронены в то же время и по тем же причинам, что и сосуществовавшие останки вымерших видов животных. [112] В 1848 году Гибралтар 1 из карьера Форбса был подарен Научному обществу Гибралтара его секретарем лейтенантом Эдмундом Генри Рене Флинтом, но считался черепом современного человека. [113] В 1856 году местный школьный учитель Иоганн Карл Фульротт распознал кости из пещеры Кляйне Фельдхофер в долине Неандер — неандертальца 1 ( образец голотипа ) — в отличие от костей современных людей. [и] и передал их немецкому антропологу Герману Шаафхаузену для изучения в 1857 году. Он включал череп, бедренные кости, правую руку, левую плечевую и локтевую кости , левую подвздошную кость (тазовую кость), часть правой лопатки и части ребер . [111] [114] Следуя Чарльза Дарвина книге «Происхождение видов» , Фульротт и Шаафхаузен утверждали, что кости представляют собой древнюю современную человеческую форму; [26] [111] [115] [116] Шаафхаузен, социальный дарвинист , считал, что люди линейно эволюционировали от дикарей к цивилизованным, и поэтому пришел к выводу, что неандертальцы были варварскими пещерными жителями. [26] Фульротт и Шаафхаузен встретили сопротивление, в частности, со стороны плодовитого патологоанатома Рудольфа Вирхова , который выступал против определения новых видов на основе только одной находки. В 1872 году Вирхов ошибочно истолковал характеристики неандертальцев как свидетельство старости , болезней и уродств, а не архаичности. [117] что остановило исследования неандертальцев до конца века. [26] [115]
К началу 20 века было сделано множество других открытий неандертальцев, сделавших H. neanderthalensis законным видом. Самым влиятельным экземпляром был La Chapelle-aux-Saints 1 («Старик») из Ла-Шапель-о-Сент , Франция. Французский палеонтолог Марселлен Буль является автором нескольких публикаций, одной из первых, кто утвердил палеонтологию как науку, подробно описав образец, но реконструировал его как сутулого, обезьяноподобного и лишь отдаленно связанного с современными людьми. «Открытие» пилтдаунского человека в 1912 году (мистификация), который выглядел гораздо более похожим на современных людей, чем на неандертальцев, было использовано в качестве доказательства существования множества различных и несвязанных между собой ветвей примитивных людей и поддержало реконструкцию Буля H. neanderthalensis как очень далекого вида. относительный и эволюционный тупик . [26] [118] [119] [120] Он способствовал распространению популярного образа неандертальцев как варварских, сутулящихся, владеющих дубинками примитивов; этот образ воспроизводился в течение нескольких десятилетий и популяризировался в научно-фантастических произведениях, таких как « В поисках огня» года Ж.-Х. 1911 Росни Айне и «Ужасный народ» Герберта Уэллса 1927 года , в котором они изображены в виде монстров. [26] В 1911 году шотландский антрополог Артур Кейт реконструировал Ла-Шапель-о-Сент-1 как непосредственного предшественника современных людей, сидящих рядом с огнем, производящих инструменты, носящих ожерелье и имеющих более человеческую позу, но это не помогло собрать много научных данных. взаимопонимание, и Кейт позже отказался от своей диссертации в 1915 году. [26] [115] [121]
К середине столетия, основываясь на разоблачении Пилтдаунского человека как мистификации, а также на повторном обследовании Ла Шапель-о-Сент-1 (у которого был остеоартрит , из-за которого он сутулился в жизни) и новых открытиях, научное сообщество начало перерабатывать его понимание неандертальцев. Обсуждались такие идеи, как поведение, интеллект и культура неандертальцев, и возник их более человеческий образ. В 1939 году американский антрополог Карлтон Кун реконструировал неандертальца в современном деловом костюме и шляпе, чтобы подчеркнуть, что они были бы более или менее неотличимы от современных людей, если бы дожили до наших дней. В романе Уильяма Голдинга » 1955 года «Наследники неандертальцы изображены гораздо более эмоциональными и цивилизованными. [25] [26] [120] Однако образ Буля продолжал влиять на произведения вплоть до 1960-х годов. В наши дни реконструкции неандертальцев часто очень похожи на человеческие. [115] [120]
−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
Гибридизация между неандертальцами и ранними современными людьми была предложена еще раньше. [122] например, английский антрополог Томас Хаксли в 1890 году, [123] Датский этнограф Ганс Педер Стенсби в 1907 году. [124] и Кун в 1962 году. [125] В начале 2000-х годов были обнаружены предполагаемые гибридные экземпляры: Lagar Velho 1. [126] [127] [128] [129] и Муиерии 1 . [130] Однако подобная анатомия могла быть вызвана адаптацией к сходной среде, а не скрещиванием. [100] Примесь неандертальцев была обнаружена в современных популяциях в 2010 году при картировании первой последовательности генома неандертальца. [87] Это было основано на трех образцах из пещеры Виндия в Хорватии, которые содержали почти 4% архаичной ДНК (что позволяет почти полностью секвенировать геном). ) приходилось примерно 1 ошибка Однако на каждые 200 букв ( пар оснований из-за невероятно высокой частоты мутаций, вероятно, из-за сохранности образца. В 2012 году британско-американский генетик Грэм Куп выдвинул гипотезу, что вместо этого они обнаружили доказательства скрещивания различных архаичных видов человека с современными людьми, что было опровергнуто в 2013 году секвенированием высококачественного генома неандертальца, сохранившегося в кости пальца ноги из Денисовой пещеры. Сибирь. [100]
Классификация
[ редактировать ]
| |||||||||||||||||||||||||||
| Филогения 2019 г. основана на сравнении древних протеомов и геномов с протеомами и геномами современных видов. [131] |
Неандертальцы — гоминиды рода Homo Homo , люди, и обычно классифицируются как отдельный вид , H. neanderthalensis , хотя иногда и как подвид современного человека, как sapiens neanderthalensis . Это потребовало бы классификации современных людей как H. sapiens sapiens . [132]
Большая часть разногласий проистекает из расплывчатости термина «вид», поскольку он обычно используется для различения двух генетически изолированных популяций, но известно, что смешение между современными людьми и неандертальцами имело место. [132] [133] Однако отсутствие отцовской Y-хромосомы и матрилинейной митохондриальной ДНК (мтДНК) неандертальского происхождения у современных людей, а также недостаточная представленность ДНК Х-хромосомы неандертальцев может означать снижение плодовитости или частую стерильность некоторых гибридных скрещиваний. [89] [134] [135] [136] представляющий собой частичный биологический репродуктивный барьер между группами и, следовательно, видовое различие. [89] В 2014 году генетик Сванте Пэабо резюмировал противоречие, назвав такие « таксономические войны» неразрешимыми, «поскольку не существует определения вида, идеально описывающего этот случай». [132]
Считается, что неандертальцы были более тесно связаны с денисовцами, чем с современными людьми. Точно так же неандертальцы и денисовцы имеют более позднего последнего общего предка (LCA), чем современные люди, на основе ядерной ДНК (нДНК). Однако неандертальцы и современные люди имеют общий более поздний митохондриальный LCA (наблюдаемый при изучении мтДНК) и LCA Y-хромосомы. [137] Вероятно, это произошло в результате скрещивания, последовавшего за расколом неандертальцев и денисовцев. Это включало либо интрогрессию неизвестного архаичного человека в денисовцев, либо [92] [93] [131] [138] [139] или интрогрессия более ранней неопознанной современной человеческой волны из Африки в неандертальцев. [137] [140] [141] Тот факт, что мтДНК раннего архаического человека неандертальской линии возрастом около 430 000 лет из Сима-де-лос-Уэсос в Испании более тесно связана с мтДНК денисовцев, чем с другими неандертальцами или современными людьми, был приведен в качестве доказательства в пользу последней гипотезы. . [137] [14] [140]
Эволюция
[ редактировать ]



Принято считать, что H. heidelbergensis был последним общим предком неандертальцев, денисовцев и современных людей до того, как популяции были изолированы в Европе, Азии и Африке соответственно. [143] Таксономическое различие между H. heidelbergensis и неандертальцами в основном основано на разрыве в окаменелостях в Европе между 300 и 243 000 лет назад, во время 8-й стадии морских изотопов . «Неандертальцами» принято называть окаменелости, датируемые после этого разрыва. [12] [25] [142] ДНК архаичных людей из 430 000-летнего поселения Сима-де-лос-Уэсос в Испании указывает на то, что они более тесно связаны с неандертальцами, чем с денисовцами, а это указывает на то, что раскол между неандертальцами и денисовцами должен был произойти еще до этого времени. [14] [144] [145] возрастом 400 000 лет Череп Ароэйры 3 также может представлять собой одного из ранних представителей линии неандертальцев. [146] Вполне возможно, что поток генов между Западной Европой и Африкой во время среднего плейстоцена мог скрыть неандертальские характеристики у некоторых экземпляров европейских гомининов среднего плейстоцена, например, из Чепрано , Италия, и ущелья Сичево , Сербия. [14] Летопись окаменелостей гораздо более полна, начиная с 130 000 лет назад. [147] и экземпляры этого периода составляют основную часть известных скелетов неандертальцев. [148] [149] Остатки зубов из итальянских стоянок Висольяно и Фонтана-Рануччо указывают на то, что особенности зубов неандертальцев развились примерно 450–430 000 лет назад во время среднего плейстоцена . [150]
Существуют две основные гипотезы относительно эволюции неандертальцев после разделения неандертальцев и человека: двухфазная и аккреционная. Двухфазный утверждает, что одно-единственное крупное экологическое событие, такое как оледенение Заале , привело к быстрому увеличению размера и прочности тела европейского H. heidelbergensis , а также к удлинению головы (фаза 1), что затем привело к другим изменения анатомии черепа (2 этап). [128] Однако анатомия неандертальцев, возможно, не была полностью обусловлена адаптацией к холодной погоде. [66] что неандертальцы медленно эволюционировали с течением времени от предка H. heidelbergensis , разделенного на четыре стадии: ранние пренеандертальцы ( MIS 12 , Эльстерское оледенение ), пренеандертальцы (MIS 11–9 MIS Аккреция утверждает , , голштинское межледниковье ), ранние неандертальцы ( 7–5 ), и , оледенение Заале – эмское классические неандертальцы (МИС 4–3, оледенение Вюрма ). [142]
Было предложено множество дат разделения неандертальцев и людей. Дата около 250 000 лет назад указывает на то, что « H. Helmei » был последним общим предком (LCA), а раскол связан с леваллуазской техникой изготовления каменных орудий. Для даты около 400 000 лет назад используется H. heidelbergensis в качестве LCA . По оценкам, сделанным 600 000 лет назад, предполагается, что « H. rhodesiensis » был LCA, который разделился на современную человеческую линию и линию неандертальцев/ H. heidelbergensis . [151] Восемьсот тысяч лет назад в качестве LCA использовался H. antecessor , но различные варианты этой модели отодвинули бы дату на 1 миллион лет назад. [14] [151] Однако анализ H. antecessor, эмали протеомов проведенный в 2020 году, предполагает, что H. antecessor является родственным, но не прямым предком. [152] Исследования ДНК дали различные результаты для времени расхождения неандертальцев и человека, например, 538–315, [12] 553–321, [153] 565–503, [154] 654–475, [151] 690–550, [155] 765–550, [14] [92] 741–317, [156] и 800–520 000 лет назад; [157] и стоматологический анализ завершился более 800 000 лет назад. [13]
Неандертальцы и денисовцы более тесно связаны друг с другом, чем с современными людьми, а это означает, что раскол неандертальцев и денисовцев произошел после их раскола с современными людьми. [14] [92] [138] [158] Предполагая частоту мутаций 1 × 10 −9 или 0,5 × 10 −9 на пару оснований (п.н.) в год раскол неандертальцев и денисовцев произошел примерно 236–190 000 или 473–381 000 лет назад соответственно. [92] Использование 1,1 × 10 −8 на поколение, новое поколение каждые 29 лет, время — 744 000 лет назад. Использование 5 × 10 −10 нуклеотидных сайтов в год, это 616 000 лет назад. Если использовать последние даты, раскол, вероятно, уже произошел к тому времени, когда гоминиды распространились по Европе, а уникальные черты неандертальцев начали развиваться 600–500 000 лет назад. [138] Перед расколом неандертальцы/денисовцы (или «неандерсовцы»), мигрировавшие из Африки в Европу, по-видимому, скрещивались с неопознанным «суперархаичным» человеческим видом, который уже присутствовал там; эти суперархаики были потомками очень ранней миграции из Африки около 1,9 млн лет назад. [159]
Демография
[ редактировать ]Диапазон
[ редактировать ]
Пре- и ранние неандертальцы, жившие до эемского межледниковья (130 000 лет назад), мало известны и происходят в основном из западноевропейских стоянок. Начиная со 130 000 лет назад, качество летописи окаменелостей резко возрастает с появлением классических неандертальцев, которые зарегистрированы в Западной, Центральной, Восточной и Средиземноморской Европе. [15] а также Юго-Западную , Центральную и Северную Азию до Горного Алтая на юге Сибири. С другой стороны, пре- и ранние неандертальцы, по-видимому, постоянно оккупировали только Францию, Испанию и Италию, хотя некоторые из них, по-видимому, покинули эту «основную территорию», чтобы сформировать временные поселения на востоке (хотя и не покидая Европу). Тем не менее, на юго-западе Франции самая высокая плотность стоянок до-, ранних и классических неандертальцев. [160] Неандертальцы были первым человеческим видом, постоянно населявшим Европу, поскольку более ранние люди на континенте были лишь время от времени. [161]
Самая южная находка была зафиксирована в пещере Шукба , Левант; [162] сообщения о неандертальцах из североафриканского Джебель-Ирхуда [163] и Хауа Фтеах [164] были повторно идентифицированы как H. sapiens . Их самое восточное присутствие зафиксировано в Денисовой пещере , Сибирь, 85° в.д .; Юго-восточный китайский человек Маба , череп, имеет некоторые общие физические признаки с неандертальцами, хотя они могут быть результатом конвергентной эволюции, а не расширения неандертальцами своего ареала до Тихого океана. [165] Самая северная граница обычно считается 55° с.ш. , а однозначные места известны между 50–53 , хотя это трудно оценить, поскольку наступление ледников уничтожает большую часть ° с.ш. человеческих останков, а палеоантрополог Трине Келлберг Нильсен утверждает, что отсутствие доказательств Заселение южных скандинавов (по крайней мере, во время эемского межледниковья) связано с первым объяснением и отсутствием исследований в этом районе. [166] [167] Артефакты среднего палеолита обнаружены до 60° с.ш. на равнинах России. [168] [169] [170] но они, скорее всего, приписываются современным людям. [171] Исследование 2017 года показало присутствие Homo возрастом 130 000 лет на территории калифорнийского мастодонта Черутти в Северной Америке. [172] но это во многом считается неправдоподобным. [173] [174] [175]
Неизвестно, как быстро меняющийся климат последнего ледникового периода ( события Дансгаарда-Эшгера ) повлиял на неандертальцев, поскольку периоды потепления приводили к более благоприятным температурам, но способствовали росту лесов и сдерживали мегафауну, тогда как холодные периоды приводили к противоположному результату. [176] Однако неандертальцы, возможно, предпочитали лесной ландшафт. [66] Стабильная среда с умеренными среднегодовыми температурами, возможно, была наиболее подходящей средой обитания для неандертальцев. [177] Пик численности популяций мог достигаться в холодные, но не экстремальные периоды, например, на 8-й и 6-й стадиях морских изотопов (соответственно 300 000 и 191 000 лет назад во время оледенения Заале). Возможно, их ареал расширялся и сужался по мере отступания и роста льда, соответственно, чтобы избежать зон вечной мерзлоты , находящихся в определенных зонах убежища во время максимумов ледников. [176] возрастом от 140 до 120 000 лет В 2021 году израильский антрополог Исраэль Гершковитц и его коллеги предположили, что останки израильского Нешера Рамлы , в которых сочетаются черты неандертальца и более древних черт H. erectus , представляют собой одну из таких исходных популяций, которые повторно заселили Европу после ледникового периода. . [178]

Население
[ редактировать ]Как и современные люди, неандертальцы, вероятно, произошли от очень небольшой популяции с эффективной популяцией (количеством особей, способных выносить или стать отцом детей) примерно от 3000 до 12 000 человек. Однако неандертальцы сохранили эту очень низкую популяцию, размножая слабо вредные гены из-за снижения эффективности естественного отбора . [82] [179] Различные исследования с использованием анализа мтДНК дают разные эффективные популяции. [176] например, от около 1000 до 5000; [179] от 5000 до 9000 остается постоянным; [180] или от 3000 до 25 000, которые постепенно увеличивались до 52 000 лет назад, а затем начали сокращаться вплоть до исчезновения. [84] Археологические данные свидетельствуют о десятикратном увеличении численности современного человека в Западной Европе в период перехода от неандертальцев к современному человеку. [181] а неандертальцы, возможно, находились в невыгодном демографическом положении из-за более низкого уровня рождаемости, более высокого уровня детской смертности или комбинации этих двух факторов. [182] По оценкам, общая численность населения превышает десятки тысяч человек. [138] оспариваются. [179] Постоянно низкую численность населения можно объяснить в контексте « ловушки Бозерупа популяции »: пропускная способность ограничена количеством пищи, которую она может получить, что, в свою очередь, ограничено ее технологией. Инновации растут вместе с населением, но если население слишком мало, инновации не будут происходить очень быстро, и население останется низким. Это согласуется с очевидным 150 000-летним застоем неандертальской каменной технологии. [176]
В выборке из 206 неандертальцев, исходя из численности молодых и зрелых взрослых по сравнению с представителями других возрастных групп, около 80% из них в возрасте старше 20 лет умерли, не дожив до 40 лет. Этот высокий уровень смертности, вероятно, был обусловлен их сильным стрессом. среда. [86] Однако было также подсчитано, что возрастные пирамиды неандертальцев и современных современных людей были одинаковыми. [176] По оценкам, детская смертность у неандертальцев была очень высокой - около 43% в Северной Евразии. [183]


Анатомия
[ редактировать ]Строить
[ редактировать ]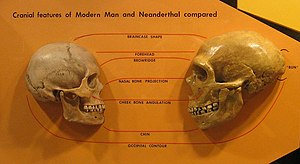
Неандертальцы имели более крепкое и коренастое телосложение, чем типичные современные люди. [70] более широкие и бочкообразные грудные клетки; более широкий таз; [25] [184] и пропорционально более короткие предплечья и передние ноги. [66] [185]
На основе 45 длинных костей неандертальцев , взятых у 14 мужчин и 7 женщин, средний рост составлял от 164 до 168 см (от 5 футов 5 дюймов до 5 футов 6 дюймов) для мужчин и от 152 до 156 см (от 5 футов 0 дюймов до 5 футов 1 дюйм). для женщин. [70] Для сравнения, средний рост 20 мужчин и 10 женщин людей верхнего палеолита составляет соответственно 176,2 см (5 футов 9,4 дюйма) и 162,9 см (5 футов 4,1 дюйма), хотя ближе к концу он уменьшается на 10 см (4 дюйма). за период на основе 21 мужчины и 15 женщин; [186] а средний показатель в 1900 году составлял 163 см (5 футов 4 дюйма) и 152,7 см (5 футов 0 дюймов) соответственно. [187] Летопись окаменелостей показывает, что рост взрослых неандертальцев варьировался от 147,5 до 177 см (от 4 футов 10 дюймов до 5 футов 10 дюймов), хотя некоторые из них могли вырасти намного выше (от 73,8 до 184,8 см в зависимости от длины следа и от 65,8 до 189,3 см). в зависимости от ширины следа). [188] Что касается веса неандертальцев, образцы из 26 экземпляров обнаружили в среднем 77,6 кг (171 фунт) для мужчин и 66,4 кг (146 фунтов) для женщин. [189] Используя 76 кг (168 фунтов), индекс массы тела мужчин-неандертальцев составил 26,9–28,2, что у современных людей коррелирует с избыточным весом . Это указывает на очень надежную конструкцию. [70] Ген неандертальского LEPR, отвечающий за накопление жира и выработку тепла телом, аналогичен гену шерстистого мамонта и поэтому, вероятно, был адаптацией к холодному климату. [67]

Шейные позвонки неандертальцев толще спереди назад и в поперечном направлении, чем у (большинства) современных людей, что приводит к стабильности и, возможно, позволяет приспособиться к другой форме и размеру головы. [190] неандертальца Хотя грудная клетка (там, где находится грудная клетка ) по размеру была аналогична размерам современных людей, более длинные и прямые ребра соответствовали расширенной средней и нижней части грудной клетки и более сильному дыханию в нижней части грудной клетки, что указывает на большую диафрагму и, возможно, на более крупную диафрагму. большая емкость легких . [184] [191] [192] Емкость легких Кебары 2 оценивалась в 9,04 л (2,39 галлона США) по сравнению со средней емкостью легких человека 6 л (1,6 галлона США) для мужчин и 4,7 л (1,2 галлона США) для женщин. Грудь неандертальца также была более выраженной (расширенной спереди назад или переднезадне). Крестец лордоз (там, где таз соединяется с позвоночником ) был более наклонен вертикально и располагался ниже по отношению к тазу, в результате чего позвоночник был менее изогнут (проявлялся меньший ) и несколько сгибался (чтобы быть инвагинированным). . В современных популяциях это состояние затрагивает лишь часть населения и известно как поясничный крестец. [193] Такие модификации позвоночника будут способствовать усилению бокового (медиолатерального) сгибания , что позволит лучше поддерживать более широкую нижнюю часть грудной клетки. Некоторые утверждают, что эта особенность была бы нормальной для всех Homo , даже адаптированных к тропическим условиям Homo ergaster или erectus , при этом более узкая грудная клетка у большинства современных людей является уникальной характеристикой. [184]
Пропорции тела обычно называют «гиперарктическими» как приспособление к холоду, поскольку они аналогичны пропорциям человеческих популяций, развившихся в холодном климате. [194] — телосложение неандертальцев наиболее похоже на телосложение инуитов и сибирских юпиков среди современных людей. [195] — а более короткие конечности приводят к более высокому сохранению тепла тела. [185] [194] [196] Тем не менее, неандертальцы из более умеренного климата, таких как Иберия, все еще сохраняют «гиперарктическое» телосложение. [197] В 2019 году английский антрополог Джон Стюарт и его коллеги предположили, что неандертальцы вместо этого были приспособлены к бегу на короткие дистанции, поскольку имеются доказательства того, что неандертальцы предпочитают более теплые лесные районы более холодным гигантским степям , а анализ ДНК указывает на более высокую долю быстросокращающихся мышечных волокон у неандертальцев, чем у современных. люди. Он объяснил их пропорции тела и большую мышечную массу адаптацией к спринтерскому бегу, в отличие от телосложения современного человека, ориентированного на выносливость . [66] поскольку настойчивая охота может быть эффективной только в жарком климате, где охотник может довести добычу до теплового истощения ( гипертермии ). У них были длиннее пяточные кости , [198] снижая их способность к бегу на выносливость, а их более короткие конечности имели бы уменьшенный момент силы в конечностях, что позволяло бы создавать большую чистую вращательную силу в запястьях и лодыжках, вызывая более быстрое ускорение. [66] В 1981 году американский палеоантрополог Эрик Тринкаус обратил внимание на это альтернативное объяснение, но посчитал его менее вероятным. [185] [199]
Лицо
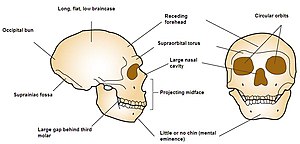
[ редактировать ]
У неандертальцев были менее развитые подбородки, покатые лбы и более длинные, широкие и выступающие носы. Череп неандертальца обычно более удлиненный, но при этом более широкий и менее шаровидный, чем у большинства современных людей, и имеет гораздо больше сходства с затылочным бугорком . [200] или «шиньон», выступ на задней части черепа, хотя у современных людей он находится в пределах вариаций. Это вызвано тем, что основание черепа и височные кости расположены выше и ближе к передней части черепа, а также более плоской черепной коробкой . [201]
Лицо неандертальца характеризуется прогнатизмом средней части лица , при котором скуловые дуги расположены назад по сравнению с современными людьми, в то время как их верхнечелюстные кости и носовые кости, по сравнению с ними, расположены более вперед. [202] Глазные яблоки неандертальцев крупнее, чем у современного человека. В одном исследовании было высказано предположение, что это произошло из-за того, что у неандертальцев были улучшены зрительные способности за счет неокортикального и социального развития. [203] Однако это исследование было отвергнуто другими исследователями, которые пришли к выводу, что размер глазного яблока не является доказательством когнитивных способностей неандертальцев или современных людей. [204]
Проецируемый неандертальский нос и околоносовые пазухи обычно объяснялись тем, что воздух нагревался при попадании в легкие и сохранял влагу (гипотеза «носового радиатора»); [205] если бы их носы были шире, это отличалось бы от обычно суженной формы у существ, адаптированных к холоду, и это было бы вызвано генетическим дрейфом . Кроме того, широко реконструированные пазухи не очень велики и сравнимы по размеру с таковыми у современного человека. Однако если размер пазух не является важным фактором для дыхания холодным воздухом, то фактическая функция будет неясна, поэтому они не могут быть хорошим индикатором эволюционного давления, необходимого для развития такого носа. [206] Кроме того, компьютерная реконструкция носа неандертальца и предсказанные структуры мягких тканей показывают некоторое сходство с носами современных арктических народов, что потенциально означает, что носы обеих популяций конвергентно эволюционировали для дыхания холодным и сухим воздухом. [68]
У неандертальцев была довольно большая челюсть, которая когда-то рассматривалась как реакция на большую силу укуса , о чем свидетельствует сильное изнашивание передних зубов неандертальцев (гипотеза «передней нагрузки на зубы»), но аналогичные тенденции изнашивания наблюдаются и у современных людей. Он также мог бы эволюционировать, чтобы соответствовать более крупным зубам в челюсти, которые лучше сопротивлялись бы износу и истиранию. [205] [207] а повышенный износ передних зубов по сравнению с задними, вероятно, связан с повторяющимся использованием. Образцы износа зубов неандертальцев больше всего похожи на таковые у современных инуитов. [205] Резцы большие и имеют лопатовидную форму, и, по сравнению с современными людьми, наблюдается необычно высокая частота тауродонтизма — состояния, при котором коренные зубы становятся более объемными из-за увеличенной пульпы (сердцевины зуба). Когда-то считалось, что тауродонтизм был отличительной чертой неандертальцев, которая давала некоторые механические преимущества или возникала в результате повторяющегося использования, но, скорее всего, была просто продуктом генетического дрейфа. [208] Сейчас считается, что сила укуса неандертальцев и современных людей примерно одинакова. [205] около 285 Н (64 фунта-силы) и 255 Н (57 фунтов-силы) у современных мужчин и женщин соответственно. [209]

Мозг
[ редактировать ]Размер черепной коробки неандертальца составляет в среднем 1640 см. 3 (100 у.е. дюймов) для мужчин и 1460 см. 3 (89 у.е. дюймов) для самок, [72] [73] что значительно превышает средние показатели для всех групп современного человека; [74] например, средний рост современных европейских мужчин составляет 1362 см. 3 (83,1 куб. Дюймов) и самки 1201 см. 3 (73,3 куб. Дюймов). [210] Для 28 современных человеческих экземпляров, живших от 190 000 до 25 000 лет назад, средний рост составлял около 1478 см. 3 (90,2 куб. Дюймов), независимо от пола, и предполагается, что размер мозга современного человека уменьшился со времен верхнего палеолита. [211] Самый большой мозг неандертальца, Амуд 1 , по расчетам, имел размер 1736 см. 3 (105,9 куб. Дюймов), один из крупнейших, когда-либо зарегистрированных у гоминид. [73] И неандертальцы, и человеческие младенцы имеют рост около 400 см. 3 (24 куб.дюйма). [212]
Если смотреть сзади, черепная коробка неандертальца выглядит ниже, шире и округлее, чем у анатомически современных людей. Эта характерная форма называется «en бомбе» (похожая на бомбу) и является уникальной для неандертальцев, при этом все другие виды гоминидов (включая большинство современных людей) обычно имеют узкие и относительно вертикальные своды черепа, если смотреть сзади. [213] [214] [215] [216] Мозг неандертальца характеризовался относительно меньшими теменными долями. [80] и больший мозжечок . [80] [217] Мозг неандертальцев также имеет более крупные затылочные доли (что связано с классическим появлением затылочного бугорка в анатомии черепа неандертальцев, а также с большей шириной их черепов), что подразумевает внутренние различия в пропорциональности внутренних областей мозга по сравнению с Homo sapiens. , что согласуется с внешними измерениями, полученными на ископаемых черепах. [203] [218] Их мозг также имеет более крупные полюса височных долей. [217] более широкая орбитофронтальная кора, [219] и более крупные обонятельные луковицы, [220] предполагая потенциальные различия в понимании языка и ассоциациях с эмоциями ( временные функции ), принятием решений ( орбитофронтальная кора ) и обонянием ( обонятельные луковицы ). Их мозг также демонстрирует разные темпы роста и развития. [221] Такие различия, хотя и незначительные, были бы заметны в ходе естественного отбора и могут лежать в основе и объяснять различия в материальных показателях в таких вещах, как социальное поведение, технологические инновации и художественные произведения. [18] [222]
Цвет волос и кожи
[ редактировать ]Недостаток солнечного света, скорее всего, привел к распространению более светлой кожи у неандертальцев. [223] хотя недавно утверждалось, что светлая кожа у современных европейцев не была особенно плодовитой, возможно, до бронзового века . [224] Генетически BNC2 присутствовал у неандертальцев, что связано со светлым цветом кожи; однако также присутствовал второй вариант BNC2, который в современных популяциях связан с более темным цветом кожи в Биобанке Великобритании . [223] Анализ ДНК трех женщин-неандертальцев из юго-восточной Европы показывает, что у них были карие глаза, темный цвет кожи и каштановые волосы, причем у одной из них были рыжие волосы. [225] [226]
У современных людей цвет кожи и волос регулируется меланоцитстимулирующим гормоном , который увеличивает соотношение эумеланина (черного пигмента) и феомеланина (красного пигмента), который кодируется геном MC1R. У современных людей известно пять вариантов этого гена, которые вызывают потерю функции и связаны со светлым цветом кожи и волос, а также еще один неизвестный вариант у неандертальцев (вариант R307G), который может быть связан с бледной кожей и рыжими волосами. Вариант R307G был идентифицирован у неандертальца из Монти Лессини , Италия, и, возможно, из Куэва-дель-Сидрон, Испания. [227] Однако, как и у современных людей, красный цвет волос, вероятно, не был очень распространенным, потому что этот вариант не присутствует у многих других секвенированных неандертальцев. [223]
Метаболизм
[ редактировать ]Максимальная естественная продолжительность жизни и сроки взросления, менопаузы и беременности, скорее всего, были очень похожи на современных людей. [176] была выдвинута гипотеза Однако на основе скорости роста зубов и зубной эмали : [228] [229] что неандертальцы взрослели быстрее современных людей, хотя это не подтверждается возрастными биомаркерами . [86] Основными различиями в созревании являются атлант шеи, а также средние грудные позвонки, сросшиеся у неандертальцев примерно на 2 года позже, чем у современных людей, но это, скорее, было вызвано различиями в анатомии, а не скоростью роста. [230] [231]
неандертальцев в Как правило, модели потребностей калориях сообщают о значительно более высоком потреблении калорий, чем у современных людей, поскольку они обычно предполагают, что у неандертальцев был более высокий уровень основного обмена (BMR) из-за более высокой мышечной массы, более высоких темпов роста и большего производства тепла телом в условиях холода; [232] [233] [234] и более высокие уровни ежедневной физической активности (PAL) из-за больших ежедневных путешествий во время поиска пищи. [233] [234] Однако, используя высокие показатели BMR и PAL, американский археолог Брайан Хокетт подсчитал, что беременная неандерталец потребляла бы 5500 калорий в день, что потребовало бы сильной зависимости от мяса крупной дичи; такая диета могла бы вызвать многочисленные недостатки или отравления питательными веществами, поэтому он пришел к выводу, что эти предположения необоснованны. [234]
Неандертальцы, возможно, были более активны в условиях слабого освещения, а не среди дневного света, потому что они жили в регионах с сокращенным световым днем зимой, охотились на крупную дичь (такие хищники обычно охотятся ночью, чтобы улучшить тактику засады) и имели большие глаза и зрительную обработку. нервные центры. Генетически дальтонизм (который может усиливать мезопическое зрение ) обычно коррелирует с популяциями северных широт, а у неандертальцев из пещеры Виндия в Хорватии были некоторые замены в генах опсина , которые могли повлиять на цветовое зрение. Однако функциональные последствия этих замен неубедительны. [235] Аллели неандертальского происхождения рядом с ASB1 и EXOC6 связаны с вечерним поведением , нарколепсией и дневным сном. [223]
Патология
[ редактировать ]Неандертальцы часто получали травматические повреждения: по оценкам, у 79–94% особей наблюдались признаки заживления серьезных травм, из которых 37–52% были серьезно ранены, а 13–19% получили травмы, не достигнув совершеннолетия. [236] Крайним примером является Шанидар 1 , у которого наблюдаются признаки ампутации правой руки, вероятно, из-за несращения после перелома кости в подростковом возрасте, остеомиелита (инфекция кости) левой ключицы , неправильной походки , проблем со зрением левого глаза. и возможная потеря слуха [237] (возможно, ухо пловца ). [238] В 1995 году Тринкаус подсчитал, что около 80% скончались от полученных травм и умерли, не дожив до 40 лет, и таким образом предположил, что неандертальцы использовали рискованную стратегию охоты (гипотеза «наездника на родео»). [86] Однако частота черепно-мозговых травм существенно не различается между неандертальцами и современными людьми среднего палеолита (хотя у неандертальцев, похоже, был более высокий риск смертности). [239] мало экземпляров как современных людей верхнего палеолита, так и неандертальцев, умерших после 40 лет, [182] и в целом между ними есть схожие модели травм. В 2012 году Тринкаус пришел к выводу, что неандертальцы травмировали себя так же, как и современные люди, например, в результате межличностного насилия. [240] Исследование, проведенное в 2016 году на 124 экземплярах неандертальцев, показало, что высокий уровень травм был вызван нападениями животных , и обнаружило, что около 36% выборки стали жертвами нападений медведей , 21% нападений больших кошек и 17% нападений волков (всего 92 положительных результата). случаев, 74%). Случаев нападения гиен не было, хотя гиены все же, вероятно, нападали на неандертальцев, по крайней мере, оппортунистически. [241] Такое интенсивное хищничество, вероятно, возникло из-за обычных конфронтаций из-за конкуренции за пищу и пространство в пещерах, а также из-за охоты неандертальцев на этих хищников. [241]

Низкая популяция вызвала низкое генетическое разнообразие и, возможно, инбридинг, что снизило способность популяции фильтровать вредные мутации ( инбредная депрессия ). Однако неизвестно, как это повлияло на генетическое бремя одного неандертальца и, следовательно, вызвало ли это более высокий уровень врожденных дефектов, чем у современных людей. [242] Однако известно, что у 13 жителей пещеры Сидрон в совокупности наблюдалось 17 различных врожденных дефектов, вероятно, связанных с инбридингом или рецессивными нарушениями . [243] Вероятно, из-за преклонного возраста (60 или 70 лет) у Ла Шапель-о-Сент 1 были признаки болезни Бааструпа , поражающей позвоночник, и остеоартрит. [244] У Шанидара 1, который, вероятно, умер примерно в 30 или 40 лет, был диагностирован самый древний случай диффузного идиопатического гиперостоза скелета (DISH), дегенеративного заболевания, которое может ограничивать движения, что, если оно правильное, указывает на умеренно высокий уровень заболеваемости среди пожилых людей. Неандертальцы. [245]
Неандертальцы были подвержены нескольким инфекционным заболеваниям и паразитам. Современные люди, вероятно, передали им болезни; Одним из возможных кандидатов является желудочная бактерия Helicobacter pylori . [246] Современный вариант вируса папилломы человека 16А может произойти от интрогрессии неандертальцев. [247] У неандертальца из Куэва-дель-Сидрон, Испания, обнаружены признаки желудочно-кишечной инфекции Enterocytozoon bieneusi . [248] Кости ног французского La Ferrassie 1 имеют поражения, которые соответствуют периоститу — воспалению ткани, окружающей кость, — вероятно, результату гипертрофической остеоартропатии , которая в первую очередь вызвана инфекцией грудной клетки или раком легких . [249] была ниже, У неандертальцев частота кариеса чем у современных людей, несмотря на то, что некоторые популяции потребляли в больших количествах продукты, обычно вызывающие кариес, что может указывать на отсутствие бактерий полости рта, вызывающих кариес, а именно Streptococcus mutans . [250]
Два ребенка-неандерталоида возрастом 250 000 лет из Пайре , Франция, представляют собой самые ранние известные случаи воздействия свинца на гомининов. Они подверглись воздействию в двух разных случаях: либо съели или выпили зараженную пищу или воду, либо вдыхали свинцовый дым от пожара. В радиусе 25 км (16 миль) от объекта находятся две свинцовые шахты. [251]
Культура
[ редактировать ]Социальная структура
[ редактировать ]Групповая динамика
[ редактировать ]
Неандертальцы, вероятно, жили более редко распределёнными группами, чем современные современные люди. [176] но считается, что размер группы в среднем составлял от 10 до 30 особей, как и у современных охотников-собирателей. [31] Надежные доказательства состава группы неандертальцев получены из Куэва-дель-Сидрон, Испания, и следов в Ле-Розеле, Франция: [188] на первом изображены 7 взрослых, 3 подростка, 2 подростка и младенец; [252] тогда как последний, исходя из размера следа, показывает группу из 10–13 человек, из которых 90% составляют несовершеннолетние и подростки. [188]
Анализ зубов неандертальского ребенка в 2018 году показал, что он был отнят от груди через 2,5 года, как и современные охотники-собиратели, и родился весной, что согласуется с современными людьми и другими млекопитающими, чьи циклы рождения совпадают с циклами окружающей среды. [251] что связано с различными заболеваниями, возникающими в результате сильного стресса в молодом возрасте, такими как задержка роста ; Британский археолог Пол Петтитт предположил, что детей обоих полов заставляют работать сразу после отлучения от груди, [183] и Тринкаус сказал, что по достижении подросткового возраста можно было ожидать, что человек присоединится к охоте на крупную и опасную дичь. [86] Однако травма кости сравнима с травмой современных инуитов, что может указывать на схожее детство неандертальцев и современных современных людей. [253] Кроме того, такая задержка роста могла быть также результатом суровых зим и нехватки пищевых ресурсов. [251]
Места, свидетельствующие о присутствии не более трех человек, могли представлять собой нуклеарные семьи или временные места для палаток для групп специального назначения (например, охотничьего отряда). [31] Группы, вероятно, перемещались между определенными пещерами в зависимости от сезона, на что указывают остатки сезонных материалов, таких как определенные продукты питания, и возвращались в одни и те же места поколение за поколением. Некоторые места могли использоваться более 100 лет. [254] Пещерные медведи, возможно, сильно конкурировали с неандертальцами за место в пещере. [255] и наблюдается сокращение популяций пещерных медведей, начиная с 50 000 лет назад (хотя их вымирание произошло намного позже вымирания неандертальцев). [256] [257] Неандертальцы также отдавали предпочтение пещерам, выходы которых были обращены на юг. [258] Хотя обычно считается, что неандертальцы были пещерными обитателями, а «родной базой» была пещера, поселения под открытым небом вблизи одновременно населенных пещерных систем в Леванте могут указывать на мобильность между пещерными и открытыми базами в этой области. Свидетельства существования долговременных поселений под открытым небом известны из городища Эйн-Кашиш в Израиле. [259] [260] и Молдова I в Украине. Хотя неандертальцы, судя по всему, обладали способностью населять различные среды обитания, включая равнины и плато, места обитания неандертальцев под открытым небом обычно интерпретируются как места для убоя скота, а не как жилые помещения. [85]
In 2022, remains of the first-known Neanderthal family (six adults and five children) were excavated from Chagyrskaya Cave in the Altai Mountains of southern Siberia in Russia. The family, which included a father, a daughter, and what appear to be cousins, most likely died together, presumably from starvation.[261][262]
Neanderthals, like contemporaneous modern humans, were most likely polygynous, based on their low second-to-fourth digit ratios, a biomarker for prenatal androgen effects that corresponds to high incidence of polygyny in haplorhine primates.[263]
Inter-group relations
[edit]
Canadian ethnoarchaeologist Brian Hayden calculated a self-sustaining population that avoids inbreeding to consist of about 450–500 individuals, which would necessitate these bands to interact with 8–53 other bands, but more likely the larger estimate given low population density.[31] Analysis of the mtDNA of the Neanderthals of Cueva del Sidrón, Spain, showed that the three adult men belonged to the same maternal lineage, while the three adult women belonged to different ones. This suggests a patrilocal residence (that a woman moved out of her group to live with her partner).[264] However, the DNA of a Neanderthal from Denisova Cave, Russia, shows that she had an inbreeding coefficient of 1⁄8 (her parents were either half-siblings with a common mother, double first cousins, an uncle and niece or aunt and nephew, or a grandfather and granddaughter or grandmother and grandson)[92] and the inhabitants of Cueva del Sidrón show several defects, which may have been caused by inbreeding or recessive disorders.[243]
Considering most Neanderthal artifacts were sourced no more than 5 km (3.1 mi) from the main settlement, Hayden considered it unlikely these bands interacted very often,[31] and mapping of the Neanderthal brain and their small group size and population density could indicate that they had a reduced ability for inter-group interaction and trade.[203] However, a few Neanderthal artefacts in a settlement could have originated 20, 30, 100 and 300 km (12.5, 18.5, 60 and 185 mi) away. Based on this, Hayden also speculated that macro-bands formed which functioned much like those of the low-density hunter-gatherer societies of the Western Desert of Australia. Macro-bands collectively encompass 13,000 km2 (5,000 sq mi), with each band claiming 1,200–2,800 km2 (460–1,080 sq mi), maintaining strong alliances for mating networks or to cope with leaner times and enemies.[31] Similarly, British anthropologist Eiluned Pearce and Cypriot archaeologist Theodora Moutsiou speculated that Neanderthals were possibly capable of forming geographically expansive ethnolinguistic tribes encompassing upwards of 800 people, based on the transport of obsidian up to 300 km (190 mi) from the source compared to trends seen in obsidian transfer distance and tribe size in modern hunter-gatherers. However, according to their model Neanderthals would not have been as efficient at maintaining long-distance networks as modern humans, probably due to a significantly lower population.[265] Hayden noted an apparent cemetery of six or seven individuals at La Ferrassie, France, which, in modern humans, is typically used as evidence of a corporate group which maintained a distinct social identity and controlled some resource, trading, manufacturing and so on. La Ferrassie is also located in one of the richest animal-migration routes of Pleistocene Europe.[31]

Genetic analysis indicates there were at least three distinct geographical groups—Western Europe, the Mediterranean coast, and east of the Caucasus—with some migration among these regions.[84] Post-Eemian Western European Mousterian lithics can also be broadly grouped into three distinct macro-regions: Acheulean-tradition Mousterian in the southwest, Micoquien in the northeast, and Mousterian with bifacial tools (MBT) in between the former two. MBT may actually represent the interactions and fusion of the two different cultures.[83] Southern Neanderthals exhibit regional anatomical differences from northern counterparts: a less protrusive jaw, a shorter gap behind the molars, and a vertically higher jawbone.[266] These all instead suggest Neanderthal communities regularly interacted with neighbouring communities within a region, but not as often beyond.[83]
Nonetheless, over long periods of time, there is evidence of large-scale cross-continental migration. Early specimens from Mezmaiskaya Cave in the Caucasus[139] and Denisova Cave in the Siberian Altai Mountains[90] differ genetically from those found in Western Europe, whereas later specimens from these caves both have genetic profiles more similar to Western European Neanderthal specimens than to the earlier specimens from the same locations, suggesting long-range migration and population replacement over time.[90][139] Similarly, artefacts and DNA from Chagyrskaya and Okladnikov Caves, also in the Altai Mountains, resemble those of eastern European Neanderthal sites about 3,000–4,000 km (1,900–2,500 mi) away more than they do artefacts and DNA of the older Neanderthals from Denisova Cave, suggesting two distinct migration events into Siberia.[267] Neanderthals seem to have suffered a major population decline during MIS 4 (71–57,000 years ago), and the distribution of the Micoquian tradition could indicate that Central Europe and the Caucasus were repopulated by communities from a refuge zone either in eastern France or Hungary (the fringes of the Micoquian tradition) who dispersed along the rivers Prut and Dniester.[268]
There is also evidence of inter-group conflict: a skeleton from La Roche à Pierrot, France, showing a healed fracture on top of the skull apparently caused by a deep blade wound,[269] and another from Shanidar Cave, Iraq, found to have a rib lesion characteristic of projectile weapon injuries.[270]
Social hierarchy
[edit]
It is sometimes suggested that, since they were hunters of challenging big game and lived in small groups, there was no sexual division of labour as seen in modern hunter-gatherer societies. That is, men, women and children all had to be involved in hunting, instead of men hunting with women and children foraging. However, with modern hunter-gatherers, the higher the meat dependency, the higher the division of labour.[31] Further, tooth-wearing patterns in Neanderthal men and women suggest they commonly used their teeth for carrying items, but men exhibit more wearing on the upper teeth, and women the lower, suggesting some cultural differences in tasks.[271]
It is controversially proposed that some Neanderthals wore decorative clothing or jewellery—such as a leopard skin or raptor feathers—to display elevated status in the group. Hayden postulated that the small number of Neanderthal graves found was because only high-ranking members would receive an elaborate burial, as is the case for some modern hunter-gatherers.[31] Trinkaus suggested that elderly Neanderthals were given special burial rites for lasting so long given the high mortality rates.[86] Alternatively, many more Neanderthals may have received burials, but the graves were infiltrated and destroyed by bears.[272] Given that 20 graves of Neanderthals aged under 4 have been found—over a third of all known graves—deceased children may have received greater care during burial than other age demographics.[253]
Looking at Neanderthal skeletons recovered from several natural rock shelters, Trinkaus said that, although Neanderthals were recorded as bearing several trauma-related injuries, none of them had significant trauma to the legs that would debilitate movement. He suggested that self worth in Neanderthal culture derived from contributing food to the group; a debilitating injury would remove this self-worth and result in near-immediate death, and individuals who could not keep up with the group while moving from cave to cave were left behind.[86] However, there are examples of individuals with highly debilitating injuries being nursed for several years, and caring for the most vulnerable within the community dates even further back to H. heidelbergensis.[42][253] Especially given the high trauma rates, it is possible that such an altruistic strategy ensured their survival as a species for so long.[42]
Food
[edit]Hunting and gathering
[edit]
Neanderthals were once thought of as scavengers, but are now considered to have been apex predators.[273][274] In 1980, it was hypothesised that two piles of mammoth skulls at La Cotte de St Brelade, Jersey, at the base of a gulley were evidence of mammoth drive hunting (causing them to stampede off a ledge),[275] but this is contested.[276] Living in a forested environment, Neanderthals were likely ambush hunters, getting close to and attacking their target—a prime adult—in a short burst of speed, thrusting in a spear at close quarters.[66][277] Younger or wounded animals may have been hunted using traps, projectiles, or pursuit.[277] Some sites show evidence that Neanderthals slaughtered whole herds of animals in large, indiscriminate hunts and then carefully selected which carcasses to process.[278] Nonetheless, they were able to adapt to a variety of habitats.[51][276] They appear to have eaten predominantly what was abundant within their immediate surroundings,[53] with steppe-dwelling communities (generally outside of the Mediterranean) subsisting almost entirely on meat from large game, forest-dwelling communities consuming a wide array of plants and smaller animals, and waterside communities gathering aquatic resources, although even in more southerly, temperate areas such as the southeastern Iberian Peninsula, large game still featured prominently in Neanderthal diets.[279] Contemporary humans, in contrast, seem to have used more complex food extraction strategies and generally had a more diverse diet.[280] Nonetheless, Neanderthals still would have had to have eaten a varied enough diet to prevent nutrient deficiencies and protein poisoning, especially in the winter when they presumably ate mostly lean meat. Any food with high contents of other essential nutrients not provided by lean meat would have been vital components of their diet, such as fat-rich brains,[42] carbohydrate-rich and abundant underground storage organs (including roots and tubers),[281] or, like modern Inuit, the stomach contents of herbivorous prey items.[282]
For meat, they appear to have fed predominantly on hoofed mammals,[283] namely red deer and reindeer as these two were the most abundant game,[46][284] but also on other Pleistocene megafauna such as chamois,[285] ibex,[286] wild boar,[285] steppe wisent,[287] aurochs,[285] Irish elk,[288] woolly mammoth,[289] straight-tusked elephant,[290] woolly rhinoceros,[291] Merck's rhinoceros[292] the narrow-nosed rhinoceros,[293] wild horse,[284] and so on.[25][47][294] There is evidence of directed cave and brown bear hunting both in and out of hibernation, as well as butchering.[295] Analysis of Neanderthal bone collagen from Vindija Cave, Croatia, shows nearly all of their protein needs derived from animal meat.[47] Some caves show evidence of regular rabbit and tortoise consumption. At Gibraltar sites, there are remains of 143 different bird species, many ground-dwelling such as the common quail, corn crake, woodlark, and crested lark.[51] Scavenging birds such as corvids and eagles were commonly exploited.[296] Neanderthals also exploited marine resources on the Iberian, Italian and Peloponnesian Peninsulas, where they waded or dived for shellfish,[51][297][298] as early as 150,000 years ago at Cueva Bajondillo, Spain, similar to the fishing record of modern humans.[299] At Vanguard Cave, Gibraltar, the inhabitants consumed Mediterranean monk seal, short-beaked common dolphin, common bottlenose dolphin, Atlantic bluefin tuna, sea bream and purple sea urchin;[51][300] and at Gruta da Figueira Brava, Portugal, there is evidence of large-scale harvest of shellfish, crabs and fish.[301] Evidence of freshwater fishing was found in Grotte di Castelcivita, Italy, for trout, chub and eel;[298] Abri du Maras, France, for chub and European perch; Payré, France;[302] and Kudaro Cave, Russia, for Black Sea salmon.[303]
Edible plant and mushroom remains are recorded from several caves.[49] Neanderthals from Cueva del Sidrón, Spain, based on dental tartar, likely had a meatless diet of mushrooms, pine nuts and moss, indicating they were forest foragers.[248] Remnants from Amud Cave, Israel, indicates a diet of figs, palm tree fruits and various cereals and edible grasses.[50] Several bone traumas in the leg joints could possibly suggest habitual squatting, which, if the case, was likely done while gathering food.[304] Dental tartar from Grotte de Spy, Belgium, indicates the inhabitants had a meat-heavy diet including woolly rhinoceros and mouflon sheep, while also regularly consuming mushrooms.[248] Neanderthal faecal matter from El Salt, Spain, dated to 50,000 years ago—the oldest human faecal matter remains recorded—show a diet mainly of meat but with a significant component of plants.[305] Evidence of cooked plant foods—mainly legumes and, to a far lesser extent, acorns—was discovered at the Kebara Cave site in Israel, with its inhabitants possibly gathering plants in spring and fall and hunting in all seasons except fall, although the cave was probably abandoned in late summer to early fall.[40] At Shanidar Cave, Iraq, Neanderthals collected plants with various harvest seasons, indicating they scheduled returns to the area to harvest certain plants, and that they had complex food-gathering behaviours for both meat and plants.[48]
Food preparation
[edit]Neanderthals probably could employ a wide range of cooking techniques, such as roasting, and they may have been able to heat up or boil soup, stew, or animal stock.[44] The abundance of animal bone fragments at settlements may indicate the making of fat stocks from boiling bone marrow, possibly taken from animals that had already died of starvation. These methods would have substantially increased fat consumption, which was a major nutritional requirement of communities with low carbohydrate and high protein intake.[44][306] Neanderthal tooth size had a decreasing trend after 100,000 years ago, which could indicate an increased dependence on cooking or the advent of boiling, a technique that would have softened food.[307]

At Cueva del Sidrón, Spain, Neanderthals likely cooked and possibly smoked food,[45] as well as used certain plants—such as yarrow and camomile—as flavouring,[44] although these plants may have instead been used for their medicinal properties.[39] At Gorham's Cave, Gibraltar, Neanderthals may have been roasting pinecones to access pine nuts.[51]
At Grotte du Lazaret, France, a total of twenty-three red deer, six ibexes, three aurochs, and one roe deer appear to have been hunted in a single autumn hunting season, when strong male and female deer herds would group together for rut. The entire carcasses seem to have been transported to the cave and then butchered. Because this is such a large amount of food to consume before spoilage, it is possible these Neanderthals were curing and preserving it before winter set in. At 160,000 years old, it is the oldest potential evidence of food storage.[43] The great quantities of meat and fat which could have been gathered in general from typical prey items (namely mammoths) could also indicate food storage capability.[308] With shellfish, Neanderthals needed to eat, cook, or in some manner preserve them soon after collection, as shellfish spoils very quickly. At Cueva de los Aviones, Spain, the remains of edible, algae eating shellfish associated with the alga Jania rubens could indicate that, like some modern hunter gatherer societies, harvested shellfish were held in water-soaked algae to keep them alive and fresh until consumption.[309]
Competition
[edit]
Competition from large Ice Age predators was rather high. Cave lions likely targeted horses, large deer and wild cattle; and leopards primarily reindeer and roe deer; which heavily overlapped with Neanderthal diet. To defend a kill against such ferocious predators, Neanderthals may have engaged in a group display of yelling, arm waving, or stone throwing; or quickly gathered meat and abandoned the kill. However, at Grotte de Spy, Belgium, the remains of wolves, cave lions and cave bears—which were all major predators of the time—indicate Neanderthals hunted their competitors to some extent.[52]
Neanderthals and cave hyenas may have exemplified niche differentiation, and actively avoided competing with each other. Although they both mainly targeted the same groups of creatures—deer, horses and cattle—Neanderthals mainly hunted the former and cave hyenas the latter two. Further, animal remains from Neanderthal caves indicate they preferred to hunt prime individuals, whereas cave hyenas hunted weaker or younger prey, and cave hyena caves have a higher abundance of carnivore remains.[46] Nonetheless, there is evidence that cave hyenas stole food and leftovers from Neanderthal campsites and scavenged on dead Neanderthal bodies.[310]
Cannibalism
[edit]
There are several instances of Neanderthals practising cannibalism across their range.[312][313] The first example came from the Krapina, Croatia site, in 1899,[120] and other examples were found at Cueva del Sidrón[266] and Zafarraya in Spain; and the French Grotte de Moula-Guercy,[314] Les Pradelles, and La Quina. For the five cannibalised Neanderthals at the Grottes de Goyet, Belgium, there is evidence that the upper limbs were disarticulated, the lower limbs defleshed and also smashed (likely to extract bone marrow), the chest cavity disemboweled, and the jaw dismembered. There is also evidence that the butchers used some bones to retouch their tools. The processing of Neanderthal meat at Grottes de Goyet is similar to how they processed horse and reindeer.[312][313] About 35% of the Neanderthals at Marillac-le-Franc, France, show clear signs of butchery, and the presence of digested teeth indicates that the bodies were abandoned and eaten by scavengers, likely hyaenas.[315]
These cannibalistic tendencies have been explained as either ritual defleshing, pre-burial defleshing (to prevent scavengers or foul smell), an act of war, or simply for food. Due to a small number of cases, and the higher number of cut marks seen on cannibalised individuals than animals (indicating inexperience), cannibalism was probably not a very common practice, and it may have only been done in times of extreme food shortages as in some cases in recorded human history.[313]
The arts
[edit]Personal adornment
[edit]Neanderthals used ochre, a clay earth pigment. Ochre is well documented from 60 to 45,000 years ago in Neanderthal sites, with the earliest example dating to 250–200,000 years ago from Maastricht-Belvédère, the Netherlands (a similar timespan to the ochre record of H. sapiens).[316] It has been hypothesised to have functioned as body paint, and analyses of pigments from Pech de l'Azé, France, indicates they were applied to soft materials (such as a hide or human skin).[317] However, modern hunter gatherers, in addition to body paint, also use ochre for medicine, for tanning hides, as a food preservative, and as an insect repellent, so its use as decorative paint for Neanderthals is speculative.[316] Containers apparently used for mixing ochre pigments were found in Peștera Cioarei, Romania, which could indicate modification of ochre for solely aesthetic purposes.[318]

Neanderthals collected uniquely shaped objects and are suggested to have modified them into pendants, such as a fossil Aspa marginata sea snail shell possibly painted red from Grotta di Fumane, Italy, transported over 100 km (62 mi) to the site about 47,500 years ago;[319] three shells, dated to about 120–115,000 years ago, perforated through the umbo belonging to a rough cockle, a Glycymeris insubrica, and a Spondylus gaederopus from Cueva de los Aviones, Spain, the former two associated with red and yellow pigments, and the latter a red-to-black mix of hematite and pyrite; and a king scallop shell with traces of an orange mix of goethite and hematite from Cueva Antón, Spain. The discoverers of the latter two claim that pigment was applied to the exterior to make it match the naturally vibrant inside colouration.[56][309] Excavated from 1949 to 1963 from the French Grotte du Renne, Châtelperronian beads made from animal teeth, shells and ivory were found associated with Neanderthal bones, but the dating is uncertain and Châtelperronian artefacts may actually have been crafted by modern humans and simply redeposited with Neanderthal remains.[320][321][322][323]

Gibraltarian palaeoanthropologists Clive and Geraldine Finlayson suggested that Neanderthals used various bird parts as artistic media, specifically black feathers.[324] In 2012, the Finlaysons and colleagues examined 1,699 sites across Eurasia, and argued that raptors and corvids, species not typically consumed by any human species, were overrepresented and show processing of only the wing bones instead of the fleshier torso, and thus are evidence of feather plucking of specifically the large flight feathers for use as personal adornment. They specifically noted the cinereous vulture, red-billed chough, kestrel, lesser kestrel, alpine chough, rook, jackdaw and the white tailed eagle in Middle Palaeolithic sites.[325] Other birds claimed to present evidence of modifications by Neanderthals are the golden eagle, rock pigeon, common raven and the bearded vulture.[326] The earliest claim of bird bone jewellery is a number of 130,000-year-old white tailed eagle talons found in a cache near Krapina, Croatia, speculated, in 2015, to have been a necklace.[327][328] A similar 39,000-year-old Spanish imperial eagle talon necklace was reported in 2019 at Cova Foradà in Spain, though from the contentious Châtelperronian layer.[329] In 2017, 17 incision-decorated raven bones from the Zaskalnaya VI rock shelter, Ukraine, dated to 43–38,000 years ago were reported. Because the notches are more-or-less equidistant to each other, they are the first modified bird bones that cannot be explained by simple butchery, and for which the argument of design intent is based on direct evidence.[54]
Discovered in 1975, the so-called Mask of la Roche-Cotard, a mostly flat piece of flint with a bone pushed through a hole on the midsection—dated to 32, 40, or 75,000 years ago[330]—has been purported to resemble the upper half of a face, with the bone representing eyes.[331][332] It is contested whether it represents a face, or if it even counts as art.[333] In 1988, American archaeologist Alexander Marshack speculated that a Neanderthal at Grotte de L'Hortus, France, wore a leopard pelt as personal adornment to indicate elevated status in the group based on a recovered leopard skull, phalanges and tail vertebrae.[31][334]
Abstraction
[edit]
As of 2014, 63 purported engravings have been reported from 27 different European and Middle Eastern Lower-to-Middle Palaeolithic sites, of which 20 are on flint cortexes from 11 sites, 7 are on slabs from 7 sites, and 36 are on pebbles from 13 sites. It is debated whether or not these were made with symbolic intent.[58] In 2012, deep scratches on the floor of Gorham's Cave, Gibraltar, were discovered, dated to older than 39,000 years ago, which the discoverers have interpreted as Neanderthal abstract art.[335][336] The scratches could have also been produced by a bear.[272] In 2021, an Irish elk phalanx with five engraved offset chevrons stacked above each other was discovered at the entrance to the Einhornhöhle cave in Germany, dating to about 51,000 years ago.[337]
In 2018, some red-painted dots, disks, lines and hand stencils on the cave walls of the Spanish La Pasiega, Maltravieso, and Doña Trinidad were dated to be older than 66,000 years ago, at least 20,000 years prior to the arrival of modern humans in Western Europe. This would indicate Neanderthal authorship, and similar iconography recorded in other Western European sites—such as Les Merveilles, France, and Cueva del Castillo, Spain—could potentially also have Neanderthal origins.[61][62][338] However, the dating of these Spanish caves, and thus attribution to Neanderthals, is contested.[60]
Neanderthals are known to have collected a variety of unusual objects—such as crystals or fossils—without any real functional purpose or any indication of damage caused by use. It is unclear if these objects were simply picked up for their aesthetic qualities, or if some symbolic significance was applied to them. These items are mainly quartz crystals, but also other minerals such as cerussite, iron pyrite, calcite and galena. A few findings feature modifications, such as a mammoth tooth with an incision and a fossil nummulite shell with a cross etched in from Tata, Hungary; a large slab with 18 cupstones hollowed out from a grave in La Ferrassie, France;[57] and a geode from Peștera Cioarei, Romania, coated with red ochre.[339] A number of fossil shells are also known from French Neanderthals sites, such as a rhynchonellid and a Taraebratulina from Combe Grenal; a belemnite beak from Grottes des Canalettes; a polyp from Grotte de l'Hyène; a sea urchin from La Gonterie-Boulouneix; and a rhynchonella, feather star and belemnite beak from the contentious Châtelperronian layer of Grotte du Renne.[57]
Music
[edit]
Purported Neanderthal bone flute fragments made of bear long bones were reported from Potočka zijalka, Slovenia, in the 1920s, and Istállós-kői-barlang, Hungary,[340] and Mokriška jama, Slovenia, in 1985; but these are now attributed to modern human activities.[341][342]
The 43,000-year-old Divje Babe flute from Slovenia, found in 1995, has been attributed by some researchers to Neanderthals, though its status as a flute is heavily disputed. Many researchers consider it to be most likely the product of a carnivorous animal chewing the bone,[343][342][344] but its discoverer Ivan Turk and other researchers have maintained an argument that it was manufactured by Neanderthal as a musical instrument.[59]
Technology
[edit]Despite the apparent 150,000-year stagnation in Neanderthal lithic innovation,[176] there is evidence that Neanderthal technology was more sophisticated than was previously thought.[64] However, the high frequency of potentially debilitating injuries could have prevented very complex technologies from emerging, as a major injury would have impeded an expert's ability to effectively teach a novice.[236]
Stone tools
[edit]

Neanderthals made stone tools, and are associated with the Mousterian industry.[27] The Mousterian is also associated with North African H. sapiens as early as 315,000 years ago[345] and was found in Northern China about 47–37,000 years ago in caves such as Jinsitai or Tongtiandong.[346] It evolved around 300,000 years ago with the Levallois technique which developed directly from the preceding Acheulean industry (invented by H. erectus about 1.8 mya). Levallois made it easier to control flake shape and size, and as a difficult-to-learn and unintuitive process, the Levallois technique may have been directly taught generation to generation rather than via purely observational learning.[28]
There are distinct regional variants of the Mousterian industry, such as: the Quina and La Ferrassie subtypes of the Charentian industry in southwestern France, Acheulean-tradition Mousterian subtypes A and B along the Atlantic and northwestern European coasts,[347] the Micoquien industry of Central and Eastern Europe and the related Sibiryachikha variant in the Siberian Altai Mountains,[267] the Denticulate Mousterian industry in Western Europe, the racloir industry around the Zagros Mountains, and the flake cleaver industry of Cantabria, Spain, and both sides of the Pyrenees. In the mid-20th century, French archaeologist François Bordes debated against American archaeologist Lewis Binford to explain this diversity (the "Bordes–Binford debate"), with Bordes arguing that these represent unique ethnic traditions and Binford that they were caused by varying environments (essentially, form vs. function).[347] The latter sentiment would indicate a lower degree of inventiveness compared to modern humans, adapting the same tools to different environments rather than creating new technologies.[53] A continuous sequence of occupation is well documented in Grotte du Renne, France, where the lithic tradition can be divided into the Levallois–Charentian, Discoid–Denticulate (43,300 ±929 – 40,900 ±719 years ago), Levallois Mousterian (40,200 ±1,500 – 38,400 ±1,300 years ago) and Châtelperronian (40,930 ±393 – 33,670 ±450 years ago).[348]
There is some debate if Neanderthals had long-ranged weapons.[349][350] A wound on the neck of an African wild ass from Umm el Tlel, Syria, was likely inflicted by a heavy Levallois-point javelin,[351] and bone trauma consistent with habitual throwing has been reported in Neanderthals.[349][350] Some spear tips from Abri du Maras, France, may have been too fragile to have been used as thrusting spears, possibly suggesting their use as darts.[302]
Organic tools
[edit]The Châtelperronian in central France and northern Spain is a distinct industry from the Mousterian, and is controversially hypothesised to represent a culture of Neanderthals borrowing (or by process of acculturation) tool-making techniques from immigrating modern humans, crafting bone tools and ornaments. In this frame, the makers would have been a transitional culture between the Neanderthal Mousterian and the modern human Aurignacian.[352][353][354][355][356] The opposing viewpoint is that the Châtelperronian was manufactured by modern humans instead.[357] Abrupt transitions similar to the Mousterian/Châtelperronian could also simply represent natural innovation, like the La Quina–Neronian transition 50,000 years ago featuring technologies generally associated with modern humans such as bladelets and microliths. Other ambiguous transitional cultures include the Italian Uluzzian industry,[358] and the Balkan Szeletian industry.[359]
Before immigration, the only evidence of Neanderthal bone tools are animal rib lissoirs—which are rubbed against hide to make it more supple or waterproof—although this could also be evidence for modern humans immigrating earlier than expected. In 2013, two 51,400- to 41,100-year-old deer rib lissoirs were reported from Pech-de-l'Azé and the nearby Abri Peyrony in France.[354][100][100] In 2020, five more lissoirs made of aurochs or bison ribs were reported from Abri Peyrony, with one dating to about 51,400 years ago and the other four to 47,700–41,100 years ago. This indicates the technology was in use in this region for a long time. Since reindeer remains were the most abundant, the use of less abundant bovine ribs may indicate a specific preference for bovine ribs. Potential lissoirs have also been reported from Grosse Grotte, Germany (made of mammoth), and Grottes des Canalettes, France (red deer).[360]

The Neanderthals in 10 coastal sites in Italy (namely Grotta del Cavallo and Grotta dei Moscerini) and Kalamakia Cave, Greece, are known to have crafted scrapers using smooth clam shells, and possibly hafted them to a wooden handle. They probably chose this clam species because it has the most durable shell. At Grotta dei Moscerini, about 24% of the shells were gathered alive from the seafloor, meaning these Neanderthals had to wade or dive into shallow waters to collect them. At Grotta di Santa Lucia, Italy, in the Campanian volcanic arc, Neanderthals collected the porous volcanic pumice, which, for contemporary humans, was probably used for polishing points and needles. The pumices are associated with shell tools.[298]
At Abri du Maras, France, twisted fibres and a 3-ply inner-bark-fibre cord fragment associated with Neanderthals show that they produced string and cordage, but it is unclear how widespread this technology was because the materials used to make them (such as animal hair, hide, sinew, or plant fibres) are biodegradable and preserve very poorly. This technology could indicate at least a basic knowledge of weaving and knotting, which would have made possible the production of nets, containers, packaging, baskets, carrying devices, ties, straps, harnesses, clothes, shoes, beds, bedding, mats, flooring, roofing, walls and snares, and would have been important in hafting, fishing and seafaring. Dating to 52–41,000 years ago, the cord fragment is the oldest direct evidence of fibre technology, although 115,000-year-old perforated shell beads from Cueva Antón possibly strung together to make a necklace are the oldest indirect evidence.[36][302] In 2020, British archaeologist Rebecca Wragg Sykes expressed cautious support for the genuineness of the find, but pointed out that the string would have been so weak that it would have had limited functions. One possibility is as a thread for attaching or stringing small objects.[361]
The archaeological record shows that Neanderthals commonly used animal hide and birch bark, and may have used them to make cooking containers, although this is based largely on circumstantial evidence, because neither fossilizes well.[307] It is possible that the Neanderthals at Kebara Cave in Israel, used the shells of the spur-thighed tortoise as containers.[362]
At the Italian Poggetti Vecchi site, there is evidence that they used fire to process boxwood branches to make digging sticks, a common implement in hunter-gatherer societies.[363]
The Schöningen spears are a collection of wooden spears probably made by early Neanderthals found in Germany, dating to around 300,000 years ago. They were likely both thrown and used as handheld thrusting spears. The tools were specifically made of spruce, (or possibly larch in some specimens) and pine despite their uncommonness in the environment, suggesting that they had been deliberately selected for their material properties. The spears had been deliberately debarked, followed by the ends being sharpened using cutting and scraping. Other wooden tools made of split wood were also found at the site, some rounded and some pointed, which may have functioned for domestic tasks, like serving as awls (used to make holes) and hide smoothers for the pointed and rounded types respectively. The wooden artifacts show evidence of being repurposed and reshaped.[364]
Fire and construction
[edit]Many Mousterian sites have evidence of fire, some for extended periods of time, though it is unclear whether they were capable of starting fire or simply scavenged from naturally occurring wildfires. Indirect evidence of fire-starting ability includes pyrite residue on a couple of dozen bifaces from late Mousterian (c. 50,000 years ago) northwestern France (which could indicate they were used as percussion fire starters), and collection of manganese dioxide by late Neanderthals which can lower the combustion temperature of wood.[29][30][365] They were also capable of zoning areas for specific activities, such as for knapping, butchering, hearths and wood storage. Many Neanderthal sites lack evidence for such activity perhaps due to natural degradation of the area over tens of thousands of years, such as by bear infiltration after abandonment of the settlement.[272]
In a number of caves, evidence of hearths has been detected. Neanderthals likely considered air circulation when making hearths as a lack of proper ventilation for a single hearth can render a cave uninhabitable in several minutes. Abric Romaní rock shelter, Spain, indicates eight evenly spaced hearths lined up against the rock wall, likely used to stay warm while sleeping, with one person sleeping on either side of the fire.[31][32] At Cueva de Bolomor, Spain, with hearths lined up against the wall, the smoke flowed upwards to the ceiling, and led to outside the cave. In Grotte du Lazaret, France, smoke was probably naturally ventilated during the winter as the interior cave temperature was greater than the outside temperature; likewise, the cave was likely only inhabited in the winter.[32]

In 1990, two 176,000-year-old ring structures, several metres wide, made of broken stalagmite pieces, were discovered in a large chamber more than 300 m (980 ft) from the entrance within Grotte de Bruniquel, France. One ring was 6.7 m × 4.5 m (22 ft × 15 ft) with stalagmite pieces averaging 34.4 cm (13.5 in) in length, and the other 2.2 m × 2.1 m (7.2 ft × 6.9 ft) with pieces averaging 29.5 cm (11.6 in). There were also four other piles of stalagmite pieces for a total of 112 m (367 ft) or 2.2 t (2.4 short tons) worth of stalagmite pieces. Evidence of the use of fire and burnt bones also suggest human activity. A team of Neanderthals was likely necessary to construct the structure, but the chamber's actual purpose is uncertain. Building complex structures so deep in a cave is unprecedented in the archaeological record, and indicates sophisticated lighting and construction technology, and great familiarity with subterranean environments.[366]
The 44,000-year-old Moldova I open-air site, Ukraine, shows evidence of a 7 m × 10 m (23 ft × 33 ft) ring-shaped dwelling made out of mammoth bones meant for long-term habitation by several Neanderthals, which would have taken a long time to build. It appears to have contained hearths, cooking areas and a flint workshop, and there are traces of woodworking. Upper Palaeolithic modern humans in the Russian plains are thought to have also made housing structures out of mammoth bones.[85]
Birch tar
[edit]Neanderthal produced the adhesive birch bark tar, using the bark of birch trees, for hafting.[367] It was long believed that birch bark tar required a complex recipe to be followed, and that it thus showed complex cognitive skills and cultural transmission. However, a 2019 study showed it can be made simply by burning birch bark beside smooth vertical surfaces, such as a flat, inclined rock.[34] Thus, tar making does not require cultural processes per se. However, at Königsaue (Germany), Neanderthals did not make tar with such an aboveground method but rather employed a technically more demanding underground production method. This is one of our best indicators that some of their techniques were conveyed by cultural processes.[368]
Clothes
[edit]Neanderthals were likely able to survive in a similar range of temperatures to modern humans while sleeping: about 32 °C (90 °F) while naked in the open and windspeed 5.4 km/h (3.4 mph), or 27–28 °C (81–82 °F) while naked in an enclosed space. Since ambient temperatures were markedly lower than this—averaging, during the Eemian interglacial, 17.4 °C (63.3 °F) in July and 1 °C (34 °F) in January and dropping to as a low as −30 °C (−22 °F) on the coldest days—Danish physicist Bent Sørensen hypothesised that Neanderthals required tailored clothing capable of preventing airflow to the skin. Especially during extended periods of travelling (such as a hunting trip), tailored footwear completely enwrapping the feet may have been necessary.[369]

Nonetheless, as opposed to the bone sewing-needles and stitching awls assumed to have been in use by contemporary modern humans, the only known Neanderthal tools that could have been used to fashion clothes are hide scrapers, which could have made items similar to blankets or ponchos, and there is no direct evidence they could produce fitted clothes.[35][370] Indirect evidence of tailoring by Neanderthals includes the ability to manufacture string, which could indicate weaving ability,[302] and a naturally-pointed horse metatarsal bone from Cueva de los Aviones, Spain, which was speculated to have been used as an awl, perforating dyed hides, based on the presence of orange pigments.[309] Whatever the case, Neanderthals would have needed to cover up most of their body, and contemporary humans would have covered 80–90%.[370][371]
Since human/Neanderthal admixture is known to have occurred in the Middle East, and no modern body louse species descends from their Neanderthal counterparts (body lice only inhabit clothed individuals), it is possible Neanderthals (and/or humans) in hotter climates did not wear clothes, or Neanderthal lice were highly specialised.[371]
Seafaring
[edit]Remains of Middle Palaeolithic stone tools on Greek islands indicate early seafaring by Neanderthals in the Ionian Sea possibly starting as far back as 200–150,000 years ago. The oldest stone artefacts from Crete date to 130–107,000 years ago, Cephalonia 125,000 years ago, and Zakynthos 110–35,000 years ago. The makers of these artefacts likely employed simple reed boats and made one-day crossings back and forth.[37] Other Mediterranean islands with such remains include Sardinia, Melos, Alonnisos,[38] and Naxos (although Naxos may have been connected to land),[372] and it is possible they crossed the Strait of Gibraltar.[38] If this interpretation is correct, Neanderthals' ability to engineer boats and navigate through open waters would speak to their advanced cognitive and technical skills.[38][372]
Medicine
[edit]Given their dangerous hunting and extensive skeletal evidence of healing, Neanderthals appear to have lived lives of frequent traumatic injury and recovery. Well-healed fractures on many bones indicate the setting of splints. Individuals with severe head and rib traumas (which would have caused massive blood loss) indicate they had some manner of dressing major wounds, such as bandages made from animal skin. By and large, they appear to have avoided severe infections, indicating good long-term treatment of such wounds.[42]
Their knowledge of medicinal plants was comparable to that of contemporary humans.[42] An individual at Cueva del Sidrón, Spain, seems to have been medicating a dental abscess using poplar—which contains salicylic acid, the active ingredient in aspirin—and there were also traces of the antibiotic-producing Penicillium chrysogenum.[248] They may also have used yarrow and camomile, and their bitter taste—which should act as a deterrent as it could indicate poison—means it was likely a deliberate act.[39] At the Kebara Cave in Israel, plant remains which have historically been used for their medicinal properties were found, including the common grape vine, the pistachios of the Persian turpentine tree, ervil seeds and oak acorns.[40]
Language
[edit]
The degree of language complexity is difficult to establish, but given that Neanderthals achieved some technical and cultural complexity, and interbred with humans, it is reasonable to assume they were at least fairly articulate, comparable to modern humans. Some researchers have argued, a somewhat complex language—possibly using syntax—was likely necessary to survive in their harsh environment, with Neanderthals needing to communicate about topics such as locations, hunting and gathering, and tool-making techniques.[64][373][374] The FOXP2 gene in modern humans is associated with speech and language development. FOXP2 was present in Neanderthals,[375] but not the gene's modern human variant.[376] Neurologically, Neanderthals had an expanded Broca's area—operating the formulation of sentences, and speech comprehension, but out of a group of 48 genes believed to affect the neural substrate of language, 11 had different methylation patterns between Neanderthals and modern humans. This could indicate a stronger ability in modern humans than in Neanderthals to express language.[377]
In 1971, cognitive scientist Philip Lieberman attempted to reconstruct the Neanderthal vocal tract and concluded that it was similar to that of a newborn and incapable of producing a large range of speech sounds, due to the large size of the mouth and the small size of the pharyngeal cavity (according to his reconstruction), thus no need for a descended larynx to fit the entire tongue inside the mouth. He claimed that they were anatomically unable to produce the sounds /a/, /i/, /u/, /ɔ/, /g/, and /k/ and thus lacked the capacity for articulate speech, though were still able to speak at a level higher than non-human primates.[378][379][380] However, the lack of a descended larynx does not necessarily equate to a reduced vowel capacity.[381] The 1983 discovery of a Neanderthal hyoid bone—used in speech production in humans—in Kebara 2 which is almost identical to that of humans suggests Neanderthals were capable of speech. Also, the ancestral Sima de los Huesos hominins had humanlike hyoid and ear bones, which could suggest the early evolution of the modern human vocal apparatus. However, the hyoid does not definitively provide insight into vocal tract anatomy.[65] Subsequent studies reconstruct the Neanderthal vocal apparatus as comparable to that of modern humans, with a similar vocal repertoire.[382] In 2015, Lieberman hypothesized that Neanderthals were capable of syntactical language, although nonetheless incapable of mastering any human dialect.[383]
It is debated if behavioural modernity is a recent and uniquely modern human innovation, or if Neanderthals also possessed it.[55][374][384][385]
Religion
[edit]Funerals
[edit]
Although Neanderthals did bury their dead, at least occasionally—which may explain the abundance of fossil remains[53]—the behavior is not indicative of a religious belief of life after death because it could also have had non-symbolic motivations, such as great emotion[386] or the prevention of scavenging.[387]
Estimates made regarding the number of known Neanderthal burials range from thirty-six to sixty.[388][389][390][391] The oldest confirmed burials do not seem to occur before approximately 70,000 years ago.[392] The small number of recorded Neanderthal burials implies that the activity was not particularly common. The setting of inhumation in Neanderthal culture largely consisted of simple, shallow graves and pits.[393] Sites such as La Ferrassie in France or Shanidar in Iraq may imply the existence of mortuary centers or cemeteries in Neanderthal culture due to the number of individuals found buried at them.[393]
The debate on Neanderthal funerals has been active since the 1908 discovery of La Chapelle-aux-Saints 1 in a small, artificial hole in a cave in southwestern France, very controversially postulated to have been buried in a symbolic fashion.[394][395][396] Another grave at Shanidar Cave, Iraq, was associated with the pollen of several flowers that may have been in bloom at the time of deposition—yarrow, centaury, ragwort, grape hyacinth, joint pine and hollyhock.[397] The medicinal properties of the plants led American archaeologist Ralph Solecki to claim that the man buried was some leader, healer, or shaman, and that "The association of flowers with Neanderthals adds a whole new dimension to our knowledge of his humanness, indicating that he had 'soul' ".[398] However, it is also possible the pollen was deposited by a small rodent after the man's death.[399]
The graves of children and infants, especially, are associated with grave goods such as artefacts and bones. The grave of a newborn from La Ferrassie, France, was found with three flint scrapers, and an infant from Dederiyeh Cave, Syria, was found with a triangular flint placed on its chest. A 10-month-old from Amud Cave, Israel, was associated with a red deer mandible, likely purposefully placed there given other animal remains are now reduced to fragments. Teshik-Tash 1 from Uzbekistan was associated with a circle of ibex horns, and a limestone slab argued to have supported the head.[253] A child from Kiik-Koba, Crimea, Ukraine, had a flint flake with some purposeful engraving on it, likely requiring a great deal of skill.[58] Nonetheless, these contentiously constitute evidence of symbolic meaning as the grave goods' significance and worth are unclear.[253]
Cults
[edit]It was once argued that the bones of the cave bear, particularly the skull, in some European caves were arranged in a specific order, indicating an ancient bear cult that killed bears and then ceremoniously arranged the bones. This would be consistent with bear-related rituals of modern human Arctic hunter-gatherers, but the alleged peculiarity of the arrangement could also be sufficiently explained by natural causes,[63][386] and bias could be introduced as the existence of a bear cult would conform with the idea that totemism was the earliest religion, leading to undue extrapolation of evidence.[400]
It was also once thought that Neanderthals ritually hunted, killed and cannibalised other Neanderthals and used the skull as the focus of some ceremony.[313] In 1962, Italian palaeontologist Alberto Blanc believed a skull from Grotta Guattari, Italy, had evidence of a swift blow to the head—indicative of ritual murder—and a precise and deliberate incising at the base to access the brain. He compared it to the victims of headhunters in Malaysia and Borneo,[401] putting it forward as evidence of a skull cult.[386] However, it is now thought to have been a result of cave hyaena scavengery.[402] Although Neanderthals are known to have practiced cannibalism, there is unsubstantial evidence to suggest ritual defleshing.[312]
In 2019, Gibraltarian palaeoanthropologists Stewart, Geraldine and Clive Finlayson and Spanish archaeologist Francisco Guzmán speculated that the golden eagle had iconic value to Neanderthals, as exemplified in some modern human societies because they reported that golden eagle bones had a conspicuously high rate of evidence of modification compared to the bones of other birds. They then proposed some "Cult of the Sun Bird" where the golden eagle was a symbol of power.[55][326] There is evidence from Krapina, Croatia, from wear use and even remnants of string, that suggests that raptor talons were worn as personal ornaments.[403]
Interbreeding
[edit]Interbreeding with modern humans
[edit]
The first Neanderthal genome sequence was published in 2010, and strongly indicated interbreeding between Neanderthals and early modern humans.[87][405][406][407] The genomes of all studied modern populations contain Neanderthal DNA.[87][89][408][409][410] Various estimates exist for the proportion, such as 1–4%[87] or 3.4–7.9% in modern Eurasians,[411] or 1.8–2.4% in modern Europeans and 2.3–2.6% in modern East Asians.[412] Pre-agricultural Europeans appear to have had similar, or slightly higher,[410] percentages to modern East Asians, and the numbers may have decreased in the former due to dilution with a group of people which had split off before Neanderthal introgression.[100] Typically, studies have reported finding no significant levels of Neanderthal DNA in Sub-Saharan Africans, but a 2020 study detected 0.3-0.5% in the genomes of five African sample populations, likely the result of Eurasians back-migrating and interbreeding with Africans, as well as human-to-neanderthal gene flow from dispersals of Homo sapiens preceding the larger Out-of-Africa migration, and also showed more equal Neanderthal DNA percentages for European and Asian populations.[410] Such low percentages of Neanderthal DNA in all present day populations indicate infrequent past interbreeding,[413] unless interbreeding was more common with a different population of modern humans which did not contribute to the present day gene pool.[100] Of the inherited Neanderthal genome, 25% in modern Europeans and 32% in modern East Asians may be related to viral immunity.[414] In all, approximately 20% of the Neanderthal genome appears to have survived in the modern human gene pool.[94]

However, due to their small population and resulting reduced effectivity of natural selection, Neanderthals accumulated several weakly harmful mutations, which were introduced to and slowly selected out of the much larger modern human population; the initial hybridised population may have experienced up to a 94% reduction in fitness compared to contemporary humans. By this measure, Neanderthals may have substantially increased in fitness.[82] A 2017 study focusing on archaic genes in Turkey found associations with coeliac disease, malaria severity and Costello syndrome.[416] Nonetheless, some genes may have helped modern East Asians adapt to the environment; the putatively Neanderthal Val92Met variant of the MC1R gene, which may be weakly associated with red hair and UV radiation sensitivity,[417] is primarily found in East Asian, rather than European, individuals.[418] Some genes related to the immune system appear to have been affected by introgression, which may have aided migration,[96] such as OAS1,[419] STAT2,[420] TLR6, TLR1, TLR10,[421] and several related to immune response.[95][f] In addition, Neanderthal genes have also been implicated in the structure and function of the brain,[g] keratin filaments, sugar metabolism, muscle contraction, body fat distribution, enamel thickness and oocyte meiosis.[99] Nonetheless, a large portion of surviving introgression appears to be non-coding ("junk") DNA with few biological functions.[100]
There is considerably less Neanderthal ancestry on the X-chromosome as compared to the autosomal chromosomes. This has led to suggestions that admixture with modern humans was sex biased, and primarily the result of mating between modern human females and Neanderthal males. Other authors have suggested that this may be due to negative selection against Neanderthal alleles, however these two proposals are not mutually exclusive.[141] A 2023 study confirmed that the low level of Neanderthal ancestry on the X-chromosomes is best explained by sex bias in the admixture events, and these authors also found evidence for negative selection on archaic genes.[423]
Neanderthal mtDNA (which is passed on from mother to child) is absent in modern humans.[136][155][424] This is evidence that interbreeding occurred mainly between Neanderthal males and modern human females.[425] According to Svante Pääbo, it is not clear that modern humans were socially dominant over Neanderthals, which may explain why the interbreeding occurred primarily between Neanderthal males and modern human females.[426] Furthermore, even if Neanderthal women and modern human males had interbred, Neanderthal mtDNA lineages may have gone extinct if women who carried them only gave birth to sons.[426]
Due to the lack of Neanderthal-derived Y-chromosomes in modern humans (which is passed on from father to son), it has also been suggested that the hybrids that contributed ancestry to modern populations were predominantly females, or the Neanderthal Y-chromosome was not compatible with H. sapiens and became extinct.[100][427]
According to linkage disequilibrium mapping, the last Neanderthal gene flow into the modern human genome occurred 86–37,000 years ago, but most likely 65–47,000 years ago.[428] It is thought that Neanderthal genes which contributed to the present day human genome stemmed from interbreeding in the Near East rather than the entirety of Europe. However, interbreeding still occurred without contributing to the modern genome.[100] The approximately 40,000-year-old modern human Oase 2 was found, in 2015, to have had 6–9% (point estimate 7.3%) Neanderthal DNA, indicating a Neanderthal ancestor up to four to six generations earlier, but this hybrid population does not appear to have made a substantial contribution to the genomes of later Europeans.[415] In 2016, the DNA of Neanderthals from Denisova Cave revealed evidence of interbreeding 100,000 years ago, and interbreeding with an earlier dispersal of H. sapiens may have occurred as early as 120,000 years ago in places such as the Levant.[91] The earliest H. sapiens remains outside of Africa occur at Misliya Cave 194–177,000 years ago, and Skhul and Qafzeh 120–90,000 years ago.[429] The Qafzeh humans lived at approximately the same time as the Neanderthals from the nearby Tabun Cave.[430] The Neanderthals of the German Hohlenstein-Stadel have deeply divergent mtDNA compared to more recent Neanderthals, possibly due to introgression of human mtDNA between 316,000 and 219,000 years ago, or simply because they were genetically isolated.[90] Whatever the case, these first interbreeding events have not left any trace in modern human genomes.[431]
Genetic evidence suggests that following their split from Denisovans, Neanderthals experienced gene flow (around 3% of their genome) from the lineage leading to modern humans prior to the expansion of modern humans outside of Africa during the Last Glacial Period, with this interbreeding suggested to have taken place around 200-300,000 years ago.[141]
Detractors of the interbreeding model argue that the genetic similarity is only a remnant of a common ancestor instead of interbreeding,[432] although this is unlikely as it fails to explain why sub-Saharan Africans do not have Neanderthal DNA.[406]
Interbreeding with Denisovans
[edit]
Although nDNA confirms that Neanderthals and Denisovans are more closely related to each other than they are to modern humans, Neanderthals and modern humans share a more recent maternally-transmitted mtDNA common ancestor, possibly due to interbreeding between Denisovans and some unknown human species. The 400,000-year-old Neanderthal-like humans from Sima de los Huesos in northern Spain, looking at mtDNA, are more closely related to Denisovans than Neanderthals. Several Neanderthal-like fossils in Eurasia from a similar time period are often grouped into H. heidelbergensis, of which some may be relict populations of earlier humans, which could have interbred with Denisovans.[434] This is also used to explain an approximately 124,000-year-old German Neanderthal specimen with mtDNA that diverged from other Neanderthals (except for Sima de los Huesos) about 270,000 years ago, while its genomic DNA indicated divergence less than 150,000 years ago.[90]
Sequencing of the genome of a Denisovan from Denisova Cave has shown that 17% of its genome derives from Neanderthals.[93] This Neanderthal DNA more closely resembled that of a 120,000-year-old Neanderthal bone from the same cave than that of Neanderthals from Vindija Cave, Croatia, or Mezmaiskaya Cave in the Caucasus, suggesting that interbreeding was local.[92]
For the 90,000-year-old Denisova 11, it was found that her father was a Denisovan related to more recent inhabitants of the region, and her mother a Neanderthal related to more recent European Neanderthals at Vindija Cave, Croatia. Given how few Denisovan bones are known, the discovery of a first-generation hybrid indicates interbreeding was very common between these species, and Neanderthal migration across Eurasia likely occurred sometime after 120,000 years ago.[435]
Extinction
[edit]Transition
[edit]
The extinction of Neanderthals was part of the broader Late Pleistocene megafaunal extinction event.[436] Whatever the cause of their extinction, Neanderthals were replaced by modern humans, indicated by near full replacement of Middle Palaeolithic Mousterian stone technology with modern human Upper Palaeolithic Aurignacian stone technology across Europe (the Middle-to-Upper Palaeolithic Transition) from 41,000 to 39,000 years ago.[8][9][11][437] By between 44,200 to 40,600 BP, Neanderthals vanished from northwestern Europe.[438] However, it is postulated that Iberian Neanderthals persisted until about 35,000 years ago, as indicated by the date range of transitional lithic assemblages—Châtelperronian, Uluzzian, Protoaurignacian and Early Aurignacian. The latter two are attributed to modern humans, but the former two have unconfirmed authorship, potentially products of Neanderthal/modern human cohabitation and cultural transmission. Further, the appearance of the Aurignacian south of the Ebro River has been dated to roughly 37,500 years ago, which has prompted the "Ebro Frontier" hypothesis which states that the river presented a geographic barrier preventing modern human immigration, and thus prolonging Neanderthal persistence.[439][440] However, the dating of the Iberian Transition is debated, with a contested timing of 43,000–40,800 years ago at Cueva Bajondillo, Spain.[441][442][443][444] The Châtelperronian appears in northeastern Iberia about 42,500–41,600 years ago.[439]
Some Neanderthals in Gibraltar were dated to much later than this—such as Zafarraya (30,000 years ago)[445] and Gorham's Cave (28,000 years ago)[446]—which may be inaccurate as they were based on ambiguous artefacts instead of direct dating.[11] A claim of Neanderthals surviving in a polar refuge in the Ural Mountains[169] is loosely supported by Mousterian stone tools dating to 34,000 years ago from the northern Siberian Byzovaya site at a time when modern humans may not yet have colonised the northern reaches of Europe;[171] however, modern human remains are known from the nearby Mamontovaya Kurya site dating to 40,000 years ago.[447] Indirect dating of Neanderthals remains from Mezmaiskaya Cave reported a date of about 30,000 years ago, but direct dating instead yielded 39,700 ±1,100 years ago, more in line with trends exhibited in the rest of Europe.[10]

The earliest indication of Upper Palaeolithic modern human immigration into Europe is the Balkan Bohunician industry beginning 48,000 years ago, likely deriving from the Levantine Emiran industry,[359] and the earliest bones in Europe date to roughly 45–43,000 years ago in Bulgaria,[448] Italy,[449] and Britain.[450] This wave of modern humans replaced Neanderthals.[8] However, Neanderthals and H. sapiens have a much longer contact history. DNA evidence indicates H. sapiens contact with Neanderthals and admixture as early as 120–100,000 years ago. A 2019 reanalysis of 210,000-year-old skull fragments from the Greek Apidima Cave assumed to have belonged to a Neanderthal concluded that they belonged to a modern human, and a Neanderthal skull dating to 170,000 years ago from the cave indicates H. sapiens were replaced by Neanderthals until returning about 40,000 years ago.[451] This identification was refuted by a 2020 study.[452] Archaeological evidence suggests that Neanderthals displaced modern humans in the Near East around 100,000 years ago until about 60–50,000 years ago.[100]
Cause
[edit]Modern humans
[edit]
Historically, modern human technology was viewed as vastly superior to that of Neanderthals, with more efficient weaponry and subsistence strategies, and Neanderthals simply went extinct because they could not compete.[19]
The discovery of Neanderthal/modern human introgression has caused the resurgence of the multiregional hypothesis, wherein the present day genetic makeup of all humans is the result of complex genetic contact among several different populations of humans dispersed across the world. By this model, Neanderthals and other recent archaic humans were simply assimilated into the modern human genome – that is, they were effectively bred out into extinction.[19] Modern humans coexisted with Neanderthals in Europe for around 3,000 to 5,000 years.[453]
Climate change
[edit]Their ultimate extinction coincides with Heinrich event 4, a period of intense seasonality; later Heinrich events are also associated with massive cultural turnovers when European human populations collapsed.[20][21] This climate change may have depopulated several regions of Neanderthals, like previous cold spikes, but these areas were instead repopulated by immigrating humans, leading to Neanderthal extinction.[454] In southern Iberia, there is evidence that Neanderthal populations declined during H4 and the associated proliferation of Artemisia-dominated desert-steppes.[455]

It has also been proposed that climate change was the primary driver, as their low population left them vulnerable to any environmental change, with even a small drop in survival or fertility rates possibly quickly leading to their extinction.[456] However, Neanderthals and their ancestors had survived through several glacial periods over their hundreds of thousands of years of European habitation.[280] It is also proposed that around 40,000 years ago, when Neanderthal populations may have already been dwindling from other factors, the Campanian Ignimbrite Eruption in Italy could have led to their final demise, as it produced 2–4 °C (3.6–7.2 °F) cooling for a year and acid rain for several more years.[22][457]
Disease
[edit]Modern humans may have introduced African diseases to Neanderthals, contributing to their extinction. A lack of immunity, compounded by an already low population, was potentially devastating to the Neanderthal population, and low genetic diversity could have also rendered fewer Neanderthals naturally immune to these new diseases ("differential pathogen resistance" hypothesis). However, compared to modern humans, Neanderthals had a similar or higher genetic diversity for 12 major histocompatibility complex (MHC) genes associated with the adaptive immune system, casting doubt on this model.[24]
Low population and inbreeding depression may have caused maladaptive birth defects, which could have contributed to their decline (mutational meltdown).[243]
In late-20th-century New Guinea, due to cannibalistic funerary practices, the Fore people were decimated by transmissible spongiform encephalopathies, specifically kuru, a highly virulent disease spread by ingestion of prions found in brain tissue. However, individuals with the 129 variant of the PRNP gene were naturally immune to the prions. Studying this gene led to the discovery that the 129 variant was widespread among all modern humans, which could indicate widespread cannibalism at some point in human prehistory. Because Neanderthals are known to have practised cannibalism to an extent and to have co-existed with modern humans, British palaeoanthropologist Simon Underdown speculated that modern humans transmitted a kuru-like spongiform disease to Neanderthals, and, because the 129 variant appears to have been absent in Neanderthals, it quickly killed them off.[23][458]
In popular culture
[edit]
Neanderthals have been portrayed in popular culture including appearances in literature, visual media and comedy. The "caveman" archetype often mocks Neanderthals and depicts them as primitive, hunchbacked, knuckle-dragging, club-wielding, grunting, nonsocial characters driven solely by animal instinct. "Neanderthal" can also be used as an insult.[25]
In literature, they are sometimes depicted as brutish or monstrous, such as in H. G. Wells' The Grisly Folk and Elizabeth Marshall Thomas' The Animal Wife, but sometimes with a civilised but unfamiliar culture, as in William Golding's The Inheritors, Björn Kurtén's Dance of the Tiger, and Jean M. Auel's Clan of the Cave Bear and her Earth's Children series.[26]
See also
[edit]- Denisovan – Asian archaic human
- Early human migrations
- Early European modern humans – Earliest anatomically modern humans in Europe
- Homo floresiensis – Archaic human from Flores, Indonesia
- Homo luzonensis – Archaic human from Luzon, Philippines
- Homo naledi – South African archaic human species
- Human timeline
Footnotes
[edit]- ^ After being mined for limestone, the cave caved in and was lost by 1900. It was rediscovered in 1997 by archaeologists Ralf Schmitz and Jürgen Thissen.[101]
- ^ The German spelling Thal ("valley") was current until 1901 but has been Tal since then. (The German noun is cognate with English dale.) The German /t/ phoneme was frequently spelled th from the 15th to 19th centuries, but the spelling Tal became standardized in 1901 and the old spellings of the German names Neanderthal for the valley and Neanderthaler for the species were both changed to the spellings without h.[102][103]
- ^ In Mettmann, "Neander Valley", there is a local idiosyncrasy in use of the outdated spellings with th, such as with the Neanderthal Museum (but the name is in English [German would require Neandertalermuseum]), the Neanderthal station (Bahnhof Neanderthal), and some other rare occasions meant for tourists. Beyond these, city convention is to use th when referring to the species.[103]
- ^ King made a typo and said "theositic".
- ^ The bones were discovered by workers of Wilhelm Beckershoff and Friedrich Wilhelm Pieper. Initially, the workers threw the bones out as debris, but Beckershoff then told them to store the bones. Pieper asked Fuhlrott to come up to the cave and investigate the bones, which Beckershoff and Pieper believed belonged to a cave bear.[101]
- ^ OAS1[419] and STAT2[420] both are associated with fighting viral inflections (interferons), and the listed toll-like receptors (TLRs)[421] allow cells to identify bacterial, fungal, or parasitic pathogens. African origin is also correlated with a stronger inflammatory response.[95]
- ^ Higher levels of Neanderthal-derived genes are associated with an occipital and parietal bone shape reminiscent to that of Neanderthals, as well as modifications to the visual cortex and the intraparietal sulcus (associated with visual processing).[422]
- ^ Homo floresiensis originated in an unknown location from unknown ancestors and reached remote parts of Indonesia. Homo erectus spread from Africa to western Asia, then east Asia and Indonesia; its presence in Europe is uncertain, but it gave rise to Homo antecessor, found in Spain. Homo heidelbergensis originated from Homo erectus in an unknown location and dispersed across Africa, southern Asia and southern Europe (other scientists interpret fossils, here named heidelbergensis, as late erectus). Modern humans spread from Africa to western Asia and then to Europe and southern Asia, eventually reaching Australia and the Americas. In addition to Neanderthals and Denisovans, a third gene flow of archaic Africa origin is indicated at the right.[433] The chart is missing superarchaic (which diverged from erectus 1.9 mya) introgression into Neanderthal/Denisovan common ancestor.[159]
References
[edit]- ^ Jump up to: a b Haeckel, E. (1895). Systematische Phylogenie: Wirbelthiere (in German). G. Reimer. p. 601.
- ^ Schwalbe, G. (1906). Studien zur Vorgeschichte des Menschen [Studies on the pre-history of man] (in German). Stuttgart, E. Nägele. doi:10.5962/bhl.title.61918. hdl:2027/uc1.b4298459.
- ^ Klaatsch, H. (1909). "Preuves que l'Homo Mousteriensis Hauseri appartient au type de Neandertal" [Evidence that Homo Mousteriensis Hauseri belongs to the Neanderthal type]. L'Homme Préhistorique (in French). 7: 10–16.
- ^ Romeo, L. (1979). Ecce Homo!: a lexicon of man. John Benjamins Publishing Company. p. 92. ISBN 978-90-272-2006-6.
- ^ Jump up to: a b c d e McCown, T.; Keith, A. (1939). The stone age of Mount Carmel. The fossil human remains from the Levalloisso-Mousterian. Vol. 2. Clarenden Press.
- ^ Szalay, F. S.; Delson, E. (2013). Evolutionary history of the Primates. Academic Press. p. 508. ISBN 978-1-4832-8925-0.
- ^ Wells, J. (2008). Longman pronunciation dictionary (3rd ed.). Harlow, England: Pearson Longman. ISBN 978-1-4058-8118-0.
- ^ Jump up to: a b c Higham, T.; Douka, K.; Wood, R.; Ramsey, C. B.; Brock, F.; Basell, L.; Camps, M.; Arrizabalaga, A.; Baena, J.; Barroso-Ruíz, C.; C. Bergman; C. Boitard; P. Boscato; M. Caparrós; N.J. Conard; C. Draily; A. Froment; B. Galván; P. Gambassini; A. Garcia-Moreno; S. Grimaldi; P. Haesaerts; B. Holt; M.-J. Iriarte-Chiapusso; A. Jelinek; J.F. Jordá Pardo; J.-M. Maíllo-Fernández; A. Marom; J. Maroto; M. Menéndez; L. Metz; E. Morin; A. Moroni; F. Negrino; E. Panagopoulou; M. Peresani; S. Pirson; M. de la Rasilla; J. Riel-Salvatore; A. Ronchitelli; D. Santamaria; P. Semal; L. Slimak; J. Soler; N. Soler; A. Villaluenga; R. Pinhasi; R. Jacobi; et al. (2014). "The timing and spatiotemporal patterning of Neanderthal disappearance". Nature. 512 (7514): 306–309. Bibcode:2014Natur.512..306H. doi:10.1038/nature13621. hdl:1885/75138. PMID 25143113. S2CID 205239973.
We show that the Mousterian [the Neanderthal tool-making tradition] ended by 41,030–39,260 calibrated years BP (at 95.4% probability) across Europe. We also demonstrate that succeeding 'transitional' archaeological industries, one of which has been linked with Neanderthals (Châtelperronian), end at a similar time.
- ^ Jump up to: a b Higham, T. (2011). "European Middle and Upper Palaeolithic radiocarbon dates are often older than they look: problems with previous dates and some remedies". Antiquity. 85 (327): 235–249. doi:10.1017/s0003598x00067570. S2CID 163207571.
Few events of European prehistory are more important than the transition from ancient to modern humans about 40,000 years ago, a period that unfortunately lies near the limit of radiocarbon dating. This paper shows that as many as 70 per cent of the oldest radiocarbon dates in the literature may be too young, due to contamination by modern carbon.
- ^ Jump up to: a b Pinhasi, R.; Higham, T. F. G.; Golovanova, L. V.; Doronichev, V. B. (2011). "Revised age of late Neanderthal occupation and the end of the Middle Palaeolithic in the northern Caucasus". Proceedings of the National Academy of Sciences. 108 (21): 8611–8616. Bibcode:2011PNAS..108.8611P. doi:10.1073/pnas.1018938108. PMC 3102382. PMID 21555570.
The direct date of the fossil (39,700±1,100 14C BP) is in good agreement with the probability distribution function, indicating at a high level of probability that Neanderthals did not survive at Mezmaiskaya Cave after 39 kya cal BP. [...] This challenges previous claims for late Neanderthal survival in the northern Caucasus. [...] Our results confirm the lack of reliably dated Neanderthal fossils younger than ≈40 kya cal BP in any other region of Western Eurasia, including the Caucasus.
- ^ Jump up to: a b c Galván, B.; Hernández, C. M.; Mallol, C.; Mercier, N.; Sistiaga, A.; Soler, V. (2014). "New evidence of early Neanderthal disappearance in the Iberian Peninsula". Journal of Human Evolution. 75: 16–27. Bibcode:2014JHumE..75...16G. doi:10.1016/j.jhevol.2014.06.002. PMID 25016565.
- ^ Jump up to: a b c Stringer, C. (2012). "The status of Homo heidelbergensis (Schoetensack 1908)". Evolutionary Anthropology. 21 (3): 101–107. doi:10.1002/evan.21311. PMID 22718477. S2CID 205826399.
- ^ Jump up to: a b Gómez-Robles, A. (2019). "Dental evolutionary rates and its implications for the Neanderthal–modern human divergence". Science Advances. 5 (5): eaaw1268. Bibcode:2019SciA....5.1268G. doi:10.1126/sciadv.aaw1268. PMC 6520022. PMID 31106274.
- ^ Jump up to: a b c d e f g Meyer, M.; Arsuaga, J.; de Filippo, C.; Nagel, S. (2016). "Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins". Nature. 531 (7595): 504–507. Bibcode:2016Natur.531..504M. doi:10.1038/nature17405. PMID 26976447. S2CID 4467094.
- ^ Jump up to: a b Klein, R. G. (1983). "What Do We Know About Neanderthals and Cro-Magnon Man?". Anthropology. 52 (3): 386–392. JSTOR 41210959.
- ^ Vaesen, Krist; Dusseldorp, Gerrit L.; Brandt, Mark J. (2021). "An emerging consensus in palaeoanthropology: Demography was the main factor responsible for the disappearance of Neanderthals". Scientific Reports. 11 (1): 4925. Bibcode:2021NatSR..11.4925V. doi:10.1038/s41598-021-84410-7. ISSN 2045-2322. PMC 7921565. PMID 33649483.
- ^ Vaesen, Krist; Dusseldorp, Gerrit L.; Brandt, Mark J. (2021). "Author correction: 'An Emerging Consensus in Palaeoanthropology: Demography Was the Main Factor Responsible for the Disappearance of Neanderthals'". Scientific Reports. 11 (1): 8450. Bibcode:2021NatSR..11.8450V. doi:10.1038/s41598-021-88189-5. PMC 8044239. PMID 33850254. S2CID 233232999.
- ^ Jump up to: a b Wynn, Thomas; Overmann, Karenleigh A; Coolidge, Frederick L (2016). "The false dichotomy: A refutation of the Neandertal indistinguishability claim". Journal of Anthropological Sciences. 94 (94): 201–221. doi:10.4436/jass.94022 (inactive March 6, 2024). PMID 26708102. S2CID 15817439.
{{cite journal}}: CS1 maint: DOI inactive as of March 2024 (link) - ^ Jump up to: a b c Villa, P.; Roebroeks, W. (2014). "Neandertal demise: an archaeological analysis of the modern human superiority complex". PLOS ONE. 9 (4): e96424. Bibcode:2014PLoSO...996424V. doi:10.1371/journal.pone.0096424. PMC 4005592. PMID 24789039.
- ^ Jump up to: a b Bradtmöller, M.; Pastoors, A.; Weninger, B.; Weninger, G. (2012). "The repeated replacement model – Rapid climate change and population dynamics in Late Pleistocene Europe". Quaternary International. 247: 38–49. Bibcode:2012QuInt.247...38B. doi:10.1016/j.quaint.2010.10.015.
- ^ Jump up to: a b Wolf, D.; Kolb, T.; Alcaraz-Castaño, M.; Heinrich, S. (2018). "Climate deteriorations and Neanderthal demise in interior Iberia". Scientific Reports. 8 (1): 7048. Bibcode:2018NatSR...8.7048W. doi:10.1038/s41598-018-25343-6. PMC 5935692. PMID 29728579.
- ^ Jump up to: a b c Black, B. A.; Neely, R. R.; Manga, M. (2015). "Campanian Ignimbrite volcanism, climate, and the final decline of the Neanderthals" (PDF). Geology. 43 (5): 411–414. Bibcode:2015Geo....43..411B. doi:10.1130/G36514.1. OSTI 1512181. S2CID 128647846.
- ^ Jump up to: a b Underdown, S. (2008). "A potential role for transmissible spongiform encephalopathies in Neanderthal extinction". Medical Hypotheses. 71 (1): 4–7. doi:10.1016/j.mehy.2007.12.014. PMID 18280671.
- ^ Jump up to: a b Sullivan, A. P.; de Manuel, M.; Marques-Bonet, T.; Perry, G. H. (2017). "An evolutionary medicine perspective on Neandertal extinction" (PDF). Journal of Human Evolution. 108: 62–71. Bibcode:2017JHumE.108...62S. doi:10.1016/j.jhevol.2017.03.004. PMID 28622932. S2CID 4592682.
- ^ Jump up to: a b c d e f g Papagianni & Morse 2013.
- ^ Jump up to: a b c d e f g h i Drell, J. R. R. (2000). "Neanderthals: a history of interpretation". Oxford Journal of Archaeology. 19 (1): 1–24. doi:10.1111/1468-0092.00096. S2CID 54616107.
- ^ Jump up to: a b Shaw, I.; Jameson, R., eds. (1999). A dictionary of archaeology. Blackwell. p. 408. ISBN 978-0-631-17423-3.
- ^ Jump up to: a b Lycett, S. J.; von Cramon-Taubadel, N. (2013). "A 3D morphometric analysis of surface geometry in Levallois cores: patterns of stability and variability across regions and their implications". Journal of Archaeological Science. 40 (3): 1508–1517. Bibcode:2013JArSc..40.1508L. doi:10.1016/j.jas.2012.11.005.
- ^ Jump up to: a b Sorensen, A. C.; Claud, E.; Soressi, M. (2018). "Neandertal fire-making technology inferred from microwear analysis". Scientific Reports. 8 (1): 10065. Bibcode:2018NatSR...810065S. doi:10.1038/s41598-018-28342-9. ISSN 2045-2322. PMC 6053370. PMID 30026576.
- ^ Jump up to: a b Brittingham, A.; Hren, M. T.; Hartman, G.; Wilkinson, K. N.; Mallol, C.; Gasparyan, B.; Adler, D. S. (2019). "Geochemical evidence for the control of fire by Middle Palaeolithic hominins". Scientific Reports. 9 (1): 15368. Bibcode:2019NatSR...915368B. doi:10.1038/s41598-019-51433-0. PMC 6814844. PMID 31653870.
- ^ Jump up to: a b c d e f g h i j k Hayden, B. (2012). "Neandertal social structure?". Oxford Journal of Archaeology. 31 (1): 1–26. doi:10.1111/j.1468-0092.2011.00376.x.
- ^ Jump up to: a b c Kedar, Y.; Barkai, R. (2019). "The significance of air circulation and hearth location at Paleolithic cave sites". Open Quaternary. 5 (1): 4. doi:10.5334/oq.52.
- ^ Angelucci, Diego E.; Nabais, Mariana; Zilhão, João (2023). "Formation processes, fire use, and patterns of human occupation across the Middle Palaeolithic (MIS 5a-5b) of Gruta da Oliveira (Almonda karst system, Torres Novas, Portugal)". PLOS ONE. 18 (10): e0292075. Bibcode:2023PLoSO..1892075A. doi:10.1371/journal.pone.0292075. PMC 10566745. PMID 37819902.
- ^ Jump up to: a b Schmidt, P.; Blessing, M.; Rageot, M.; Iovita, R.; Pfleging, J.; Nickel, K. G.; Righetti, L.; Tennie, C. (2019). "Birch tar production does not prove Neanderthal behavioral complexity". Proceedings of the National Academy of Sciences. 116 (36): 17707–17711. Bibcode:2019PNAS..11617707S. doi:10.1073/pnas.1911137116. PMC 6731756. PMID 31427508.
- ^ Jump up to: a b Hoffecker, J. F. (2009). "The spread of modern humans in Europe". Proceedings of the National Academy of Sciences. 106 (38): 16040–16045. Bibcode:2009PNAS..10616040H. doi:10.1073/pnas.0903446106. PMC 2752585. PMID 19571003.
- ^ Jump up to: a b Hardy, B. L.; Moncel, M.-H.; Kerfant, C.; et al. (2020). "Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications". Scientific Reports. 10 (4889): 4889. Bibcode:2020NatSR..10.4889H. doi:10.1038/s41598-020-61839-w. PMC 7145842. PMID 32273518.
- ^ Jump up to: a b Ferentinos, G.; Gkioni, M.; Geraga, M.; Papatheodorou, G. (2012). "Early seafaring activity in the southern Ionian Islands, Mediterranean Sea". Journal of Archaeological Science. 39 (7): 2167–2176. Bibcode:2011JQS....26..553S. doi:10.1016/j.jas.2012.01.032.
- ^ Jump up to: a b c d Strasser, T. F.; Runnels, C.; Wegmann, K. W.; Panagopoulou, E. (2011). "Dating Palaeolithic sites in southwestern Crete, Greece". Journal of Quaternary Science. 26 (5): 553–560. Bibcode:2011JQS....26..553S. doi:10.1016/j.jas.2012.01.032.
- ^ Jump up to: a b c Buckley, S.; Hardy, K.; Huffman, M. (2013). "Neanderthal self-medication in context". Antiquity. 87 (337): 873–878. doi:10.1017/S0003598X00049528. S2CID 160563162.
- ^ Jump up to: a b c Lev, E.; Kislev, M. E.; Bar-Yosef, O. (2005). "Mousterian vegetal food in Kebara Cave, Mt. Carmel". Journal of Archaeological Science. 32 (3): 475–484. Bibcode:2005JArSc..32..475L. doi:10.1016/j.jas.2004.11.006.
- ^ Power, R. C.; Salazar-García, D. C.; Rubini, M.; Darlas, A.; Harvati, K.; Walker, M.; Hublin, J.; Henry, A. G. (2018). "Dental calculus indicates widespread plant use within the stable Neanderthal dietary niche". Journal of Human Evolution. 119: 27–41. Bibcode:2018JHumE.119...27P. doi:10.1016/j.jhevol.2018.02.009. hdl:10550/65536. ISSN 0047-2484. PMID 29685752. S2CID 13831823.
- ^ Jump up to: a b c d e f Spikins, P.; Needham, A.; Wright, B. (2019). "Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare". Quaternary Science Reviews. 217: 98–118. Bibcode:2019QSRv..217...98S. doi:10.1016/j.quascirev.2018.08.011.
- ^ Jump up to: a b c Valensi, P.; Michel, V.; et al. (2013). "New data on human behavior from a 160,000 year old Acheulean occupation level at Lazaret cave, south-east France: An archaeozoological approach". Quaternary International. 316: 123–139. Bibcode:2013QuInt.316..123V. doi:10.1016/j.quaint.2013.10.034.
- ^ Jump up to: a b c d Krief, S.; Daujeard, C.; Moncel, M.; Lamon, N.; Reynolds, V. (2015). "Flavouring food: the contribution of chimpanzee behaviour to the understanding of Neanderthal calculus composition and plant use in Neanderthal diets". Antiquity. 89 (344): 464–471. doi:10.15184/aqy.2014.7. S2CID 86646905.
- ^ Jump up to: a b Hardy, K.; Buckley, S.; Collins, M. J.; Estalrrich, A. (2012). "Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus". The Science of Nature. 99 (8): 617–626. Bibcode:2012NW.....99..617H. doi:10.1007/s00114-012-0942-0. PMID 22806252. S2CID 10925552.
- ^ Jump up to: a b c d Dusseldorp, G. L. (2013). "Neanderthals and Cave Hyenas: Co-existence, Competition or Conflict?" (PDF). In Clark, J. L.; Speth, J. D. (eds.). Zooarchaeology and Modern Human Origins. Vertebrate paleobiology and paleoanthropology. Springer Science+Business Media Dordrecht. pp. 191–208. doi:10.1007/978-94-007-6766-9_12. ISBN 978-94-007-6765-2.
- ^ Jump up to: a b c Richards, M. P.; Pettitt, P. B.; Trinkaus, E.; Smith, F. H.; Paunović, M.; Karavanić, I. (2000). "Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes". Proceedings of the National Academy of Sciences of the United States of America. 97 (13): 7663–7666. Bibcode:2000PNAS...97.7663R. doi:10.1073/pnas.120178997. JSTOR 122870. PMC 16602. PMID 10852955.
- ^ Jump up to: a b Henry, A. G.; Brooks, A. S.; Piperno, D. R. (2011). "Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium)". Proceedings of the National Academy of Sciences. 108 (2): 486–491. Bibcode:2011PNAS..108..486H. doi:10.1073/pnas.1016868108. PMC 3021051. PMID 21187393.
- ^ Jump up to: a b Shipley, G. P.; Kindscher, K. (2016). "Evidence for the paleoethnobotany of the Neanderthal: a review of the literature". Scientifica. 2016: 1–12. doi:10.1155/2016/8927654. PMC 5098096. PMID 27843675.
- ^ Jump up to: a b Madella, M.; Jones, M. K.; Goldberg, P.; Goren, Y.; Hovers, E. (2002). "The Exploitation of plant resources by Neanderthals in Amud Cave (Israel): the evidence from phytolith studies". Journal of Archaeological Science. 29 (7): 703–719. Bibcode:2002JArSc..29..703M. doi:10.1006/jasc.2001.0743. S2CID 43217031.
- ^ Jump up to: a b c d e f Brown 2011.
- ^ Jump up to: a b Shipman 2015, pp. 120–143.
- ^ Jump up to: a b c d Tattersall 2015.
- ^ Jump up to: a b d’Errico, F.; Tsvelykh, A. (2017). "A decorated raven bone from the Zaskalnaya VI (Kolosovskaya) Neanderthal site, Crimea". PLOS ONE. 12 (3): e0173435. Bibcode:2017PLoSO..1273435M. doi:10.1371/journal.pone.0173435. PMC 5371307. PMID 28355292.
- ^ Jump up to: a b c Finlayson 2019.
- ^ Jump up to: a b Hoffman, D. L.; Angelucci, D. E.; Villaverde, V.; Zapata, Z.; Zilhão, J. (2018). "Symbolic use of marine shells and mineral pigments by Iberian Neandertals 115,000 years ago". Science Advances. 4 (2): eaar5255. Bibcode:2018SciA....4.5255H. doi:10.1126/sciadv.aar5255. PMC 5833998. PMID 29507889.
- ^ Jump up to: a b c Moncel, M.-H.; Chiotti, L.; Gaillard, C.; Onoratini, G.; Pleurdeau, D. (2012). "Non utilitarian objects in the Palaeolithic: emergence of the sense of precious?". Archaeology, Ethnology & Anthropology of Eurasia. 401: 25–27. doi:10.1016/j.aeae.2012.05.004.
- ^ Jump up to: a b c Majkić, A.; d’Errico, F.; Stepanchuk, V. (2018). "Assessing the significance of Palaeolithic engraved cortexes. A case study from the Mousterian site of Kiik-Koba, Crimea". PLOS ONE. 13 (5): e0195049. Bibcode:2018PLoSO..1395049M. doi:10.1371/journal.pone.0195049. PMC 5931501. PMID 29718916.
- ^ Jump up to: a b Turk, M.; Turk, I.; Dimkaroski, L. (2018). "The Mousterian musical instrument from the Divje Babe I Cave (Slovenia): arguments on the material evidence for Neanderthal musical behaviour". L'Anthropologie. 122 (4): 1–28. doi:10.1016/j.anthro.2018.10.001. S2CID 133682741. Archived from the original on October 20, 2021. Retrieved January 9, 2020.
- ^ Jump up to: a b Aubert, M.; Brumm, A.; Huntley, J. (2018). "Early dates for 'Neanderthal cave art' may be wrong". Journal of Human Evolution. 125: 215–217. Bibcode:2018JHumE.125..215A. doi:10.1016/j.jhevol.2018.08.004. PMID 30173883. S2CID 52145541.
- ^ Jump up to: a b Pike, A. W.; Hoffmann, D. L.; Pettitt, P. B.; García-Diez, M.; Zilhão, J. (2017). "Dating Palaeolithic cave art: Why U–Th is the way to go" (PDF). Quaternary International. 432: 41–49. Bibcode:2017QuInt.432...41P. doi:10.1016/j.quaint.2015.12.013. S2CID 130629624.
- ^ Jump up to: a b Hoffmann, D. L.; Standish, C. D.; García-Diez, M.; Pettitt, P. B.; Milton, J. A.; Zilhão, J.; Alcolea-González, J. J.; Cantalejo-Duarte, P.; Collado, H.; de Balbín, R.; Lorblanchet, M.; Ramos-Muñoz, J.; Weniger, G.-C.; Pike, A. W. G. (2018). "U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art". Science. 359 (6378): 912–915. Bibcode:2018Sci...359..912H. doi:10.1126/science.aap7778. hdl:10498/21578. PMID 29472483.
- ^ Jump up to: a b Wunn, I. (2000). "Beginning of religion". Numen. 47 (4): 417–452. doi:10.1163/156852700511612.
- ^ Jump up to: a b c Dediu, D.; Levinson, S. C. (2018). "Neanderthal language revisited: not only us". Current Opinion in Behavioral Sciences. 21: 49–55. doi:10.1016/j.cobeha.2018.01.001. hdl:21.11116/0000-0000-1667-4. S2CID 54391128.
- ^ Jump up to: a b D’Anastasio, R.; Wroe, S.; Tuniz, C.; Mancini, L.; Cesana, D. T. (2013). "Micro-biomechanics of the Kebara 2 hyoid and its implications for speech in Neanderthals". PLOS ONE. 8 (12): e82261. Bibcode:2013PLoSO...882261D. doi:10.1371/journal.pone.0082261. PMC 3867335. PMID 24367509.
- ^ Jump up to: a b c d e f g Stewart, J.R.; García-Rodríguez, O.; Knul, M.V.; Sewell, L.; Montgomery, H.; Thomas, M.G.; Diekmann, Y. (2019). "Palaeoecological and genetic evidence for Neanderthal power locomotion as an adaptation to a woodland environment". Quaternary Science Reviews. 217: 310–315. Bibcode:2019QSRv..217..310S. doi:10.1016/j.quascirev.2018.12.023. S2CID 133980969.
- ^ Jump up to: a b Kislev, M.; Barkai, R. (2018). "Neanderthal and woolly mammoth molecular resemblance". Human Biology. 90 (2): 115–128. doi:10.13110/humanbiology.90.2.03. PMID 33951886. S2CID 106401104.
- ^ Jump up to: a b de Azevedo, S.; González, M. F.; Cintas, C.; et al. (2017). "Nasal airflow simulations suggest convergent adaptation in Neanderthals and modern humans". Proceedings of the National Academy of Sciences. 114 (47): 12442–12447. Bibcode:2017PNAS..11412442D. doi:10.1073/pnas.1703790114. PMC 5703271. PMID 29087302.
- ^ Rae, T. C.; Koppe, T.; Stringer, C. B. (2011). "The Neanderthal face is not cold adapted". Journal of Human Evolution. 60 (2): 234–239. Bibcode:2011JHumE..60..234R. doi:10.1016/j.jhevol.2010.10.003. PMID 21183202.
- ^ Jump up to: a b c d Helmuth, H. (1998). "Body height, body mass and surface area of the Neanderthals". Zeitschrift für Morphologie und Anthropologie. 82 (1): 1–12. doi:10.1127/zma/82/1998/1 (inactive March 6, 2024). JSTOR 25757530. PMID 9850627.
{{cite journal}}: CS1 maint: DOI inactive as of March 2024 (link) - ^ Stringer, C. (1984). "Human evolution and biological adaptation in the Pleistocene". In Foley, R. (ed.). Hominid evolution and community ecology. Academic Press. ISBN 978-0-12-261920-5.
- ^ Jump up to: a b Holloway, R. L. (1985). "The poor brain of Homo sapiens neanderthalensis: see what you please". In Delson, E. (ed.). Ancestors: The hard evidence. Alan R. Liss. ISBN 978-0-471-84376-4.
- ^ Jump up to: a b c Amano, H.; Kikuchi, T.; Morita, Y.; Kondo, O.; Suzuki, H.; et al. (2015). "Virtual reconstruction of the Neanderthal Amud 1 cranium" (PDF). American Journal of Physical Anthropology. 158 (2): 185–197. doi:10.1002/ajpa.22777. hdl:10261/123419. PMID 26249757. S2CID 36974955.
- ^ Jump up to: a b Beals, K.; Smith, C.; Dodd, S. (1984). "Brain size, cranial morphology, climate, and time machines" (PDF). Current Anthropology. 12 (3): 301–30. doi:10.1086/203138. S2CID 86147507.
- ^ Bruner, Emiliano (2004). "Geometric Morphometrics and Paleoneurology: Brain Shape Evolution in the Genus Homo". Journal of Human Evolution. 47 (5): 279–303. Bibcode:2004JHumE..47..279B. doi:10.1016/j.jhevol.2004.03.009. PMID 15530349.
- ^ Bruner, Emiliano (2010). "Morphological Differences in the Parietal Lobes with the Human Genus: A Neurofunctional Perspective". Current Anthropology. 51 (1): S77–S88. doi:10.1086/650729. S2CID 146587487.
- ^ Bruner, Emiliano; Manzi, Giorgio; Arsuaga, Juan Luis (2003). "Encephalization and Allometric Trajectories in the Genus Homo: Evidence from the Neandertal and Modern Lineages". Proceedings of the National Academy of Sciences of the United States of America. 100 (26): 15335–15340. Bibcode:2003PNAS..10015335B. doi:10.1073/pnas.2536671100. PMC 307568. PMID 14673084.
- ^ Hublin, Jean-Jacques; Neubauer, Simon; Gunz, Philipp (2015). "Brain Ontogeny and Life History in Pleistocene Hominins". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1663): 1–11. doi:10.1098/rstb.2014.0062. PMC 4305163. PMID 25602066. S2CID 1518695.
- ^ Weaver, Anne (2005). "Reciprocal Evolution of the Cerebellum and Neocortex in Fossil Humans". Proceedings of the National Academy of Sciences of the United States of America. 102 (10): 3576–3580. Bibcode:2005PNAS..102.3576W. doi:10.1073/pnas.0500692102. PMC 553338. PMID 15731345.
- ^ Jump up to: a b c Bruner, Emiliano; Ogihara, Naomichi; Tanabe, Hiroki C. (December 28, 2017). Digital Endocasts: From Skulls to Brains. Springer. p. 25. ISBN 978-4-431-56582-6.
- ^ Nishiaki, Yoshihiro; Jöris, Olaf (November 27, 2019). Learning Among Neanderthals and Palaeolithic Modern Humans: Archaeological Evidence. Springer Nature. p. 19. ISBN 978-981-13-8980-1. "indicates that the volumes of the parietal cortex and cerebellum in the Neanderthal brain seemed to be smaller than those in modern humans, while the region from the posterior temporal to occipital cortex seemed to be larger in Neanderthals..."
- ^ Jump up to: a b c d Juric, I.; Aeschbacher, S.; Coop, G. (2016). "The strength of selection against Neanderthal introgression". PLOS Genetics. 12 (11): e1006340. doi:10.1371/journal.pgen.1006340. PMC 5100956. PMID 27824859.
- ^ Jump up to: a b c Ruebens, K. (2013). "Regional behaviour among late Neanderthal groups in Western Europe: A comparative assessment of late Middle Palaeolithic bifacial tool variability". Journal of Human Evolution. 65 (4): 341–362. Bibcode:2013JHumE..65..341R. doi:10.1016/j.jhevol.2013.06.009. PMID 23928352.
- ^ Jump up to: a b c d Fabre, V.; Condemi, S.; Degioanni, A. (2009). "Genetic evidence of geographical groups among Neanderthals". PLOS ONE. 4 (4): e5151. Bibcode:2009PLoSO...4.5151F. doi:10.1371/journal.pone.0005151. PMC 2664900. PMID 19367332.
- ^ Jump up to: a b c Demay, L.; Péan, S.; Patou-Mathis, M. (2012). "Mammoths used as food and building resources by Neanderthals: zooarchaeological study applied to layer 4, Molodova I (Ukraine)" (PDF). Quaternary International. 276–277: 212–226. Bibcode:2012QuInt.276..212D. doi:10.1016/j.quaint.2011.11.019. hdl:2268/190618.
- ^ Jump up to: a b c d e f g Trinkaus, E. (1995). "Neanderthal mortality patterns". Journal of Archaeological Science. 22 (1): 121–142. Bibcode:1995JArSc..22..121T. doi:10.1016/S0305-4403(95)80170-7.
- ^ Jump up to: a b c d e Green, R. E.; Krause, J.; Briggs, A. W.; et al. (2010). "A draft sequence of the Neandertal genome". Science. 328 (5979): 710–722. Bibcode:2010Sci...328..710G. doi:10.1126/science.1188021. PMC 5100745. PMID 20448178.
- ^ Sankararaman, S.; Mallick, S.; Patterson, N.; Reich, D. (2016). "The combined landscape of Denisovan and Neanderthal ancestry in present-day humans". Current Biology. 26 (9): 1241–1247. Bibcode:2016CBio...26.1241S. doi:10.1016/j.cub.2016.03.037. PMC 4864120. PMID 27032491.
- ^ Jump up to: a b c d Sankararaman, S.; Mallick, S.; Dannemann, M.; Prüfer, K.; Kelso, J.; Pääbo, S.; Patterson, N.; Reich, D. (2014). "The genomic landscape of Neanderthal ancestry in present-day humans". Nature. 507 (7492): 354–357. Bibcode:2014Natur.507..354S. doi:10.1038/nature12961. PMC 4072735. PMID 24476815.
- ^ Jump up to: a b c d e Peyrégne, S.; Slon, V.; Mafessoni, F.; et al. (2019). "Nuclear DNA from two early Neandertals reveals 80 ka of genetic continuity in Europe". Science Advances. 5 (6): eaaw5873. Bibcode:2019SciA....5.5873P. doi:10.1126/sciadv.aaw5873. PMC 6594762. PMID 31249872.
- ^ Jump up to: a b Kuhlwilm, M. (2016). "Ancient gene flow from early modern humans into eastern Neanderthals". Nature. 530 (7591): 429–433. Bibcode:2016Natur.530..429K. doi:10.1038/nature16544. PMC 4933530. PMID 26886800.
- ^ Jump up to: a b c d e f g Prüfer, K.; et al. (2014). "The complete genome sequence of a Neanderthal from the Altai Mountains". Nature. 505 (7481): 43–49. Bibcode:2014Natur.505...43P. doi:10.1038/nature12886. PMC 4031459. PMID 24352235.
- ^ Jump up to: a b c Pennisi, E. (2013). "More genomes from Denisova Cave show mixing of early human groups". Science. 340 (6134): 799. Bibcode:2013Sci...340..799P. doi:10.1126/science.340.6134.799. PMID 23687020.
- ^ Jump up to: a b Vernot, B.; Akey, J. M. (2014). "Resurrecting surviving Neandertal lineages from modern human genomes". Science. 343 (6174): 1017–1021. Bibcode:2014Sci...343.1017V. doi:10.1126/science.1245938. PMID 24476670. S2CID 23003860.
- ^ Jump up to: a b c Nédélec, Y.; Sanz, J.; Baharian, G.; et al. (2016). "Genetic ancestry and natural selection drive population differences in immune responses to pathogens". Cell. 167 (3): 657–669. doi:10.1016/j.cell.2016.09.025. PMID 27768889.
- ^ Jump up to: a b Ségurel, L.; Quintana-Murci, L. (2014). "Preserving immune diversity through ancient inheritance and admixture". Current Opinion in Immunology. 30: 79–84. doi:10.1016/j.coi.2014.08.002. PMID 25190608.
- ^ Zeberg, Hugo; Pääbo, Svante (March 2, 2021). "A genomic region associated with protection against severe COVID-19 is inherited from Neandertals". Proceedings of the National Academy of Sciences. 118 (9): e2026309118. Bibcode:2021PNAS..11826309Z. doi:10.1073/pnas.2026309118. ISSN 0027-8424. PMC 7936282. PMID 33593941. S2CID 231943450.
- ^ Zeberg, Hugo; Pääbo, Svante (September 30, 2020). "The major genetic risk factor for severe COVID-19 is inherited from Neanderthals". Nature. 587 (7835): 610–612. Bibcode:2020Natur.587..610Z. doi:10.1038/s41586-020-2818-3. hdl:21.11116/0000-0007-0F26-F. ISSN 1476-4687. PMID 32998156. S2CID 222148977.
- ^ Jump up to: a b Dolgova, O.; Lao, O. (2018). "Evolutionary and medical consequences of archaic introgression into modern human genomes". Genes. 9 (7): 358. doi:10.3390/genes9070358. PMC 6070777. PMID 30022013.
- ^ Jump up to: a b c d e f g h i j k Reich 2018.
- ^ Jump up to: a b c Schmitz, R. W.; Serre, D.; Bonani, G.; et al. (2002). "The Neandertal type site revisited: interdisciplinary investigations of skeletal remains from the Neander Valley, Germany". Proceedings of the National Academy of Sciences. 99 (20): 13342–13347. Bibcode:2002PNAS...9913342S. doi:10.1073/pnas.192464099. PMC 130635. PMID 12232049.
- ^ Howell, F. C. (1957). "The evolutionary significance of variation and varieties of 'Neanderthal' man". The Quarterly Review of Biology. 32 (4): 330–347. doi:10.1086/401978. JSTOR 2816956. PMID 13506025. S2CID 10857962.
- ^ Jump up to: a b "Neandertal oder Neanderthal? Was ist denn nun richtig?" [Neandertal or Neanderthal? So which is actually right?]. Kreisstadt Mettmann. Retrieved February 1, 2017.
Heute sollten Ortsbezeichnungen das 'Neandertal' ohne 'h' bezeichnen. Alle Namen, die sich auf den prähistorischen Menschen beziehen, führen das 'h'. (Nowadays, place names should refer to the Neander Valley ['Neandertal'] without an 'h'. All names referring to the prehistoric humans have the 'h'.)
- ^ "Neanderthal". Collins English Dictionary. Retrieved February 18, 2020.
- ^ "Neanderthal". Merriam-Webster Dictionary. Retrieved February 18, 2020.
- ^ "Neanderthal". American Heritage Dictionary. Retrieved February 18, 2020.
- ^ Vogt, K. C. (1864). Lectures on man: his place in creation, and in the history of the earth. London, UK: Longman, Green, Longman and Roberts. pp. 302, 473.
- ^ King, W. (1864). "On the Neanderthal skull, or reasons for believing it to belong to the Clydian Period and to a species different from that represented by man". Report of the British Association for the Advancement of Science, Notices and Abstracts, Newcastle-upon-Tyne, 1863. 33: 81–82 – via Biodiversity Heritage Library.
- ^ Murray, J.; Nasheuer, H. P.; Seoighe, C.; McCormack, G. P.; Williams, D. M.; Harper, D. A. T. (2015). "The contribution of William King to the early development of palaeoanthropology". Irish Journal of Earth Sciences. 33: 1–16. doi:10.3318/ijes.2015.33.1. JSTOR 10.3318/ijes.2015.33.1. S2CID 131804686.
- ^ Winner, A. K. (1964). "Terminology". Current Anthropology. 5 (2): 119–122. doi:10.1086/200469. JSTOR 2739959. S2CID 224796921.
- ^ Jump up to: a b c King, W. (1864). "The reputed fossil man of the Neanderthal" (PDF). Quarterly Journal of Science. 1: 96.
- ^ Schmerling, P. (1834). Recherches sur les ossemens fossiles découverts dans les cavernes de la province de Liége [Research on the fossil specimens discovered in the caves of Liège]. P. J. Collardin. pp. 30–32. hdl:2268/207986.
- ^ Menez, A. (2018). "Custodian of the Gibraltar skull: the history of the Gibraltar Scientific Society". Earth Sciences History. 37 (1): 34–62. Bibcode:2018ESHis..37...34M. doi:10.17704/1944-6178-37.1.34.
- ^ Schaaffhausen, H. (1858). "Zur Kenntnis der ältesten Rassenschädel" [Acknowledging the oldest racial skull]. Archiv für Anatomie, Physiologie und Wissenschaftliche Medicin (in German): 453–478.
- ^ Jump up to: a b c d Schlager, S.; Wittwer-Backofen, U. (2015). "Images in Paleoanthropology: Facing Our Ancestors". In Henke, W.; Tattersall, I. (eds.). Handbook of Paleoanthropology. Springer-Verlag Berlin Heidelberg. pp. 1019–1027. doi:10.1007/978-3-642-39979-4_70. ISBN 978-3-642-39978-7.
- ^ Fuhlrott, J. C. (1859). "Menschliche Überreste aus einer Felsengrotte des Düsselthales" [Human remains from a rock grotto in Düsseltal] (PDF). Verh Naturhist Ver Preuss Rheinl (in German). 16: 131–153.
- ^ Virchow, R. (1872). "Untersuchung des Neanderthal-Schädels" [Examinations on the Neandertal skull]. Verh Berl Anthrop Ges (in German). 4: 157–165.
- ^ Boule, M. (1911). L'homme fossile de La Chapelle-aux-Saints [Fossil man from La Chapelle-aux-Saints] (in French). Masson. pp. 1–62.
- ^ Van Reybrouck, D. (2002). "Boule's error: on the social context of scientific knowledge". Antiquity. 76 (291): 158–164. doi:10.1017/S0003598X00089936. S2CID 164060946.
- ^ Jump up to: a b c d Langdon, J. H. (2016). "Case study 18. Neanderthals in the mirror: imagining our relatives". The science of human evolution: getting it right. Springer. ISBN 978-3-319-41584-0.
- ^ Sommer, M. (2006). "Mirror, mirror on the wall: Neanderthal as image and 'distortion' in early 20th-century French science and press" (PDF). Social Studies of Science. 36 (2): 207–240. doi:10.1177/0306312706054527. S2CID 145778787.
- ^ Cairney, C. T. (1989). Clans and families of Ireland and Scotland, an ethnography of the Gael. McFarland. p. 14. ISBN 978-0-89950-362-2.
- ^ Huxley, T. (1891). "The Aryan question and pre-historic man". The Popular Science Monthly. 38: 512–516.
- ^ Steensby, H. P. (1907). "Racestudier i Danmark" [Race Studies in Denmark] (PDF). Geographical Journal (in Danish). 9. Geografisk Tidsskrift.
- ^ Coon, C. S. (1962). "The origin of races". Science. 140 (3563). Knopf: 548–549. doi:10.1126/science.140.3563.208. PMID 14022816.
- ^ Tattersall, I.; Schwartz, J. H. (1999). "Hominids and hybrids: The place of Neanderthals in human evolution". Proceedings of the National Academy of Sciences. 96 (13): 7117–19. Bibcode:1999PNAS...96.7117T. doi:10.1073/pnas.96.13.7117. JSTOR 48019. PMC 33580. PMID 10377375.
- ^ Duarte, C.; Maurício, J.; Pettitt, P. B.; Souto, P.; Trinkaus, E.; van der Plicht, H.; Zilhão, J. (1999). "The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia". Proceedings of the National Academy of Sciences of the United States of America. 96 (13): 7604–7609. Bibcode:1999PNAS...96.7604D. doi:10.1073/pnas.96.13.7604. PMC 22133. PMID 10377462.
- ^ Jump up to: a b Hublin, J. J. (2009). "The origin of Neandertals". Proceedings of the National Academy of Sciences. 106 (38): 16022–16027. Bibcode:2009PNAS..10616022H. doi:10.1073/pnas.0904119106. JSTOR 40485013. PMC 2752594. PMID 19805257.
- ^ Harvati, K.; Frost, S. R.; McNulty, K. P. (2004). "Neanderthal taxonomy reconsidered: implications of 3D primate models of intra- and interspecific differences". Proceedings of the National Academy of Sciences. 101 (5): 1147–52. Bibcode:2004PNAS..101.1147H. doi:10.1073/pnas.0308085100. PMC 337021. PMID 14745010.
- ^ Soficaru, A.; Dobos, A.; Trinkaus, E. (2006). "Early modern humans from the Peștera Muierii, Baia de Fier, Romania". Proceedings of the National Academy of Sciences. 103 (46): 17196–17201. Bibcode:2006PNAS..10317196S. doi:10.1073/pnas.0608443103. JSTOR 30052409. PMC 1859909. PMID 17085588.
- ^ Jump up to: a b Chen, F.; Welker, F.; Shen, C. (2019). "A late Middle Pleistocene Denisovan mandible from the Tibetan Plateau" (PDF). Nature. 569 (7, 756): 409–412. Bibcode:2019Natur.569..409C. doi:10.1038/s41586-019-1139-x. PMID 31043746. S2CID 141503768.
- ^ Jump up to: a b c Pääbo, S. (2014). Neanderthal man: in search of lost genomes. New York: Basic Books. p. 237.
- ^ Hofreiter, M. (2011). "Drafting human ancestry: What does the Neanderthal genome tell us about hominid evolution? Commentary on Green et al. (2010)". Human Biology. 83 (1): 1–11. doi:10.3378/027.083.0101. PMID 21453001. S2CID 15005225.
- ^ Currat, M.; Excoffier, L. (2004). "Modern humans did not admix with Neanderthals during their range expansion into Europe". PLOS Biology. 2 (12): e421. doi:10.1371/journal.pbio.0020421. PMC 532389. PMID 15562317.
- ^ Mendez, F. L.; Poznik, G. D.; Castellano, S.; Bustamante, C. D. (2016). "The divergence of Neandertal and modern human Y chromosomes". American Journal of Human Genetics. 98 (4): 728–734. doi:10.1016/j.ajhg.2016.02.023. PMC 4833433. PMID 27058445.
- ^ Jump up to: a b Serre, D.; Langaney, A.; Chech, M.; Teschler-Nicola, M.; Paunovic, M.; Mennecier, P.; Hofreiter, M.; Possnert, G.; Pääbo, S. (2004). "No evidence of Neandertal mtDNA contribution to early modern humans". PLOS Biology. 2 (3): e57. doi:10.1371/journal.pbio.0020057. PMC 368159. PMID 15024415.
- ^ Jump up to: a b c Petr, Martin; Hajdinjak, Mateja; Fu, Qiaomei; Essel, Elena; Rougier, Hélène; Crevecoeur, Isabelle; Semal, Patrick; Golovanova, Liubov V.; Doronichev, Vladimir B.; Lalueza-Fox, Carles; de la Rasilla, Marco; Rosas, Antonio; Shunkov, Michael V.; Kozlikin, Maxim B.; Derevianko, Anatoli P. (September 25, 2020). "The evolutionary history of Neanderthal and Denisovan Y chromosomes". Science. 369 (6511): 1653–1656. Bibcode:2020Sci...369.1653P. doi:10.1126/science.abb6460. hdl:21.11116/0000-0007-11C2-A. ISSN 0036-8075. PMID 32973032.
- ^ Jump up to: a b c d Rogers, A. R.; Bohlender, R. J.; Huff, C. D. (2017). "Early history of Neanderthals and Denisovans". Proceedings of the National Academy of Sciences. 114 (37): 9859–9863. Bibcode:2017PNAS..114.9859R. doi:10.1073/pnas.1706426114. PMC 5604018. PMID 28784789.
- ^ Jump up to: a b c Hajdinjak, M.; Fu, Q.; Hübner, A. (2018). "Reconstructing the genetic history of late Neanderthals". Nature. 555 (7698): 652–656. Bibcode:2018Natur.555..652H. doi:10.1038/nature26151. PMC 6485383. PMID 29562232.
- ^ Jump up to: a b Posth, C.; Wißing, C.; Kitagawa, K.; et al. (2017). "Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals". Nature Communications. 8: 16046. Bibcode:2017NatCo...816046P. doi:10.1038/ncomms16046. PMC 5500885. PMID 28675384.
- ^ Jump up to: a b c Reilly, Patrick F.; Tjahjadi, Audrey; Miller, Samantha L.; Akey, Joshua M.; Tucci, Serena (September 2022). "The contribution of Neanderthal introgression to modern human traits". Current Biology. 32 (18): R970–R983. Bibcode:2022CBio...32.R970R. doi:10.1016/j.cub.2022.08.027. PMC 9741939. PMID 36167050.
- ^ Jump up to: a b c Dean, D.; Hublin, J.-J.; Holloway, R.; Ziegler, R. (1998). "On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany". Journal of Human Evolution. 34 (5): 485–508. Bibcode:1998JHumE..34..485D. doi:10.1006/jhev.1998.0214. PMID 9614635. S2CID 6996233.
- ^ Ko, K. W. (2016). "Hominin interbreeding and the evolution of human variation". Journal of Biological Research-Thessaloniki. 23: 17. doi:10.1186/s40709-016-0054-7. PMC 4947341. PMID 27429943.
- ^ Bischoff, J. L.; Shamp, D. D.; Aramburu, A.; et al. (2003). "The Sima de los Huesos hominids date to beyond U/Th equilibrium (>350kyr) and perhaps to 400–500kyr: new radiometric dates". Journal of Archaeological Science. 30 (3): 275–280. Bibcode:2003JArSc..30..275B. doi:10.1006/jasc.2002.0834.
- ^ Arsuaga, J. L.; Martínez, I.; Gracia, A.; Lorenzo, C. (1997). "The Sima de los Huesos crania (Sierra de Atapuerca, Spain). A comparative study". Journal of Human Evolution. 33 (2–3): 219–281. Bibcode:1997JHumE..33..219A. doi:10.1006/jhev.1997.0133. PMID 9300343.
- ^ Daura, J.; Sanz, M.; Arsuaga, J. L.; Hoffman, D. L. (2017). "New Middle Pleistocene hominin cranium from Gruta da Aroeira (Portugal)". Proceedings of the National Academy of Sciences. 114 (13): 3397–3402. Bibcode:2017PNAS..114.3397D. doi:10.1073/pnas.1619040114. PMC 5380066. PMID 28289213.
- ^ Stringer 1993, pp. 65–70.
- ^ Vandermeersch, B.; Garralda, M. D. (2011). "Neanderthal Geographical and Chronological Variation". In S. Condemi; G.-C. Weniger (eds.). Continuity and Discontinuity in the Peopling of Europe. Vertebrate Paleobiology and Paleoanthropology. Springer Netherlands. pp. 113–125. doi:10.1007/978-94-007-0492-3_10. ISBN 978-94-007-0491-6.
- ^ Richter, J. (2011). "When Did the Middle Paleolithic Begin?". In Conard, N. J.; Richter, J. (eds.). Neanderthal Lifeways, Subsistence and Technology. Vertebrate paleobiology and paleoanthropology. Vol. 19. Springer. pp. 7–14. doi:10.1007/978-94-007-0415-2_2. ISBN 978-94-007-0414-5.
- ^ Zanolli, C.; Martinón-Torres, M.; Bernardini, F.; et al. (2018). "The Middle Pleistocene (MIS 12) human dental remains from Fontana Ranuccio (Latium) and Visogliano (Friuli-Venezia Giulia), Italy. A comparative high resolution endostructural assessment". PLOS ONE. 13 (10): e0189773. Bibcode:2018PLoSO..1389773Z. doi:10.1371/journal.pone.0189773. PMC 6169847. PMID 30281595.
- ^ Jump up to: a b c Endicott, P.; Ho, S. Y. W.; Stringer, C. (2010). "Using genetic evidence to evaluate four palaeoanthropological hypotheses for the timing of Neanderthal and modern human origins" (PDF). Journal of Human Evolution. 59 (1): 87–95. Bibcode:2010JHumE..59...87E. doi:10.1016/j.jhevol.2010.04.005. PMID 20510437.
- ^ Welker, F.; et al. (2020). "The dental proteome of Homo antecessor". Nature. 580 (7802): 235–238. Bibcode:2020Natur.580..235W. doi:10.1038/s41586-020-2153-8. PMC 7582224. PMID 32269345. S2CID 214736611.
- ^ Briggs, A. W.; Good, J. M.; Green, R. E. (2009). "Targeted retrieval and analysis of five Neandertal mtDNA genomes" (PDF). Science. 325 (5, 938): 318–321. Bibcode:2009Sci...325..318B. doi:10.1126/science.1174462. PMID 19608918. S2CID 7117454. Archived from the original (PDF) on March 7, 2019.
- ^ Hajdinjak, M.; Fu, Q.; Hübner, A.; et al. (March 1, 2018). "Reconstructing the genetic history of late Neanderthals". Nature. 555 (7698): 652–656. Bibcode:2018Natur.555..652H. doi:10.1038/nature26151. ISSN 1476-4687. PMC 6485383. PMID 29562232.
- ^ Jump up to: a b Krings, M.; Stone, A.; Schmitz, R. W.; Krainitzki, H.; Stoneking, M.; Pääbo, S. (1997). "Neandertal DNA sequences and the origin of modern humans". Cell. 90 (1): 19–30. doi:10.1016/s0092-8674(00)80310-4. hdl:11858/00-001M-0000-0025-0960-8. PMID 9230299. S2CID 13581775.
- ^ Krings, M.; Geisert, H.; Schmitz, R. W.; Krainitzki, H.; Pääbo, S. (1999). "DNA sequence of the mitochondrial hypervariable region II from the Neandertal type specimen". Proceedings of the National Academy of Sciences. 96 (10): 5581–5585. Bibcode:1999PNAS...96.5581K. doi:10.1073/pnas.96.10.5581. PMC 21903. PMID 10318927.
- ^ Green, R. E.; Malaspinas, A. S.; Krause, J.; Briggs, A. W.; et al. (2008). "A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing". Cell. 134 (3): 416–426. doi:10.1016/j.cell.2008.06.021. PMC 2602844. PMID 18692465.
- ^ Sawyer, S.; Renaud, G.; Viola, B.; Hublin, J. J. (2015). "Nuclear and mitochondrial DNA sequences from two Denisovan individuals". Proceedings of the National Academy of Sciences. 112 (51): 15696–15700. Bibcode:2015PNAS..11215696S. doi:10.1073/pnas.1519905112. PMC 4697428. PMID 26630009.
- ^ Jump up to: a b Rogers, A. R.; Harris, N. S.; Achenbach, A. A. (2020). "Neanderthal-Denisovan ancestors interbred with a distantly related hominin". Science Advances. 6 (8): eaay5483. Bibcode:2020SciA....6.5483R. doi:10.1126/sciadv.aay5483. PMC 7032934. PMID 32128408.
- ^ Serangeli, J.; Bolus, M. (2008). "Out of Europe - The dispersal of a successful European hominin form" (PDF). Quartär. 55: 83–98. Archived from the original (PDF) on February 29, 2020. Retrieved October 11, 2022.
- ^ French 2021, p. 133.
- ^ Callander, J. (2004). "Dorothy Garrod's excavations in the Late Mousterian of Shukbah Cave in Palestine reconsidered". Proceedings of the Prehistoric Society. 70: 207–231. doi:10.1017/S0079497X00001171. S2CID 191630165.
- ^ Smith, T. M.; Tafforeau, P.; Reid, D. J.; et al. (2007). "Earliest evidence of modern human life history in North African early Homo sapiens". Proceedings of the National Academy of Sciences of the United States of America. 104 (15): 6128–6133. Bibcode:2007PNAS..104.6128S. doi:10.1073/pnas.0700747104. PMC 1828706. PMID 17372199.
- ^ Douka, K.; J., Zenobia; Lane, C.; et al. (2014). "The chronostratigraphy of the Haua Fteah cave (Cyrenaica, northeast Libya)". Journal of Human Evolution. 66: 39–63. Bibcode:2014JHumE..66...39D. doi:10.1016/j.jhevol.2013.10.001. PMID 24331954.
- ^ Wu, X.-J.; Bruner, E. (2016). "The endocranial anatomy of Maba 1". American Journal of Physical Anthropology. 160 (4): 633–643. doi:10.1002/ajpa.22974. PMID 26972814.
- ^ Nielsen, T. K.; Benito, B. M.; et al. (2017). "Investigating Neanderthal dispersal above 55°N in Europe during the Last Interglacial Complex". Quaternary International. 431: 88–103. Bibcode:2017QuInt.431...88N. doi:10.1016/j.quaint.2015.10.039.
- ^ Nielsen, T. K.; Riede, F. (2018). "On research history and Neanderthal occupation at its northern margins". European Journal of Archaeology. 21 (4): 506–527. doi:10.1017/eaa.2018.12. S2CID 165849999.
- ^ Pavlov, P.; Roebroeks, W.; Svendsen, J. I. (2004). "The Pleistocene colonization of northeastern Europe: a report on recent research". Journal of Human Evolution. 47 (1–2): 3–17. Bibcode:2004JHumE..47....3P. doi:10.1016/j.jhevol.2004.05.002. PMID 15288521.
- ^ Jump up to: a b Slimak, L.; Svendsen, J. I.; Mangerud, J.; Plisson, H. (2011). "Late Mousterian persistence near the Arctic Circle". Science. 332 (6031): 841–845. Bibcode:2011Sci...332..841S. doi:10.1126/science.1203866. JSTOR 29784275. PMID 21566192. S2CID 24688365.
- ^ Slimak, L. (2012). "Response to "Comment on Late Mousterian persistence near the Arctic Circle"". Science. 335 (6065): 167. Bibcode:2012Sci...335..167S. doi:10.1126/science.1210211. PMID 22246757.
- ^ Jump up to: a b Zwyns, N. (2012). "Comment on Late Mousterian persistence near the Arctic Circle". Science. 335 (6065): 167. Bibcode:2012Sci...335..167Z. doi:10.1126/science.1209908. PMID 22246757.
- ^ Holan, S. R.; Deméré, T. A.; Fisher, D. C.; Fullager, R.; Paces, J. B.; Jefferson, G. T. (2017). "A 130,000-year-old archaeological site in southern California, USA". Nature. 544 (7651): 479–483. Bibcode:2017Natur.544..479H. doi:10.1038/nature22065. PMID 28447646. S2CID 205255425.
- ^ Sutton, M. Q.; Parkinson, J.; Rosen, M. D. (2019). "Observations regarding the Cerutti Mastodon". PaleoAmerica. 5 (1): 8–15. doi:10.1080/20555563.2019.1589409. S2CID 155596679.
- ^ Eren, M. I.; Bebber, M. R. (2019). "The Cerutti Mastodon site and experimental archaeology's quiet coming of age". Antiquity. 93 (369): 796–797. doi:10.15184/aqy.2019.50. S2CID 197853460.
- ^ Ferrel, P. M. (2019). "The Cerutti Mastodon site reinterpreted with reference to freeway construction plans and methods". PaleoAmerica. 5 (1): 1–7. doi:10.1080/20555563.2019.1589663. S2CID 167172979.
- ^ Jump up to: a b c d e f g h Bocquet-Appel, J.; Degioanni, A. (2013). "Neanderthal demographic estimates". Current Anthropology. 54: 202–214. doi:10.1086/673725. S2CID 85090309.
- ^ Pederzani, Sarah; Britton, Kate; Jones, Jennifer Rose; Agudo Pérez, Lucía; Geiling, Jeanne Marie; Marín-Arroyo, Ana B. (July 17, 2023). "Late Pleistocene Neanderthal exploitation of stable and mosaic ecosystems in northern Iberia shown by multi-isotope evidence". Quaternary Research. 116: 108–132. Bibcode:2023QuRes.116..108P. doi:10.1017/qua.2023.32. hdl:2164/21236. ISSN 0033-5894. Retrieved February 7, 2024.
- ^ Hershkovitz, I.; May, H.; Sarig, R.; et al. (2021). "A Middle Pleistocene Homo from Nesher Ramla, Israel". Science. 372 (6549): 1424–1428. Bibcode:2021Sci...372.1424H. doi:10.1126/science.abh3169. S2CID 235628111.
- ^ Jump up to: a b c Mafessoni, F.; Prüfer, K. (2017). "Better support for a small effective population size of Neandertals and a long shared history of Neandertals and Denisovans". Proceedings of the National Academy of Sciences. 114 (48): 10256–10257. Bibcode:2017PNAS..11410256M. doi:10.1073/pnas.1716918114. PMC 5715791. PMID 29138326.
- ^ Lalueza-Fox, C.; Sampietro, M. L.; Caramelli, D.; Puder, Y. (2013). "Neandertal evolutionary genetics: mitochondrial DNA data from the iberian peninsula". Molecular Biology and Evolution. 22 (4): 1077–1081. doi:10.1093/molbev/msi094. PMID 15689531.
- ^ Mellars, P.; French, J. C. (2011). "Tenfold population increase in Western Europe at the Neandertal-to-modern human transition". Science. 333 (6042): 623–627. Bibcode:2011Sci...333..623M. doi:10.1126/science.1206930. PMID 21798948. S2CID 28256970.
- ^ Jump up to: a b Trinkaus, E. (2011). "Late Pleistocene adult mortality patterns and modern human establishment". Proceedings of the National Academy of Sciences. 108 (4): 1267–1271. Bibcode:2011PNAS..108.1267T. doi:10.1073/pnas.1018700108. PMC 3029716. PMID 21220336.
- ^ Jump up to: a b Pettitt, R. B. (2000). "Neanderthal lifecycles: developmental and social phases in the lives of the last archaics". World Archaeology. 31 (3): 351–366. doi:10.1080/00438240009696926. JSTOR 125106. PMID 16475295. S2CID 43859422.
- ^ Jump up to: a b c Gómez-Olivencia, A.; Barash, A.; García-Martínez, D.; et al. (2018). "3D virtual reconstruction of the Kebara 2 Neandertal thorax". Nature Communications. 9 (4387): 4387. Bibcode:2018NatCo...9.4387G. doi:10.1038/s41467-018-06803-z. PMC 6207772. PMID 30377294.
- ^ Jump up to: a b c Trinkaus, E. (1981). "Neanderthal limb proportions and cold adaptation". In Stringer, C. B. (ed.). Aspects of human evolution. Taylor and Francis Ltd.
- ^ Formicola, V.; Giannecchini, M. (1998). "Evolutionary trends of stature in Upper Paleolithic and Mesolithic Europe". Journal of Human Evolution. 36 (3): 325. doi:10.1006/jhev.1998.0270. PMID 10074386.
- ^ Roser, M.; Appel, C.; Ritchie, H. (2013). "Human height". Our World in Data. Retrieved June 16, 2020.
- ^ Jump up to: a b c Duveau, J.; Berillon, G.; Verna, C.; Laisné, G.; Cliquet, D. (2019). "The composition of a Neandertal social group revealed by the hominin footprints at Le Rozel (Normandy, France)". Proceedings of the National Academy of Sciences. 116 (39): 19409–19414. Bibcode:2019PNAS..11619409D. doi:10.1073/pnas.1901789116. PMC 6765299. PMID 31501334.
- ^ Froehle, A. W.; Churchill, S. E. (2009). "Energetic competition between Neandertals and anatomically modern humans" (PDF). PaleoAnthropology: 96–116.
- ^ Gómez-Olivencia, A.; Been, E.; Arsuaga, J. L.; Stock, J. T. (2013). "The Neandertal vertebral column 1: the cervical spine". Journal of Human Evolution. 64 (6): 604–630. Bibcode:2013JHumE..64..608G. doi:10.1016/j.jhevol.2013.02.008. PMID 23541382.
- ^ García-Martínez, D.; Bastir, M.; Huguet, R. (2017). "The costal remains of the El Sidrón Neanderthal site (Asturias, northern Spain) and their importance for understanding Neanderthal thorax morphology". Journal of Human Evolution. 111: 85–101. Bibcode:2017JHumE.111...85G. doi:10.1016/j.jhevol.2017.06.003. hdl:10261/158977. PMID 28874276. S2CID 4634345.
- ^ Gómez-Olivencia, A.; Holliday, T.; Madelaine, M. (2018). "The costal skeleton of the Regourdou 1 Neandertal". Journal of Human Evolution. 130: 151–171. doi:10.1016/j.jhevol.2017.12.005. PMID 29496322.
- ^ "What is Lumbarization?". January 13, 2016.
- ^ Jump up to: a b Weaver, T. D. (2009). "The meaning of Neandertal skeletal morphology". Proceedings of the National Academy of Sciences. 106 (38): 16, 028–16, 033. doi:10.1073/pnas.0903864106. PMC 2752516. PMID 19805258.
- ^ Holliday, T. W. (1997). "Postcranial evidence of cold adaptation in European Neandertals". American Journal of Physical Anthropology. 104 (2): 245–258. doi:10.1002/(SICI)1096-8644(199710)104:2<245::AID-AJPA10>3.0.CO;2-#. PMID 9386830.
- ^ De Groote, I. (2011). "The Neanderthal lower arm". Journal of Human Evolution. 61 (4): 396–410. Bibcode:2011JHumE..61..396D. doi:10.1016/j.jhevol.2011.05.007. PMID 21762953.
- ^ Walker, M. J.; Ortega, J.; Parmová, K.; López, M. V.; Trinkaus, E. (2011). "Morphology, body proportions, and postcranial hypertrophy of a Neandertal female from the Sima de las Palomas, southeastern Spain". Proceedings of the National Academy of Sciences. 108 (25): 10, 087–10, 091. Bibcode:2011PNAS..10810087W. doi:10.1073/pnas.1107318108. PMC 3121844. PMID 21646528.
- ^ Raichlen, D.; Armstrong, H.; Lieberman, D. E. (2011). "Calcaneus length determines running economy: Implications for endurance running performance in modern humans and Neandertals". Journal of Human Evolution. 60 (3): 299–308. Bibcode:2011JHumE..60..299R. doi:10.1016/j.jhevol.2010.11.002. PMID 21269660.
- ^ Lee, S. S. M.; Piazza, S. J. (2009). "Built for speed: musculoskeletal structure and sprinting ability" (PDF). Journal of Experimental Biology. 212 (22): 3700–3707. doi:10.1242/jeb.031096. PMID 19880732. S2CID 10139786. Archived from the original (PDF) on December 14, 2019.
- ^ Gunz, P.; Tilot, A. K.; Wittfeld, K.; et al. (2019). "Neandertal introgression sheds light on modern human endocranial globularity". Current Biology. 29 (1): 120–127. Bibcode:2019CBio...29E.120G. doi:10.1016/j.cub.2018.10.065. PMC 6380688. PMID 30554901.
- ^ Gunz, P.; Harvati, K. (2007). "The Neanderthal "chignon": variation, integration, and homology". Journal of Human Evolution. 52 (3): 262–274. Bibcode:2007JHumE..52..262G. doi:10.1016/j.jhevol.2006.08.010. PMID 17097133.
- ^ Romagnoli, Rivals & Benazzi 2022, p. 75
- ^ Jump up to: a b c Pearce, E.; Stringer, C.; Dunbar, R. I. M. (2013). "New insights into differences in brain organization between Neanderthals and anatomically modern humans". Proceedings of the Royal Society B. 280 (1758): 20130168. doi:10.1098/rspb.2013.0168. PMC 3619466. PMID 23486442.
- ^ Finlayson 2019, p. 95: "Why did Neanderthals have such large eyes? A recent paper compared Neanderthal and modern human eyes and concluded that Neanderthals had invested in enhanced vision (assumed to correlate with eye size) at the expense of the development of the neocortex, which is so important to cognition.¹ Modern humans, on the other hand, had invested in the neocortex instead and this permitted them to live in larger groups and interact socially with a greater number of individuals than Neanderthals could "..." The paper was rebutted on methodological grounds and concluded that the larger orbits of Neanderthals compared to the modern human population could not permit any interpretation of cognitive ability based on group size."
- ^ Jump up to: a b c d Clement, A. F.; Hillson, S. W.; Aiello, L. C. (2012). "Tooth wear, Neanderthal facial morphology and the anterior dental loading hypothesis". Journal of Human Evolution. 62 (3): 367–376. Bibcode:2012JHumE..62..367C. doi:10.1016/j.jhevol.2011.11.014. PMID 22341317.
- ^ Holton, N. E.; Yokley, T. R.; Franciscus, R. G. (2011). "Climatic adaptation and Neandertal facial evolution: A comment on Rae et al. (2011)". Journal of Human Evolution. 61 (5): 624–627. Bibcode:2011JHumE..61..624H. doi:10.1016/j.jhevol.2011.08.001. PMID 21920585.
- ^ O'Connor, C. F.; Franciscus, R. G.; Holton, N. E. (2005). "Bite force production capability and efficiency in Neandertals and modern humans". American Journal of Physical Anthropology. 127 (2): 129–151. doi:10.1002/ajpa.20025. PMID 15558614.
- ^ Benazzi, S.; Nguyen, H. H.; Kullmer, O.; Hublin, J.-J. (2014). "Exploring the biomechanics of taurodontism". Journal of Anatomy. 226 (2): 180–188. doi:10.1111/joa.12260. PMC 4304574. PMID 25407030.
- ^ Takaki, P.; Vieira, M.; Bommarito, S. (2014). "Maximum bite force analysis in different age groups". International Archives of Otorhinolaryngology. 18 (3): 272–276. doi:10.1055/s-0034-1374647. PMC 4297017. PMID 25992105.
- ^ Allen, J. S.; Damasio, H.; Grabowski, T. J. (2002). "Normal neuroanatomical variation in the human brain: an MRI-volumetric study". American Journal of Physical Anthropology. 118 (4): 341–358. doi:10.1002/ajpa.10092. PMID 12124914. S2CID 21705705.
- ^ Balzeau, A.; Grimaud-Hervé, D.; Detroit, F.; Holloway, R. L. (2013). "First description of the Cro-Magnon 1 endocast and study of brain variation and evolution in anatomically modern Homo sapiens". Bulletins et Mémoires de la Société d anthropologie de Paris. 25 (1–2): 11–12. doi:10.1007/s13219-012-0069-z. S2CID 14675512.
- ^ Ponce de León, M. S.; Golovanova, L.; Doronichev, V. (2008). "Neanderthal brain size at birth provides insights into the evolution of human life history". Proceedings of the National Academy of Sciences. 105 (37): 13764–13768. doi:10.1073/pnas.0803917105. PMC 2533682. PMID 18779579.
- ^ Cartmill, Matt; Smith, Fred H. (September 20, 2011). The Human Lineage. John Wiley & Sons. p. 500. ISBN 978-1-118-21145-8. "The increased cranial breadth in Neandertals and its low position give the braincase a distinctive oval silhouette in rear view. This vault shape, conventionally described as “en bombe” (French, “bomb-like”), is regarded by some as a uniquely Neandertal apomorohy (Hublin 1980)."
- ^ Narayan, Sachindra (1999). Anthropological and Archaeological Research and Development. Gyan Publishing House. p. 50. ISBN 978-81-212-0619-8. "...the skull presents a typical rounded profile (called en bombe)..."
- ^ Dean, David; Hublin, Jean-Jacques; Holloway, Ralph; Ziegler, Reinhard (1998). "On the phylogenetic position of the pre-Neandertal specimen from Reilingen, Germany". Journal of Human Evolution. 34 (5): 485–508. Bibcode:1998JHumE..34..485D. doi:10.1006/jhev.1998.0214. PMID 9614635.
- ^ Buzi, Costantino; Di Vincenzo, Fabio; Profico, Antonio; Harvati, Katerina; Melchionna, Marina; Raia, Pasquale; Manzi, Giorgio (2021). "Retrodeformation of the Steinheim cranium: Insights into the evolution of Neanderthals". Symmetry. 13 (9): 1611. Bibcode:2021Symm...13.1611B. doi:10.3390/sym13091611. hdl:11573/1690670.
- ^ Jump up to: a b Hublin, Jean-Jacques; Neubauer, Simon; Gunz, Philipp (2015). "Brain Ontogeny and Life History in Pleistocene Hominins". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1663): 1–11. doi:10.1098/rstb.2014.0062. PMC 4305163. PMID 25602066.
- ^ Kochiyama, T.; Ogihara, N.; Tanabe, H. C. (2018). "Reconstructing the Neanderthal brain using computational anatomy". Scientific Reports. 8 (6296): 6296. Bibcode:2018NatSR...8.6296K. doi:10.1038/s41598-018-24331-0. PMC 5919901. PMID 29700382.
- ^ Bastir, Markus; Rosas, Antonio; Lieberman, Daniel E; O’Higgins, Paul (2008). "Middle Cranial Fossa Anatomy and the Origin of Modern Humans". The Anatomical Record. 291 (2): 130–140. doi:10.1002/ar.20636. PMID 18213701. S2CID 9755048.
- ^ Bastir, Markus; Rosas, Antonio; Gunz, Philipp; Peña-Melian, Angel; Manzi, Giorgio; Harvati, Katerina; Kruszynski, Robert; Stringer, Chris; Hublin, Jean-Jacques (2011). "Evolution of the Base of the Brain in Highly Encephalized Human Species". Nature Communications. 2 (2): 588. Bibcode:2011NatCo...2..588B. doi:10.1038/ncomms1593. hdl:10261/123641. PMID 22158443.
- ^ Gunz, Philipp; Maureille, Bruno; Hublin, Jean-Jacques (2010). "Brain Development after Birth Differs between Neanderthals and Modern Humans". Current Biology. 20 (21): R921–R922. Bibcode:2010CBio...20.R921G. doi:10.1016/j.cub.2010.10.018. PMID 21056830. S2CID 29295311.
- ^ Coolidge, Frederick L.; Wynn, Thomas; Overmann, Karenleigh A. (2024). "The Expert Neandertal Mind and Brain, Revisited". In Wynn, Thomas; Overmann, Karenleigh A.; Coolidge, Frederick L. (eds.). The Oxford Handbook of Cognitive Archaeology. Oxford University Press. pp. 131–149. ISBN 9780192895950.
- ^ Jump up to: a b c d Dannemann, M.; Kelso, J. (2017). "The contribution of Neanderthals to phenotypic variation in modern humans". The American Journal of Human Genetics. 101 (4): 578–589. doi:10.1016/j.ajhg.2017.09.010. PMC 5630192. PMID 28985494.
- ^ Allentoft, M. E.; Sikora, M. (2015). "Population genomics of Bronze Age Eurasia". Nature. 522 (7, 555): 167–172. Bibcode:2015Natur.522..167A. doi:10.1038/nature14507. PMID 26062507. S2CID 4399103.
- ^ Watson, Traci. "Were Some Neandertals Brown-Eyed Girls?". science.org. "According to this method, all three Neandertals had a dark complexion and brown eyes, and although one was red-haired, two sported brown locks"
- ^ Cerqueira, C. C.; Piaxão-Côrtes, V. R.; Zambra, F. M. B.; Hünemeier, T.; Bortolini, M. (2012). "Predicting Homo pigmentation phenotype through genomic data: From neanderthal to James Watson". American Journal of Human Biology. 24 (5): 705–709. doi:10.1002/ajhb.22263. PMID 22411106. S2CID 25853632.
- ^ Lalueza-Fox, C.; Rompler, H.; Caramelli, D.; et al. (2007). "A melanocortin 1 receptor allele suggests varying pigmentation among Neanderthals". Science. 318 (5855): 1453–1455. Bibcode:2007Sci...318.1453L. doi:10.1126/science.1147417. PMID 17962522. S2CID 10087710.
- ^ Smith, T. M.; Tafforeau, P.; Reid, D. J. (2010). "Dental evidence for ontogenetic differences between modern humans and Neanderthals". Proceedings of the National Academy of Sciences. 107 (49): 20923–20928. Bibcode:2010PNAS..10720923S. doi:10.1073/pnas.1010906107. PMC 3000267. PMID 21078988.
- ^ Guatelli-Steinberg, D. (2009). "Recent studies of dental development in Neandertals: Implications for Neandertal life histories". Evolutionary Biology. 18 (1): 9–20. doi:10.1002/evan.20190. S2CID 84273699.
- ^ Rosas, A.; Ríos, L.; Estalrrich, A.; et al. (2017). "The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain)". Science. 357 (6537): 1282–1287. Bibcode:2017Sci...357.1282R. doi:10.1126/science.aan6463. PMID 28935804.
- ^ DeSilva, J. M. (2018). "Comment on 'The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain)'". Science. 359 (6380): eaar3611. doi:10.1126/science.aar3611. PMID 29590012.
- ^ Froehle, A. W.; Churchill, S. E. (2009). "Energetic competition between Neanderthals and anatomically modern humans" (PDF). PaleoAnthropology: 96–116.
- ^ Jump up to: a b Snodgrass, J. J.; Leonard, W. R. (2009). "Neandertal energetics revisited: insights into population dynamics and life history evolution" (PDF). PaleoAnthropology: 220–237. doi:10.4207/PA.2009.ART31. S2CID 16044193. Archived from the original (PDF) on August 1, 2020.
- ^ Jump up to: a b c Hockett, B. (2012). "The consequences of Middle Paleolithic diets on pregnant Neanderthal women". Quaternary International. 264: 78–82. Bibcode:2012QuInt.264...78H. doi:10.1016/j.quaint.2011.07.002.
- ^ Taylor, J. S.; Reimchen, T. E. (2016). "Opsin gene repertoires in northern archaic hominids". Genome. 59 (8): 541–549. doi:10.1139/gen-2015-0164. hdl:1807/73317. PMID 27463216.
- ^ Jump up to: a b Nakahashi, W. (2017). "The effect of trauma on Neanderthal culture: A mathematical analysis". Homo. 68 (2): 83–100. doi:10.1016/j.jchb.2017.02.001. PMID 28238406.
- ^ Trinkaus, E.; Villotte, S (2017). "External auditory exostoses and hearing loss in the Shanidar 1 Neandertal". PLOS ONE. 12 (10): e0186684. Bibcode:2017PLoSO..1286684T. doi:10.1371/journal.pone.0186684. PMC 5650169. PMID 29053746.
- ^ Trinkaus, E.; Samsel, M.; Villotte, S. (2019). "External auditory exostoses among western Eurasian late Middle and Late Pleistocene humans". PLOS ONE. 14 (8): e0220464. Bibcode:2019PLoSO..1420464T. doi:10.1371/journal.pone.0220464. PMC 6693685. PMID 31412053.
- ^ Beier, J.; Anthes, N.; Wahl, J.; Harvati, K. (2018). "Similar cranial trauma prevalence among Neanderthals and Upper Palaeolithic modern humans". Nature. 563 (7733): 686–690. Bibcode:2018Natur.563..686B. doi:10.1038/s41586-018-0696-8. PMID 30429606. S2CID 53306963.
- ^ Trinkaus, E. (2012). "Neandertals, early modern humans, and rodeo riders". Journal of Archaeology. 39 (2): 3691–3693. Bibcode:2012JArSc..39.3691T. doi:10.1016/j.jas.2012.05.039.
- ^ Jump up to: a b Camarós, E.; Cueto, M.; Lorenzo, C.; Villaverde, V. (2016). "Large carnivore attacks on hominins during the Pleistocene: a forensic approach with a Neanderthal example". Archaeological and Anthropological Sciences. 8 (3): 635–646. Bibcode:2016ArAnS...8..635C. doi:10.1007/s12520-015-0248-1. hdl:10550/54275. S2CID 82001651.
- ^ Sánchez-Quinto, F.; Lalueza-Fox, C. (2015). "Almost 20 years of Neanderthal palaeogenetics: adaptation, admixture, diversity, demography and extinction". Philosophical Transactions of the Royal Society B. 370 (1660): 20130374. doi:10.1098/rstb.2013.0374. PMC 4275882. PMID 25487326.
- ^ Jump up to: a b c Ríos, L.; Kivell, T. L.; Lalueza-Fox, C.; Estalrrich, A. (2019). "Skeletal anomalies in the Neandertal family of El Sidrón (Spain) support a role of inbreeding in Neandertal extinction". Scientific Reports. 9 (1): 1697. Bibcode:2019NatSR...9.1697R. doi:10.1038/s41598-019-38571-1. PMC 6368597. PMID 30737446.
- ^ Haeusler, M.; Trinkaus, E.; Fornai, C.; Müller, J.; Bonneau, N.; Boeni, T.; Frater, N. (2019). "Morphology, pathology, and the vertebral posture of the La Chapelle-aux-Saints Neandertal". Proceedings of the National Academy of Sciences. 116 (11): 4923–4927. Bibcode:2019PNAS..116.4923H. doi:10.1073/pnas.1820745116. PMC 6421410. PMID 30804177.
- ^ Crubézy, E.; Trinkaus, E. (1992). "Shanidar 1: a case of hyperostotic disease (DISH) in the middle Paleolithic". American Journal of Physical Anthropology. 89 (4): 411–420. doi:10.1002/ajpa.1330890402. PMID 1463085.
- ^ Houldcroft, C. J.; Underdown, S. J. (2016). "Neanderthal genomics suggests a pleistocene time frame for the first epidemiologic transition" (PDF). American Journal of Physical Anthropology. 160 (3): 379–388. doi:10.1002/ajpa.22985. PMID 27063929.
- ^ Pimenoff, V. N.; de Oliveira, C. M.; Bravo, I. G. (2017). "Transmission between archaic and modern human ancestors during the evolution of the oncogenic human papillomavirus 16". Molecular Biology and Evolution. 34 (1): 4–19. doi:10.1093/molbev/msw214. PMC 5854117. PMID 28025273.
- ^ Jump up to: a b c d Weyrich, L. S.; Duchene, S.; Soubrier, J.; et al. (2017). "Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus". Nature. 544 (7650): 357–361. Bibcode:2017Natur.544..357W. doi:10.1038/nature21674. hdl:10261/152016. PMID 28273061. S2CID 4457717.
- ^ Fennel, K. J.; Trinkaus, E. (1997). "Bilateral femoral and tibial periostitis in the La Ferrassie 1 Neanderthal". Journal of Archaeological Science. 24 (11): 985–995. Bibcode:1997JArSc..24..985F. doi:10.1006/jasc.1996.0176.
- ^ Soltysiak, A. (2012). "Comment: low dental caries rate in Neandertals: the result of diet or the oral flora composition?". Homo. 63 (2): 110–113. doi:10.1016/j.jchb.2012.02.001. PMID 22409830.
- ^ Jump up to: a b c Smith, T. A.; Austin, C.; Green, D. R.; et al. (2018). "Wintertime stress, nursing, and lead exposure in Neanderthal children". Science Advances. 4 (10): eaau9483. Bibcode:2018SciA....4.9483S. doi:10.1126/sciadv.aau9483. PMC 6209393. PMID 30402544.
- ^ Rosas, A.; Estalrrich, A.; García-Vargas, S.; García-Tabernero, A.; Huguet, R.; Lalueza-Fox, C.; de la Rasilla, M. (2013). "Identification of Neandertal individuals in fragmentary fossil assemblages by means of tooth associations: The case of El Sidrón (Asturias, Spain)". Comptes Rendus Palevol. 12 (5): 279–291. Bibcode:2013CRPal..12..279R. doi:10.1016/j.crpv.2013.06.003.
- ^ Jump up to: a b c d e Spikins, P.; Hitchens, G.; Needham, A.; et al. (2014). "The Cradle of Thought: Growth, Learning, Play and Attachment in Neanderthal Children" (PDF). Oxford Journal of Archaeology. 33 (2): 111–134. doi:10.1111/ojoa.12030.
- ^ Farizy, C. (1994). "Spatial patterning of Middle Paleolithic sites". Journal of Anthropological Archaeology. 13 (2): 153–160. doi:10.1006/jaar.1994.1010.
- ^ Torres, Trinidad; Ortiz, José E.; Cobo, Rafael; de Hoz, Pedro; García-Redondo, Ana; Grün, Rainer (January 1, 2007). "Hominid exploitation of the environment and cave bear populations. The case of Ursus spelaeus Rosenmüller-Heinroth in Amutxate cave (Aralar, Navarra-Spain)". Journal of Human Evolution. 52 (1): 1–15. Bibcode:2007JHumE..52....1T. doi:10.1016/j.jhevol.2006.07.013. ISSN 0047-2484. PMID 16996576. Retrieved February 25, 2024 – via Elsevier Science Direct.
- ^ Fortes, G. G.; Grandal-d'Anglade, A.; Kolbe, B. (2016). "Ancient DNA reveals differences in behaviour and sociality between brown bears and extinct cave bears" (PDF). Molecular Ecology. 25 (19): 4907–4918. Bibcode:2016MolEc..25.4907F. doi:10.1111/mec.13800. PMID 27506329. S2CID 18353913.
- ^ Stiller, M.; Baryshnikov, G.; Bocherens, H. (2010). "Withering away—25,000 years of genetic decline preceded cave bear extinction". Molecular Biology and Evolution. 27 (5): 975–978. doi:10.1093/molbev/msq083. PMID 20335279.
- ^ Trájer, Attila J. (June 15, 2023). "Ecological evaluation of the development of Neanderthal niche exploitation". Quaternary Science Reviews. 310: 108127. Bibcode:2023QSRv..31008127T. doi:10.1016/j.quascirev.2023.108127. ISSN 0277-3791.
- ^ Ekshtain, R.; Malinsky-Buller, A.; Greenbaum, N.; et al. (2019). "Persistent Neanderthal occupation of the open-air site of 'Ein Qashish, Israel". PLOS ONE. 14 (6): e0215668. Bibcode:2019PLoSO..1415668E. doi:10.1371/journal.pone.0215668. PMC 6594589. PMID 31242180.
- ^ Бин, Э.; Ховерс, Э.; Экштайн, Р. (2017). «Останки первого неандертальца из стоянки среднего палеолита под открытым небом в Леванте» . Научные отчеты . 7 (2958): 2958. Бибкод : 2017НатСР...7.2958Б . дои : 10.1038/s41598-017-03025-z . ПМК 5462778 . ПМИД 28592838 .
- ^ Циммер, Карл (19 октября 2022 г.). «Первая известная семья неандертальцев, найденная в русской пещере» . Нью-Йорк Таймс . ISSN 0362-4331 . Проверено 21 октября 2022 г.
- ^ Сков, Лауритс; Пейрень, Стефан; Попли, Дивьяратхан; Яссы, Леонардо Н.М.; Девьезе, Тибо; Слоан, Вивиан; Завала, Елена Ивановна; Хайдиняк, Мэтью; Шумер, Арев П.; Гроте, Штеффи; Боссомс Меса, Альба; Лопес Эрраес, Дэвид; Никель, Биргит; Нагель, Сара; Рихтер, Юлия (октябрь 2022 г.). «Генетический взгляд на социальную организацию неандертальцев» . Природа . 610 (7932): 519–525. Бибкод : 2022Nature.610..519S . дои : 10.1038/s41586-022-05283-y . ISSN 1476-4687 . ПМЦ 9581778 . ПМИД 36261548 .
- ^ Нельсон, Эмма; Ролиан, Кэмпбелл; Кэшмор, Лиза; Шульц, Сюзанна (3 ноября 2010 г.). «Соотношение цифр предсказывает полигинию у ранних обезьян, ардипитеков, неандертальцев и ранних современных людей, но не у австралопитеков» . Труды Королевского общества B: Биологические науки . 278 (1711): 1556–1563. дои : 10.1098/rspb.2010.1740 . ISSN 0962-8452 . ПМК 3081742 . ПМИД 21047863 .
- ^ Лалуэса-Фокс, К.; Росас, А.; Эсталррих, А. (2011). «Генетические доказательства патрилокального брачного поведения среди групп неандертальцев» . Труды Национальной академии наук . 108 (1): 250–253. дои : 10.1073/pnas.1011553108 . ПМК 3017130 . ПМИД 21173265 .
- ^ Пирс, Э.; Мусиу, Т. (2015). «Использование расстояний передачи обсидиана для изучения поддержания социальных сетей у охотников-собирателей позднего плейстоцена» . Журнал антропологии и археологии . 36 : 12–20. дои : 10.1016/j.jaa.2014.07.002 . ПМК 4157217 . ПМИД 25214705 .
- ^ Перейти обратно: а б Росас, А.; Бастир, М.; Мартинес-Маса, К. (2006). «Палеобиология и сравнительная морфология образца позднего неандертальца из Эль-Сидрона, Астурия, Испания» . Труды Национальной академии наук . 103 (51): 19266–19271. Бибкод : 2006PNAS..10319266R . дои : 10.1073/pnas.0609662104 . ПМК 1748215 . ПМИД 17164326 .
- ^ Перейти обратно: а б Колобова К.А.; Робертс, Р.Г.; Чабай, вице-президент; и др. (2020). «Археологические свидетельства двух отдельных расселений неандертальцев в южной Сибири» . ПНАС . 117 (6): 2979–2885. Бибкод : 2020PNAS..117.2879K . дои : 10.1073/pnas.1918047117 . ПМК 7022189 . ПМИД 31988114 .
- ^ Пицин А.; Хайдиняк, М.; Новачевска, В. (2020). «Новые взгляды на расселение и перемещение неандертальцев из пещеры Стайня (Польша)» . Научные отчеты . 10 (14778): 14778. Бибкод : 2020NatSR..1014778P . дои : 10.1038/s41598-020-71504-x . ПМЦ 7479612 . ПМИД 32901061 .
- ^ Золликофер, CPE; Понсе де Леон, MS; Вандермерш, Б.; Левек, Ф. (2002). «Доказательства межличностного насилия у неандертальца Святого Сезара» . Труды Национальной академии наук . 99 (9): 6444–6448. Бибкод : 2002PNAS...99.6444Z . дои : 10.1073/pnas.082111899 . ПМК 122968 . ПМИД 11972028 .
- ^ Черчилль, SE; Франциск, Р.Г.; Маккин-Пераса, штат ХА; Дэниел, Дж.А.; Уоррен, БР (2009). «Колотое ребро Шанидара 3 неандертальца и палеолитическое вооружение». Журнал эволюции человека . 57 (2): 163–178. Бибкод : 2009JHumE..57..163C . дои : 10.1016/j.jhevol.2009.05.010 . ПМИД 19615713 .
- ^ Эсталррих, А.; Росас, А. (2015). «Разделение труда по полу и возрасту у неандертальцев: подход на основе изучения износа зубов, связанного с деятельностью». Журнал эволюции человека . 80 : 51–63. Бибкод : 2015JHumE..80...51E . дои : 10.1016/j.jhevol.2014.07.007 . ПМИД 25681013 .
- ^ Перейти обратно: а б с Камарос, Э.; Куэто, М.; Тейра, Л.; Мюнцель, Южная Каролина (2017). «Медведи на сцене: сложные взаимодействия плейстоцена с последствиями, касающимися изучения поведения неандертальцев» . Четвертичный интернационал . 435 : 237–246. Бибкод : 2017QuInt.435..237C . дои : 10.1016/j.quaint.2015.11.027 .
- ^ Бохеренс, Х.; Друкер, генеральный директор; Биллиу, Д.; Пату-Матис, М.; Вандермеерш, Б. (2005). «Изотопные данные о диете и образе жизни неандертальца Сен-Сезера I: обзор и использование модели смешивания из нескольких источников». Журнал эволюции человека . 49 (1): 71–87. Бибкод : 2005JHumE..49...71B . дои : 10.1016/j.jhevol.2005.03.003 . ПМИД 15869783 .
- ^ Джауэн, К.; и др. (2019). «Исключительно высокие значения δ15N в отдельных аминокислотах коллагена подтверждают, что неандертальцы были хищниками высокого трофического уровня» . Труды Национальной академии наук . 116 (11): 4928–4933. Бибкод : 2019PNAS..116.4928J . дои : 10.1073/pnas.1814087116 . ПМК 6421459 . ПМИД 30782806 .
- ^ Скотт, К. (1980). «Два охотничьих эпизода среднего палеолита в Ла-Котт-де-Сен-Брелад, Джерси (Нормандские острова)». Мировая археология . 12 (2): 137–152. дои : 10.1080/00438243.1980.9979788 .
- ^ Перейти обратно: а б Скотт, Б.; Бейтс, М.; Бейтс, ЧР; Коннеллер, К. (2015). «Новый вид из Ла-Котт-де-Сент-Брелад, Джерси» . Античность . 88 (339): 13–29. дои : 10.1017/S0003598X00050195 . hdl : 11858/00-001M-0000-002E-32BE-E .
- ^ Перейти обратно: а б Марин, Дж.; Саладье, П.; Родригес-Идальго, А.; Карбонелл, Э. (2017). «Стратегии охоты неандертальцев, выведенные из профилей смертности в последовательности Abric Romaní» . ПЛОС ОДИН . 12 (11): e0186970. Бибкод : 2017PLoSO..1286970M . дои : 10.1371/journal.pone.0186970 . ПМК 5699840 . ПМИД 29166384 .
- ^ Уайт, Марк; Петтитт, Пол; Шреве, Даниэль (15 мая 2016 г.). «Сначала стреляйте, потом задавайте вопросы: интерпретирующие рассказы об охоте неандертальцев» . Четвертичные научные обзоры . 140 : 1–20. Бибкод : 2016QSRv..140....1W . doi : 10.1016/j.quascirev.2016.03.004 . ISSN 0277-3791 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Саласар-Гарсия, Доминго К.; Пауэр, Роберт С.; Санчис Серра, Альфред; Вильяверде, Валентин; Уокер, Майкл Дж.; Генри, Аманда Г. (18 декабря 2013 г.). «Питание неандертальцев в центральном и юго-восточном Средиземноморье Иберии» . Четвертичный интернационал . Палеолитическая экодинамика на юге Иберии. 318 : 3–18. Бибкод : 2013QuInt.318....3S . дои : 10.1016/j.quaint.2013.06.007 . hdl : 10550/42242 . ISSN 1040-6182 . Получено 11 февраля 2024 г. - через Elsevier Science Direct.
- ^ Перейти обратно: а б Эль Заатари, С.; Грин, FE; Унгар, PS; Хублин, Ж.-Ж. (2016). «Реакция питания неандертальцев и современного человека на климатические колебания» . ПЛОС ОДИН . 11 (4): e0153277. Бибкод : 2016PLoSO..1153277E . дои : 10.1371/journal.pone.0153277 . ПМЦ 4847867 . ПМИД 27119336 .
- ^ Харди, Б.Л. (2010). «Климатическая изменчивость и распространение растительной пищи в плейстоценовой Европе: последствия для диеты и существования неандертальцев». Четвертичные научные обзоры . 29 (5–6): 662–679. Бибкод : 2010QSRv...29..662H . doi : 10.1016/j.quascirev.2009.11.016 .
- ^ Бак, Лейтенант; Стрингер, CB (2014). «Иметь на это смелость: вклад в диету неандертальцев?» (PDF) . Четвертичные научные обзоры . 96 : 161–167. Бибкод : 2014QSRv...96..161B . doi : 10.1016/j.quascirev.2013.09.003 .
- ^ Каррион, Хосе С.; Уокер, Майкл Дж. (1 августа 2019 г.). «Предыстория присутствия неандертальцев в Западно-Средиземноморской Европе» . Четвертичные научные обзоры . Неандертальцы: экология и эволюция. 217 : 7–44. Бибкод : 2019QSRv..217....7C . дои : 10.1016/j.quascirev.2018.10.011 . ISSN 0277-3791 . S2CID 135133190 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Перейти обратно: а б Дожер, К.; Веттезе, Д.; Бриттон, К.; Беарес, П.; Бульбес, Н.; Крегу-Боннур, Э.; Дескло, Э.; Латер, Н.; Пайк-Тэй, А.; Соперники, Ф.; Аллюэ, Э.; Чакон, Миннесота; Пуо, С.; Ричард, М.; Корти, Массачусетс; Галлотти, Р.; Харди, Б.; Бахейн, Джей-Джей; Фальгер, К.; Понс-Бранчу, Э.; Валладас, Х.; Монсель, М.-Х. (1 марта 2019 г.). «Неандертальская выборочная охота на оленей? На примере Абри дю Мара (юго-восток Франции)» . Археологические и антропологические науки . 11 (3): 985–1011. Бибкод : 2019АрАнС..11..985Д . дои : 10.1007/s12520-017-0580-8 . hdl : 2164/11676 . ISSN 1866-9565 . S2CID 134607529 . Проверено 25 февраля 2024 г.
- ^ Перейти обратно: а б с Ливраги, Алессандра; Фанфарилло, Габриэле; Колле, Маурицио Даль; Романдини, Маттео; Пересани, Марко (10 июня 2021 г.). «Экология неандертальцев и эксплуатация оленей и быков в начале MIS4: исследование пещеры Де Надале, Италия» . Четвертичный интернационал . 586 : 24–41. Бибкод : 2021QuInt.586...24L . дои : 10.1016/j.quaint.2019.11.024 . hdl : 11392/2414685 . S2CID 213340432 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Ираведра, Хосе; Кобо-Санчес, Люсия (1 января 2015 г.). «Неандертальская эксплуатация козлов и серн в юго-западной Европе» . Журнал эволюции человека . 78 : 12–32. Бибкод : 2015JHumE..78...12Y . дои : 10.1016/j.jhevol.2014.10.002 . ISSN 0047-2484 . ПМИД 25481629 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Терлато, Габриэле; Ливраги, Алессандра; Романдини, Маттео; Пересани, Марко (1 июня 2019 г.). «Крупные быки в меню неандертальцев: эксплуатация Bison priscus и Bos primigenius на северо-востоке Италии» . Журнал археологической науки . 25 : 129–143. Бибкод : 2019JArSR..25..129T . дои : 10.1016/j.jasrep.2019.04.006 . ISSN 2352-409X . S2CID 146317863 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Ливраги, Алессандра; Фанфарилло, Габриэле; Колле, Маурицио Даль; Романдини, Маттео; Пересани, Марко (июнь 2021 г.). «Экология неандертальцев и эксплуатация оленей и быков в начале MIS4: исследование пещеры Де Надале, Италия» . Четвертичный интернационал . 586 : 24–41. Бибкод : 2021QuInt.586...24L . дои : 10.1016/j.quaint.2019.11.024 . hdl : 11392/2414685 .
- ^ Жермонпре, Митье; Удреску, Мирча; Фирс, Эвелин (9 июля 2014 г.). «Возможные свидетельства охоты на мамонтов на стоянке неандертальцев Шпи (Бельгия)» . Четвертичный интернационал . Экологическая и культурная динамика в Западной и Центральной Европе в верхнем плейстоцене. 337 : 28–42. Бибкод : 2014QuInt.337...28G . дои : 10.1016/j.quaint.2012.10.035 . ISSN 1040-6182 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Гаудзински-Виндхойзер, Сабина; Киндлер, Лутц; Макдональд, Кэтрин; Робрукс, Уил (3 февраля 2023 г.). «Охота и обработка слонов с прямыми бивнями 125 000 лет назад: последствия для поведения неандертальцев» . Достижения науки . 9 (5): eadd8186. Бибкод : 2023SciA....9D8186G . дои : 10.1126/sciadv.add8186 . ISSN 2375-2548 . ПМЦ 9891704 . ПМИД 36724231 .
- ^ Бочеренс, Эрве; Друкер, Дороти Г.; Биллиу, Дэниел; Пату-Матис, Мэрилен; Вандермерш, Бернард (1 июля 2005 г.). «Изотопные данные о диете и образе жизни неандертальца Сен-Сезера I: обзор и использование модели смешивания из нескольких источников» . Журнал эволюции человека . 49 (1): 71–87. Бибкод : 2005JHumE..49...71B . дои : 10.1016/j.jhevol.2005.03.003 . ISSN 0047-2484 . ПМИД 15869783 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Братлунд, Б. 1999. Возвращение к Таубаху . Ежегодник Римско-германского центрального музея Майнца 46: 61-174.
- ^ Моклан, Абель; Юге, Роза; Маркес, Белен; Лаплана, Цезарь; Галиндо-Пеллисена, Мария Анхелес; Гарсия, Нурия; Блен, Хьюг-Александр; Альварес-Лао, Диего Дж.; Арсуага, Хуан Луис; Перес-Гонсалес, Альфредо; Бакедано, Энрике (октябрь 2021 г.). «Охотничий лагерь неандертальцев в центральной системе Пиренейского полуострова: зооархеологический и тафономический анализ каменного убежища Навальмайо (Пинилья-дель-Валле, Испания)» . Четвертичные научные обзоры . 269 : 107142. Бибкод : 2021QSRv..26907142M . doi : 10.1016/j.quascirev.2021.107142 .
- ^ Фиоренца, Л.; Бенацци, С.; Тауш, Дж.; Куллмер, О.; Бромедж, Т.Г.; Шренк, Ф. (2011). Розенберг, Карен (ред.). «Молярная макроодежда раскрывает эколого-географические различия в питании неандертальцев» . ПЛОС ОДИН . 6 (3): e14769. Бибкод : 2011PLoSO...614769F . дои : 10.1371/journal.pone.0014769 . ПМК 3060801 . ПМИД 21445243 .
- ^ Романдини, М.; Терлато, Г.; Наннини, Н. (2018). «Медведи и люди, неандертальская сказка. Реконструкция необычного поведения на основе зооархеологических данных в южной Европе». Журнал археологической науки . 90 : 71–91. Бибкод : 2018JArSc..90...71R . дои : 10.1016/j.jas.2017.12.004 . hdl : 11392/2381729 . S2CID 53410125 .
- ^ Финлейсон, Стюарт; Финлейсон, Клайв (9 ноября 2016 г.). «Люди-птицы плейстоцена: О взаимоотношениях неандертальцев и птиц-падальщиков» . Четвертичный интернационал . Человечество и птичий мир: зооархеологические данные, позволяющие сделать вывод о поведенческих эволюционных признаках. 421 : 78–84. Бибкод : 2016QuInt.421...78F . дои : 10.1016/j.quaint.2015.12.057 . ISSN 1040-6182 . Получено 25 февраля 2024 г. - через Elsevier Science Direct.
- ^ Колонезе, AC; Маннино, Массачусетс; Майер, DE (2011). «Эксплуатация морских моллюсков в предыстории Средиземноморья: обзор». Четвертичный интернационал . 239 (1–2): 86–103. Бибкод : 2011QuInt.239...86C . дои : 10.1016/j.quaint.2010.09.001 .
- ^ Перейти обратно: а б с Вилла, П.; Сориано, С.; Поллароло, Л. (2020). «Неандертальцы на пляже: использование морских ресурсов в Гроте деи Москерини (Лацио, Италия)» . ПЛОС ОДИН . 15 (1): e0226690. Бибкод : 2020PLoSO..1526690V . дои : 10.1371/journal.pone.0226690 . ПМК 6961883 . ПМИД 31940356 .
- ^ Кортес-Санчес, М.; Моралес-Муньис, А.; и др. (2011). «Самое раннее известное использование морских ресурсов неандертальцами» . ПЛОС ОДИН . 6 (9): e24026. Бибкод : 2011PLoSO...624026C . дои : 10.1371/journal.pone.0024026 . ПМЦ 3173367 . ПМИД 21935371 .
- ^ Стрингер, CB ; Финлейсон, К. ; Бартон, РНЕ (2008). «Неандертальская эксплуатация морских млекопитающих в Гибралтаре» . Труды Национальной академии наук . 105 (38): 14319–14324. Бибкод : 2008PNAS..10514319S . дои : 10.1073/pnas.0805474105 . ПМК 2567146 . ПМИД 18809913 .
- ^ Зильян, Ж.; Анджелуччи, Делавэр; Играя, Массачусетс; и др. (2020). «Последние межледниковые иберийские неандертальцы как рыбаки-охотники-собиратели» (PDF) . Наука . 367 (6485): eaaz7943. дои : 10.1126/science.aaz7943 . hdl : 2445/207289 . ПМИД 32217702 . S2CID 214671143 . Архивировано из оригинала (PDF) 8 ноября 2020 г. Проверено 4 октября 2020 г.
- ^ Перейти обратно: а б с д Харди, БЛ; Монсель, М.-Х.; Дожер, К.; и др. (2013). «Невозможные неандертальцы? Изготовление веревки, метание снарядов и ловля мелкой дичи на этапе 4 морского изотопа (Абри-дю-Марас, Франция)». Четвертичные научные обзоры . 82 : 23–40. Бибкод : 2013QSRv...82...23H . doi : 10.1016/j.quascirev.2013.09.028 .
- ^ Бохеренс, Х.; Барышников Г.; Ван Нир, В. (2014). «Принимали ли участие в накоплении лосося в среднем палеолите Кавказа медведи или львы? Изотопное исследование в пещере Кударо 3». Четвертичный интернационал . 339–340: 112–118. Бибкод : 2014QuInt.339..112B . дои : 10.1016/j.quaint.2013.06.026 .
- ^ Тринкаус, Э. (1975). «Сквоттинг среди неандертальцев: проблема поведенческой интерпретации морфологии скелета». Журнал археологической науки . 2 (4): 327–351. Бибкод : 1975JArSc...2..327T . дои : 10.1016/0305-4403(75)90005-9 .
- ^ Систиага, А.; и др. (2014). «Неандертальская еда: новый взгляд на фекальные биомаркеры» . ПЛОС ОДИН . 9 (6): е101045. Бибкод : 2014PLoSO...9j1045S . дои : 10.1371/journal.pone.0101045 . ПМК 4071062 . ПМИД 24963925 .
- ^ Даугард, К. (2008). «Освоение окружающей среды неандертальцами на юго-востоке Франции» . Бюллетень Французского доисторического общества (на французском языке). 106 (4): 818–819.
- ^ Перейти обратно: а б Спет, доктор юридических наук (2015). «Когда люди научились варить?» (PDF) . Палеоантропология : 54–67.
- ^ Агам, А.; Баркай, Р. (2018). «Охота на слонов и мамонтов во время палеолита: обзор соответствующих археологических, этнографических и этноисторических данных» . Четвертичный период . 1 (3): 19. дои : 10,3390/quat1010003 .
- ^ Перейти обратно: а б с Зильян, Ж.; Анджелуччи, Делавэр; Бадал-Гарсия, Э. (2010). «Символическое использование морских раковин и минеральных пигментов иберийскими неандертальцами» . Труды Национальной академии наук . 107 (3): 1023–1028. Бибкод : 2010PNAS..107.1023Z . дои : 10.1073/pnas.0914088107 . ПМЦ 2824307 . ПМИД 20080653 .
- ^ Дидрих, CG (2010). « Популяция Crocuta crocuta spelaea (Goldfuss 1823) и ее добыча из логова гиен в позднеплейстоценовой пещере Тойфельскаммер, помимо знаменитой палеолитической неандертальской пещеры (Северный Рейн-Вестфалия, северо-запад Германии)». Историческая биология . 23 (2–3): 237–270. дои : 10.1080/08912963.2010.530348 . S2CID 128758869 .
- ^ Ружье, Элен; Кревкер, Изабель (6 июля 2016 г.). «Неандертальский каннибализм и кости неандертальцев, используемые в качестве инструментов в Северной Европе» . Научные отчеты . 6 (1): 29005. Бибкод : 2016NatSR...629005R . дои : 10.1038/srep29005 . ISSN 2045-2322 . ПМЦ 4933918 . ПМИД 27381450 .
Наши результаты показывают, что неандертальцы из пещеры Труазием в Гойе были убиты, и гипотеза об их использовании в качестве источника пищи является наиболее скудным объяснением наблюдаемых модификаций поверхности костей. Гойе предоставляет первые недвусмысленные доказательства каннибализма неандертальцев в Северной Европе, и, учитывая даты, полученные на останках неандертальцев, наиболее вероятно, что они были обработаны их собратьями-неандертальцами, поскольку в то время в этом регионе не было современных людей. Однако имеющиеся данные не позволяют определить, представляют ли изменения, наблюдаемые на материале скелетов неандертальцев, символические практики или просто являются результатом обработки непосредственно доступных источников пищи.
- ^ Перейти обратно: а б с Ружье, Х.; Кревкер, И.; Боваль, К. (2016). «Неандертальский каннибализм и кости неандертальцев, используемые в качестве инструментов в Северной Европе» . Научные отчеты . 6 (29, 005): 29005. Бибкод : 2016NatSR...629005R . дои : 10.1038/srep29005 . ПМЦ 4933918 . ПМИД 27381450 .
- ^ Перейти обратно: а б с д Ираведра, Дж.; Юстос, М. (2015). «Каннибализм в неандертальском мире: исчерпывающий пересмотр» . Журнал тафономии . 13 (1): 33–52.
- ^ Дефлер, А.; Уайт, Т.; Валенси, П.; Слимак, Л. (1999). «Неандертальский каннибализм в Мула-Герси, Ардеш, Франция» . Наука . 286 (5437): 128–131. дои : 10.1126/science.286.5437.128 . ПМИД 10506562 .
- ^ Морей, Б.; Сандрин, К.; Боваль, К.; Манн, А.Е. (2017). «Проблемы идентификации частично переваренных человеческих зубов: первое описание останков неандертальцев из мустьерской стоянки Марильяк (Марийак-ле-Фран, Шаранта, Франция) и значение для палеоантропологических исследований» . Палео . 28 : 201–212. дои : 10.4000/палео.3448 .
- ^ Перейти обратно: а б Робрукс, В.; Сир, MJ; Нильсен, ТК; и др. (2012). «Использование красной охры ранними неандертальцами» . Труды Национальной академии наук Соединенных Штатов Америки . 109 (6): 1889–1894. Бибкод : 2012PNAS..109.1889R . дои : 10.1073/pnas.1112261109 . ПМК 3277516 . ПМИД 22308348 .
- ^ д'Эррико, Ф.; Ванхаерен, М.; Хеншилвуд, К. (2009). «От происхождения языка к разнообразию языков: что говорят археология и палеоантропология?». Становление красноречием: достижения в развитии языка, человеческого познания и современных культур . Издательство Джона Бенджамина. п. 25. ISBN 978-90-272-3269-4 .
- ^ Марин, К.; Монселе, М.-Х.; Ангелину, М.; Карчиумару, Р. (2002). «Пещера Чоарей-Бороштень (Карпаты, Румыния): находки среднего палеолита и технологический анализ каменных комплексов». Античность . 76 (293): 681–690. дои : 10.1017/S0003598X00091122 . S2CID 130918977 .
- ^ Пересани, М.; Ванхаерен, М.; Кваджотто, Э.; и др. (2013). «Охристая ископаемая морская раковина из мустьерского периода пещеры Фумане, Италия» . ПЛОС ОДИН . 8 (7): e68572. Бибкод : 2013PLoSO...868572P . дои : 10.1371/journal.pone.0068572 . ПМЦ 3707824 . ПМИД 23874677 .
- ^ Хайэм, Т.; Якоби, Р.; Жюльен, М.; Дэвид, Ф.; Бэселл, Л.; Вуд, Р.; Дэвис, В.; Рэмси, CB (2010). «Хронология Грота дю Ренн (Франция) и значение контекста украшений и человеческих останков в Шательперрониане» . Труды Национальной академии наук . 107 (47): 20234–20239. Бибкод : 2010PNAS..10720234H . дои : 10.1073/pnas.1007963107 . ПМЦ 2996711 . ПМИД 20956292 .
- ^ Мелларс, П. (2010). «Неандертальская символика и производство украшений: лопнувший пузырь?» . Труды Национальной академии наук Соединенных Штатов Америки . 107 (47): 20147–20148. Бибкод : 2010PNAS..10720147M . дои : 10.1073/pnas.1014588107 . ПМК 2996706 . ПМИД 21078972 .
- ^ Хаблин, Ж.-Ж.; Таламо, С.; Жюльен, М.; Дэвид, Ф.; Коннет, Н.; Боду, П.; Вандермерш, Б.; Ричардс, член парламента (2012). «Радиоуглеродные данные из Грот-дю-Ренн и Сен-Сезер подтверждают неандертальское происхождение шательперронцев» . Труды Национальной академии наук США . 109 (46): 18743–18748. Бибкод : 2012PNAS..10918743H . дои : 10.1073/pnas.1212924109 . ПМЦ 3503158 . ПМИД 23112183 .
- ^ Велкера, Ф.; Хайдиняк, М.; Таламо, С.; Джауэн, К.; Даннеманн, М.; Дэвид, Ф.; Жюльен, М.; Мейер, М.; Келсо, Дж.; Барнс, И.; Брейс, С.; Камминга, П.; Фишер, Р.; Кесслер, Б.М.; Стюарт-младший; Паабо, С. ; Коллинз, MJ; Хублин, Ж.-Ж. (2016). «Палеопротеомные данные идентифицируют архаичных гомининов, связанных с Шательперроном в Гроте дю Ренн» . Труды Национальной академии наук США . 113 (40): 11162–11167. Бибкод : 2016PNAS..11311162W . дои : 10.1073/pnas.1605834113 . ПМК 5056053 . ПМИД 27638212 .
- ^ Финлейсон 2019 , стр. 129–132.
- ^ Финлейсон, К. ; Браун, К.; Бласко, Р.; Роселл, Дж.; Блэк, Дж. Джозеф; Бортолотти, Греция; Финлейсон, Дж .; Санчес Марко, А.; Джайлс Пачеко, Ф.; Родригес Видаль, младший; Каррион, Дж.С.; Сказочный; Родригес Льянес, JM (2012). «Рыбак рыбака: эксплуатация неандертальцами хищников и врановых» . ПЛОС ОДИН . 7 (9): e45927. Бибкод : 2012PLoSO...745927F . дои : 10.1371/journal.pone.0045927 . ПМЦ 3444460 . ПМИД 23029321 .
- ^ Перейти обратно: а б Финлейсон, С.; Финлейсон, Дж .; Гузман, ФГ; Финлейсон, К. (2019). «Неандертальцы и культ солнечной птицы». Четвертичные научные обзоры . 217 : 217–224. Бибкод : 2019QSRv..217..217F . doi : 10.1016/j.quascirev.2019.04.010 . S2CID 149949579 .
- ^ Радовчич, Д.; Сршень, АО; Радовчич Ю.; Фрайер, Д.В.; Петралья, доктор медицины (2015). «Свидетельства неандертальских украшений: видоизмененные когти орлана-белохвоста в Крапине» . ПЛОС ОДИН . 10 (3): e0119802. Бибкод : 2015PLoSO..1019802R . дои : 10.1371/journal.pone.0119802 . ПМЦ 4356571 . ПМИД 25760648 .
- ^ Каллауэй, Э. (2015). «Неандертальцы носили орлиные когти в качестве украшений» . Природа . дои : 10.1038/nature.2015.17095 . S2CID 191368693 .
- ^ Родригес-Идальго, А.; Моралес, Дж.И.; Себриа, А.; и др. (2019). «Шательперронские неандертальцы из Кова Форадада (Калафель, Испания) использовали фаланги императорских орлов в символических целях» . Достижения науки . 5 (11): eaax1984. Бибкод : 2019SciA....5.1984R . дои : 10.1126/sciadv.aax1984 . ПМК 6824858 . ПМИД 31701003 .
- ^ Марке, Дж.; Лорбланше, М.; Оберлин, К.; Тамо-Бозсо, Э.; Обри, Т. (2016). «Новая датировка «маски» Ла Рош-Котар (Ланже, Эндр и Луара, Франция)» . Палео журнал доисторической археологии . 27 : 253–263.
- ^ Марке, Ж.-К.; Лорбланше, М. (2003). «Лицо неандертальца? Протофигурка из Ла-Рош-Котар, Ланже (Эндр и Луара, Франция)». Античность . 77 (298): 661–670. дои : 10.1017/s0003598x00061627 . ISSN 0003-598X . S2CID 162609308 .
- ^ Петтитт, ПБ (2003). «Это зарождение искусства? Или искусство младенца? Возможное лицо неандертальца из Ла-Рош-Котар, Франция» (PDF) . До земледелия . 11 (3). Архивировано из оригинала (PDF) 2 октября 2011 г.
- ^ «Но разве это искусство?». Наука . 302 (5652): 1890а–1890. 2003. doi : 10.1126/science.302.5652.1890a . S2CID 149146480 .
- ^ Маршак, А. (1988). «Неандертальцы и способность человека к символическому мышлению». Ин Отте, М. (ред.). L'Homme de Néandertal: la pensée [ Неандерталец: мысль ]. Том. 5. Льежский университет. п. 71.
- ^ Э. Каллауэй (2014). «Неандертальцы создали одни из древнейших произведений искусства Европы» . Новости природы . дои : 10.1038/nature.2014.15805 . S2CID 163840392 .
- ^ Родригес-Видаль, Дж.; д'Эррико, Ф.; Пачеко, ФГ (2014). «Наскальная гравюра, сделанная неандертальцами в Гибралтаре» . Труды Национальной академии наук . 111 (37): 13301–13306. Бибкод : 2014PNAS..11113301R . дои : 10.1073/pnas.1411529111 . ПМК 4169962 . ПМИД 25197076 .
- ^ Ледер, Д.; Германн, Р.; Хюльс, М.; и др. (2021). «Гравированная кость возрастом 51 000 лет демонстрирует способность неандертальцев к символическому поведению». Экология и эволюция природы . 5 (9): 1273–1282. Бибкод : 2021NatEE...5.1273L . дои : 10.1038/s41559-021-01487-z . ПМИД 34226702 . S2CID 235746596 .
- ^ Маррис, Э. (2018). «Неандертальские художники создали древнейшие из известных наскальных рисунков». Новости природы . дои : 10.1038/d41586-018-02357-8 .
- ^ Карсиумару, М.; Ницу, Э.-К.; Кирстина, О. (2015). «Жеода, расписанная охрой неандертальцем». Comptes Рендус Палевол . 14 (2): 31–41. Бибкод : 2015CRPal..14...31C . дои : 10.1016/j.crpv.2014.05.003 .
- ^ Добоси, В.Т. (1985). «Ювелирные изделия, музыкальные инструменты и экзотические предметы венгерского палеолита». Фолиа археологическая . 36 : 7–29.
- ^ Витальм, Г. (2004). «Новые свидетельства охоты на пещерного медведя из пещеры Поточка Зиялка, Словения» . Сообщения Комиссии по четвертичным исследованиям Австрийской академии наук . 13 :219-234.
- ^ Перейти обратно: а б Дидрих, CG (2015). « «Неандертальские костяные флейты»: просто продукты ледникового периода, когда пятнистые гиены добывали мусор на пещерных медвежатах в европейских пещерных медвежьих берлогах» . Королевское общество открытой науки . 2 (4): 140022. Цифровой код : 2015RSOS....240022D . дои : 10.1098/rsos.140022 . ПМЦ 4448875 . ПМИД 26064624 .
- ^ Морли, Иэн (2006). «Мустьерское музыкальное искусство? Случай с Divje Babe I Bone» . Оксфордский журнал археологии . 25 (4): 317–333. дои : 10.1111/j.1468-0092.2006.00264.x . Проверено 30 мая 2024 г.
- ^ Бродар, Митя (26 сентября 2008 г.). « Пищелка» из Дивих баб ни неандертальска» [Дивье Бабе «Флейта» не неандерталец] (на словенском языке). Архивировано из оригинала 28 июля 2011 года.
- ^ Рихтер, Д.; Грюн, Р.; Жоаннес-Бойо, Р.; Стил; и др. (2017). «Возраст окаменелостей гомининов из Джебель-Ирхуда, Марокко, и истоки среднего каменного века». Природа . 546 (7657): 293–296. Бибкод : 2017Natur.546..293R . дои : 10.1038/nature22335 . ПМИД 28593967 . S2CID 205255853 .
- ^ Ли, Ф.; Кун, СЛ; Чен, Ф. (2018). «Самый восточный средний палеолит (мустьерский период) из пещеры Цзиньситай, Северный Китай». Журнал эволюции человека . 114 : 76–84. Бибкод : 2018JHumE.114...76L . дои : 10.1016/j.jhevol.2017.10.004 . ПМИД 29447762 .
- ^ Перейти обратно: а б Роллан, Н.; Диббл, HL (1990). «Новый синтез изменчивости среднего палеолита». Американская древность . 55 (3): 480–499. дои : 10.2307/281279 . JSTOR 281279 . S2CID 161990108 .
- ^ Рокка, Р.; Коннет, Н.; Ломме, В. (2017). «До перехода? Последняя каменная индустрия среднего палеолита из Грота дю Ренн (слой XI) в Арси-сюр-Кюр (Бургундия, Франция)» . Comptes Рендус Палевол . 16 (8): 878–893. Бибкод : 2017CRPal..16..878R . дои : 10.1016/j.crpv.2017.04.003 .
- ^ Перейти обратно: а б Милкс, А.; Паркер, Д.; Поуп, М. (2019). «Внешняя баллистика плейстоценовых копий, брошенных вручную: экспериментальные данные и последствия для эволюции человека» . Научные отчеты . 9 (1): 820. Бибкод : 2019НатСР...9..820М . дои : 10.1038/s41598-018-37904-w . ПМК 6347593 . ПМИД 30683877 .
- ^ Перейти обратно: а б Февр, Ж.-П.; Морей, Б.; Бэйл, П.; Кревекёр, И. (2014). «Человеческие останки среднего плейстоцена из Турвиль-ла-Ривьер (Нормандия, Франция) и их археологический контекст» . ПЛОС ОДИН . 9 (10): е104111. Бибкод : 2014PLoSO...9j4111F . дои : 10.1371/journal.pone.0104111 . ПМЦ 4189787 . ПМИД 25295956 .
- ^ Боэда, Э.; Женесте, Дж. М.; Григго, К.; Мерсье, Н.; Мухесен, С.; Рейсс, Дж.Л.; Таха, А.; Валладас, Х. (2015). «Острие леваллуа, вставленное в позвонок дикого осла ( Equus africanus ): древко, снаряды и мустьерское охотничье оружие». Античность . 73 (280): 394–402. CiteSeerX 10.1.1.453.29 . дои : 10.1017/S0003598X00088335 . S2CID 163560577 .
- ^ Руссель, М.; Соресси, М.; Хублин, Ж.-Ж. (2016). «Загадка Шательперрона: каменные технологии клинков и лезвий из Кенсе, Франция». Журнал эволюции человека . 95 : 13–32. Бибкод : 2016JHumE..95...13R . дои : 10.1016/j.jhevol.2016.02.003 . ПМИД 27260172 .
- ^ Хаблин, Ж.-Ж.; Таламо, С.; Жюльен, М.; Дэвид, Ф.; Коннет, Н.; Боду, П.; Вандермерш, Б.; Ричардс, член парламента (2012). «Радиоуглеродные данные из Грот-дю-Ренн и Сен-Сезер подтверждают неандертальское происхождение шательперронцев» . Труды Национальной академии наук . 190 (46): 18743–18748. Бибкод : 2012PNAS..10918743H . дои : 10.1073/pnas.1212924109 . ПМЦ 3503158 . ПМИД 23112183 .
- ^ Перейти обратно: а б Соресси, М.; Макферрон, СП; Ленуар, М.; и др. (2013). «Неандертальцы изготовили первые в Европе специализированные костяные орудия» . Труды Национальной академии наук . 110 (35): 14186–14190. Бибкод : 2013PNAS..11014186S . дои : 10.1073/pnas.1302730110 . ПМЦ 3761603 . ПМИД 23940333 .
- ^ Кэрон, Ф.; д'Эррико, Ф.; Дель Мораль, П.; Сантос, Ф.; Зильяо, Дж. (2011). «Реальность символического поведения неандертальцев в Гроте дю Ренн, Арси-сюр-Кюр, Франция» . ПЛОС ОДИН . 6 (6): e21545. Бибкод : 2011PLoSO...621545C . дои : 10.1371/journal.pone.0021545 . ПМК 3126825 . ПМИД 21738702 .
- ^ Мелларс, П .; Гравина, Б.; Рэмси, CB (2007). «Подтверждение интерстратификации неандертальцев и современных людей на типовом участке Шательперрон» . Труды Национальной академии наук . 104 (9): 3657–3662. Бибкод : 2007PNAS..104.3657M . дои : 10.1073/pnas.0608053104 . ПМК 1805566 . ПМИД 17360698 .
- ^ Гравина, Б.; Бачеллери, Ф.; Ко, С. (2018). «Нет надежных доказательств связи неандертальцев и шательперронцев в Ла-Рош-а-Пьеро, Сен-Сезер» . Научные отчеты . 8 (1): 15134. Бибкод : 2018НатСР...815134Г . дои : 10.1038/s41598-018-33084-9 . ПМК 6181958 . ПМИД 30310091 .
- ^ Вилла, П.; Поллароло, Л.; Конфорти, Дж.; и др. (2018). «От неандертальцев до современных людей: новые данные об улузцах» . ПЛОС ОДИН . 13 (5): e0196786. Бибкод : 2018PLoSO..1396786V . дои : 10.1371/journal.pone.0196786 . ПМЦ 5942857 . ПМИД 29742147 .
- ^ Перейти обратно: а б Хоффекер, Дж. Ф. (2009). «Распространение современного человека в Европе» . Труды Национальной академии наук Соединенных Штатов Америки . 106 (38): 16040–16045. Бибкод : 2009PNAS..10616040H . дои : 10.1073/pnas.0903446106 . ПМЦ 2752585 . ПМИД 19571003 .
- ^ Мартисиус, Нидерланды; Велкер, Ф.; Доганджич, Т.; и др. (2020). «Неразрушающая идентификация ZooMS выявила стратегический выбор сырья для костных орудий неандертальцами» . Научные отчеты . 10 (1): 7746. Бибкод : 2020НатСР..10.7746М . doi : 10.1038/s41598-020-64358-w . ПМК 7210944 . ПМИД 32385291 .
- ^ Сайкс 2020 , стр. 260–261.
- ^ Спет, доктор медицинских наук; Чернов, Е. (2002). «Использование черепах среднего палеолита в пещере Кебара (Израиль)». Журнал археологической науки . 29 (5): 471–483. Бибкод : 2002JArSc..29..471S . дои : 10.1006/jasc.2001.0740 .
- ^ Арангурен, Б.; Реведин А.; Амико, Н.; Кавулли, Ф.; Джачи, Г.; Гримальди, С.; Маккиони, Н.; Сантаньелло, Ф. (2018). «Деревянные орудия и огневая техника на ранней неандертальской стоянке Поггетти-Векки (Италия)» . Труды Национальной академии наук . 115 (9): 2054–2059. Бибкод : 2018PNAS..115.2054A . дои : 10.1073/pnas.1716068115 . ПМЦ 5834685 . ПМИД 29432163 .
- ^ Ледер, Дирк; Леманн, Йенс; Милкс, Аннемике; Кодденберг, Тим; Зитц, Майкл; Фогель, Матиас; Бёнер, Утц; Тербергер, Томас (9 апреля 2024 г.). «Деревянные артефакты из Шёнингенского копьегоризонта и их место в эволюции человека» . Труды Национальной академии наук . 121 (15): e2320484121. Бибкод : 2024PNAS..12120484L . дои : 10.1073/pnas.2320484121 . ISSN 0027-8424 . ПМЦ 11009636 . ПМИД 38557183 .
- ^ Привет, Пи Джей; Анастасакис, К.; де Йонг, В.; ван Хозель, А.; Робрукс, В.; Соресси, М. (2016). «Выбор и использование диоксида марганца неандертальцами» . Научные отчеты . 6 (1): 22 159. Бибкод : 2016NatSR...622159H . дои : 10.1038/srep22159 . ПМЦ 4770591 . ПМИД 26922901 .
- ^ Жобер, Дж.; Верхейден, С.; Дженти, Д.; и др. (2016). «Ранние неандертальские постройки глубоко в пещере Брюникель на юго-западе Франции». Природа . 534 (7605): 111–114. Бибкод : 2016Natur.534..111J . дои : 10.1038/nature18291 . ПМИД 27251286 . S2CID 205249458 .
- ^ Дегано, И.; Сориано, С.; Вилла, П.; Поллароло, Л.; Лукейко, Джей Джей; Джейкобс, З.; Дука, К.; Витальяно, С.; Тоцци, К. (2019). «Хафтинг орудий среднего палеолита в Лациуме (центральная Италия): новые данные из пещер Фосселлоне и Сант-Агостино» . ПЛОС ОДИН . 14 (6): e0213473. Бибкод : 2019PLoSO..1413473D . дои : 10.1371/journal.pone.0213473 . ПМК 6586293 . ПМИД 31220106 .
- ^ Шмидт, П.; Кох, Т.; Блессинг, М.; Каракостис, Ф.А.; Харвати, К.; Дрезели В. и Шарри-Дюо А. (2023). «Метод производства березового дегтя Кенигсауэ документирует кумулятивную культуру неандертальцев» . Археологические и антропологические науки . 15 (6): 84. Бибкод : 2023АрАнС..15...84С . дои : 10.1007/s12520-023-01789-2 . ПМЦ 10202989 . ПМИД 37228449 .
- ^ Соренсен, Б. (2009). «Использование энергии Эмом Неандертальцами». Журнал археологической науки . 36 (10): 2201–2205. Бибкод : 2009JArSc..36.2201S . дои : 10.1016/j.jas.2009.06.003 . S2CID 44218631 .
- ^ Перейти обратно: а б Коллард, М.; Тарле, Л.; Сандгэт, Д.; Аллан, А. (2016). «Фаунистические свидетельства разницы в использовании одежды между неандертальцами и ранними современными людьми в Европе». Журнал антропологической археологии . 44 : 235–246. дои : 10.1016/j.jaa.2016.07.010 . hdl : 2164/9989 .
- ^ Перейти обратно: а б Уэльс, Н. (2012). «Моделирование одежды неандертальцев по этнографическим аналогам». Журнал эволюции человека . 63 (6): 781–795. Бибкод : 2012JHumE..63..781W . дои : 10.1016/j.jhevol.2012.08.006 . ПМИД 23084621 .
- ^ Перейти обратно: а б Картер, Т.; Контрерас, Д.А.; Холкомб, Дж.; Михайлович, Д.Д. (2019). «Самая ранняя оккупация Центрального Эгейского моря (Наксос), Греция: последствия для поведения и расселения гомининов и Homo sapiens » . Достижения науки . 5 (10): eaax0997. Бибкод : 2019SciA....5..997C . дои : 10.1126/sciadv.aax0997 . ПМК 6795523 . ПМИД 31663021 .
- ^ Йоханссон, С. (2015). «Языковые способности неандертальцев» . Ежегодный обзор лингвистики . 1 : 311–322. doi : 10.1146/annurev-linguist-030514-124945 .
- ^ Перейти обратно: а б Уайтинг, К.; Константакос, Л.; Сэдлер, Г.; Гилл, К. (2018). «Были ли неандертальцы разумны? Стоический подход» . Гуманитарные науки . 7 (2): 39. дои : 10.3390/h7020039 .
- ^ Краузе, Дж.; Лалуэса-Фокс, К.; Орландо, Л. (2007). «Производный вариант FOXP2 современных людей был использован неандертальцами». Современная биология . 17 (21): 1, 908–1, 912. Бибкод : 2007CBio...17.1908K . дои : 10.1016/j.cub.2007.10.008 . hdl : 11858/00-001M-0000-000F-FED3-1 . ПМИД 17949978 . S2CID 9518208 .
- ^ Моцци, А.; Форни, Д.; Клеричи, М.; Поццоли, У.; Маскеретти, С. (2016). «Эволюционная история генов, участвующих в устной и письменной речи: за пределами FOXP2» . Научные отчеты . 6 : 22157. Бибкод : 2016NatSR...622157M . дои : 10.1038/srep22157 . ПМЦ 4766443 . ПМИД 26912479 .
- ^ Мерфи, Э.; Бенитес-Буррако, А. (2017). «Палеоосцилломика: выявление аспектов языковых способностей неандертальцев на основе генной регуляции нейронных колебаний» (PDF) . Журнал антропологических наук . 96 (96): 111–124. дои : 10.4436/JASS.96010 . ПМИД 30566085 . S2CID 56479752 .
- ^ Либерман, П .; Крелин, ЕС (1971). «О речи неандертальца». Лингвистический запрос . 2 (2): 203–222. дои : 10.1515/9783110819434-007 . JSTOR 4177625 .
- ^ Либерман, П. (1992). «О речи неандертальцев и вымирании неандертальцев». Современная антропология . 33 (4): 409–410. дои : 10.1086/204092 . S2CID 144005974 .
- ^ Либерман, П. (2007). «Современные взгляды на речевые способности неандертальцев: ответ Бо и др. (2002)». Журнал фонетики . 35 (4): 552–563. дои : 10.1016/j.wocn.2005.07.002 .
- ^ Боэ, Л.-Ж.; Хайм, Ж.-Л.; Хонда, К.; Маэда, С. (2002). «Потенциальное пространство гласных неандертальцев было таким же большим, как и у современных людей». Журнал фонетики . 30 (3): 465–484. дои : 10.1006/jpho.2002.0170 . S2CID 16685699 .
- ^ Боэ, LJ; Хейм, Дж.Л.; Хонда, К.; Маэда, С.; Бадин, П.; Абри, К. (2007). «Голосовой тракт новорожденных людей и неандертальцев: акустические возможности и последствия для дискуссии о происхождении языка. Ответ Либерману». Журнал фонетики . 35 (4): 564–581. дои : 10.1016/j.wocn.2007.06.006 .
- ^ Либерман, П. (2015). «Язык не возник 100 000 лет назад» . ПЛОС Биология . 13 (2): e1002064. дои : 10.1371/journal.pbio.1002064 . ПМЦ 4332681 . ПМИД 25679377 .
- ^ Кисель, М.; Фуэнтес, А. (2018). « Поведенческая современность» как процесс, а не событие в человеческой нише» . Время и разум . 11 (2): 163–183. дои : 10.1080/1751696X.2018.1469230 . S2CID 165770309 .
- ^ Стерельный, К. (2011). «От гомининов к людям: как сапиенсы стали поведенчески современными» . Философские труды Королевского общества Б. 366 (1566): 809–822. дои : 10.1098/rstb.2010.0301 . ПМК 3048993 . ПМИД 21320896 .
- ^ Перейти обратно: а б с Вунн, И. (2001). «Поклонение пещерному медведю в палеолите» (PDF) . Блокноты из лаборатории Laxe Xeolóxico . 26 : 457–463.
- ^ Вышедский, А. (2017). «Эволюция языка к революции: от медленно развивающейся конечной системы общения с множеством слов к бесконечному современному языку» . Идеи и результаты исследования . 5 : е38546. bioRxiv 10.1101/166520 . дои : 10.3897/rio.5.e38546 .
- ^ Дефлер, Альбан (1993). Мустьерские погребения . Париж: CNRS. ISBN 978-2-222-04651-6 . Проверено 29 июля 2022 г.
- ^ Харрольд, Фрэнсис Б. (октябрь 1980 г.). «Сравнительный анализ палеолитических погребений Евразии» . Мировая археология . 12 (2): 195–211. дои : 10.1080/00438243.1980.9979792 . Проверено 29 июля 2022 г.
- ^ Отте, Марсель (1996). Нижний и средний палеолит в Европе . Париж: Арман Колен. ISBN 2-200-01389-2 . Проверено 29 июля 2022 г.
- ^ Смирнов, Юрий (июнь 1989 г.). «Намеренное захоронение людей: начало среднего палеолита (последнего оледенения)» . Журнал мировой предыстории . 3 (2): 199–233. дои : 10.1007/BF00975761 . S2CID 163801437 . Проверено 29 июля 2022 г.
- ^ Петтит, Пол (январь 2002 г.). «Мертвые неандертальцы: изучение изменчивости погребений в Среднепалеолитической Евразии». До земледелия . 2002 (1): 1–26. дои : 10.3828/BFARM.2002.1.4 . S2CID 52906473 .
- ^ Перейти обратно: а б Петтитт, Пол (2017). Оксфордский справочник по археологии . Нью-Йорк: Издательство Оксфордского университета. стр. 354–355. ISBN 978-0-19-885520-0 . Проверено 30 июля 2022 г.
- ^ Ренду, В.; Боваль, К.; Кревкер, И.; Бэйл, П.; Бальзо, А.; Висмут, Т.; Бургиньон, Л.; Дельфур, Г.; Февр, Ж.-П.; Лакрамп-Куяубер, Ф.; Мут, X.; Пасти, С.; Семаль, П.; Тавормина, К.; Тодиско, Д.; Турк, А.; Морей, Б. (2016). «Пусть мертвые говорят… комментарии к ответу Диббла и др. на «Доказательства, подтверждающие преднамеренное захоронение в Ла-Шапель-о-Сент» ». Журнал археологической науки . 69 : 12–20. Бибкод : 2016JArSc..69...12R . дои : 10.1016/j.jas.2016.02.006 .
- ^ Ренду, В.; Боваль, К.; Кревкер, И.; и др. (2014). «Доказательства преднамеренного захоронения неандертальцев в Ла-Шапель-о-Сент» . Труды Национальной академии наук . 111 (1): 81–86. Бибкод : 2014PNAS..111...81R . дои : 10.1073/pnas.1316780110 . ПМЦ 3890882 . ПМИД 24344286 .
- ^ Диббл, Х.; Алдеяс, В.; Гольдберг, П.; Сандгэт, Д.; Стил, TE (2015). «Критический взгляд на доказательства из Ла-Шапель-о-Сент, подтверждающие преднамеренное захоронение». Журнал археологической науки . 53 : 649–657. дои : 10.1016/j.jas.2014.04.019 .
- ^ Леруа-Гуран, А. (1975). «Цветы, найденные в Шанидаре IV, захоронении неандертальцев в Ираке». Наука . 190 (4214): 562–564. Бибкод : 1975Sci...190..562L . дои : 10.1126/science.190.4214.562 . S2CID 140686473 .
- ^ Солецкий, Р.С. (1975). «Шанидар IV: цветочное захоронение неандертальцев в северном Ираке». Наука . 190 (4217): 880–881. Бибкод : 1975Sci...190..880S . дои : 10.1126/science.190.4217.880 . S2CID 71625677 .
- ^ Соммер, Джей Ди (1999). «Цветочное захоронение Шанидара IV: переоценка погребального ритуала неандертальцев». Кембриджский археологический журнал . 9 (1): 127–129. дои : 10.1017/s0959774300015249 . S2CID 162496872 .
- ^ Кэрролл, член парламента (1986). «Культ медведя, которого не было: исследование психоистории антропологии». Журнал психоаналитической антропологии . 19 (1): 19–34.
- ^ Блан, AC (1962). «Некоторые свидетельства ранних идеологий человека» . В Уошберне, СЛ (ред.). Социальная жизнь древнего человека . Рутледж . стр. 124–126. ISBN 978-1-136-54361-6 .
- ^ Уайт, ТД; Тот, Н.; Чейз, ПГ (1991). «Вопрос ритуального каннибализма в Гроте Гваттари». Современная антропология . 32 (7): 118–138. дои : 10.1086/203931 . JSTOR 2743640 . S2CID 144628134 .
- ^ Романьоли, Rivals & Benazzi 2022 , стр. 267–268
- ^ Черчилль, Стивен Э.; Киз, Камрин; Росс, Энн Х. (август 2022 г.). «Морфология средней части лица и скрещивание неандертальца и современного человека» . Биология . 11 (8): 1163. doi : 10.3390/biology11081163 . ISSN 2079-7737 . ПМЦ 9404802 . ПМИД 36009790 .
- ^ Шанкарараман, С.; Паттерсон, Н.; Ли, Х.; Паабо, С. ; Райх, Д; Эйки, Дж. М. (2012). «Дата скрещивания неандертальцев и современных людей» . ПЛОС Генетика . 8 (10): e1002947. arXiv : 1208.2238 . Бибкод : 2012arXiv1208.2238S . дои : 10.1371/journal.pgen.1002947 . ПМЦ 3464203 . ПМИД 23055938 .
- ^ Перейти обратно: а б Ян, Массачусетс; Маласпинас, А.С.; Дюран, EY; Слаткин, М. (2012). «Древняя структура Африки вряд ли может объяснить генетическое сходство неандертальцев и неафриканских людей» . Молекулярная биология и эволюция . 29 (10): 2, 987–2, 995. doi : 10.1093/molbev/mss117 . ПМЦ 3457770 . ПМИД 22513287 .
- ^ Санчес-Кинто, Ф.; Ботиг, LR; Сивит, С.; Аренас, Дж.; Авила-Аркос, MC; Бустаманте, CD; Комас, Д.; Лалуэса-Фокс, К.; Карамелли, Д. (2012). «Население Северной Африки несет в себе признак смешения с неандертальцами» . ПЛОС ОДИН . 7 (10): е47765. Бибкод : 2012PLoSO...747765S . дои : 10.1371/journal.pone.0047765 . ПМЦ 3474783 . ПМИД 23082212 .
- ^ Йотова В.; Лефевр, Ж.-Ф.; Моро, К.; и др. (2011). «Х-сцепленный гаплотип неандертальского происхождения присутствует среди всех неафриканских популяций» . Молекулярная биология и эволюция . 28 (7): 1957–1962. дои : 10.1093/molbev/msr024 . ПМИД 21266489 .
- ^ Фу, К.; Ли, Х.; Мурджани, П.; и др. (2014). «Последовательность генома современного человека возрастом 45 000 лет из Западной Сибири» . Природа . 514 (7523): 445–449. Бибкод : 2014Natur.514..445F . дои : 10.1038/nature13810 . ПМЦ 4753769 . ПМИД 25341783 .
- ^ Перейти обратно: а б с Чен, Л.; Вольф, АБ; Фу, В.; Эйки, Дж. М. (2020). «Выявление и интерпретация очевидного неандертальского происхождения у африканских людей» . Клетка . 180 (4): 677–687.e16. дои : 10.1016/j.cell.2020.01.012 . ПМИД 32004458 . S2CID 210955842 .
- ^ Лозе, К.; Франц, LAF (2013). «Доказательства максимального правдоподобия примеси неандертальцев в евразийских популяциях на основе трех геномов». Популяции и эволюция . 1307 : 8263. arXiv : 1307.8263 . Бибкод : 2013arXiv1307.8263L .
- ^ Прюфер, К.; де Филиппо, К.; Гроте, С.; Мафессони, Ф.; Корлевич, П.; Хайдиняк, М.; и др. (2017). «Геном неандертальца с высоким охватом из пещеры Виндия в Хорватии» . Наука . 358 (6363): 655–658. Бибкод : 2017Sci...358..655P . дои : 10.1126/science.aao1887 . ПМК 6185897 . ПМИД 28982794 .
- ^ Паабо, С. (2015). «Разнообразное происхождение генофонда человека». Обзоры природы Генетика . 16 (6): 313–314. дои : 10.1038/nrg3954 . ПМИД 25982166 . S2CID 5628263 .
- ^ Энард, Д.; Петров, Д.А. (2018). «Доказательства того, что РНК-вирусы способствовали адаптивной интрогрессии между неандертальцами и современными людьми» . Клетка . 175 (2): 360–371. дои : 10.1016/j.cell.2018.08.034 . ПМК 6176737 . ПМИД 30290142 .
- ^ Перейти обратно: а б Фу, К.; Хайдиняк, М.; Молдавский, ОТ; и др. (2015). «Ранний современный человек из Румынии с недавним предком-неандертальцем» . Природа . 524 (7564): 216–219. Бибкод : 2015Natur.524..216F . дои : 10.1038/nature14558 . ПМЦ 4537386 . ПМИД 26098372 .
- ^ Таскент, РО; Алиоглу, Северная Дакота; Фер, Э.; и др. (2017). «Вариации и функциональное влияние происхождения неандертальцев в Западной Азии» . Геномная биология и эволюция . 9 (12): 3516–3624. дои : 10.1093/gbe/evx216 . ПМК 5751057 . ПМИД 29040546 .
- ^ Зорина-Лихтенвальтер, К.; Лихтенвальтер, Р.Н.; Зайкин Д.В.; и др. (2019). «Исследование в алом цвете: MC1R как главный предиктор рыжих волос и образец эффекта шлепанья» . Молекулярная генетика человека . 28 (12): 2093–2106. дои : 10.1093/hmg/ddz018 . ПМК 6548228 . ПМИД 30657907 .
- ^ Дин, К.; Ху, Ю.; Сюй, С.; Ван, К.-К.; Ли, Х.; Чжан, Р.; Ян, С.; Ван, Дж.; Джин, Л. (2014). «Неандертальское происхождение гаплотипов, несущих функциональный вариант Val92Met в MC1R у современных людей» . Молекулярная биология и эволюция . 31 (8): 1994–2003. дои : 10.1093/molbev/msu180 . ПМИД 24916031 . «Кроме того, мы обнаружили, что все предполагаемые интрогрессивные гаплотипы неандертальцев несут вариант Val92Met, вариант с потерей функции в MC1R, который связан с множеством дерматологических признаков, включая цвет кожи и фотостарение. Частота этой неандертальской интрогрессии у европейцев низкая (~ 5%), умеренный у аборигенов континентальной Восточной Азии (~30%) и высокий у аборигенов Тайваня (60–70%).
- ^ Перейти обратно: а б Мендес, Флорида; Уоткинс, Дж. К.; Хаммер, МФ (2013). «Неандертальское происхождение генетических вариаций в кластере генов иммунитета к ОАС» (PDF) . Молекулярная биология и эволюция . 30 (4): 798–801. дои : 10.1093/molbev/mst004 . ПМИД 23315957 . S2CID 2839679 . Архивировано из оригинала (PDF) 23 февраля 2019 г.
- ^ Перейти обратно: а б Мендес, Флорида; Уоткинс, Дж. К.; Хаммер, МФ (2012). «Гаплотип STAT2 интрогрессировал от неандертальцев и служит кандидатом положительного отбора в Папуа-Новой Гвинее» . Американский журнал генетики человека . 91 (2): 265–274. дои : 10.1016/j.ajhg.2012.06.015 . ПМЦ 3415544 . ПМИД 22883142 .
- ^ Перейти обратно: а б Даннеманн, М.; Андрес, А.А.; Келсо, Дж. (2016). «Интрогрессия неандертальских и денисовских гаплотипов способствует адаптивным изменениям в толл-подобных рецепторах человека» . Американский журнал генетики человека . 98 (1): 22–33. дои : 10.1016/j.ajhg.2015.11.015 . ПМЦ 4716682 . ПМИД 26748514 .
- ^ Грегори, доктор медицины; Киппенхан, Дж. С.; Айзенберг, ДП; и др. (2017). «Генетические вариации, происходящие от неандертальцев, формируют череп и мозг современного человека» . Научные отчеты . 7 (1): 6308. Бибкод : 2017НатСР...7.6308Г . дои : 10.1038/s41598-017-06587-0 . ПМЦ 5524936 . ПМИД 28740249 .
- ^ Чеви, Элизабет Т.; Уэрта-Санчес, Эмилия; Рамачандран, Сохини (14 августа 2023 г.). «Интеграция гендерных предубеждений в исследования архаической интрогрессии хромосомы X» . ПЛОС Генетика . 19 (8): e1010399. дои : 10.1371/journal.pgen.1010399 . ISSN 1553-7404 . ПМЦ 10449224 . ПМИД 37578977 .
Мы показали, что наблюдаемый низкий уровень архаического покрытия хромосомы X можно объяснить просто снижением влияния гетерозиса и половых предубеждений в событиях интрогрессии, без привлечения более сложной модели гибридной несовместимости. Наша работа также предполагает, что отрицательный отбор, вероятно, действовал на архаичные варианты, и предоставляет соответствующий набор нулевых моделей для оценки положительного отбора на интрогрессивных сегментах хромосомы X.
- ^ Луо, С.; Валенсия, Калифорния; Чжан, Дж.; Ли, Северная Каролина; Слон, Дж.; Гуй, Б.; Ван, X.; Ли, З.; Делл, С.; Браун, Дж.; Чен, С.М.; Чиен, Ю.-Х.; Хву, В.-Л.; Фан, П.-К.; Вонг, Л.-Дж.; Атвал, PS; Хуанг, Т. (2018). «Биродительское наследование митохондриальной ДНК у человека» . Труды Национальной академии наук . 115 (51): 13039–13044. Бибкод : 2018PNAS..11513039L . дои : 10.1073/pnas.1810946115 . ПМК 6304937 . ПМИД 30478036 .
- ^ Папини, М. (2020). Сравнительная психология: эволюция и развитие мозга и поведения, 3-е издание . Тейлор и Фрэнсис. п. 619. ИСБН 978-1-000-17770-1 . Проверено 28 марта 2024 г.
Есть свидетельства несколько избирательного скрещивания. мтДНК неандертальцев отсутствует у современных людей. Поскольку мтДНК переносится только самками, скрещивание могло происходить в основном между самцами-неандертальцами и современными самками (Krings, Stone, Schmitz, Krainitzki, Stoneking & Pääbo, 1997).
- ^ Перейти обратно: а б Паабо, С. (2014). Неандерталец: в поисках утраченных геномов . Основные книги. п. 19. ISBN 978-0-465-02083-6 . Проверено 28 марта 2024 г.
- ^ Мендес, Флорида; Позник, Г.Д.; Кастеллано, С.; Бустаманте, компакт-диск (2016). «Расхождение Y-хромосом неандертальца и современного человека» . Американский журнал генетики человека . 98 (4): 728–734. дои : 10.1016/j.ajhg.2016.02.023 . ПМЦ 4833433 . ПМИД 27058445 .
- ^ Шанкарараман, С.; Паттерсон, Н.; Ли, Х.; Паабо, С. ; Райх, Д. (2012). «Дата скрещивания неандертальцев и современных людей» . ПЛОС Генетика . 8 (10): e1002947. arXiv : 1208.2238 . Бибкод : 2012arXiv1208.2238S . дои : 10.1371/journal.pgen.1002947 . ПМЦ 3464203 . ПМИД 23055938 .
- ^ Гершковитц, И.; Вебер, Г.В.; Куам, Р.; Дюваль, М.; Грюн, Р. (2018). «Самые ранние современные люди за пределами Африки» . Наука . 359 (6374): 459. Бибкод : 2018Sci...359..456H . дои : 10.1126/science.aap8369 . hdl : 10072/372670 . ПМИД 29371468 .
- ^ Макдермотт, Ф.; Грюн, Р.; Стрингер, CB ; Хоксворт, CJ (1993). «Масс-спектрометрические даты U-серии для стоянок израильских неандертальцев и ранних современных гоминидов». Природа . 363 (6426): 252–255. Бибкод : 1993Natur.363..252M . дои : 10.1038/363252a0 . ПМИД 8387643 . S2CID 4234812 .
- ^ Пагани, Л. (2016). «Геномный анализ дает информацию о миграционных событиях во время заселения Евразии» . Природа . 538 (7624): 238–242. Бибкод : 2016Natur.538..238P . дои : 10.1038/nature19792 . ПМК 5164938 . ПМИД 27654910 .
- ^ Лоури, РК; Урибе, Г.; Хименес, Е.Б.; Вайс, Массачусетс; Эррера, К.Дж.; Регейру, М.; Эррера, Р.Дж. (2013). «Генетическое сходство неандертальцев и денисовых с современными людьми: интрогрессия против общих предковых полиморфизмов». Джин . 530 (1): 83–94. дои : 10.1016/j.gene.2013.06.005 . ПМИД 23872234 .
- ^ Стрингер, К. (2012). «Эволюция: что делает современного человека» . Природа . 485 (7396): 33–35. Бибкод : 2012Natur.485...33S . дои : 10.1038/485033а . ПМИД 22552077 . S2CID 4420496 .
- ^ Паабо, С. (2014). «Последовательность митохондриального генома гоминина из Сима-де-лос-Уэсос» (PDF) . Природа . 505 (7483): 403–406. Бибкод : 2014Natur.505..403M . дои : 10.1038/nature12788 . ПМИД 24305051 . S2CID 4456221 .
- ^ Уоррен, Мэтью (2018). «Мама — неандерталец, папа — денисовец: первое открытие гибрида древнего человека» . Новости природы . 560 (7719): 417–418. Бибкод : 2018Natur.560..417W . дои : 10.1038/d41586-018-06004-0 . ПМИД 30135540 .
- ^ Ортола, Поликарп; Мартинес-Наварро, Бьенвенидо (8 мая 2013 г.). «Четвертичное вымирание мегафауны и судьба неандертальцев: интегративная рабочая гипотеза» . Четвертичный интернационал . Восток встречается с Западом: первые поселения и эволюция человека в Евразии. 295 : 69–72. Бибкод : 2013QuInt.295...69H . дои : 10.1016/j.quaint.2012.02.037 . ISSN 1040-6182 . Получено 5 февраля 2024 г. - через Elsevier Science Direct.
- ^ Агусти, Дж.; Рубио-Кампильо, X. (2017). «Были ли неандертальцы ответственны за свое вымирание?». Четвертичный интернационал . 431 : 232–237. Бибкод : 2017QuInt.431..232A . дои : 10.1016/j.quaint.2016.02.017 .
- ^ Девьезе, Тибо; Абрамс, Грегори; Хайдиняк, Матея; Пирсон, Стефан; Де Гроот, Изабель; Ди Модика, Кевин; Туссен, Мишель; Фишер, Валентин; Комески, Дэн; Шпиндлер, Люк; Мейер, Матиас; Семаль, Патрик; Хайэм, Том (23 марта 2021 г.). «Переоценка времени исчезновения неандертальцев в Северо-Западной Европе» . Труды Национальной академии наук Соединенных Штатов Америки . 118 (12). Бибкод : 2021PNAS..11822466D . дои : 10.1073/pnas.2022466118 . ISSN 0027-8424 . ПМЦ 7999949 . ПМИД 33798098 .
- ^ Перейти обратно: а б Зильян, Ж.; Анесин Д.; Обри, Т.; и др. (2017). «Точная датировка перехода от среднего к верхнему палеолиту в Мурсии (Испания) подтверждает существование позднего неандертальца в Иберии» . Гелион . 3 (1): E00435. Бибкод : 2017Heliy...300435Z . doi : 10.1016/j.heliyon.2017.e00435 . ПМЦ 5696381 . ПМИД 29188235 .
- ^ Моралес, Дж. И.; Себриа, А.; Бурге-Кока, А.; и др. (2019). «Заселение переходного периода от среднего к верхнему палеолиту из Кова Форадада (Калафель, северо-восточная Иберия)» . ПЛОС ОДИН . 14 (5): e0215832. Бибкод : 2019PLoSO..1415832M . дои : 10.1371/journal.pone.0215832 . ПМК 6522054 . ПМИД 31095578 .
- ^ Кортес-Санчес, М.; Хименес-Эспехо, Ф.Дж.; Симон-Валлехо, доктор медицины; Стрингер, К. ; и др. (2019). «Раннее прибытие ориньякцев в юго-западную Европу». Экология и эволюция природы . 3 (2): 207–212. Бибкод : 2019NatEE...3..207C . дои : 10.1038/s41559-018-0753-6 . ПМИД 30664696 . S2CID 58571988 .
- ^ Андерсон, Л.; Рейнольдс, Н.; Тейсандье, Н. (2019). «Нет надежных свидетельств существования очень раннего ориньяка в Южной Иберии» . Экология и эволюция природы . 3 (5): 713. Бибкод : 2019NatEE...3..713A . дои : 10.1038/s41559-019-0885-3 . ПМИД 30988496 . S2CID 115153460 .
- ^ де ла Пенья, П. (2019). «Свидания сами по себе не могут определить модели занятий гоминидов» . Экология и эволюция природы . 3 (5): 712. Бибкод : 2019NatEE...3..712D . дои : 10.1038/s41559-019-0886-2 . ПМИД 30988497 . S2CID 116861216 .
- ^ Кортес-Санчес, М.; Хименес-Эспехо, Ф.Дж.; Симон-Валлехо, доктор медицины; Стрингер, К. ; и др. (2019). «Ответ на вопросы: «Датирование само по себе не может определить характер занятий гоминидов» и «Нет надежных доказательств существования очень раннего ориньякца в Южной Иберии» ». Экология и эволюция природы . 3 (5): 714–715. Бибкод : 2019NatEE...3..714C . дои : 10.1038/s41559-019-0887-1 . ПМИД 30988498 . S2CID 115153407 .
- ^ Фонтюнь, М.; Рейсс, Дж.Л.; Руис, CB; Лара, премьер-министр (1995). «Мустьерская стоянка Сафаррайя (Гранада, Испания): датировка и влияние на палеолитические процессы заселения Западной Европы». Comptes Rendus de l'Académie des Sciences . 321 (10): 931–937.
- ^ Финлейсон, К. ; Пачеко, ФГ (2006). «Позднее выживание неандертальцев на самой южной окраине Европы». Природа . 443 (7, 113): 850–853. Бибкод : 2006Natur.443..850F . дои : 10.1038/nature05195 . ПМИД 16971951 . S2CID 4411186 .
- ^ Павлов П.; Свендсен, Дж.И.; Индрелид, С. (2001). «Присутствие человека в Европейской Арктике почти 40 000 лет назад». Природа . 413 (6, 851): 64–67. Бибкод : 2001Natur.413...64P . дои : 10.1038/35092552 . ПМИД 11544525 . S2CID 1986562 .
- ^ Хаблин, Ж.-Ж.; Сираков Н.; и др. (2020). «Первоначальный человек верхнего палеолита Homo sapiens из пещеры Бачо Киро, Болгария» (PDF) . Природа . 581 (7808): 299–302. Бибкод : 2020Natur.581..299H . дои : 10.1038/s41586-020-2259-z . hdl : 11585/770553 . ПМИД 32433609 . S2CID 218592678 .
- ^ Бенацци, С.; Дука, К.; Форнаи, К.; Бауэр, CC; Куллмер, О.; Свобода, Ж.И.; Пап, И.; Маллени, Ф.; Бэйл, П.; Кокерель, М.; Кондеми, С.; Рончителли, А.; Харвати, К.; Вебер, GW (2011). «Раннее расселение современных людей в Европе и последствия для поведения неандертальцев». Природа . 479 (7374): 525–528. Бибкод : 2011Natur.479..525B . дои : 10.1038/nature10617 . ПМИД 22048311 . S2CID 205226924 .
- ^ Хайэм, Т.; Комптон, Т.; Стрингер, К.; Якоби, Р.; Шапиро, Б.; Тринкаус, Э.; Чендлер, Б.; Грёнинг, Ф.; Коллинз, К.; Хиллсон, С.; о'Хиггинс, П.; Фицджеральд, К.; Фэган, М. (2011). «Самые ранние свидетельства существования анатомически современных людей в северо-западной Европе». Природа . 479 (7374): 521–524. Бибкод : 2011Natur.479..521H . дои : 10.1038/nature10484 . ПМИД 22048314 . S2CID 4374023 .
- ^ Харвати, К.; и др. (2019). «Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии» . Природа . 571 (7766): 500–504. дои : 10.1038/s41586-019-1376-z . ПМИД 31292546 . S2CID 195873640 .
- ^ де Ламли, Массачусетс; Гиперт, Г.; де Ламли, Х .; Протопапа, Н.; Питсиос, Т. (2020). «Апидима 1 и Апидима 2: два черепа антенеандерта на Пелопоннесе, Греция». Л'Антропология . 124 (1): 102743. doi : 10.1016/j.anthro.2019.102743 . S2CID 216449347 .
- ^ Прюфер, Кей; Пост, Козимо (июнь 2021 г.). «Последовательность генома черепа современного человека возрастом более 45 000 лет из Златы-Куна в Чехии» . Экология и эволюция природы . 5 (6): 820–825. Бибкод : 2021NatEE...5..820P . дои : 10.1038/s41559-021-01443-x . ISSN 2397-334X . ПМЦ 8175239 . ПМИД 33828249 .
- ^ Штаубвассер, М.; Дрэгушин, В.; Онак, БП (2018). «Влияние изменения климата на переход неандертальцев к современным людям в Европе» . Труды Национальной академии наук Соединенных Штатов Америки . 115 (37): 9116–9121. Бибкод : 2018PNAS..115.9116S . дои : 10.1073/pnas.1808647115 . ПМК 6140518 . ПМИД 30150388 .
- ^ д'Эррико, Франческо; Санчес Гоньи, Мария Фернанда (апрель 2003 г.). «Вымирание неандертальцев и климатическая изменчивость OIS 3 в тысячелетнем масштабе» . Четвертичные научные обзоры . 22 (8–9): 769–788. Бибкод : 2003QSRv...22..769D . дои : 10.1016/S0277-3791(03)00009-X . Получено 14 марта 2024 г. - через Elsevier Science Direct.
- ^ Дегиоанни, А.; Боненфант, К.; Кабут, С.; Кондеми, С. (2019). «Жизнь на грани: была ли демографическая слабость причиной гибели неандертальцев?» . ПЛОС ОДИН . 14 (5). е0216742. Бибкод : 2019PLoSO..1416742D . дои : 10.1371/journal.pone.0216742 . ПМК 6541251 . ПМИД 31141515 .
- ^ Марти, А.; Фолч, А.; Коста, А.; Энгвелл, С. (2016). «Реконструкция плинианских и сопутствующих игнимбритов источников крупных извержений вулканов: новый подход к кампанскому игнимбриту» . Научные отчеты . 6 (21220): 1–11. Бибкод : 2016НатСР...621220М . дои : 10.1038/srep21220 . ПМЦ 4756320 . ПМИД 26883449 .
- ^ Либерски, П. (2013). «Куру: путешествие во времени от Папуа-Новой Гвинеи до вымирания неандертальцев» . Патогены . 2 (3): 472–505. doi : 10.3390/pathogens2030472 . ПМЦ 4235695 . ПМИД 25437203 .
Источники
[ редактировать ]- Браун, К.; Фа, Д.А.; Финлейсон, Дж .; Финлейсон, К. (2011). «Мелкая дичь и эксплуатация морских ресурсов неандертальцами: свидетельства из Гибралтара» . Поход по берегу: смена береговых линий и древность прибрежных поселений . Междисциплинарный вклад в археологию. Спрингер. ISBN 978-1-4419-8218-6 .
- Романьоли, Франческа; Соперники, Флоран; Бенацци, Стефано (2022). Обновление неандертальцев: понимание сложности поведения в позднем среднем палеолите . Академическая пресса. ISBN 978-0-12-821429-9 .
- Финлейсон, К. (2019). Умный неандерталец: ловля птиц, наскальное искусство и когнитивная революция . Издательство Оксфордского университета. ISBN 978-0-19-251812-5 .
- Френч, Дженнифер (2021). Палеолитическая Европа: демографическая и социальная предыстория . Кембридж, Великобритания: Издательство Кембриджского университета. ISBN 978-1-108-49206-5 .
- Папаянни, Д.; Морс, Массачусетс (2013). — Все еще с нами? . Неандертальцы открыли заново: как современная наука переписывает их историю . Темза и Гудзон. ISBN 978-0-500-77311-6 .
- Райх, Д. (2018). «Встречи с неандертальцами». Кто мы и как мы сюда попали: древняя ДНК и новая наука о человеческом прошлом . Издательство Оксфордского университета. ISBN 978-0-19-882125-0 .
- Шипман, П. (2015). «Как люди и их собаки привели неандертальцев к вымиранию». Захватчики: как люди и их собаки привели неандертальцев к вымиранию . Издательство Гарвардского университета. doi : 10.2307/j.ctvjf9zbs . ISBN 978-0-674-42538-5 . JSTOR j.ctvjf9zbs .
- Стрингер, К. ; Гэмбл, К. (1993). В поисках неандертальцев . Темза и Гудзон. ISBN 978-0-500-05070-5 .
- Сайкс, Ребекка Рэгг (2020). Сородичи: неандертальская жизнь, любовь, смерть и искусство . Лондон, Великобритания: Блумсбери Сигма. ISBN 978-1-4729-3749-0 .
- Таттерсолл, И. (2015). «Неандертальцы, ДНК и творчество». Странный случай с шатким казаком: и другие поучительные истории из эволюции человека . Издательская группа Святого Мартина. ISBN 978-1-4668-7943-0 .
Дальнейшее чтение
[ редактировать ]- Хант, Крис; и др. (28 августа 2023 г.). «Shanidar et ses fleurs? Размышления о палинологии неандертальской гипотезы «цветочных захоронений»» . Журнал археологической науки . 159 : 105822. Бибкод : 2023JArSc.159j5822H . дои : 10.1016/j.jas.2023.105822 . S2CID 261325698 .
Внешние ссылки
[ редактировать ]- Доказательство того, что неандертальцы ели крабов, — еще один «гвоздь в гроб» стереотипов о первобытных пещерных обитателях — Phys.org, 7 февраля 2023 г.
- « Человек неандертальский » . Смитсоновский институт. 14 февраля 2010 г.
- Хронология человечества (интерактивная) – Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).
- «ДНК неандертальца» . Международное общество генетической генеалогии. Архивировано из оригинала 17 июня 2006 г .: включает последовательности мтДНК неандертальцев.
- Записи GenBank для H.s. neanderthalensis, поддерживаемый Национальным центром биотехнологической информации (NCBI).
- Алекс, Бриджит (21 февраля 2024 г.). «Что стоит за эволюцией портретов неандертальцев» . САПИЕНС .