Хронология эволюции человека

Хронология эволюции человека описывает основные события в эволюционной линии современного человеческого вида , Homo sapiens . на протяжении всей истории жизни , начиная примерно 4 миллиарда лет назад, вплоть до недавней эволюции во H. sapiens время и после последнего ледникового периода .
Он включает краткие объяснения различных таксономических рангов человеческого происхождения. Временная шкала отражает основные взгляды в современной таксономии , основанные на принципе филогенетической номенклатуры ; в случае открытых вопросов и отсутствия четкого консенсуса кратко излагаются основные конкурирующие возможности.
Обзор таксономических рангов
[ редактировать ]Табличный обзор таксономического ранга Homo sapiens (с оценками возраста для каждого ранга) показан ниже.
| Классифицировать | Имя | Общее имя | Миллионы лет назад (начало) |
|---|---|---|---|
| Жизнь | 4,200 | ||
| Архея | 3,700 | ||
| Домен | Эукариоты | Эукариоты | 2,100 |
| Опимода | Исключает растения и их родственники. | 1,540 | |
| Аморфея | |||
| яичник | Исключает амебу (амёбы) | ||
| Опистоконт | Holozoa + Holomycota (Cristidicoidea и Fungi ) | 1,300 | |
| Голозоа | Исключает голомикоту | 1,100 | |
| Философия | Хоанозоа + Филастерия | ||
| Чоанозоа | Хоанофлагелляты + Животные | 900 | |
| Королевство | животное | Животные | 610 |
| Подкоролевство | Эуметазоа | Исключает Porifera (губки) | |
| Парахоксозоа | Исключает гребневики (гребневики) | ||
| Искатель | Триплобласты/черви | 560 | |
| Нефрозоа | |||
| вторичноротые | Отделение от протостомов | ||
| Тип | Хордовые | Хордовые (позвоночные и близкородственные беспозвоночные) | 530 |
| Ольфакторы | Исключает головохордовых (ланцетников). | ||
| Подтип | Позвоночные животные | Рыбы/Позвоночные животные | 505 |
| Инфрафилум | челюстноротые | Челюстная рыба | 460 |
| Телеостома | Костистая рыба | 420 | |
| Саркоптеригии | Рыба с лопастными плавниками | ||
| Суперкласс | Четвероногие | Четвероногие (животные с четырьмя конечностями) | 395 |
| Амниота | Амниоты (полностью наземные четвероногие, яйца которых «оснащены амнионом» ) | 340 | |
| Сторона синапса | Прото-млекопитающие | 308 | |
| Терапсид | Конечности под телом и другие черты млекопитающих | 280 | |
| Сорт | Млекопитающие | Млекопитающие | 220 |
| Подкласс | Терия | Млекопитающие, рождающие живых детенышей (т. е. не откладывающие яйца) | 160 |
| Инфракласс | Эутерия | Плацентарные млекопитающие (т.е. несумчатые) | 125 |
| Оптовые заказы | Бореоэтерия | Надприматы, (большинство) копытных млекопитающих, (большинство) плотоядных млекопитающих, китообразных и летучих мышей. | 124–101 |
| Суперзаказ | Эуархонтоглирес | Надприматные: приматы, колуго, землеройки, грызуны и кролики. | 100 |
| Грандордер | Эуархонта | Приматы , колуго и землеройки | 99–80 |
| Зеркальный заказ | Приматоморфа | Приматы и колуго | 79.6 |
| Заказ | Приматы | Приматы / Plesiadapiformes | 66 |
| Подотряд | Хаплоррини | «Сухоносые» (буквально «простоносые») приматы: долгопяты и обезьяны (в т.ч. обезьяны) | 63 |
| Инфрапорядок | Симииформы | обезьяны (в т.ч. обезьяны) | 40 |
| Парвордер | Катар | Приматы с «опущенным вниз носом»: человекообразные обезьяны и мартышки Старого Света | 30 |
| Суперсемья | Гоминодея | Обезьяны : человекообразные и низшие обезьяны ( гиббоны ). | 22-20 |
| Семья | Гоминиды | Человекообразные обезьяны : люди , шимпанзе , гориллы и орангутанги — гоминиды. | 20–15 |
| Подсемейство | Люди | Люди, шимпанзе и гориллы (африканские обезьяны) [ 1 ] | 14–12 |
| Племя | Люди | Включает как Homo , Pan (шимпанзе), но не Gorilla . | 10–8 |
| Подтриба | Люди | Род Homo и близкие родственники и предки человека после отделения от Пана — гоминины | 8–4 [ 2 ] |
| (Род) | Ардипитек с.л. | 6-4 | |
| (Род) | Австралопитек | 3 | |
| Род | Человек (H. habilis) | Люди | 2.5 |
| (Разновидность) | Х. прямоходящий сл | ||
| (Разновидность) | H. heidelbergensis sl | ||
| Разновидность | человек Мудрый | Анатомически современный человек | 0.8–0.3 [ 3 ] |
Хронология
[ редактировать ]−10 — – −9 — – −8 — – −7 — – −6 — – −5 — – −4 — – −3 — – −2 — – −1 — – 0 — | ( О. praegens ) ( О. тугененсис ) ( Ар. кадабба ) ( Ар. ramidus ) |
| ||||||||||||||||||||||||||||
−4500 — – — – −4000 — – — – −3500 — – — – −3000 — – — – −2500 — – — – −2000 — – — – −1500 — – — – −1000 — – — – −500 — – — – 0 — |
| |||||||||||||||||||||||||||||||||||||||||||||
Одноклеточная жизнь
[ редактировать ]| Дата | Событие |
|---|---|
| 4.3-4.1 Перейти |
Самая ранняя жизнь появляется, возможно, в виде протоклеток . Их генетический материал, вероятно, состоял из РНК , способной как к саморепликации, так и к ферментативной активности; их мембраны состояли из липидов . Гены представляли собой отдельные цепи, транслированные в белки и часто обменивавшиеся между протоклетками. |
| 4,0-3,8 млрд лет | Появляются прокариотические клетки ; их генетический материал состоит из более стабильной ДНК , и они используют белки по разным причинам, в первую очередь для того, чтобы помочь ДНК реплицироваться с помощью белковых ферментов (РНК сейчас действует как посредник в этой центральной догме потока генетической информации клеточной жизни); гены теперь связаны в последовательности, поэтому вся информация передается потомкам. У них были клеточные стенки и внешние мембраны , и они, вероятно, изначально были термофилами . |
| 3,5 млрд лет | Это знаменует собой первое появление цианобактерий и их метода кислородного фотосинтеза и, следовательно, первое появление атмосферного кислорода на Земле.
Еще миллиард лет прокариоты будут продолжать спокойно диверсифицироваться. |
| 2,5-2,2 млрд лет | Первые организмы, использующие кислород. К 2400 млн лет назад, во время так называемого Великого события окисления (GOE), большая часть докислородных анаэробных форм жизни была уничтожена производителями кислорода. |
| 2,2-1,8 млрд лет | Происхождение эукариот : организмы с ядрами , эндомембранными системами (в том числе митохондриями ) и сложным цитоскелетом ; они сплайсировали мРНК между транскрипцией и трансляцией (сплайсинг происходит и у прокариот, но только некодирующих РНК ). Считается, что эволюция эукариот и, возможно, пол связана с GOE, поскольку он, вероятно, заставил две или три линии прокариот (включая аэробную , которая позже стала митохондриями) зависеть друг от друга, что привело к эндосимбиозу . Ранние эукариоты потеряли клеточные стенки и внешние мембраны. |
| 1,2 млрд лет | развивается половое размножение ( митоз и мейоз ), что приводит к более быстрой эволюции. По крайней мере к этому времени [ 4 ] где гены смешиваются в каждом поколении, что обеспечивает большую вариативность для последующего отбора. |
| 1,2-0,8 млрд лет |  Линия эукариот Holozoa развивает множество особенностей создания клеточных колоний и, наконец, приводит к предку животных (многоклеточных животных) и хоанофлагеллят . [ 5 ] [ 6 ] Протероспонгии (представители хоанофлагеллат) — лучшие живые примеры того, как могли выглядеть предки всех животных. Они живут колониями и демонстрируют примитивный уровень клеточной специализации для выполнения различных задач. |
животное
[ редактировать ]| Дата | Событие |
|---|---|
| 800–650 млн лет назад |  Урметазой : Первые окаменелости, которые могут представлять животных, возрастом 665 миллионов лет появляются в породах формации Трезона в Южной Австралии . Эти окаменелости интерпретируются как ранние губки. [ 7 ] Многоклеточные животные могли существовать с 800 млн лет назад. Отделение от линии Porifera ( губок ). Eumetazoa / Diploblast : отделение от линии Ctenophora («гребневики»). Planulozoa / ParaHoxozoa : отделение от линий Placozoa и Cnidaria . Все диплобласты обладают эпителием , нервами , мышцами , соединительной тканью и ртом и, за исключением плакозой, имеют некоторую форму симметрии, причем их предки, вероятно, имели радиальную симметрию, как у книдарий. Диплобласты разделили свои ранние эмбриональные клетки на два зародышевых листка ( экто- и энтодерму ). Фоторецепторные глазные пятна развиваются. |
| 650-600 млн лет назад |  Урбилатерий : последний общий предок ксенацеломорфов, протостомов (включая членистоногих [насекомое, ракообразные, пауки], моллюсков [кальмар, улитка, моллюск] и кольчатых червей линии [дождевые черви]) и вторичноротых (включая линию позвоночных [человек]) (последний два более родственны друг другу и называются Nephrozoa ). У всех ксенацеломорфов есть гонопор для изгнания гамет , но у нефрозойных особей он слит с анусом . Самое раннее развитие двусторонней симметрии , мезодермы , головы (передняя цефализация ) и различных мышц кишечника (и, следовательно, перистальтики ), а у Nephrozoa - нефридиев (предшественников почек), целома (или, возможно, псевдоцелома ), отдельных рта и ануса (эволюция сквозных кишечник ) и, возможно, даже нервные связки и кровеносные сосуды . [ 8 ] Репродуктивная ткань, вероятно, концентрируется в паре гонад, соединяющихся непосредственно перед задним отверстием. Развиваются «чашеобразные глаза» и органы равновесия (функция слуха добавляется позже, по мере развития более сложного внутреннего уха у позвоночных). Нефрозойная сквозная кишка имела в передней части более широкую часть, называемую глоткой . Покровы ) или кожа состоят из эпителиального слоя ( эпидермиса и соединительного слоя. |
| 600-540 млн лет назад |  Большинство известных типов животных появились в летописи окаменелостей как морские виды во время эдиакарско - кембрийского взрыва , вероятно, вызванного крупномасштабной оксигенацией примерно с 585 млн лет назад (иногда называемой неопротерозойским событием оксигенации или NOE), а также притоком океанических минералов. Deuterostomes , последний общий предок линии Chordata [человека], Hemichordata ( желудевые черви и граптолиты ) и Echinodermata ( морские звезды , морские ежи , морские огурцы и т. д.), вероятно, имели как вентральные, так и дорсальные нервные шнуры , как современные желудевые черви. Архаичным выжившим на этой стадии является желудевый червь , имеющий открытую систему кровообращения (с менее разветвленными кровеносными сосудами) и сердце, которое также функционирует как почка. Желудевые черви имеют сплетения, сосредоточенные как в дорсальных, так и в вентральных нервных шнурах. Спинной канатик достигает хоботка и частично отделен от эпидермиса в этой области. Эта часть спинного нервного канатика часто полая и вполне может быть гомологична мозгу позвоночных. [ 9 ] У вторичноротых также появились глоточные щели , которые, вероятно, использовались для фильтрационного питания, как у полу- и протохордовых. |
Хордовые
[ редактировать ]| Дата | Событие |
|---|---|
| 540-520 млн лет назад |  Повышенное количество кислорода приводит к тому, что многие эукариоты, включая большинство животных, становятся облигатными аэробами . От предка хордовых произошли ланцетники (Amphioxii) и Olfactores. У предков хордовых развились постанальный хвост , хорда и эндостиль (предшественник щитовидной железы). Глоточные щели (или жабры ) теперь поддерживаются соединительной тканью и используются для фильтрационного питания и, возможно, дыхания . [ 10 ] Другие, более ранние предшественники хордовых, включают Myllokunmingia fengjiaoa , [ 11 ] Хайкоуэлла ланцетная , [ 12 ] и Haikouichthys ercaicunensis . [ 13 ] Вероятно, они потеряли свой вентральный нервный канатик и развили особую область спинного нерва , называемую мозгом , где глия стала постоянно ассоциироваться с нейронами . Вероятно, у них появились первые клетки крови (вероятно, ранние лейкоциты , что указывает на развитый врожденный иммунитет ), которые они образовали вокруг глотки и кишечника. [ 14 ] Все хордовые, за исключением оболочников, имеют сложную замкнутую систему кровообращения с сильно разветвленными кровеносными сосудами. Обонятельные , последний общий предок оболочников и позвоночных, у которых развилось обоняние (запах) . Поскольку у ланцетников нет сердца , возможно, оно появилось у этого предка (ранее сами кровеносные сосуды были сократительными), хотя у ланцетников оно могло быть утрачено после развития ранних вторичноротых (сердце есть у полухордовых и иглокожих). |
| 520-480 млн лет назад |  Появляются первые позвоночные (« рыбы »): остракодермы . Haikouichthys и Myllokunmingia являются примерами этих бесчелюстных рыб, или Agnatha ; бесчелюстные круглоротые на этом этапе расходятся. У них не было челюстей, они имели семь пар глоточных дуг , как и их сегодняшние потомки, а их эндоскелет состоял только из хондрокраниума был хрящевым (тогда он /мозговой оболочки и позвонков ). Соединительная ткань под эпидермисом дифференцируется на дерму и гиподерму . [ 15 ] Дыхание зависело от жабр , и у них развилось уникальное чувство вкуса (оставшееся ощущение кожи, теперь называемое « осязанием »), эндотелий , глаза -камеры и внутренние уши (способные слышать и балансировать; каждое из них состоит из лагены , отолитового органа). и два полукружных канала ), а также печень , щитовидная железа , почки и двухкамерное сердце (одно предсердие и один желудочек ). У них был хвостовой плавник , но не было парных (грудных и брюшных) плавников, как у более продвинутых рыб. Мозг разделен на три части (дальнейшее разделение привело к созданию отдельных областей в зависимости от функции). Шишковидная железа головного мозга проникает до уровня кожи головы, создавая впечатление третьего глаза . У них появились первые эритроциты и тромбоциты . [ 16 ] |
| 460-430 млн лет назад |  Плакодермы Gnathostomata были первыми челюстными рыбами ( ; ) их челюсти из первой жаберной /глоточной дуги, они в значительной степени заменили эндоскелетный хрящ костью развились и развили грудные и тазовые плавники. Кости первой жаберной дуги стали верхней и нижней челюстью, а кости второй дуги — гиомандибулой , цератохиальной и базихиальной; это закрыло две из семи пар жабр. Зазор между первой и второй дугами чуть ниже черепной коробки (сросшейся с верхней челюстью) образовал пару дыхалец , которые открывались в коже и вели в глотку (через них проходила вода и выходила через жабры). Плакодермы конкурировали с предыдущими доминирующими животными, головоногими моллюсками и морскими скорпионами , и сами стали доминировать. Некоторые из них, вероятно, развились в костных и хрящевых рыб после развития чешуи , зубов (что позволило перейти к полному хищничеству ), желудков , селезенки , тимуса , миелиновых оболочек , гемоглобина и развитого адаптивного иммунитета (последние два возникли независимо у миноги и миксины). У челюстных рыб также имеется третий, латеральный полукружный канал, а их отолиты разделены между собой мешочек и маточка . |
| 430-410 млн лет назад |  |
Четвероногие
[ редактировать ]| Дата | Событие |
|---|---|
| 390 млн лет назад |  У некоторых пресноводных лопастных рыб (sarcopterygii) развиваются конечности и дают начало Tetrapodomorpha . Эти рыбы эволюционировали в мелководных и болотистых пресноводных средах обитания , где у них появились большие глаза и дыхальца. Примитивные четвероногие («рыбаподы») произошли от четвероногих с двухдольным мозгом , уплощенным черепом, широкой пастью и средней мордой, чьи обращенные вверх глаза показывают, что это был донный обитатель, и у которых уже развились приспособления плавники с мясистым основанием и костями . («Живое ископаемое» целакант — это родственная рыба с лопастными плавниками, не имеющая подобных приспособлений для мелководья.) Четвероногие рыбы использовали свои плавники в качестве лопастей в мелководных местах обитания, забитых растениями и детритом . Универсальные характеристики четвероногих: передние конечности, сгибающиеся назад в локтях, и задние конечности, сгибающиеся вперед в коленях, вероятно, можно отнести к ранним четвероногим, обитавшим на мелководье. [ 18 ] Пандерихтис - рыба длиной 90–130 см (35–50 дюймов), обитающая в позднем девоне (380 млн лет назад ). У него большая голова, напоминающая четвероногих. Panderichthys демонстрирует переходные черты между лопастноперыми рыбами и ранними четвероногими. Отпечатки следов, оставленные чем-то, напоминающим конечности Ихтиостеги, образовались 390 млн лет назад в польских морских приливных отложениях. Это предполагает, что эволюция четвероногих старше, чем датированные окаменелости от Пандерихтиса до Ихтиостеги . |
| 375-350 млн лет назад |  Тиктаалик — род саркоптеригийных (лопастных) рыб позднего девона со многими чертами, напоминающими четвероногих. Это показывает четкую связь между Panderichthys и Acanthostega .   Акантостега — вымершее четвероногое животное , одно из первых животных, у которых были узнаваемые конечности. Это кандидат на звание одного из первых позвоночных, способных выходить на сушу. У него не было запястий, и в целом он был плохо приспособлен к жизни на суше. Конечности не могли выдержать вес животного. У Acanthostega были как легкие, так и жабры, что также указывает на то, что это было связующее звено между лопастными рыбами и наземными позвоночными. Спинная пара ребер образует грудную клетку, поддерживающую легкие, а вентральная пара исчезает. Ихтиостега — еще одно вымершее четвероногое животное. Будучи одним из первых животных, имевших всего две пары конечностей (также уникальных, поскольку они заканчиваются пальцами и имеют кости), Ихтиостега рассматривается как промежуточное звено между рыбой и амфибией. У Ихтиостеги были конечности, но они, вероятно, не использовались для ходьбы. Они, возможно, очень короткое время проводили вне воды и использовали свои конечности, чтобы пробираться сквозь грязь . [ 19 ] У них обоих было более пяти пальцев (восемь или семь) на концах каждой конечности, а их тела не имели чешуи (за исключением живота, где они остались в виде гастралий ). На этом этапе произошло множество эволюционных изменений: веки и слезные железы развились, чтобы предохранять глаза от попадания воды, а глаза стали соединяться с глоткой для отвода жидкости; гиомандибула (теперь называемая колумеллой ) сжалась в дыхальце, которое теперь также соединялось с внутренним ухом с одной стороны и с глоткой с другой, становясь евстахиевой трубой (колумелла помогала слышать); ранняя барабанная перепонка (участок соединительной ткани) развилась на конце каждой трубки (так называемая ушная вырезка ); а цератохиальная и базихиальная кости слились в подъязычную кость . Эти «рыбоногие» имели более окостеневшие и более сильные кости, позволяющие им удерживаться на суше (особенно кости черепа и конечностей ). Кости челюсти срастаются, а жаберные и крышечные кости исчезают. |
| 350-330 млн лет назад |  Педерпес, датируемый примерно 350 млн лет назад, указывает на то, что стандартное число из 5 цифр возникло в раннем карбоне , когда современные четвероногие (или « амфибии ») разделились на два направления (одно привело к современным амфибиям, а другое — к амниотам). На этом этапе у наших предков развились сошниково-носовые органы , слюнные железы , язык , паращитовидные железы , трехкамерное сердце (с двумя предсердиями и одним желудочком) и мочевые пузыри , а к зрелому возрасту они полностью удалили свои жабры. развивается Голосовая щель , чтобы предотвратить попадание пищи в дыхательные пути . Легкие и тонкая влажная кожа позволяли им дышать; вода также была необходима для рождения яиц без скорлупы и для раннего развития. Спинной, анальный и хвостовой плавники исчезли. Лиссамфибии (современные амфибии) сохраняют многие черты ранних амфибий, но имеют только четыре пальца ( у червяг нет). |
| 330-300 млн лет назад |  От земноводных произошли первые рептилии: Hylonomus — самая ранняя известная рептилия . Его длина составляла 20 см (8 дюймов) (включая хвост) и, вероятно, он был очень похож на современных ящериц . У него были маленькие острые зубы, и он, вероятно, питался мелкими многоножками и насекомыми . Это предшественник более поздних амниот (в самом широком смысле слова «рептилия»). альфа-кератин впервые развивается Здесь ; он используется в когтях современных амниот и в волосах млекопитающих, указывая на когти и другой тип чешуи, развившийся у амниот (также полная потеря жабр). [ 20 ] Эволюция амниотического яйца приводит к появлению амниот, четвероногих, которые могут размножаться на суше и откладывать яйца в скорлупе на суше. Им не нужно было возвращаться в воду для размножения или дыхания. Эта адаптация и устойчивая к высыханию чешуя дали им возможность впервые заселить возвышенности, хотя и заставляли их пить воду через рот. На этом этапе ткань надпочечников может сконцентрироваться в отдельных железах . У амниотов развитая нервная система с двенадцатью парами черепных нервов , в отличие от низших позвоночных. У них также развилась настоящая грудина , но они потеряли барабанные перепонки и ушные вырезки (слух только за счет костной проводимости колумеллы). |
Млекопитающие
[ редактировать ]| Дата | Событие |
|---|---|
| 300-260 млн лет назад | Вскоре после появления первых рептилий откололись две ветви. Одна ветвь — Sauropsida , от которой произошли современные рептилии и птицы. Другая ветвь — Synapsida, от которой произошли современные млекопитающие. У обоих были височные окна — пара отверстий в черепе за глазами, которые использовались для увеличения пространства для челюстных мышц. Синапсиды имели по одному отверстию с каждой стороны, а диапсиды (ветвь зауропсид) — два. Ранняя, неэффективная версия диафрагмы , возможно, развилась в синапсидах.
Самыми ранними «рептилиями, похожими на млекопитающих», являются пеликозавры . Пеликозавры были первыми животными, у которых были височные окна. Пеликозавры были не терапсидами , а их предками. Терапсиды, в свою очередь, были предками млекопитающих . У терапсидов височные окна были крупнее и больше напоминали млекопитающих, чем у пеликозавров, их зубы демонстрировали более серийную дифференциацию, их походка была полупрямой, а у более поздних форм развилось вторичное небо . Вторичное небо позволяет животному есть и дышать одновременно и является признаком более активного, возможно, теплокровного образа жизни. [ 21 ] Они лишились гастралий и, возможно, чешуи. |
| 260-230 млн лет назад |  Одна подгруппа терапсидов, цинодонты, потеряла шишковидное тело и поясничные ребра и, весьма вероятно, стала теплокровной . Нижние дыхательные пути образуют в паренхиме легких сложные ветви , заканчивающиеся сильно васкуляризированными альвеолами . Эритроциты и тромбоциты теряют свои ядра, в то время как лимфатическая система и высокий иммунитет возникают. Возможно, у них также была более толстая дерма , как у современных млекопитающих. Челюсти цинодонтов напоминали челюсти современных млекопитающих; передняя часть, зубная кость, содержала дифференцированные зубы. В эту группу животных, вероятно, входит вид, который является предком всех современных млекопитающих. Их височные окна слились с орбитами . Их задние конечности стали прямыми, а задние кости челюсти постепенно сжались до области колумеллы . [ 22 ] |
| 230-170 млн лет назад |  От Eucynodontia произошли первые млекопитающие . Большинство ранних млекопитающих были мелкими землероеподобными животными, питавшимися насекомыми и перешедшими к ночному образу жизни , чтобы избежать конкуренции с доминирующими архозаврами — это привело к утрате зрения красного и ультрафиолетового света (наследственная тетрахромация позвоночных редуцировалась до дихромазии ). Хотя в летописи окаменелостей нет никаких свидетельств, вполне вероятно, что эти животные имели постоянную температуру тела , волосы и молочные железы у своих детенышей (железы произошли от молочной линии ). Неокортекс . (часть головного мозга) развивается у млекопитающих при редукции тектума ( необонятельные ощущения, которые здесь обрабатывались, интегрировались в неокортекс, но обоняние стало основным чувством) Начало предстательной железы и пара отверстий, открывающихся в колумеллу и близлежащие сморщивающиеся кости челюсти; новые барабанные перепонки располагаются впереди колумеллы и евстахиевой трубы. Кожа становится волосистой, железистой (железы, выделяющие кожное сало и пот ) и терморегуляторной. Зубы полностью дифференцируются в резцы , клыки , премоляры и коренные зубы ; млекопитающие становятся дифиодонтными и обладают развитой диафрагмой, а у самцов — внутренними пенисами . Все млекопитающие имеют четырехкамерное сердце (с двумя предсердиями и двумя желудочками) и лишены шейных ребер (сейчас у млекопитающих есть только грудные ребра). Однопроходные — яйцекладущая группа млекопитающих, представленная сегодня утконосом и ехидной . Недавнее секвенирование генома утконоса показывает, что его половые гены ближе к генам птиц, чем к генам терианских (живорожденных) млекопитающих. Сравнивая это с другими млекопитающими, можно сделать вывод, что первые млекопитающие, получившие половую дифференциацию благодаря наличию или отсутствию гена SRY (обнаруженного в y-хромосоме), эволюционировали только у териан. Ранние млекопитающие и, возможно, их предки-эуцинодонты имели надлобковые кости , которые у современных сумчатых (обоих полов) служат для удержания сумки. |
| 170-120 млн лет назад |  Эволюция живорождения ( живорождение ), при этом ранние терианы, вероятно, имели мешочки для хранения неразвитого детеныша, как у современных сумчатых . Соски торчали из терианских молочных линий. разделяется Заднее отверстие на анальное и мочеполовое отверстия; самцы обладают внешним пенисом. Однопроходные и терии независимо отделяют молоточек и наковальню от зубной кости (нижней челюсти) и соединяют их с сморщенной колумеллой (теперь называемой стременем ) в барабанной полости позади барабанной перепонки (которая соединена с молоточком и удерживается другой костью, отделенной от барабанной перепонки). зубная кость, барабанная и эктотимпанальная части ) и скручивают свою лагену ( улитку ), чтобы улучшить слух, с У терианцев в дальнейшем развиваются внешние ушные раковины и прямые передние конечности. У самок плацентарных млекопитающих нет мешочков и надлобковых костей, но вместо этого имеется развитая плацента , которая проникает в стенки матки (в отличие от сумчатых), что обеспечивает более длительную беременность ; у них также есть отдельные мочевые и половые отверстия. [ 23 ] |
| 100-90 млн лет назад | Последний общий предок грызунов хищников , кроликов , копытных , и человека , летучих мышей , землероек (основа клады Boreoeutheria ; у самцов теперь есть наружные яички ). |
Приматы
[ редактировать ]| Дата | Событие |
|---|---|
| 90–66 млн лет назад |   Группа мелких, ночных, древесных и насекомоядных млекопитающих под названием Euarchonta начинает видообразование, которое приведет к приматов , отрядам землероек и летающих лемуров . Они сократили количество молочных желез до двух пар (на груди). Primatomorpha — подразделение Euarchonta, включающее приматов и их предков-стволовых приматов Plesiadapiformes . Ранний стеблевой примат Plesiadapis еще имел когти и глаза по бокам головы, что позволяло передвигаться по земле быстрее, чем по деревьям, но он начал проводить длительное время на нижних ветвях, питаясь плодами и листьями. Plesiadapiformes, скорее всего, содержат виды-предки всех приматов. [ 24 ] Впервые они появились в летописи окаменелостей около 66 миллионов лет назад, вскоре после мел-палеогенового вымирания , которое уничтожило около трех четвертей видов растений и животных на Земле, включая большинство динозавров. [ 25 ] [ 26 ] Один из последних Plesiadapiformes — Carpolestes simpsoni , имеющий хватательные пальцы, но не обращенные вперед глаза. |
| 66-56 млн лет назад | Приматы делятся на подотряды Strepsirrini (приматы с мокрым носом) и Haplorrini (приматы с сухим носом). Мозг расширяется и большой мозг делится на 4 пары долей . Заглазничная перекладина развивается и отделяет орбиту от височных ямок , когда зрение вновь обретает свое положение в качестве основного чувства; глаза стали обращены вперед. Strepsirrini содержат большинство полуобезьян ; современные примеры включают лемуров и лори . К гаплорринам относятся две современные группы: прозимийские долгопяты и обезьянообразные обезьяны , включая человекообразных обезьян . Метаболизм хаплоррини утратил способность вырабатывать витамин С , что вынудило всех потомков включать в свой рацион фрукты, содержащие витамин С. У ранних приматов когти были только на вторых пальцах; остальные были превращены в гвозди . |
| 50-35 млн лет назад |  Обезьяны разделились на инфраотряды Платиррини и Катаррини . Они полностью перешли к дневному образу жизни , у них отсутствовали клешни и тапетум lucidum (которые неоднократно эволюционировали у различных позвоночных). Возможно, у них развились, по крайней мере, некоторые из околоносовых пазух , и они перешли от эстрального цикла к менструальному циклу . Количество молочных желез теперь сократилось до одной грудной пары. У Platyrhines, обезьян Нового Света, цепкие хвосты, а самцы не различают цветов. Предполагается, что особи, потомки которых станут Platyrrini, мигрировали в Южную Америку либо на плоту растительности , либо по сухопутному мосту (теперь эта гипотеза отдавала предпочтение [ 27 ] ). Катарины в основном остались в Африке , когда два континента разошлись. Возможными ранними предками катарнов являются Aegyptopithecus и Saadanius . |
| 35-20 млн лет назад |  Catarrini разделяется на 2 суперсемейства: обезьян Старого Света (Cercopithecoidea) и человекообразных обезьян ( Hominoidea ). цветовое зрение человека Трехцветное зародилось в этот период на генетическом уровне. Catarrhines потеряли сошниково-носовой орган (или, возможно, превратили его в рудиментарный статус). Проконсул — ранний род катариновых приматов. У них была смесь черт Старого Света и обезьян . Проконсула Обезьяноподобные черты включают тонкую зубную эмаль, легкое телосложение с узкой грудью и короткими передними конечностями, а также древесный четвероногий образ жизни. Его обезьяноподобные черты — отсутствие хвоста, обезьяньи локти и немного больший мозг по сравнению с размером тела. Proconsul africanus — возможный предок как высших, так и малых обезьян, включая человека. |
Гоминиды
[ редактировать ]| Дата | Событие |
|---|---|
| 20-15 млн лет назад | Гоминиды (предки человекообразных обезьян) произошли от предков гиббонов ( меньших обезьян) между ок. от 20 до 16 млн лет назад. Они в значительной степени уменьшили свою наследственную морду и потеряли фермент уриказу (присутствующий у большинства организмов). [ 28 ] |
| 16-12 млн лет назад | Предки гоминин произошли от предков орангутанов между ок. 18–14 млн лет назад. [ 29 ]
и Считается, что Pierolapithecus catalaunicus является других человекообразных общим предком человека обезьян или, по крайней мере, видом, который приближает нас к общему предку, чем любое предыдущее открытие окаменелостей. У него были особые приспособления для лазания по деревьям, как у современных людей и других человекообразных обезьян: широкая плоская грудная клетка , жесткая нижняя часть позвоночника , гибкие запястья и лопатки , лежащие вдоль спины. |
| 12 млн лет назад | Danuvius guggenmosi — первая обнаруженная человекообразная обезьяна позднего миоцена с сохранившимися длинными костями , которая в значительной степени объясняет анатомическое строение и передвижение современных обезьян. [ 30 ] У него были приспособления как к зависанию на деревьях ( подвешивание ), так и к хождению на двух ногах ( двуногие ноги ), тогда как среди современных гоминидов люди лучше приспособлены к последнему, а другие — к первому. Таким образом, у Данувиуса был метод передвижения, в отличие от любой ранее известной обезьяны, называемый «лазанием на расширенных конечностях», он ходил прямо по ветвям деревьев, а также использовал руки для подвешивания. Последний общий предок человека и других обезьян, возможно, имел схожий метод передвижения. |
| 12-8 млн лет назад | Клада, в настоящее время представленная людьми и родом Pan ( шимпанзе и бонобо ), отделилась от предков горилл между ок. от 12 до 8 млн лет. [ 31 ] |
| 8-6 млн лет назад |  Гоминини : последний общий предок человека . и шимпанзе По оценкам, они жили примерно от 10 до 5 миллионов лет назад. И у шимпанзе, и у людей есть гортань , которая в течение первых двух лет жизни перемещается в место между глоткой и легкими, что указывает на то, что у общих предков была эта особенность, которая является предпосылкой для вокализованной речи у людей. Видообразование могло начаться вскоре после 10 млн лет назад, но позднее смешение между линиями могло произойти и после 5 млн лет назад. Кандидаты в виды Homininae или Homininae , жившие в этот период времени, включают Уранопитек (ок. 8 млн лет назад), Грекопитек (ок. 7 млн лет назад), Sahelanthropus tchadensis (ок. 7 млн лет назад), Оррорин тугененсис (ок. 6 млн лет назад). 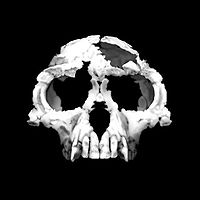 Ардипитек вел древесный образ жизни, то есть жил в основном в лесу, где конкурировал за пищу с другими лесными животными, включая, без сомнения, современных предков шимпанзе. Ардипитек, вероятно, был двуногим , о чем свидетельствуют его чашеобразный таз, угол большого затылочного отверстия и более тонкие кости запястья, хотя его ступни все еще были приспособлены для хватания, а не для ходьбы на большие расстояния. |
| 4-3,5 млн лет назад |  Представитель Australopithecus afarensis оставил следы, похожие на человеческие, на вулканическом пепле в Лаэтоли , северная Танзания , что является убедительным доказательством постоянного прямохождения. Australopithecus afarensis жил между 3,9 и 2,9 миллионами лет назад и считается одним из самых ранних гомининов — тех видов, которые развились и составили линию Homo и Homo ближайших родственников после отделения от линии шимпанзе. Считается, что A. afarensis был предком как рода Australopithecus , так и рода Homo . По сравнению с современными и вымершими человекообразными обезьянами , у A. afarensis были уменьшены клыки и коренные зубы, хотя они все же были относительно крупнее, чем у современных людей. A. afarensis также имеет относительно небольшой размер мозга (380–430 см). 3 ) и прогнатическое (выступающее вперед) лицо. Австралопитеки были обнаружены в саванне; они, вероятно, разработали свой рацион, включив в него очищенное мясо. Анализ африканского австралопитека нижних позвонков позволяет предположить, что эти кости изменились у женщин, чтобы поддерживать прямохождение даже во время беременности. |
| 3,5–3,0 млн лет назад | Kenyanthropus platyops , возможный предок Homo , произошел от австралопитека . Каменные орудия изготавливаются намеренно. [ 34 ] |
| 3 млн лет | Двуногие австралопитеки (род подтрибы Hominina ) эволюционируют в саваннах Африки, на которых охотится Мегантереон . Потеря волос на теле происходит от 3 до 2 млн лет назад, параллельно с развитием полного прямохождения и небольшим увеличением мозга. [ 35 ] |
гомо
[ редактировать ]| Дата | Событие |
|---|---|
| 2,5–2,0 млн лет назад |
Ранний Homo появляется в Восточной Африке и происходит от австралопитеков предков . Нижний палеолит определяется началом использования каменных орудий . Австралопитек гархи использовал каменные орудия примерно 2,5 млн лет назад. Homo habilis дал обозначение Homo — старейший вид , которому Лики и др. . в 1964 году. H. habilis занимает промежуточное положение между Australopithecus afarensis и H. erectus , и были предложения переклассифицировать его в пределах рода Australopithecus , как Australopithecus habilis . Каменные орудия, найденные на стоянке Шанчен в Китае и датированные 2,12 миллиона лет назад, считаются самыми ранними известными свидетельствами существования гоминидов за пределами Африки, превосходящими Дманиси в Грузии на 300 000 лет. [ 36 ] |
| 1,9–0,8 млн лет назад |  Homo erectus произошел от раннего Homo или позднего австралопитека . Homo habilis , хотя и значительно отличается по анатомии и физиологии, считается предком Homo ergaster , или африканского Homo erectus ; но известно также, что он сосуществовал с H. erectus почти полмиллиона лет (примерно до 1,5 млн лет назад). С момента своего самого раннего появления (около 1,9 млн лет назад) H. erectus распространен в Восточной Африке и Юго-Западной Азии ( Homo georgicus ). H. erectus — первый известный вид, который примерно к 1,5 млн лет назад научился контролировать огонь . Позже H. erectus мигрирует по Евразии , достигнув Юго-Восточной Азии к 0,7 млн лет назад. Описан в ряде подвидов . [ 37 ] Древние люди были социальными и сначала собирали мусор, а затем стали активными охотниками. Необходимость эффективно общаться и охотиться на добычу в новой, меняющейся среде (где необходимо запоминать и сообщать расположение ресурсов), возможно, привела к расширению мозга с 2 до 0,8 млн лет назад. Эволюция темной кожи около 1,2 млн лет назад. [ 38 ] Homo antecessor может быть общим предком человека и неандертальцев. [ 39 ] [ 40 ] По нынешним оценкам, люди имеют примерно 20 000–25 000 генов и на 99% имеют общую ДНК с ныне вымершими неандертальцами. [ 41 ] и 95–99% их ДНК принадлежит их ближайшему ныне живущему эволюционному родственнику, шимпанзе . [ 42 ] [ 43 ] Человеческий вариант гена FOXP2 (связанный с контролем речи) оказался идентичным у неандертальцев. [ 44 ] |
| 0,8–0,3 млн лет назад |  Отличие неандертальцев и денисовцев от общего предка. [ 45 ] Homo heidelbergensis (в Африке также известный как Homo rhodesiensis ) долгое время считался вероятным кандидатом на роль последнего общего предка неандертальцев и современных людей. Однако генетические данные окаменелостей Сима-де-лос-Уэсос , опубликованные в 2016 году, по-видимому, позволяют предположить, что H. heidelbergensis в целом следует включить в линию неандертальцев как «пренеандертальцев» или «ранних неандертальцев», в то время как время расхождения между Неандертальские и современные линии были отодвинуты до появления H. heidelbergensis , примерно до 600 000 800 000 лет назад — примерный возраст предшественника Homo . [ 46 ] [ 47 ] Расширение (увеличение) мозга между 0,8 и 0,2 млн лет назад могло произойти из-за вымирания большей части африканской мегафауны (из-за чего люди питались более мелкой добычей и растениями, что требовало большего интеллекта из-за большей скорости первых и неуверенности в том, были ли последние ядовиты они или нет), чрезвычайная изменчивость климата после переходного периода среднего плейстоцена (что усугубило ситуацию и привело к частым миграциям), а также общий отбор на более активную социальную жизнь (и интеллект) для большей вероятности выживание, репродуктивность и забота о матерях. Затвердевшие следы, датированные примерно 350 тыс. лет назад и связанные с H. heidelbergensis, были обнаружены на юге Италии в 2003 году. [ 48 ] H. sapiens полностью утратил надбровные дуги от своих предков-гоминидов, а также морду, хотя их носы стали выступающими (возможно, со времен H. erectus ). К 200 тыс. лет назад люди прекратили расширение своего мозга. |
Мудрый человек
[ редактировать ]| Дата | Событие |
|---|---|
| 300–130 тыс. лет назад |  Неандертальцы и денисовцы происходят из северной линии Homo heidelbergensis около 500–450 тыс. лет назад, тогда как разумные люди происходят из южной линии около 350–300 тыс. лет назад. [ 49 ] Окаменелости, приписываемые H. sapiens, вместе с каменными орудиями, датируемыми примерно 300 000 лет назад, найдены в Джебель-Ирхуде , Марокко. [ 50 ] предоставить самые ранние ископаемые свидетельства анатомически современного Homo sapiens . Современное присутствие человека в Восточной Африке ( Гадемотта ), 276 тыс. лет назад. [ 51 ] В июле 2019 года антропологи сообщили об обнаружении останков H. sapiens возрастом 210 000 лет в пещере Апидима , Пелопоннес , Греция . [ 52 ] [ 53 ] [ 54 ] по отцовской и матрилинейной линии примерно между 200 и 100 тыс. лет назад. Самые последние общие предки (MRCA) живых людей [ 55 ] [ 56 ] по некоторым оценкам, MRCA по отцовской линии несколько выше - от 250 до 500 тысяч лет назад. [ 57 ] 160 000 лет назад Homo sapiens idaltu в долине реки Аваш (недалеко от современной деревни Херто , Эфиопия) практиковал экскарнацию . [ 58 ] |
| 130–80 тыс. лет назад | Морской изотоп 5-й стадии ( Эемиан ).
Современное присутствие человека в Южной Африке и Западной Африке . [ 59 ] Появление митохондриальной гаплогруппы (мт-гаплогруппы) L2 . |
| 80–50 раз | МИС 4 , начало верхнего палеолита .
Ранние свидетельства поведенческой современности . [ 60 ] Появление мт- М и N. гаплогрупп Южное расселение, миграция из Африки , протоавстралоидное заселение Океании . [ 61 ] Архаичная примесь неандертальцев Евразии , [ 62 ] [ 63 ] от денисовцев в Океании со следами в Восточной Евразии, [ 64 ] и от неустановленной африканской линии архаичных людей в Африке к югу от Сахары, а также от смешанных видов неандертальцев и денисовцев в Азии и Океании. [ 65 ] [ 66 ] [ 67 ] [ 68 ] |
| 50–25 раз |
 Поведенческая современность развивается к этому времени или раньше, согласно теории «большого скачка вперед» . [ 69 ] Вымирание Homo floresiensis . [ 70 ] Мутация М168 (переносится всеми мужчинами неафриканского происхождения). Появление мт- U и K. гаплогрупп Заселение Европы , заселение Мамонтовой степи Северной Азии . Палеолитическое искусство . Вымирание неандертальцев и других архаичных вариантов человека (с возможным выживанием гибридных популяций в Азии и Африке). Внешний вид Y-гаплогруппы R2 ; мт- J и X. гаплогруппы |
| после 25 лет |  Последний ледниковый максимум ; Эпипалеолит / мезолит / голоцен . Заселение Америки . Внешний вид: Y-гаплогруппа R1a ; мт- V и T. гаплогруппы Различные недавние расхождения, связанные с экологическим давлением, например, светлая кожа у европейцев и жителей Восточной Азии ( KITLG , ASIP ), после 30 тыс. лет назад; [ 71 ] Адаптация инуитов к диете с высоким содержанием жиров и холодному климату, 20 тыс. лет назад. [ 72 ] Вымирание поздно сохранившихся архаичных людей в начале голоцена ( 12 тыс. лет назад). Ускоренная дивергенция из-за давления отбора в популяциях, участвовавших в неолитической революции после 12 тыс. лет назад, например Восточноазиатские типы ADH1B , связанные с одомашниванием риса . [ 73 ] или персистенция лактазы . [ 74 ] [ 75 ] Небольшое уменьшение размера мозга произошло несколько тысяч лет назад. [ нужна ссылка ] |
См. также
[ редактировать ]Ссылки
[ редактировать ]- ^ Финарелли, Джон А.; Клайд, Уильям К. (2004). «Переоценка филогении человекообразных: оценка соответствия морфологических и временных данных». Палеобиология. 30 (4): 614. doi : 10.1666/0094-8373(2004)030<0614:RHPECI>2.0.CO;2
- ^ Паттерсон Н., Рихтер Д.Д., Гнерр С., Ландер Э.С., Райх Д. (2006). «Генетические доказательства сложного видообразования человека и шимпанзе». Природа . 441 (7097): 1103–08. Бибкод : 2006Natur.441.1103P . дои : 10.1038/nature04789 . ПМИД 16710306 . S2CID 2325560 .
- ^ В зависимости от классификации линии Homo heidelbergensis ; 0,8, если неандертальцы классифицируются как H. sapiens neanderthalensis или если H. sapiens определяется кладистически на основании отличия от H. neanderthalensis , 0,3 на основе имеющихся ископаемых свидетельств.
- ^ «Эксперименты с сексом было очень трудно проводить», - сказал Годдард. «В эксперименте нужно сохранять все остальное постоянным, за исключением интересующего аспекта. Это означает, что нельзя использовать никакие высшие организмы, поскольку они заниматься сексом для размножения и, следовательно, не обеспечивать асексуального контроля».
Вместо этого Годдард и его коллеги обратились к одноклеточному организму, дрожжам, чтобы проверить идею о том, что секс позволяет популяциям адаптироваться к новым условиям быстрее, чем бесполые популяции. « Секс ускоряет эволюцию, результаты исследования (URL-адрес доступен 9 января 2005 г.) - ^ Докинз, Р. (2005), Рассказ предка: Паломничество на заре эволюции, Хоутон Миффлин Харкорт, ISBN 978-0-618-61916-0
- ^ «Протероспонгия — редкий пресноводный протист, колониальный представитель хоанофлагеллат». «Протероспонгия сама по себе не является предком губок. Однако она служит полезной моделью того, какими могли быть предки губок и других многоклеточных животных». http://www.ucmp.berkeley.edu/protista/proterospongia.html Университет Беркли
- ^ Малуф, Адам С.; Роуз, Кэтрин В.; Бич, Роберт; Сэмюэлс, Брэдли М.; Калмет, Клэр С.; Эрвин, Дуглас Х.; Пуарье, Джеральд Р.; Яо, Нан; Саймонс, Фредерик Дж. (17 августа 2010 г.). «Возможные окаменелости тел животных в домаринских известняках Южной Австралии». Природа Геонауки . 3 (9): 653–59. Бибкод : 2010NatGe...3..653M . дои : 10.1038/ngeo934 . S2CID 13171894 .
- ^ Монахан-Эрли, Р., Дворжак, AM, и Эйрд, WC (2013). Эволюционное происхождение кровеносной сосудистой системы и эндотелия. Журнал тромбозов и гемостаза , 11 (Приложение 1), 46–66. дои : 10.1111/jth.12253 . ПМИД 23809110 .
- ^ Барнс, Роберт Д. (1982). Зоология беспозвоночных . Филадельфия: Холт-Сондерс Интернэшнл. стр. 1018–26. ISBN 978-0-03-056747-6 .
- ^ «Очевидно, что у позвоночных должны были быть предки, жившие в кембрии, но предполагалось, что они были беспозвоночными предшественниками настоящих позвоночных - протохордовых. Пикайя широко пропагандировалась как старейшая ископаемая протохордовая». Ричард Докинз 2004 Рассказ предка с. 289, ISBN 0-618-00583-8
- ^ Шу, Д.Г.; Луо, ХЛ; Конвей Моррис, С.; Чжан, XL; Ху, SX; Чен, Л.; Хан, Дж.; Чжу, М.; Ли, Ю.; Чен, ЛЗ (1999). «Позвоночные животные нижнего кембрия из южного Китая». Природа . 402 (6757): 42–46. Бибкод : 1999Natur.402...42S . дои : 10.1038/46965 . S2CID 4402854 .
- ^ Чен, JY; Хуанг, ДЮ; Ли, CW (1999). «Раннекембрийское краниатоподобное хордовое». Природа . 402 (6761): 518–22. Бибкод : 1999Natur.402..518C . дои : 10.1038/990080 . S2CID 24895681 .
- ^ Шу, Д.-Г.; Конвей Моррис, С.; Хан, Дж.; Чжан, З.-Ф.; Ясуи, К.; Жанвье, П.; Чен, Л.; Чжан, X.-L.; Лю, Ж.-Н.; Ли, Ю.; Лю, H.-Q. (январь 2003 г.). «Голова и позвоночник раннекембрийского позвоночного Haikouichthys». Природа . 421 (6922): 526–529. Бибкод : 2003Natur.421..526S . дои : 10.1038/nature01264 . ПМИД 12556891 . S2CID 4401274 .
- ^ Удрою И. и Сгура А. (2017). Филогения селезенки. Ежеквартальный обзор биологии, 92 (4), 411–443. https://doi.org/10.1086/695327
- ^ Эллиот Д.Г. (2011) Функциональная морфология покровной системы рыб. В: Фаррелл А.П. (редактор), Энциклопедия физиологии рыб: от генома к окружающей среде, том 1, стр. 476–488. Сан-Диего: Академическая пресса. ISBN 9780080923239 .
- ^ У этих первых позвоночных не было челюстей, как у ныне живущих миксин и миног. Челюстные позвоночные появились 100 миллионов лет спустя, в силурийском периоде. http://www.ucmp.berkeley.edu/vertebrates/vertintro.html Университет Беркли
- ^ Ископаемая челюсть целаканта, найденная в слое возрастом 410 млн лет назад , который был собран недалеко от Бьюкена в Виктории , Восточный Гиппсленд Австралии , в настоящее время является рекордсменом по старейшим целакантам; ему было присвоено название Eoactinistia foreyi. при публикации в сентябре 2006 года [1]
- ^ «Считается, что двоякодышащие рыбы являются ближайшими ныне живущими родственниками четвероногих и имеют с ними ряд важных характеристик. Среди этих признаков - зубная эмаль, отделение легочного кровотока от кровотока тела, расположение костей черепа и наличие четырех конечностей одинакового размера с тем же положением и строением, что и четыре ноги четвероногих». http://www.ucmp.berkeley.edu/vertebrates/sarco/dipnoi.html Университет Беркли
- ^ "предок, которого амфибии делят с рептилиями и нами?" «Эти, возможно, переходные окаменелости были тщательно изучены, среди них Acanthostega , которая, по-видимому, была полностью водной, и Ichthyostega », Ричард Докинз, 2004, «История предков» , с. 250, ISBN 0-618-00583-8
- ^ Экхарт, Л.; Валле, LD; Джагер, К.; Баллаун, К.; Сабо, С.; Нарди, А.; Бухбергер, М.; Германн, М.; Алибарди, Л.; Чахлер, Э. (10 ноября 2008 г.). «Идентификация генов рептилий, кодирующих кератиноподобные белки волос, предполагает новый сценарий эволюционного происхождения волос» . Труды Национальной академии наук . 105 (47): 18419–18423. дои : 10.1073/pnas.0805154105 . ПМК 2587626 . ПМИД 19001262 .
- ^ «Во многих отношениях пеликозавры занимают промежуточное положение между рептилиями и млекопитающими» http://www.ucmp.berkeley.edu/synapsids/pelycosaurs.html Университет Беркли.
- ^ «Тринаксодон, как и любое ископаемое, следует рассматривать как двоюродного брата нашего предка, а не самого предка. Он был членом группы млекопитающих рептилий, называемых цинодонтами. Цинодонты были настолько похожи на млекопитающих, что это заманчиво называть их млекопитающими. Но какая разница, как мы их называем. Они почти идеальные промежуточные звенья». Ричард Докинз 2004 Рассказ предка с. 211, ISBN 0-618-00583-8
- ^ Вернебург, Ингмар; Спикман, Стефан Н.Ф. (2018). 4. Эмбриология и органогенез млекопитающих. В: Захос, Фрэнк; Ашер, Роберт. Эволюция, разнообразие и систематика млекопитающих. Берлин: Вальтер де Грютер, 59–116. DOI: https://doi.org/10.1515/9783110341553-004.
- ^ «Ископаемые, которые могут помочь нам реконструировать то, на что был похож Копредок 8, включают большую группу, называемую плезиадапи-формами. Они жили примерно в нужное время и обладают многими качествами, которые можно ожидать от великого предка всех приматов» Ричард Докинз 2004 Сказка предка с. 136, ISBN 0-618-00583-8
- ^ Ренне, Пол Р.; Дейно, Алан Л.; Хильген, Фредерик Дж.; Койпер, Клаудия Ф.; Марк, Даррен Ф.; Митчелл, Уильям С.; Морган, Лия Э.; Мундил, Роланд; Смит, январь (7 февраля 2013 г.). «Временные масштабы критических событий на границе мела и палеогена» . Наука . 339 (6120): 684–87. Бибкод : 2013Sci...339..684R . дои : 10.1126/science.1230492 . ПМИД 23393261 . S2CID 6112274 .
- ^ «Палеонтологи обнаруживают скелет самого примитивного примата» , Phys.org (23 января 2007 г.).
- ^ Алан де Кейрос, Путешествие обезьяны , Basic Books, 2014. ISBN 9780465020515
- ^ «Новый вид приматов у корня древа современных гоминоидов» . физ.орг . Проверено 29 мая 2020 г.
- ^ Раума, Райан; Стернера, К. (2005). «Даты дивергенции катарских приматов рассчитаны на основе полных митохондриальных геномов» (PDF) . Журнал эволюции человека . 48 (3): 237–57. дои : 10.1016/j.jhevol.2004.11.007 . ПМИД 15737392 .
- ^ Бёме, Мадлен; Спасов, Николай; Фусс, Йохен; Трёшер, Адриан; Дин, Эндрю С.; Прието, Жером; Киршер, Уве; Лехнер, Томас; Бегун, Дэвид Р. (ноябрь 2019 г.). «Новая миоценовая обезьяна и передвижение предка человекообразных обезьян и человека» . Природа . 575 (7783): 489–493. Бибкод : 2019Natur.575..489B . дои : 10.1038/s41586-019-1731-0 . ISSN 1476-4687 . ПМИД 31695194 . S2CID 207888156 .
- ^ Попадин Константин; Гунбин, Константин; Пешкин, Леонид; Аннис, София; Флейшманн, Зоя; Крайцберг, Геня; Маркузон, Наталья; Акерманн, Ребекка Р.; Храпко, Константин (19 октября 2017 г.). «Митохондриальные псевдогены предполагают повторную межвидовую гибридизацию в эволюции гоминид» . bioRxiv : 134502. doi : 10.1101/134502 . hdl : 11427/36660 .
- ^ Перлман, Дэвид (12 июля 2001 г.). «Окаменелости из Эфиопии могут быть самыми ранними предками человека» . Национальные географические новости. Архивировано из оригинала 15 июля 2001 года.
Другой соавтор — Тим Д. Уайт, палеоантрополог из Калифорнийского университета в Беркли, который в 1994 году обнаружил окаменелость дочеловеческой эпохи, названную Ardipithecus ramidus, которая на тот момент была самой древней из известных — 4,4 миллиона человек. годы.
- ^ Уайт, Тим Д.; Асфау, Берхане; Бейене, Йонас; Хайле Селассие, Джон; Лавджой, К. Оуэн; Вор, генерал; ВольдеГабриэль, Путеводитель (2009). « Ardipithecus ramidus и палеобиология ранних гоминидов». Наука 326 (5949): 75–86. Бибкод : 2009Sci...326... 75W дои : 10.1126/science.1175802 . ПМИД 19810190 . S2CID 20189444 .
- ^ Харманд, Соня; Льюис, Джейсон Э.; Фейбел, Крейг С.; Заяц, Кристофер Дж.; Прат, Сандрин; Ленобль, Арно; Боэс, Ксавье; Куинн, Ронда Л.; Брене, Мишель; Арройо, Адриан; Тейлор, Николас; Клемент, Софи; Давер, Гийом; Бругаль, Жан-Филип; Лики, Луиза; Мортлок, Ричард А.; Райт, Джеймс Д.; Локороди, Сэмми; Кирва, Кристофер; Кент, Деннис В.; Рош, Элен (2015). «Каменные орудия возрастом 3,3 миллиона лет из Ломекви 3, Западная Туркана, Кения». Природы . 521 (7552): 310–15. Бибкод : 2015Natur.521..310H . дои : 10.1038/nature14464 . ПМИД 25993961 . S2CID 1207285 .
- ^ Ракстон, Грэм Д.; Уилкинсон, Дэвид М. (27 декабря 2011 г.). «Избежание перегрева и отбор как выпадения волос, так и двуногости у гомининов» . Труды Национальной академии наук . 108 (52): 20965–20969. Бибкод : 2011PNAS..10820965R . дои : 10.1073/pnas.1113915108 . ISSN 0027-8424 . ПМК 3248486 . ПМИД 22160694 .
- ^ Чжу, Чжаоюй, Робин; Ву, И; Ян, Шися; Хоу, Ямей; Хан, Цзянвэй, Тинпин (2018). Китайского Лёссового плато примерно 2,1 миллиона лет назад». Nature . 559 (7715): 608–12. Бибкод : 2018Natur.559..608Z . doi : 10.1038/ . PMID 29995848. S2CID 49670311 s41586-018-0299-4 .
- ^ НОВА: Стать человеком. Часть 2 http://video.pbs.org/video/1319997127/
- ^ Яблонски, Нина Г. (октябрь 2004 г.). «Эволюция кожи человека и цвета кожи». Ежегодный обзор антропологии . 33 (1): 585–623. дои : 10.1146/annurev.anthro.33.070203.143955 . S2CID 53481281 .
- ^ Бермудес де Кастро, JM (30 мая 1997 г.). «Гоминид из нижнего плейстоцена Атапуэрки, Испания: возможный предок неандертальцев и современных людей». Наука . 276 (5317): 1392–1395. дои : 10.1126/science.276.5317.1392 . ПМИД 9162001 .
- ^ Грин, Ричард Э.; Краузе, Йоханнес; Птак, Сьюзен Э.; Бриггс, Адриан В.; Ронан, Майкл Т.; Саймонс, Ян Ф.; Ду, Лей; Эгхольм, Майкл; Ротберг, Джонатан М.; Паунович, Майя; Паабо, Сванте (ноябрь 2006 г.). «Анализ одного миллиона пар оснований ДНК неандертальца» . Природа . 444 (7117): 330–336. Бибкод : 2006Natur.444..330G . дои : 10.1038/nature05336 . ПМИД 17108958 . S2CID 4320907 .
- ^ " Рубин также сказал, что анализ, проведенный на данный момент, показывает, что ДНК человека и неандертальца идентичны примерно на 99,5-99,9 процента " . Кость неандертальца дает подсказки ДНК (URL-адрес доступен 16 ноября 2006 г.)
- ^ " Вывод: старая поговорка о том, что мы разделяем 98,5% последовательности нашей ДНК с шимпанзе, вероятно, ошибочна. Для этого образца лучшей оценкой было бы то, что 95% пар оснований в точности совпадают между ДНК шимпанзе и человека. " Бриттен, Р.Дж. (2002). «Расхождение между образцами последовательностей ДНК шимпанзе и человека составляет 5%, считая инделы» . ПНАС . 99 (21): 13633–35. Бибкод : 2002PNAS...9913633B . дои : 10.1073/pnas.172510699 . ПМК 129726 . ПМИД 12368483 .
- ^ « ...из трех миллиардов букв, составляющих геном человека, только 15 миллионов — менее 1 процента — изменились за шесть миллионов лет или около того с тех пор, как линии человека и шимпанзе разошлись » . Поллард, Канзас (2009). «Что делает нас людьми?». Научный американец . 300–5 (5): 44–49. Бибкод : 2009SciAm.300e..44P . doi : 10.1038/scientificamerican0509-44 . ПМИД 19438048 . S2CID 38866839 .
- ^ Краузе Дж., Лалуэса-Фокс С., Орландо Л., Энард В., Грин Р.Э., Бурбано Х.А., Хублин Дж.Дж., Ханни С., Фортеа Дж., де ла Расилья М., Бертранпетит Дж., Росас А., Паабо С. (ноябрь 2007 г.). «Производный вариант FOXP2 современных людей был использован неандертальцами». Курс. Биол . 17 (21): 1908–12. Бибкод : 2007CBio...17.1908K . дои : 10.1016/j.cub.2007.10.008 . hdl : 11858/00-001M-0000-000F-FED3-1 . ПМИД 17949978 . S2CID 9518208 .
- Николас Уэйд (19 октября 2007 г.). «У неандертальцев был важный речевой ген, как показывают данные ДНК» . Нью-Йорк Таймс .
- ^ Штейн, Ричард А. (октябрь 2015 г.). «Анализ количества копий начинает прибавляться». Новости генной инженерии и биотехнологии . 35 (17): 20, 22–23. дои : 10.1089/gen.35.17.09 .
- ^ Мейер, Матиас; Арсуага, Хуан Луис; Де Филиппо, Чезаре; Нагель, Сара; Аксиму-Петри, Айинуэр; Никель, Биргит; Мартинес, Игнасио; Грасия, Ана; Де Кастро, Хосе Мария Бермудес; Карбонелл, Эудальд; Виола, Бенс; Келсо, Джанет; Прюфер, Кей; Паабо, Сванте (март 2016 г.). «Последовательности ядерной ДНК гомининов Sima de los Huesos среднего плейстоцена». Природы . 531 (7595): 504–07. Бибкод : 2016Natur.531..504M . дои : 10.1038/nature17405 . ПМИД 26976447 . S2CID 4467094 .
- ^ Каллауэй, Юэн (2016). «ДНК древнейшего человека детализирует рассвет неандертальцев» . Природа . 531 (7594): 296–86. Бибкод : 2016Natur.531..296C . дои : 10.1038/531286a . ПМИД 26983523 . S2CID 4459329 .
- ^ Миетто, Паоло; Аванзини, Марко; Роланди, Джузеппе (2003). «Палеонтология: следы человека в вулканическом пепле плейстоцена» . Природа . 422 (6928): 133. Бибкод : 2003Natur.422..133M . дои : 10.1038/422133a . ПМИД 12634773 . S2CID 2396763 .
- ^ Тиммерманн, А., Юн, К.С., Райя, П. и др. Влияние климата на архаичные места обитания человека и смену видов. Природа 604 , 495–501 (2022). https://doi.org/10.1038/s41586-022-04600-9
- ^ Каллауэй, Юэн (7 июня 2017 г.). «Самые древние ископаемые останки Homo sapiens переписывают историю нашего вида». Природа . дои : 10.1038/nature.2017.22114 .
- ^ Трайон, Кристофер А.; Вера, Тайлер (2013). «Изменчивость среднего каменного века Восточной Африки» (PDF) . Современная антропология . 54 (8): 234–54. дои : 10.1086/673752 . S2CID 14124486 .
- ^ Циммер, Карл (10 июля 2019 г.). «Кость черепа, обнаруженная в Греции, может изменить историю предыстории человечества. Кость, найденная в пещере, является старейшим ископаемым современного человека, когда-либо обнаруженным в Европе. Она намекает на то, что люди начали покидать Африку гораздо раньше, чем когда-то считалось» . Нью-Йорк Таймс . Проверено 11 июля 2019 г.
- ^ Персонал (10 июля 2019 г.). « Самые старые останки за пределами Африки обнуляют часы миграции людей» . Физика.орг . Проверено 10 июля 2019 г.
- ^ Харвати, Катерина; и др. (10 июля 2019 г.). «Окаменелости пещеры Апидима являются самыми ранними свидетельствами существования Homo sapiens в Евразии» . Природа . 571 (7766): 500–504. дои : 10.1038/s41586-019-1376-z . ПМИД 31292546 . S2CID 195873640 .
- ^ Хайнц, Таня; Пала, Мария; Гомес-Карбалья, Альберто; Ричардс, Мартин Б.; Салас, Антонио (март 2017 г.). «Обновление дерева митохондриальной ДНК человека в Африке: актуальность для судебно-медицинской экспертизы и популяционной генетики». Международная судебно-медицинская экспертиза: Генетика . 27 : 156–159. дои : 10.1016/j.fsigen.2016.12.016 . ПМИД 28086175 .
- ^ Позник, Г.Д.; Хенн, Б.М.; Да, М.-К.; Сливерска, Э.; Ойскирхен, генеральный директор; Лин, А.А.; Снайдер, М.; Кинтана-Мурси, Л.; Кидд, Дж. М.; Андерхилл, Пенсильвания; Бустаманте, CD (1 августа 2013 г.). «Секвенирование Y-хромосом устраняет несоответствие во времени общему предку мужчин и женщин» . Наука . 341 (6145): 562–565. Бибкод : 2013Sci...341..562P . дои : 10.1126/science.1237619 . ПМК 4032117 . ПМИД 23908239 .
- ^ Кармин, Моника; Сааг, Лори; Висенте, Марио; Сэйрес, Мелисса А. Уилсон; Ярве, Мари; Талас, Ульви Герст; Рутси, Сирия; Илумяэ, Анне-Май; Магия, Ридик; Митт, Марио; Пагани, Лука; Пууранд, Тармо; Фалтыскова, Зузана; Клементе, Флориан; Кардона, Алексия; Мецпалу, Эне; Саакян, Оганес; Юнусбаев, Баязит; Худжашов, Георгий; ДеДжорджио, Майкл; Лугвяли, Ева-Лийс; Эйхштедт, Кристина; Элметс, Микк; Чаубей, Гьянешвар; Тамбетс, Кристина; Литвинов Сергей; Мормина, Мару; Сюэ, Яли; Аюб, Касим; и др. (апрель 2015 г.). «Недавнее ограничение разнообразия Y-хромосом совпало с глобальными изменениями в культуре» . Геномные исследования . 25 (4): 459–466. дои : 10.1101/гр.186684.114 . ПМЦ 4381518 . ПМИД 25770088 .
- ^ Кларк, Дж. Десмонд; Бейене, Йонас; ВольдеГабриэль, Гидеон; Харт, Уильям К.; Ренне, Пол Р.; Гилберт, Генри; Дефлер, Альбан; Вор, генерал; Като, Сигэхиро; Людвиг, Кеннет Р.; Мясная лавка, Джин Фокс; Асфау, Берхане; Уайт, Тим Д. (июнь 2003 г.). «Стратиграфический, хронологический и поведенческий контекст плейстоценового человека разумного из Среднего Аваша, Эфиопия». Природа 423 (6941): 747–752. Бибкод : 2003Nature.423..747C . дои : 10.1038/nature01670 . ПМИД 12802333 . S2CID 4312418 .
- ^ Шерри, Элеонора (2017). «Археология каменного века Западной Африки» . Оксфордская исследовательская энциклопедия истории Африки . doi : 10.1093/acrefore/9780190277734.013.137 . ISBN 9780190277734 .
- ^ Хеншилвуд, К.С. и Б. Дюбрей, 2009. Чтение артефактов: приобретение языковых навыков из среднего каменного века на юге Африки. В Р. Боте и К. Найте (ред.), «Колыбель языка». Оксфорд: Издательство Оксфордского университета, стр. 41–61. ISBN 9780191567674 .
- ^ Боулер Дж.М., Джонстон Х., Олли Дж.М., Прескотт Дж.Р., Робертс Р.Г., Шокросс В., Спунер Н.А. (2003). «Новый век человеческой деятельности и изменение климата на озере Мунго, Австралия». Природа . 421 (6925): 837–40. Бибкод : 2003Natur.421..837B . дои : 10.1038/nature01383 . ПМИД 12594511 . S2CID 4365526 .
- ^ Ричард Э. Грин; Краузе, Дж.; Бриггс, AW; Маричич, Т.; Стензель, У.; Кирхер, М.; Паттерсон, Н.; Ли, Х.; и др. (2010). «Проект последовательности генома неандертальца» . Наука . 328 (5979): 710–22. Бибкод : 2010Sci...328..710G . дои : 10.1126/science.1188021 . ПМК 5100745 . ПМИД 20448178 .
- ^ Ринкон, Пол (6 мая 2010 г.). «Гены неандертальцев «выживают в нас» » . Новости Би-би-си . Би-би-си . Проверено 7 мая 2010 г.
- ^ Шанкарараман, Шрирам; Маллик, Свапан; Паттерсон, Ник; Райх, Дэвид (2016). «Объединенный ландшафт денисовского и неандертальского происхождения у современного человека» . Современная биология . 26 (9): 1241–1247. Бибкод : 2016CBio...26.1241S . дои : 10.1016/j.cub.2016.03.037 . ПМЦ 4864120 . ПМИД 27032491 .
- ^ Каллауэй, Юэн (26 июля 2012 г.). «Геномы охотников-собирателей - кладезь генетического разнообразия». Природа . дои : 10.1038/nature.2012.11076 . S2CID 87081207 .
- ^ Лашанс, Джозеф; Верно, Бенджамин; Элберс, Клара С.; Ферверда, Барт; Фроман, Ален; Бодо, Жан-Мари; Лема, Годфри; Фу, Вэньцин; Ньямбо, Томас Б.; Реббек, Тимоти Р.; Чжан, Кун; Эйки, Джошуа М.; Тишкофф, Сара А. (август 2012 г.). «Эволюционная история и адаптация на основе полногеномных последовательностей разнообразных африканских охотников-собирателей» . Клетка . 150 (3): 457–469. дои : 10.1016/j.cell.2012.07.009 . ПМК 3426505 . ПМИД 22840920 .
- ^ Сюй, Дуо; Павлидис, Павлос; Таскент, Реджеп Озгур; Алахиотис, Николаос; Фланаган, Колин; ДеДжорджио, Майкл; Блехман, Ран; Руль, Стефан; Гоккумен, Омер (октябрь 2017 г.). «Архаичная интрогрессия гомининов в Африке способствует функциональной генетической вариации слюны MUC7» . Молекулярная биология и эволюция . 34 (10): 2704–2715. дои : 10.1093/molbev/msx206 . ПМК 5850612 . ПМИД 28957509 .
- ^ Мондал, Маюх; Бертранпети, Жауме; Лао, Оскар (16 января 2019 г.). «Приблизительные байесовские вычисления с глубоким обучением поддерживают третью архаичную интрогрессию в Азии и Океании» . Природные коммуникации . 10 (1): 246. Бибкод : 2019NatCo..10..246M . дои : 10.1038/s41467-018-08089-7 . ПМК 6335398 . ПМИД 30651539 .
- ^ Кляйн, Ричард (1995). «Анатомия, поведение и происхождение современного человека». Журнал мировой предыстории . 9 (2): 167–98. дои : 10.1007/bf02221838 . S2CID 10402296 .
- ^ Сутикна, Томас; Точери, Мэтью В.; Морвуд, Майкл Дж.; Саптомо, Э. Вахью; Джатмико; Эй, Рокус Дуэ; Васисто, Шри; Вестэуэй, Кира Э.; Обер, Максим; Ли, Бо; Чжао, Цзянь-синь; Стори, Майкл; Аллоуэй, Брент В.; Морли, Майк В.; Мейер, Ханнеке Дж. М.; ван ден Берг, Геррит Д.; Грюн, Райнер; Доссето, Энтони; Брамм, Адам ; Юнгерс, Уильям Л.; Робертс, Ричард Г. (30 марта 2016 г.). «Пересмотренная стратиграфия и хронология Homo floresiensis в Лян Буа в Индонезии» . Природа . 532 (7599): 366–69. Бибкод : 2016Natur.532..366S . дои : 10.1038/nature17179 . ПМИД 27027286 . S2CID 4469009 .
- ^ КРАСОТА, Сандра; САНТОС, AM; МакЭвой, Б.; АЛВЕС, И.; МАРТИН, К.; Кэмерон, Э.; Шрайвер, доктор медицины; ПАРРА, Э.Дж.; Роча, Дж. (2012). «Сроки осветления пигментации у европейцев» . Молекулярная биология и эволюция . 30 (1): 24–35. дои : 10.1093/molbev/mss207 . ПМЦ 3525146 . ПМИД 22923467 .
- ^ Фумагалли, М.; Мольтке, И.; Граруп, Н.; Расимо, Ф.; Бьеррегард, П.; Йоргенсен, Мэн; Корнелиуссен, Т.С.; Гербо, П.; Скотт, Л.; Линнеберг, А.; Кристенсен, К.; Брандслунд, И.; Йоргенсен, Т.; Уэрта-Санчес, Э.; Шмидт, Е.Б.; Педерсен, О.; Хансен, Т.; Альбрехцен, А.; Нильсен, Р. (17 сентября 2015 г.). «Гренландские инуиты демонстрируют генетические особенности диеты и адаптации к климату». Наука . 349 (6254): 1343–1347. Бибкод : 2015Sci...349.1343F . дои : 10.1126/science.aab2319 . hdl : 10044/1/43212 . ПМИД 26383953 . S2CID 546365 .
- ^ Пэн, Йи; Ши, Хун; Ци, Сюэ-бин; Сяо, Чун-цзе; Чжун, Хуа; Ма, Ран-лин З; Су, Бинг (2010). «Полиморфизм ADH1B Arg47His в популяциях Восточной Азии и расширение одомашнивания риса в истории» . Эволюционная биология BMC . 10 (1): 15. Бибкод : 2010BMCEE..10...15P . дои : 10.1186/1471-2148-10-15 . ПМЦ 2823730 . ПМИД 20089146 .
- ^ Сегюрель, Лор; Бон, Селин (31 августа 2017 г.). «Об эволюции персистенции лактазы у человека». Ежегодный обзор геномики и генетики человека . 18 (1): 297–319. doi : 10.1146/annurev-genom-091416-035340 . ПМИД 28426286 .
- ^ Ингрэм, Кэтрин Дж. Э.; Малкаре, Шарлотта А.; Итан, Юваль; Томас, Марк Г.; Своллоу, Даллас М. (26 ноября 2008 г.). «Переваривание лактозы и эволюционная генетика устойчивости лактазы». Генетика человека . 124 (6): 579–591. дои : 10.1007/s00439-008-0593-6 . ПМИД 19034520 . S2CID 3329285 .
Внешние ссылки
[ редактировать ]- Палеос
- Беркли Эволюция
- История эволюции животных. Архивировано 27 июня 2016 г. в Wayback Machine.
- Веб-проект «Древо жизни» – исследуйте полное филогенетическое дерево в интерактивном режиме.
- Хронология человечества (интерактивная) – Смитсоновский институт , Национальный музей естественной истории (август 2016 г.).