Плотоядный
| Хищники | |
|---|---|
 | |
| Различные хищники: кошачьи слева и псовые справа. | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Клэйд : | Карнивораморфа |
| Клэйд : | Карниворообразные |
| Заказ: | Плотоядный Боудич , 1821 г. [2] |
| Подзаказы | |
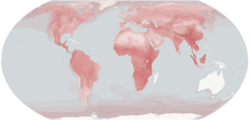 | |
| Современное распространение и плотность видов Carnivora. | |
| Синонимы | |
список синонимов: | |
Carnivora / k ɑːr ˈ n ɪ v ər ə / — отряд плацентарных млекопитающих , специализирующихся преимущественно на поедании мяса, представители которого формально называются хищниками . Отряд Carnivora — пятый по величине отряд млекопитающих, включающий не менее 279 видов на всех основных участках суши и в самых разных средах обитания, от холодных полярных регионов Земли до гиперзасушливых регионов пустыни Сахара и открытых морей. Эти млекопитающие имеют большое количество разнообразных строений тела с большим разнообразием форм и размеров.
Хищные животные делятся на два подотряда : Feliformia , в который входят настоящие кошачьи и несколько «кошачьих» животных; и Caniformia , содержащая настоящих псовых и многих «собачьих» животных.К кошачьим относятся семейства Felidae , Viverridae , гиены и мангусты , большинство из которых обитает только в Старом Свете ; кошки — единственное исключение, встречающееся в старом и новом свете и попадающее в Америку через Берингов мост . К псовым относятся Caninae , Procyonidae , медведи , куньи , скунсы и ластоногие , которые встречаются по всему миру с огромным разнообразием в их морфологии, диете и поведении.
Этимология
[ редактировать ]Слово «хищник» происходит от латинского caro (основа carn- ) «плоть» и vorāre «пожирать» и относится к любому мясоедящему организму.
Филогения
[ редактировать ]Самые старые известные млекопитающие линии хищных животных ( Carnivoramorpha ) появились в Северной Америке через 6 миллионов лет после мел-палеогенового вымирания . [22] [23] Эти ранние предки хищных животных напоминали маленьких ласок или генетоподобных млекопитающих, ведущих ночной образ жизни на лесной подстилке или на деревьях, тогда как другие группы млекопитающих, такие как мезонихи , а позже и креодонты, занимали нишу мегафауны фауноядных. Однако после вымирания мезонихий и оксиенид -креодонтов в конце эоцена в эту нишу быстро переместились хищные животные, причем такие формы, как нимравиды, были доминирующими крупнотелыми хищниками, устраивавшими засады в олигоцене наряду с гиенодонтами -креодонтами (которые аналогичным образом производили более крупных хищников). , более открытые формы в начале олигоцена). К моменту наступления миоценовой эпохи большинство, если не все, основные линии и семейства хищных животных диверсифицировались и стали наиболее доминирующей группой крупных наземных хищников в Евразии и Северной Америке, при этом различные линии успешно занимали ниши фауны мегафауны в разные периоды времени. миоцена и более поздних эпох.
Систематика
[ редактировать ]Эволюция
[ редактировать ]
Отряд Carnivora принадлежит к группе млекопитающих, известной как Laurasiatheria , которая также включает в себя другие группы, такие как летучие мыши и копытные . [24] [25] Внутри этой группы хищные животные отнесены к кладе Ferae . Ferae включает в себя ближайшего из ныне живущих родственников хищников, панголинов , а также несколько вымерших групп преимущественно палеогеновых плотоядных плацентарных, таких как креодонты , арктоционии и мезонихи . [26] Первоначально креодонты считались сестринским таксоном хищных животных, возможно, даже их предком, на основании наличия хищных зубов . [27] но природа хищных зубов у этих двух групп различна. У хищных животных хищные зубы располагаются ближе к передней части ряда коренных зубов, а у креодонтов - ближе к задней части ряда коренных зубов. [28] и это предполагает отдельную эволюционную историю и различие на уровне порядка. [29] Кроме того, недавний филогенетический анализ показывает, что креодонты более тесно связаны с панголинами, в то время как мезонихи могут быть сестринской группой хищных животных и их стволовых родственников. [26]
Ближайшие стеблево-хищные животные — миакоиды . К миакоидам относятся семейства Viverravidae и Miacidae , а вместе Carnivora и Miacoidea образуют стволовую кладу Carnivoramorpha . Миакоиды представляли собой небольшие генетически-подобные хищники, занимавшие различные ниши, такие как наземные и древесные среды обитания. Недавние исследования показали подтверждающее количество доказательств того, что Miacoidea представляет собой эволюционную степень хищных животных, которая, хотя виверравиды представляют собой монофилетическую базальную группу, миациды являются парафилетическими по отношению к Carnivora (как показано в филогении ниже). [30] [31]
Carnivoramorpha в целом впервые появились в палеоцене Северной Америки около 60 миллионов лет назад. [23] Коронные хищники впервые появились около 42 миллионов лет назад в среднем эоцене . [32] что современные Carnivora представляют собой монофилетическую группу, коронную группу Carnivoramorpha Их молекулярная филогения показывает , . [33] Отсюда хищные животные разделились на две клады в зависимости от состава костных структур, окружающих среднее ухо черепа: кошачьих кошачьих и собачьих псовых . [34] У кошачьих слуховые буллы двухкамерные, состоят из двух костей, соединенных перегородкой . Слуховые буллы псовых однокамерные или частично разделенные, состоят из одной кости. [35] Первоначально ранние представители хищных животных были небольшими, так как креодонты (в частности, оксиениды) и мезонихии доминировали в нишах вершинных хищников в течение эоцена, но в олигоцене карниворы стали доминирующей группой высших хищников вместе с нимравидами , а к миоцену большинство современных семейств хищников диверсифицировались и стали основными наземными хищниками в Северном полушарии.
Классификация современных хищников
[ редактировать ]В 1758 году шведский ботаник Карл Линней поместил всех известных в то время хищников в группу Ferae (не путать с современной концепцией Ferae, включающей также ящеров) в десятом издании своей книги Systema Naturae . Он выделил шесть родов: Canis (псовые и гиениды), Phoca (ластоногие), Felis (кошачьи), Viverra (виверровые, герпестиды и мефитиды), Mustela (куньи, не барсучьи), Ursus (медведи, крупные виды куньих и проциониды). [19] Лишь в 1821 году английский писатель и путешественник Томас Эдвард Боудич дал группе современное и общепринятое название. [2]
Первоначально современное представление о хищных было разделено на два подотряда: наземные Fissipedia и морские ластоногие . [36] Ниже приведена классификация того, как существующие семьи были связаны друг с другом после американского палеонтолога Джорджа Гейлорда Симпсона в 1945 году: [36]
- Орден Карнивора Боудича, 1821 г.
- Подотряд Fissipedia Blumenbach, 1791 г.
- Надсемейство Canoidea Г. Фишер де Вальдхайм, 1817 г.
- Семейство Canidae G. Fischer de Waldheim, 1817 – собаки.
- Семейство Ursidae G. Fischer de Waldheim, 1817 – медведи.
- Семейство Procyonidae Bonaparte, 1850 — еноты и панды.
- Семейство Mustelidae G. Fischer de Waldheim, 1817 - скунсы, барсуки, выдры и ласки.
- Надсемейство Feloidea G. Fischer de Waldheim, 1817 г.
- Семейство Viverridae JE Gray, 1821 – циветты и мангусты.
- Семейство Hyaenidae JE Grey, 1821 - гиены.
- Семейство Felidae G. Fischer de Waldheim, 1817 – кошки.
- Надсемейство Canoidea Г. Фишер де Вальдхайм, 1817 г.
- Подотряд Pinnipedia Iliger, 1811 г.
- Семейство Otariidae JE Gray, 1825 – ушастые тюлени.
- Семейство Odobenidae JA Allen, 1880 – моржи.
- Семейство Phocidae JE Grey, 1821 - безухие тюлени.
- Подотряд Fissipedia Blumenbach, 1791 г.
Однако с тех пор методы, которые маммологи используют для оценки филогенетических взаимоотношений между семействами хищных животных, были улучшены за счет более сложного и интенсивного учета генетики, морфологии и летописи окаменелостей. Исследования филогении Carnivora с 1945 года показали, что Fisspedia является парафилетической по отношению к ластоногим. [37] при этом ластоногие более тесно связаны с медведями или ласками. [38] [39] [40] [41] [42] Мелкие хищные семейства Viverridae, [43] Было обнаружено, что Procyonidae и Mustelidae являются полифилетическими :
- мангусты и несколько малагасийских Обнаружено, что эндемичных видов принадлежат к кладе с гиенами, причем малагасийские виды относятся к отдельному семейству Eupleridae . [44] [45] [46]
- Африканская пальмовая циветта — это хищный кошачий хищник. [47]
- Линсанг . более близок к кошкам [48]
- Панды не являются проционидами и не являются естественной группой. [49] Большая панда – настоящий медведь [50] [51] в то время как красная панда — это отдельное семейство. [52]
- Скунсы и барсуки-вонючки отнесены к отдельному семейству и являются сестринской группой клады, включающей Ailuridae, Procyonidae и Mustelidae sensu stricto . [53] [52]
Ниже представлена таблица современных семейств хищных животных и количества современных видов, признанных различными авторами первого (2009 г.). [54] ) и четвёртый (2014 г.) [55] ) тома Справочника млекопитающих мира :
| Плотоядный Боудич, 1821 г. | |||||
| Фелиформия Крецой, 1945 г. | |||||
| Нандиниоидея Покок, 1929 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
|---|---|---|---|---|---|
| Нандинииды Покок, 1929 г. | Африканская пальмовая циветта | Африка к югу от Сахары | 1 | Нандиния бинотата (Дж. Э. Грей, 1830 г.) | |
| Фелодея Г. Фишер де Вальдхайм, 1817 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Felidae Г. Фишер де Вальдхайм, 1817 г. | Кошки | Америка , Африка и Евразия (завезен на Мадагаскар , Австралазию и несколько островов) | 37 | Фелис катус Линней, 1758 г. |  |
| Прионодонтиды Хорсфилд, 1822 г. | Линсанг | Индомалайское царство | 2 | Прионодон линзанг (Хардвик, 1821 г.) | |
| Виверродея Дж. Э. Грей, 1821 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Viverridae Дж. Э. Грей, 1821 г. | Циветты, наездники и гуси | Южная Европа , Индомалайская область и Африка (завезена на Мадагаскар ) | 34 | Виверра Зибета Линней, 1758 г. |  |
| Герпестоидея Бонапарт, 1845 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Hyaenidae JE Grey, 1821 г. | Гиены | Африка , Ближний Восток , Кавказ , Центральная Азия и Индийский субконтинент. | 4 | Гиена гиена (Линней, 1758 г.) |  |
| Герпестиды Бонапарта, 1845 г. | Мангусты | Пиренейский полуостров , Африка , Ближний Восток , Кавказ , Средняя Азия и Индомалайское царство. | 34 | Herpestes ichneumon (Линней, 1758 г.) |  |
| Eupleridae Chenu, 1850 г. | Малагасийские мангусты и циветты | Мадагаскар | 8 | Eupleres goudotii Дойер, 1835 г. |  |
| Каниформия Крецой, 1945 г. | |||||
| Каноидея Г. Фишер де Вальдхайм, 1817 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Canidae Г. Фишер де Вальдхайм, 1817 г. | Собаки | Америка , Африка и Евразия (завезен на Мадагаскар , Австралазию и несколько островов) | 35 | Семейная собака Линнеев, 1758 г. |  |
| Урсодея Г. Фишер де Вальдхайм, 1817 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Ursidae Г. Фишер де Вальдхайм, 1817 г. | Медведи | Америки и Евразия | 8 | Урсус Арктос Линней, 1758 г. |  |
| Фокоидея Дж. Э. Грей, 1821 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Odobenidae Дж. А. Аллен, 1880 г. | Морж | Северный полюс в Северном Ледовитом океане и субарктических морях Северного полушария. | 1 | Одобенус розмарус (Линней, 1758 г.) |  |
| Otariidae JE Grey, 1825 г. | Ушастые тюлени | Приполярные, умеренные и экваториальные воды Тихого и Южного океанов, а также южной части Индийского и Атлантического океанов. | 15 | Otaria flavescens (Линней, 1758 г.) |  |
| Phocidae Дж. Э. Грей, 1821 г. | Безухие тюлени | Море озеро и Байкал | 18 | Фока витулина Линней, 1758 г. |  |
| Musteloidea Г. Фишер де Вальдхайм, 1817 г. | |||||
| Семья | Английское имя | Распределение | Количество существующих видов | Тип Таксон | Изображение Рисунок |
| Мефитиды Бонапарт, 1845 г. | Скунсы и вонючие барсуки | Америка , западные Филиппины , Индонезия и Малайзия. | 12 | Мефит Мефит (Шребер, 1776 г.) |  |
| Ailuridae JE Grey, 1843 г. | Красная Панда | Восточные Гималаи и юго-западный Китай | 1 | Ailurus fulgens Ф. Кювье, 1825 г. |  |
| Procyonidae Дж. Э. Грей, 1825 г. | Еноты | Америка (завезена в Европу , на Кавказ и в Японию ) | 12 | Процион Лотор (Линней, 1758 г.) |  |
| Mustelidae G. Фишер де Вальдхайм, 1817 г. | Ласки, выдры и барсуки | Америка , Африка и Евразия (завезен в Австралазию и на несколько островов) | 57 | Мустела горностай Линней, 1758 г. |  |
Анатомия
[ редактировать ]Череп
[ редактировать ]
Клыки обычно большие, конические, толстые и устойчивые к нагрузкам. Все наземные виды хищных животных имеют по три резца с каждой стороны каждой челюсти (исключение составляет калан ( Enhydra lutris ), у которого только два нижних резца). [56] Третий моляр утрачен. Хищная пара состоит из четвертого верхнего премоляра и первых нижних коренных зубов. Как и у большинства млекопитающих, зубной ряд гетеродонтный , хотя у некоторых видов, таких как трубкозуб ( Proteles cristata ), зубы сильно редуцированы, а щечные зубы специализированы для поедания насекомых. У ластоногих зубы гомодонтные , поскольку они эволюционировали, чтобы хватать или ловить рыбу, а щечные зубы часто утрачиваются. [57] У медведей и енотов хищная пара вторично редуцирована. [57] Череп массивный, с сильной скуловой дугой . Часто сагиттальный гребень присутствует , иногда более выраженный у видов с половым диморфизмом, таких как морские львы и морские котики , хотя у некоторых мелких хищников он также значительно редуцирован. [57] Черепная коробка увеличена, лобно-теменная кость находится спереди. У большинства видов глаза находятся на передней части лица. У псовых рострум обычно длинный с множеством зубов, а у кошачьих он короче и с меньшим количеством зубов. Хищные зубы кошачьих обычно более секционные. [ нужны разъяснения ] чем у псовых. Носовые раковины большие и сложные по сравнению с другими млекопитающими, что обеспечивает большую площадь поверхности для обонятельных рецепторов . [57]
Посткраниальная область
[ редактировать ]
Помимо совокупности характеристик зубов и черепа, не так уж много их общей анатомии объединяет хищных животных в группу. [56] Все виды хищных животных четвероногие , у большинства из них пять пальцев на передних лапах и четыре пальца на задних. У наземных хищников на ногах мягкие подушечки. Ступни могут быть пальцевыми , как у кошек, гиен и собак, или стопоходящими, как у медведей, скунсов, енотов, ласк, циветт и мангустов. У ластоногих конечности превратились в ласты .

В отличие от китообразных и сирен , у которых есть полностью функциональный хвост , помогающий им плавать, ластоногие для плавания используют свои конечности под водой. Безухие тюлени используют задние ласты; морские львы и морские котики используют передние ласты, а морж — все конечности. В результате хвосты ластоногих значительно короче, чем у других хищников.
Помимо ластоногих, собаки, медведи, гиены и кошки имеют отчетливую и узнаваемую внешность. Собаки, как правило, бегающие млекопитающие, имеют грациозный внешний вид и часто полагаются на свои зубы, чтобы удерживать добычу; медведи намного крупнее и полагаются на свою физическую силу в поисках пищи. По сравнению с собаками и медведями, кошки имеют более длинные и сильные передние конечности, вооруженные втягивающимися когтями , позволяющими удерживать добычу. Гиены — это кошачьи, похожие на собак, с покатой спиной, поскольку их передние ноги длиннее задних. Семейство енотов и красная панда — маленькие, похожие на медведя хищники с длинными хвостами. Другие мелкие семейства хищных животных Nandiniidae , Prionodontidae , Viverridae , Herpestidae , Eupleridae, Mephitidae и Mustelidae в результате конвергентной эволюции сохранили небольшой, наследственный вид миакоидов, хотя наблюдаются некоторые вариации, такие как крепкая и крепкая физическая форма барсуков и росомахи. ( Гуло гуло ). [56]
У большинства видов хищников есть четко выраженный сезон размножения . [58] Самцы хищных животных обычно имеют бакулы , отсутствующие у гиен и бинтуронгов . [59]
Длина и густота меха варьируются в зависимости от среды обитания вида. У видов теплого климата мех часто короткий и светлый. У видов с холодным климатом мех либо густой, либо длинный, часто с маслянистым веществом, помогающим сохранять тепло. Окраска шерсти у разных видов различается, часто включая черный, белый, оранжевый, желтый, красный и многие оттенки серого и коричневого. Некоторые из них полосатые, пятнистые, пятнистые, полосатые или с другим ярким рисунком. Кажется, существует корреляция между средой обитания и цветовым рисунком; например, пятнистые или полосатые виды, как правило, встречаются в густо засаженных деревьями районах. [56] Некоторые виды, такие как серый волк, полиморфны : разные особи имеют разный цвет шерсти. У песца ( Vulpes lagopus ) и горностая ( Mustela erminea ) мех меняется от белого и густого зимой до коричневого и редкого летом. У ластоногих и белых медведей толстый изолирующий слой жира помогает поддерживать температуру тела.
Половой диморфизм
[ редактировать ]

Отношения с людьми
[ редактировать ]Хищники, возможно, являются группой млекопитающих, представляющих наибольший интерес для человека. Собака примечательна не только тем , что она является первым одомашненным видом хищников, но и первым видом любого таксона. За последние 10 000–12 000 лет люди селекционно разводили собак для самых разных задач, и сегодня существует более 400 пород. Кошка — еще один одомашненный хищник, и сегодня она считается одним из самых успешных видов на планете из-за их непосредственной близости к человеку и популярности кошек в качестве домашних животных. Многие другие виды популярны и часто представляют собой харизматическую мегафауну . Многие цивилизации включили в свою культуру виды хищников: ярким примером является лев , который во многих обществах рассматривается как символ власти и королевской власти. Тем не менее, на многие виды, такие как волки и большие кошки, широко охотились, что привело к их истреблению в некоторых районах. Утрата среды обитания и вторжение человека, а также изменение климата стали основной причиной исчезновения многих видов. С 1600-х годов вымерли четыре вида хищников: Волк Фолклендских островов ( Dusicyon australis ) в 1876 г.; морская норка ( Neogale Macrodon ) в 1894 г.; японский морской лев ( Zalophus japonicus ) в 1951 году и карибский тюлень-монах ( Neomonachus тропический ) в 1952 году. [22] Некоторые виды, такие как рыжая лисица ( Vulpes vulpes ) и горностай ( Mustela erminea ), были завезены в Австралазию и привели к тому, что многие местные виды оказались под угрозой исчезновения или даже вымерли. [63]
См. также
[ редактировать ]- Классификация млекопитающих
- Карниворообразные
- Список хищников
- Список хищников по численности населения
Ссылки
[ редактировать ]- ^ Фоли, Нью-Мексико; Мейсон, ВК; Харрис, Эй Джей; Бредемейер, КР; Дамас, Дж.; Левин, ХА; Эйзирик, Э.; Гейтси, Дж.; Карлссон, ЕК; Линдблад-То, К.; Консорциум «Зоономия»; Спрингер, М.С.; Мерфи, WJ (2023). «Геномная временная шкала эволюции плацентарных млекопитающих» . Наука . 380 (6643). eabl8189. дои : 10.1126/science.abl8189 . ПМЦ 10233747 . ПМИД 37104581 .
- ^ Перейти обратно: а б Боудич, TE 1821. Анализ естественных классификаций млекопитающих для использования студентами и путешественниками Дж. Смитом в Париже. 115. (см. стр. 24, 33)
- ^ Загороднюк И. (2008) «Научные названия отрядов млекопитающих: от описательных к единообразным» Висник Львовского университета, Серия «Биология», Ис. 48. С. 33-43.
- ^ Загороднюк И. (2014) «Изменения в таксономическом разнообразии украинских млекопитающих за последние три столетия: вымершие, фантомные и чужеродные виды» Труды териологической школы, Vol. 12:3–16
- ^ Геккель, Эрнст (1866 г.) «Общая морфология организмов». Берлин: Георг Раймер.
- ^ Дж. Л. Вортман (1901). «Исследования эоценовых млекопитающих в коллекции Марша, Музей Пибоди». Американский журнал науки, серия 4 12: 193-206.
- ^ Kalandadze, N. N. and S. A. Rautian (1992.) "Systema mlekopitayushchikh i istorygeskaya zoogeographei [The system of mammals and historical zoogeography]." Sbornik Trudov Zoologicheskogo Muzeya Moskovskogo Goschdarstvennoro Universiteta 29:44–152.
- ^ Жорж Кювье, Пьер Андре Латрейль (1817 г.) «Царство животных, распределенное в соответствии с его организацией, чтобы служить основой для естественной истории животных и введения в сравнительную анатомию» Книготорговец Детервиля, Imprimerie de A. Belin, Париж, 4 тома
- ^ Феликс Вик-Дазир (1792 г.) «Методическая энциклопедия, Том 2: Анатомическая система, четвероногие» Панкук
- ^ Артур Сперри Пирс, (1936) «Зоологические названия. Список типов, классов и отрядов, подготовленный для раздела F, Американская ассоциация содействия развитию науки» Американская ассоциация развития науки
- ^ Г. Г. Симпсон (1952) «За и против единых окончаний в зоологической номенклатуре» в «Систематической зоологии, том 1, № 1 (весна, 1952 г.)», стр. 20–23, опубликовано: Taylor & Francisco, Ltd.
- ^ Кеннет Э. Кинман (1994.) «Система Кинмана: на пути к стабильной кладисто-эклектической классификации организмов: живые и вымершие, 48 типов, 269 классов, 1719 отрядов», Хейс, Канзас (почтовый ящик 1377, Hays 67601) , 88 страниц
- ^ O. S. Vyalov (1966.) "Sledy Zhiznedeyatel'nosti Organizmov i ikh Paleontologicheskoye Znacheniye [Traces of Vital Activity of Organisms and their Paleontological Significance]" Naukova Dumka, Kyiv, 1-219
- ^ Хаф, младший (1953). «Слуховая область североамериканских ископаемых кошачьих: ее значение в филогении». Профессиональные статьи Геологической службы США, 243-G, 95–115.
- ^ Рафинеск, Константин Самуэль (1815). «Анализ Природы или таблица Вселенной и организованных тел» . 1815 год . Палермо, За счет автора, 223 с.
{{cite journal}}: Для цитирования журнала требуется|journal=( помощь ) - ^ Иоганн Карл Вильгельм Иллигер (1811 г.) «Продром системы млекопитающих и птиц». Берлин: Сумптибус К. Зальфельд, XVIII, 301 страница.
- ^ Грей, Дж. Э. (1821). «О естественном расположении позвоночных животных» . Лондонский медицинский репозиторий . 15 (1): 296–310.
- ^ Мекаев, Ю.А. (2002.) «Фауногенез и классификация млекопитающих». Петрова Академия наук и искусств, Санкт-Петербург, 1–895.
- ^ Перейти обратно: а б Линней, К. (1758). Система природы через три царства Природы, по классам, отрядам, родам, видам, с разными признаками, синонимичными местами. Том I Прямые расходы. Лоуренс Сальвиус, Холмия. стр. 20–32.
- ^ Иоганн Фридрих Блюменбах (1791 г.) «Справочник по естественной истории. Четвертое издание». Геттинген, Иоганн Кристиан Дитрих, xii+704+[33] стр., 3 шт.
- ^ Леонард Радинский (1977) «Мозги ранних хищников». Палеобиология, Том 3, Выпуск 4, стр. 333–349.
- ^ Перейти обратно: а б Хантер, Л. (2018). Полевое руководство по хищникам мира . Дикая природа Блумсбери. стр. 1–271. ISBN 978-1472950796 .
- ^ Перейти обратно: а б Полли, Дэвид, Джина Д. Уэсли-Хант, Рональд Э. Генрих, Грэм Дэвис и Питер Худ (2006). «Самая ранняя известная слуховая булла хищных животных и подтверждение недавнего происхождения карниворов коронной клады (Eutheria, Mammalia)» . Палеонтология . 49 (5): 1019–1027. дои : 10.1111/j.1475-4983.2006.00586.x .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ Уодделл, Питер Дж.; Окада, Норихиро; Хасегава, Масами (1999). «На пути к разрешению межпорядковых отношений плацентарных млекопитающих» . Систематическая биология . 48 (1): 1–5. дои : 10.1093/sysbio/48.1.1 . ПМИД 12078634 .
- ^ Цагкогеорга, Г; Паркер, Дж; Ступка, Е; Коттон, Дж.А.; Росситер, С.Дж. (2013). «Филогеномный анализ проясняет эволюционные взаимоотношения летучих мышей» . Современная биология . 23 (22): 2262–2267. Бибкод : 2013CBio...23.2262T . дои : 10.1016/j.cub.2013.09.014 . ПМИД 24184098 .
- ^ Перейти обратно: а б Холлидей, Томас, доктор медицинских наук; Апчерч, Пол; Госвами, Анджали (2015). «Разрешение взаимоотношений плацентарных млекопитающих палеоцена» (PDF) . Биологические обзоры . 92 (1): 521–550. дои : 10.1111/brv.12242 . ISSN 1464-7931 . ПМК 6849585 . ПМИД 28075073 . Архивировано (PDF) из оригинала 9 октября 2022 года.
- ^ Маккенна, MC (1975). «К филогенетической классификации млекопитающих». В Лакетте, WP; Салай, Ф.С. (ред.). Филогения приматов . Нью-Йорк: Пленум. стр. 21–46.
- ^ Фельдхамер, Джордж А.; Дрикамер, Ли С.; Весси, Стивен Х.; Мерритт, Джозеф Ф.; Краевски, Кэри (2015). Маммология: адаптация, разнообразие, экология . Балтимор: Издательство Университета Джонса Хопкинса. п. 356 . ISBN 978-0801886959 .
- ^ Тернер, Алан; Антон, Маурисио (2004). Развивающийся Эдем: иллюстрированное руководство по эволюции фауны крупных млекопитающих Африки . Нью-Йорк: Издательство Колумбийского университета. п. 77. ИСБН 978-0-231-11944-3 .
- ^ Брайант, Х.Н. и М. Уолсон (2004) «Филогенетическая номенклатура хищных млекопитающих». Первое Международное совещание по филогенетической номенклатуре . Париж, Национальный музей естественной истории.
- ^ Джон Дж. Флинн; Джон А. Финарелли; Мишель Сполдинг (2010). «Филогения Carnivora и Carnivoramorpha, а также использование летописи окаменелостей для улучшения понимания эволюционных преобразований». В Анджали Госвами; Энтони Фриша (ред.). Эволюция хищников. Новые взгляды на филогению, форму и функцию . Издательство Кембриджского университета. стр. 25–63 . дои : 10.1017/CBO9781139193436.003 . ISBN 9781139193436 .
- ^ Генрих, RE; Пролив, Юг; Уд, П. (январь 2008 г.). «Самые ранние эоценовые Miacidae (Mammalia: Carnivora) из северо-западного Вайоминга». Журнал палеонтологии . 82 (1): 154–162. Бибкод : 2008JPal...82..154H . дои : 10.1666/05-118.1 . S2CID 35030667 .
- ^ Эйзирик, Э.; Мерфи, Вашингтон; Кепфли, КП; Джонсон, МЫ; Драгу, JW; О'Брайен, SJ (июль 2010 г.). «Схема и время диверсификации отряда млекопитающих Carnivora, выведенные на основе нескольких последовательностей ядерных генов» . Молекулярная филогенетика и эволюция . 56 (1): 49–63. дои : 10.1016/j.ympev.2010.01.033 . ПМК 7034395 . ПМИД 20138220 .
- ^ Ван, X.; Тедфорд, Р.Х. (2008). Собаки: их ископаемые родственники и история эволюции . Нью-Йорк: Издательство Колумбийского университета. стр. 1–232. ISBN 978-0-231-13529-0 .
- ^ РФ Эвер (1973). Хищники . Издательство Корнельского университета. ISBN 0-8014-8493-6 .
- ^ Перейти обратно: а б Симпсон, Г.Г. (1945). «Принципы классификации и классификация млекопитающих». Бюллетень АМНХ . 85 : 1–350. hdl : 2246/1104 .
- ^ Арнасон, Ю.; Гуллберг, А.; Янке, А.; Куллберг, М. (2007). «Митогеномный анализ собачьих взаимоотношений». Молекулярная филогенетика и эволюция . 45 (3): 863–74. дои : 10.1016/j.ympev.2007.06.019 . ПМИД 17919938 .
- ^ Ленто, генеральный директор; Хиксон, Р.Э.; Чемберс, ГК; Пенни, Д. (1995). «Использование спектрального анализа для проверки гипотез о происхождении ластоногих». Молекулярная биология и эволюция . 12 (1): 28–52. doi : 10.1093/oxfordjournals.molbev.a040189 . ПМИД 7877495 .
- ^ Хант, Р.М. младший; Барнс, Л.Г. (1994). «Базовые черепные доказательства родства древнейших ластоногих с урсидами» (PDF) . Труды Общества естественной истории Сан-Диего . 29 : 57–67.
- ^ Хигдон, JW; Бининда-Эмондс, Орегон; Бек, Р.М.; Фергюсон, SH (2007). «Филогения и дивергенция ластоногих (Carnivora: Mammalia) оценены с использованием набора мультигенных данных» . Эволюционная биология BMC . 7 (1): 216. Бибкод : 2007BMCEE...7..216H . дои : 10.1186/1471-2148-7-216 . ПМК 2245807 . ПМИД 17996107 .
- ^ Сато, Джей-Джей; Вулсан, М.; Сузуки, Х.; Хосода, Т.; Ямагути, Ю.; Хияма, К.; Кобаяши, М.; Минами, С. (2006). «Данные последовательностей ядерной ДНК проливают свет на филогенетические взаимоотношения ластоногих: единое происхождение со сходством с Musteloidea». Зоологическая наука . 23 (2): 125–46. дои : 10.2108/zsj.23.125 . hdl : 2115/13508 . ПМИД 16603806 . S2CID 25795496 .
- ^ Флинн, Джей-Джей; Финарелли, Дж.А.; Зер, С.; Сюй, Дж.; Недбал, Массачусетс (2005). «Молекулярная филогения хищных животных (млекопитающих): оценка влияния увеличения выборки на разрешение загадочных отношений» . Систематическая биология . 54 (2): 317–37. дои : 10.1080/10635150590923326 . ПМИД 16012099 .
- ^ Гобер, П.; Верон, Г. (2003). «Исчерпывающий набор выборок среди Viverridae показывает сестринскую группу кошачьих: линзанги как случай крайней морфологической конвергенции внутри Feliformia» . Труды Королевского общества B: Биологические науки . 270 (1532): 2523–2530. дои : 10.1098/rspb.2003.2521 . ПМК 1691530 . ПМИД 14667345 .
- ^ Энн Д. Йодер и Джон Дж. Флинн 2003: Происхождение малагасийского хищника
- ^ Йодер, А., М. Бернс, С. Зер, Т. Делефосс, Г. Верон, С. Гудман, Дж. Флинн. 2003: Единое происхождение малагасийских хищников от африканского предка – Письма к природе
- ^ Филипп Гобер, В. Крис Возенкрафт, Педро Кордейро-Эстрела и Жеральдин Верон. 2005 - Мозаика конвергенций и шума в морфологической филогении: что находится в виверридоподобном хищнике?
- ^ Эйзирик, Э.; Мерфи, Вашингтон; Кепфли, КП; Джонсон, МЫ; Драгу, JW; Уэйн, РК; О'Брайен, SJ (2010). «Схема и время диверсификации отряда млекопитающих Carnivora, выведенные на основе нескольких последовательностей ядерных генов» . Молекулярная филогенетика и эволюция . 56 (1): 49–63. дои : 10.1016/j.ympev.2010.01.033 . ПМК 7034395 . ПМИД 20138220 .
- ^ Гобер, П. (2009). «Семейство Prionodontidae (Linsangs)». В Уилсоне, Делавэр; Миттермайер, Р.А. (ред.). Справочник млекопитающих мира – Том 1 . Барселона: Lynx Ediciones. стр. 170–173. ISBN 978-84-96553-49-1 .
- ^ Салеза, М.; М. Антон; С. Пенье; Дж. Моралес (2006). «Наличие ложного большого пальца у ископаемого хищника проясняет эволюцию панд» . Труды Национальной академии наук . 103 (2): 379–382. Бибкод : 2006PNAS..103..379S . дои : 10.1073/pnas.0504899102 . ПМЦ 1326154 . ПМИД 16387860 .
- ^ Ю, Ли; Ли, И-Вэй; Райдер, Оливер А.; Чжан, Я-Пин (2007). «Анализ полных последовательностей митохондриального генома увеличивает филогенетическое разрешение медведей (Ursidae), семейства млекопитающих, которое пережило быстрое видообразование» . Эволюционная биология BMC . 7 (198): 198. Бибкод : 2007BMCEE...7..198Y . дои : 10.1186/1471-2148-7-198 . ПМК 2151078 . ПМИД 17956639 .
- ^ Краузе, Дж.; Унгер, Т.; Носон, А.; Маласпинас, А.; Колокотронис, С.; Стиллер, М.; Сойбельзон, Л.; Сприггс, Х.; Дорогой, ПХ; Бриггс, AW; Брей, SCE; О'Брайен, SJ; Рабедер, Г.; Матеус, П.; Купер, А.; Слаткин М.; Паабо, С.; Хофрейтер, М. (2008). «Митохондриальные геномы обнаруживают взрывную радиацию вымерших и современных медведей вблизи границы миоцена и плиоцена» . Эволюционная биология BMC . 8 (220): 220. Бибкод : 2008BMCEE...8..220K . дои : 10.1186/1471-2148-8-220 . ПМК 2518930 . ПМИД 18662376 .
- ^ Перейти обратно: а б Мехта, РС; Слейтер, Дж.Дж.; Закон, CJ (2018). «Разнообразие линий и неравенство размеров у Musteloidea: тестирование закономерностей адаптивной радиации с использованием молекулярных методов и методов, основанных на ископаемых» . Систематическая биология . 67 (1): 127–144. дои : 10.1093/sysbio/syx047 . ISSN 1063-5157 . ПМИД 28472434 .
- ^ Кепфли К.П., Дир К.А., Слейтер Г.Дж. и др. (2008). «Мультигенная филогения куньих: определение взаимоотношений, темпа и биогеографической истории адаптивной радиации млекопитающих» . БМК Биол . 6 : 4–5. дои : 10.1186/1741-7007-6-10 . ПМК 2276185 . ПМИД 18275614 .
- ^ Уилсон, Делавэр; Миттермайер, Р.А., ред. (2009). Справочник млекопитающих мира – Том 1 . Барселона: Lynx Ediciones. стр. 1–728. ISBN 978-84-96553-49-1 .
- ^ Уилсон, Делавэр; Миттермайер, Р.А., ред. (2014). Справочник млекопитающих мира – Том 4 . Барселона: Lynx Ediciones. стр. 1–614. ISBN 978-84-96553-93-4 .
- ^ Перейти обратно: а б с д Новак, Р.М. (2005). «Хищники мира» Уокера . Балтимор, Мэриленд: Издательство Университета Джонса Хопкинса. стр. 1 –328. ISBN 0801880335 .
- ^ Перейти обратно: а б с д Воган, штат Техас; Райан, Дж. М.; Чаплевски, Нью-Джерси (2013). Маммология . Берлингтон, Массачусетс: Jones & Bartlett Learning. стр. 1–750. ISBN 9781284032093 .
- ^ Кнобил, Эрнст (2006). Физиология репродукции Кнобила и Нила . Профессиональное издательство Персидского залива. ISBN 978-0-12-515401-7 .
- ^ Шульц, Николас Г.; Лох-Стивенс, Майкл; Абреу, Эрик; Орр, Тери; Дин, Мэтью Д. (1 июня 2016 г.). «Бакулум неоднократно приобретался и терялся в ходе эволюции млекопитающих» . Интегративная и сравнительная биология . 56 (4): 644–56. дои : 10.1093/icb/icw034 . ISSN 1540-7063 . ПМК 6080509 . ПМИД 27252214 .
- ^ Энциклопедия поведения животных . Том. 2. Академическая пресса. 21 января 2019 г. с. 7. ISBN 978-0-12-813252-4 .
- ^ Гиттлман, Дж.Л.; Валкенбург, Б. Ван (май 1997 г.). «Половой диморфизм клыков и черепов хищных животных: влияние размера, филогении и экологии поведения» . Журнал зоологии . 242 (1): 97–117. дои : 10.1111/j.1469-7998.1997.tb02932.x . ISSN 0952-8369 .
- ^ Сильвия Бруннер, Майкл М. Брайден, Питер Д. Шонесси (сентябрь 2004 г.). «Краниальный онтогенез отариидных тюленей» . Систематика и биоразнообразие . 2 (1): 83–110. Бибкод : 2004SyBio...2...83B . дои : 10.1017/S1477200004001367 . S2CID 83737300 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ «100 наихудших инвазивных видов в мире» . Группа специалистов по инвазивным видам.