Криль
| Криль | |
|---|---|

| |
| Северный криль ( Meganyctiphanes norvegica ) | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Членистоногие |
| Сорт: | Малакострака |
| Суперзаказ: | Эукарида |
| Заказ: | Евфаузиацея День , 1852 год. |
| Семьи и роды | |
| |
Криль (Еуфаузииды) , [ 1 ] ( серг.: криль) — мелкие и исключительно морские Euphausiacea , ракообразные отряда встречающиеся во всех мировых океанах. [ 2 ] Название «криль» происходит от норвежского слова krill , что означает «мелкая рыбка ». [ 3 ] что также часто относят к видам рыб.
Криль считается важным связующим звеном на трофическом уровне — в самом низу пищевой цепи . Они питаются фитопланктоном и, в меньшей степени, зоопланктоном , а также являются основным источником пищи для многих крупных животных. В Южном океане одного вида, антарктического криля , составляет биомасса около 379 миллионов тонн . [ 4 ] что делает его одним из видов с наибольшей общей биомассой. более половины этой биомассы съедают киты, тюлени , пингвины, морские птицы, кальмары Ежегодно и рыбы. Большинство видов криля совершают большие ежедневные вертикальные миграции , обеспечивая тем самым пищу хищникам у поверхности ночью и в более глубоких водах в течение дня.
Криль коммерчески вылавливается в Южном океане и в водах вокруг Японии. Общий мировой вылов составляет 150 000–200 000 тонн в год, большая часть которого добывается в море Скотия . Большая часть улова криля используется для аквакультуры и корма для аквариумов , в качестве наживки в спортивной рыбалке или в фармацевтической промышленности. В Японии, на Филиппинах и в России криль также используется в пищу человеком и известен в Японии как окиами ( オキアミ ) . Их едят как камарон в Испании и на Филиппинах. На Филиппинах криль также известен как аламан и используется для приготовления соленой пасты под названием багунг .
Криль также является основной добычей усатых китов , в том числе синего кита .
Таксономия
[ редактировать ]крупных членистоногих Криль принадлежит к подтипу , ракообразным . Самая известная и крупнейшая группа ракообразных, класс Malacostraca , включает надотряд Eucarida , включающий три отряда: Euphausiacea (криль), Decapoda (креветки, креветки, омары, крабы) и планктонные Amphionidacea .
Отряд Euphausiacea включает два семейства . Более распространенный Euphausiidae содержит 10 различных родов и в общей сложности 85 видов. Из них род Euphausia является самым крупным, насчитывающим 31 вид. [ 5 ] Менее известное семейство Bentheuphausiidae имеет только один вид — Bentheuphausia amblyops , батипелагический криль, обитающий в глубоких водах ниже 1000 м (3300 футов). Считается самым примитивным из существующих видов криля. [ 6 ]
Хорошо известные виды Euphausiidae, используемые в коммерческом промысле криля , включают антарктический криль ( Euphausia superba ), тихоокеанский криль ( E. pacifica ) и северный криль ( Meganyctiphanes norvegica ). [ 7 ]
Филогения
[ редактировать ]| Предлагаемая филогения Euphausiacea [ 8 ] | |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| Филогения получена на основе морфологических данных, (♠) придуманных названий, [ 8 ] (♣) возможно, парафилетический таксон Nematobrachion in. [ 8 ] (♦) клады отличаются от Казановы (1984), [ 9 ] где Псевдоеуфаузия — сестра Никтифана , Евфаузия — сестра Тизаноподы , а Нематобрахион — сестра Стилохейрона . |
По состоянию на 2013 год [update]Отряд Euphausiacea считается монофилетическим из-за нескольких уникальных консервативных морфологических характеристик ( аутапоморфии ), таких как голые нитевидные жабры и тонкие торакоподы. [ 10 ] и молекулярными исследованиями. [ 11 ] [ 12 ] [ 13 ]
Было много теорий о местонахождении отряда Euphausiacea. С момента первого описания Thysanopode tricuspide Анри Милном-Эдвардсом в 1830 году сходство их двуветвистых торакопод побудило зоологов сгруппировать эвфаузиид и Mysidacea в отряд Schizopoda , который был разделен Йоханом Эриком Вести Боасом в 1883 году на два отдельных отряда. [ 14 ] Позже Уильям Томас Калман (1904) причислил мизидовых к надотряду Peracarida , а эвфаузиид к надотряду Eucarida , хотя даже до 1930-х годов отстаивался отряд Schizopoda. [ 10 ] Позже было также предложено сгруппировать отряд Euphausiacea с Penaeidae (семейство креветок) в Decapoda на основании сходства в развитии, как отметили Роберт Герни и Изабелла Гордон . [ 15 ] [ 16 ] Причина этих споров в том, что криль имеет некоторые морфологические особенности десятиногих и других мизид. [ 10 ]
Молекулярные исследования не смогли однозначно сгруппировать их, возможно, из-за нехватки ключевых редких видов, таких как Bentheuphausia amblyops у криля и Amphionides reynaudii у Eucarida. Одно исследование подтверждает монофилию Eucarida (с базальной Mysida), [ 17 ] другой группирует Euphausiacea с Mysida (Schizopoda), [ 12 ] в то время как еще один группирует Euphausiacea с Hoplocarida . [ 18 ]
Хронология
[ редактировать ]Ни одна из сохранившихся окаменелостей не может быть однозначно отнесена к Euphausiacea. некоторые вымершие эумалакостраков таксоны Считалось, что относятся к эвфаузиацеям, таким как Anthracophausia , Crangopsis , которые теперь отнесены к Aeschronectida (Hoplocarida). [ 8 ] — и Палеомисис . [ 19 ] Все датировки событий видообразования были оценены с помощью методов молекулярных часов , которые установили, что последний общий предок криля семейства Euphausiidae (отряд Euphausiacea минус Bentheuphausia amblyops ) жил в нижнем меловом периоде около 130 миллионов лет назад . [ 12 ]
Распределение
[ редактировать ]Криль встречается во всем мире во всех океанах, хотя многие отдельные виды имеют эндемичное или неритическое ( т. е. прибрежное) распространение. Bentheuphausia amblyops , батипелагический вид, имеет космополитическое распространение в пределах своей глубоководной среды обитания. [ 20 ]
Виды рода Thysanoessa встречаются как в Атлантическом , так и в Тихом океане. [ 21 ] Тихий океан является домом для Euphausia pacifica . Северный криль встречается через Атлантику от Средиземного моря на север.
Виды с неритическим распространением включают четыре вида рода Nyctiphanes . [ 22 ] Они очень многочисленны в апвеллинга Калифорнии районах , Гумбольдта , Бенгелы и Канарских островов систем течений . [ 23 ] [ 24 ] [ 25 ] Другой вид, имеющий только неритическое распространение, — это E. Crystallorophias , эндемичный для побережья Антарктики. [ 26 ]
Виды с эндемичным распространением включают Nyctiphanes capensis , который встречается только в течении Бенгелы, [ 22 ] E. mucronata в течении Гумбольдта, [ 27 ] и шесть видов Euphausia, произрастающих в Южном океане.
В Антарктике известно семь видов, [ 28 ] один из рода Thysanoessa ( T. macrura ) и шесть из Euphausia . Антарктический криль ( Euphausia superba ) обычно обитает на глубинах, достигающих 100 м (330 футов). [ 29 ] тогда как ледяной криль ( Euphausia Crystallorophias ) достигает глубины 4000 м (13 100 футов), хотя обычно они обитают на глубине не более 300–600 м (1 000–2 000 футов). [ 30 ] Криль совершает вертикальные миграции (DVM) большими стаями, и акустические данные показали, что эти миграции достигают глубины до 400 метров. [ 31 ] Оба встречаются на широте южнее 55° ю.ш. , при этом E. Crystallorophias преобладает к югу от 74° ю.ш. [ 32 ] и в районах паковых льдов . Другие виды, известные в Южном океане, — E. frigida , E. longirostris , E. triacantha и E. vallentini . [ 33 ]
Анатомия и морфология
[ редактировать ]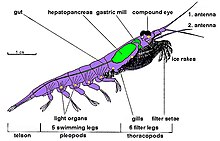

Криль относится к ракообразным и, как и все ракообразные, имеет хитиновый экзоскелет . Их анатомия аналогична стандартному десятиногому , их тела состоят из трех частей : головогрудь состоит из головы и грудной клетки , которые срослись, и брюшка , несущего десять плавательных придатков, и хвостового веера . Эта внешняя оболочка криля у большинства видов прозрачна.
У криля сложные сложные глаза . Некоторые виды адаптируются к различным условиям освещения за счет использования экранирующих пигментов . [ 34 ]
У них есть две усики и несколько пар грудных ног, называемых переоподами или торакоподами , названными так потому, что они прикреплены к грудной клетке. Их количество варьируется в зависимости от рода и вида. К этим грудным ногам относятся ноги для кормления и ноги для ухода.
Криль, вероятно, является родственной кладой десятиногих, потому что все виды имеют пять пар плавательных ног, которые вместе с последними называются «пловцами», очень похожими на ноги лобстера или пресноводного рака .
Несмотря на наличие десяти плавников, также известных как плеоподы , криль нельзя считать десятиногими. У них нет настоящих наземных ног, поскольку все их переоподы превращены в ноги для ухода и вспомогательные кормления. У Decapoda имеется десять функционирующих переоподов , что и дало им название; тогда как здесь не осталось локомотивных переоподов . И при этом вообще не существует постоянно десяти переоподов .
Длина большинства взрослых особей криля составляет около 1–2 см (0,4–0,8 дюйма). Некоторые виды вырастают до размеров порядка 6–15 сантиметров (2,4–5,9 дюйма). Самый крупный вид криля, Thysanopoda spinicaudata , обитает глубоко в открытом океане . [ 35 ] Криль можно легко отличить от других ракообразных, таких как настоящие креветки, по видимым снаружи жабрам . [ 36 ]
За исключением Bentheuphausia amblyops , криль — биолюминесцентные животные, имеющие органы, называемые фотофорами , которые могут излучать свет. Свет генерируется в результате ферментативно -катализируемой реакции хемилюминесценции , в которой люциферин (разновидность пигмента) активируется ферментом люциферазой . Исследования показывают, что люциферин многих видов криля представляет собой флуоресцентный тетрапиррол, похожий, но не идентичный динофлагеллят . люциферину [ 37 ] и что криль, вероятно, не производит это вещество сам, а приобретает его как часть своего рациона, содержащего динофлагелляты. [ 38 ] Фотофоры криля представляют собой сложные органы с линзами и способностями к фокусировке, которые можно вращать с помощью мышц. [ 39 ] Точная функция этих органов пока неизвестна; Возможности включают спаривание, социальное взаимодействие или ориентацию, а также форму камуфляжа против освещения для компенсации их тени от верхнего окружающего света. [ 40 ] [ 41 ]
Экология
[ редактировать ]
Кормление
[ редактировать ]Многие крили являются фильтраторами : [ 24 ] их передние придатки , торакоподы, образуют очень тонкие гребни, с помощью которых они могут отфильтровывать пищу из воды. Эти фильтры могут быть очень тонкими у видов (таких как Euphausia spp.), которые питаются в основном фитопланктоном , в частности, диатомовыми водорослями , которые являются одноклеточными водорослями . Криль в большинстве своем всеяден , [ 43 ] хотя некоторые виды плотоядны и питаются мелким зоопланктоном и личинками рыб . [ 44 ]
Криль является важным элементом водной пищевой цепи . Криль преобразует первичную продукцию своей добычи в форму, пригодную для потребления более крупными животными, которые не могут питаться непосредственно мельчайшими водорослями. Северный криль и некоторые другие виды имеют относительно небольшую фильтрующую корзину и активно охотятся на копепод и более крупный зоопланктон. [ 44 ]
Хищничество
[ редактировать ]Крилем питаются многие животные: от мелких животных, таких как рыбы или пингвины, до более крупных, таких как тюлени и усатые киты . [ 45 ]
Нарушения экосистемы , приводящие к сокращению популяции криля, могут иметь далеко идущие последствия. Во время цветения кокколитофор в Беринговом море в 1998 г. [ 46 ] например, концентрация диатомей упала в пострадавшем районе. Криль не может питаться более мелкими кокколитофорами, и, следовательно, популяция криля (в основном E. pacifica ) в этом регионе резко сократилась. Это, в свою очередь, повлияло на другие виды: популяция буревестников сократилась. Считалось, что этот инцидент стал одной из причин, по которой лосось не нерестился в тот сезон. [ 47 ]
Несколько одноклеточных эндопаразитоидных инфузорий рода Collinia могут заражать виды криля и опустошать пораженные популяции. Такие заболевания были зарегистрированы для Thysanoessa inermis в Беринговом море, а также для E. pacifica , Thysanoessa spinifera и T. gregaria у тихоокеанского побережья Северной Америки. [ 48 ] [ 49 ] Некоторые эктопаразиты семейства Dajidae (эпикаридовые изоподы ) поражают криль (а также креветок и мизид ); Одним из таких паразитов является Oculophryxus bicaulis , который был обнаружен на криле Stylocheron affine и S. longicorne . Он прикрепляется к глазному стеблю животного и высасывает кровь из его головы; очевидно, он подавляет размножение хозяина, поскольку ни одно из больных животных не достигло зрелости. [ 50 ]
Изменение климата представляет собой еще одну угрозу для популяций криля. [ 51 ]
Пластмассы
[ редактировать ]Предварительные исследования показывают, что криль может переваривать микропластик диаметром менее 5 мм (0,20 дюйма), расщепляя его и выводя обратно в окружающую среду в более мелкой форме. [ 52 ]
История жизни и поведение
[ редактировать ]
Жизненный цикл криля относительно хорошо изучен, несмотря на незначительные различия в деталях от вида к виду. [ 15 ] [ 24 ] После вылупления криля они проходят несколько личиночных стадий — науплиус , псевдометанауплиус , метанауплиус , калиптопсис и фурцилия , каждая из которых делится на подстадии. Стадия псевдометанауплиуса присуща только видам, которые откладывают яйца в яйцевидный мешок: так называемые «мякоть-нерестовики». Личинки растут и линяют неоднократно по мере развития, заменяя свой жесткий экзоскелет, когда он становится слишком маленьким. Мелкие животные линяют чаще, чем крупные. Запасы желтка в их организме питают личинок на стадии метанауплиуса.
На стадии калиптопсиса дифференцировка зашла достаточно далеко, чтобы у них появились рот и пищеварительный тракт, и они начали питаться фитопланктоном. К этому времени их запасы желтка исчерпаны, и личинки должны достичь фотической зоны — верхних слоев океана, где процветают водоросли. На стадиях фурцилий добавляются сегменты с парами плавников, начиная с самых передних сегментов. Каждая новая пара становится функциональной только при следующей линьке. Количество сегментов, добавляемых на любой из стадий фурцилий, может варьировать даже в пределах одного вида в зависимости от условий окружающей среды. [ 53 ] После последней стадии фурцилии незрелая молодь появляется в форме, похожей на взрослую особь, у которой впоследствии развиваются гонады и происходит половое созревание. [ 54 ]
Воспроизведение
[ редактировать ]
Во время брачного сезона, который зависит от вида и климата, самец откладывает сперматозоид в половое отверстие самки (так называемый теликум ). Самки могут нести в яичниках несколько тысяч яиц , что может составлять до трети массы тела животного. [ 55 ] Криль может иметь несколько выводков за один сезон с интервалами между выводками порядка нескольких дней. [ 25 ] [ 56 ]
Криль использует два типа механизма нереста. [ 25 ] 57 видов родов Bentheuphausia , Euphausia , Meganyctiphanes , Thysanoessa и Thysanopoda являются «широковещательными производителями»: самка выпускает оплодотворенную икру в воду, где они обычно тонут, рассеиваются и находятся самостоятельно. Эти виды обычно вылупляются на стадии науплиуса 1, но недавно было обнаружено, что они иногда вылупляются на стадии метанауплиуса или даже на стадии калиптописа. [ 57 ] Остальные 29 видов других родов являются «производителями мешков», когда самка несет с собой икру, прикрепленную к самым задним парам торакопод, пока они не вылупляются как метанауплии, хотя некоторые виды, такие как Nematoscelis difficilis, могут вылупляться как науплиусы или псевдометанауплиусы. [ 58 ]
линька
[ редактировать ]Линька происходит всякий раз, когда особь перерастает свой жесткий экзоскелет. Молодые животные, растущие быстрее, линяют чаще, чем старшие и крупные. Частота линьки широко варьируется в зависимости от вида и даже в пределах одного вида зависит от многих внешних факторов, таких как широта, температура воды и наличие пищи. Например, у субтропического вида Nyctiphanes simplex общий период между линькой составляет от двух до семи дней: личинки линяют в среднем каждые четыре дня, а молодые и взрослые особи - в среднем каждые шесть дней. Для E. superba в антарктическом море наблюдались периоды между линькой от 9 до 28 дней в зависимости от температуры от -1 до 4 °C (от 30 до 39 °F), а у Meganyctiphanes norvegica в море Северном периоды между линьками также варьируются от 9 до 28 дней, но при температуре от 2,5 до 15 ° C (от 36,5 до 59,0 ° F). [ 59 ] E. superba способен уменьшать размер своего тела, когда не хватает еды, а также линяет, когда его экзоскелет становится слишком большим. [ 60 ] Аналогичное сокращение наблюдалось и у E. pacifica , вида, обитающего в Тихом океане от полярных до умеренных зон, как адаптация к аномально высоким температурам воды. Предполагается, что сокращение численности произойдет и для других видов криля умеренной зоны. [ 61 ]
Продолжительность жизни
[ редактировать ]Некоторые высокоширотные виды криля могут жить более шести лет (например, Euphausia superba ); другие, такие как вид средних широт Euphausia pacifica , живут всего два года. [ 7 ] Продолжительность жизни субтропических или тропических видов еще короче, например Nyctiphanes simplex , который обычно живет всего шесть-восемь месяцев. [ 62 ]
Роение
[ редактировать ]
Большинство криля — роящиеся животные; размеры и плотность таких стай различаются в зависимости от вида и региона. У Euphausia superba численность стаи достигает от 10 000 до 60 000 особей на кубический метр. [ 63 ] [ 64 ] Роение — это защитный механизм, сбивающий с толку более мелких хищников, которые хотели бы выбрать особей. В 2012 году Гандоми и Алави представили, по всей видимости, успешный стохастический алгоритм для моделирования поведения стаев криля. Алгоритм основан на трех основных факторах: «(i) движение, вызванное присутствием других особей, (ii) деятельность по поиску пищи и (iii) случайное распространение». [ 65 ]
Вертикальная миграция
[ редактировать ]
Криль обычно совершает суточную вертикальную миграцию . Предполагалось, что день они проводят на большей глубине, а ночью поднимаются к поверхности. Чем глубже они заходят, тем больше они снижают свою активность, [ 66 ] очевидно, чтобы уменьшить количество встреч с хищниками и сохранить энергию. Плавательная активность криля зависит от наполненности желудка. Сытые животные, питавшиеся у поверхности, плавают менее активно и поэтому опускаются ниже перемешанного слоя. [ 67 ] По мере погружения они производят фекалии , которые играют роль в антарктическом углеродном цикле . Криль с пустым желудком плавает активнее и направляется к поверхности.
Вертикальная миграция может происходить 2–3 раза в день. Некоторые виды (например, Euphausia superba , E. pacifica , E. hanseni , Pseudeuphausia latifrons и Thysanoessa spinifera ) в течение дня образуют наземные стаи для кормления и размножения, хотя такое поведение опасно, поскольку делает их чрезвычайно уязвимыми для хищников. [ 68 ]
Экспериментальные исследования с использованием Artemia salina в качестве модели показывают, что вертикальные миграции криля на несколько сотен метров, группами глубиной в десятки метров, могут в совокупности создавать достаточное количество нисходящих струй воды, чтобы оказать существенное влияние на перемешивание океана. [ 69 ]
Плотные стаи могут спровоцировать безумную жажду кормления среди рыб, птиц и хищников-млекопитающих, особенно у поверхности. Если их потревожить, рой разбегается, а некоторые особи даже мгновенно линяют, оставляя экзувию в качестве приманки. [ 70 ]
Криль обычно плавает со скоростью 5–10 см/с (2–3 длины тела в секунду). [ 71 ] используя свои плавки для движения. Их более крупные миграции зависят от океанских течений. В случае опасности они демонстрируют реакцию бегства , называемую омаром — щелкая хвостовыми структурами, тельсонами и уроподами , они относительно быстро движутся назад по воде, достигая скорости в диапазоне от 10 до 27 длин тела в секунду, что для крупного криля например, E. superba, означает около 0,8 м/с (3 фута/с). [ 72 ] Их способность плавать побудила многих исследователей классифицировать взрослый криль как микронектонные формы жизни, т.е. мелких животных, способных индивидуально двигаться против (слабых) течений. Личиночные формы криля обычно считаются зоопланктоном. [ 73 ]
Биогеохимические циклы
[ редактировать ]
Антарктический криль является важным видом в контексте биогеохимического круговорота. [ 74 ] [ 42 ] и в пищевой сети Антарктики . [ 75 ] [ 76 ] Он играет важную роль в Южном океане из-за своей способности осуществлять круговорот питательных веществ и служить кормом для пингвинов, усатых и синих китов .

Человеческое использование
[ редактировать ]
История сбора урожая
[ редактировать ]Криль собирали в качестве источника пищи для людей и домашних животных, по крайней мере, с 19 века, а возможно, и раньше, в Японии, где он был известен как окиами . Крупномасштабный промысел развился в конце 1960-х — начале 1970-х годов и сейчас ведется только в антарктических водах и морях вокруг Японии. Исторически крупнейшими странами, занимающимися промыслом криля, были Япония и Советский Союз, а после распада последнего — Россия и Украина . [ 77 ] Пик вылова, который в 1983 году составил около 528 000 тонн только в Южном океане (из которых Советский Союз забрал 93%), теперь регулируется в качестве меры предосторожности против чрезмерного вылова рыбы. [ 78 ]
В 1993 году два события привели к сокращению вылова криля: Россия вышла из отрасли; а Конвенция о сохранении морских живых ресурсов Антарктики (АНТКОМ) определила максимальные квоты на вылов для устойчивой добычи антарктического криля. После пересмотра в октябре 2011 года Комиссия решила не менять квоту. [ 79 ]
Ежегодный вылов в Антарктике стабилизировался на уровне около 100 000 тонн, что составляет примерно одну пятидесятую квоты АНТКОМа на вылов. [ 80 ] Главным сдерживающим фактором, вероятно, были высокие затраты, а также политические и юридические проблемы. [ 81 ] Японский промысел насыщен примерно на 70 000 тонн. [ 82 ]
Хотя криль встречается по всему миру, предпочтительнее ловить рыбу в Южных океанах, поскольку в этих регионах криль более «уловист» и его больше. В частности, в антарктических морях, которые считаются нетронутыми , они считаются «чистым продуктом». [ 77 ]
В 2018 году было объявлено, что почти все компании по добыче криля, работающие в Антарктиде, с 2020 года откажутся от деятельности на огромных территориях вокруг Антарктического полуострова, включая «буферные зоны» вокруг гнездовых колоний пингвинов. [ 83 ]
Потребление человека
[ редактировать ]
Хотя общая биомасса антарктического криля может достигать 400 миллионов тонн, антропогенное воздействие на этот ключевой вид растет: за 2010–2014 годы общий вылов рыбы увеличился на 39% до 294 000 тонн. [ 80 ] Основными странами, участвующими в добыче криля, являются Норвегия (56% общего вылова в 2014 г.), Республика Корея (19%) и Китай (18%). [ 80 ]
Криль является богатым источником белка и жирных кислот омега-3 , которые в начале 21 века разрабатываются в качестве продуктов питания для человека, пищевых добавок в виде масляных капсул, корма для скота и корма для домашних животных . [ 77 ] [ 79 ] [ 84 ] Криль имеет соленый вкус и несколько более сильный рыбный вкус, чем креветки. Продукты массового потребления и продукты промышленного производства необходимо очистить от кожуры, чтобы удалить несъедобный экзоскелет . [ 84 ]
США В 2011 году Управление по санитарному надзору за качеством пищевых продуктов и медикаментов опубликовало письмо об отсутствии возражений по поводу признания произведенного из масла криля продукта безопасным (GRAS) для потребления человеком. [ 85 ]
Криль (и другие планктонные креветки , особенно виды Acetes ) наиболее широко потребляются в Юго-Восточной Азии, где его ферментируют (с неповрежденными панцирями) и обычно мелко измельчают для приготовления креветочной пасты . Его можно жарить и есть в сочетании с белым рисом или использовать для придания аромата умами широкому спектру традиционных блюд. [ 86 ] [ 87 ] Жидкость, образующуюся в процессе ферментации, также используется в качестве рыбного соуса . [ 88 ]
Биологическая робототехника
[ редактировать ]Криль — ловкие пловцы в промежуточном режиме чисел Рейнольдса , в котором не так много решений для беспилотной подводной робототехники, и он вдохновил роботизированные платформы как на изучение их передвижения, так и на поиск конструктивных решений для подводных роботов. [ 89 ]
См. также
[ редактировать ]- Антарктический криль
- Холодноводные креветки
- ракообразное
- Промысел криля
- Крилевое масло
- Северный криль
Ссылки
[ редактировать ]- ^ «Эуфаузииды (Криль)» . Правительство Канады . Рыбное хозяйство и океаны Канады. 6 апреля 2022 г. Проверено 18 апреля 2024 г.
Множество различных видов эвфаузиид обитают на восточном и западном побережьях Канады.
- ^ Ракообразные: Euphausiacea - Oxford Academic
- ^ «Криль» . Интернет-словарь этимологии . Проверено 22 июня 2010 г.
- ^ А. Аткинсон; В. Сигел; Е.А. Пахомов; М.Дж. Джессопп; В. Леб (2009). «Переоценка общей биомассы и годового производства антарктического криля» (PDF) . Глубоководные исследования . Часть I. 56 (5): 727–740. Бибкод : 2009DSRI...56..727A . дои : 10.1016/j.dsr.2008.12.007 .
- ^ Сигел В. (2011). Сигел В. (ред.). «Euphausiidae Dana, 1852» . Мировая база данных Euphausiacea . Всемирный реестр морских видов . Проверено 25 ноября 2011 г.
- ^ Э. Бринтон (1962). «Распространение тихоокеанских эвфаузиид» . Бык. Скриппс Инст. Океаногр . 8 (2): 51–270.
- ^ Перейти обратно: а б С. Николь; Ю. Эндо (1999). «Промысел криля: развитие, управление и последствия для экосистемы» . Водные живые ресурсы . 12 (2): 105–120. дои : 10.1016/S0990-7440(99)80020-5 . S2CID 84158071 .
- ^ Перейти обратно: а б с д Андреас Маас; Дитер Валошек (2001). «Развитие личинок Euphausia superba Dana, 1852 и филогенетический анализ Euphausiacea» (PDF) . Гидробиология . 448 : 143–169. дои : 10.1023/А:1017549321961 . S2CID 32997380 . Архивировано из оригинала (PDF) 18 июля 2011 года.
- ^ Бернадетт Казанова (1984). «Филогения Euphausiaceae (Crustacea: Eucarida)». Бюллетень Национального музея естественной истории (на французском языке). 4 : 1077–1089.
- ^ Перейти обратно: а б с Бернадетт Казанова (2003). «Орден Евфаузиацеи Дана, 1852 г.». Ракообразное . 76 (9): 1083–1121. дои : 10.1163/156854003322753439 . JSTOR 20105650 .
- ^ М. Евгения Д'Амато; Гордон В. Харкинс; Тулио де Оливейра; Питер Р. Теске; Марк Дж. Гиббонс (2008). «Молекулярное датирование и биогеография неритического криля Nyctiphanes » (PDF) . Морская биология . 155 (2): 243–247. дои : 10.1007/s00227-008-1005-0 . S2CID 17750015 . Архивировано из оригинала (PDF) 17 марта 2012 года . Проверено 4 июля 2010 г.
- ^ Перейти обратно: а б с Саймон Н. Джарман (2001). «Эволюционная история криля, выведенная на основе анализа последовательности ядерной большой субъединицы рДНК» . Биологический журнал Линнеевского общества . 73 (2): 199–212. дои : 10.1111/j.1095-8312.2001.tb01357.x .
- ^ Синь Шен; Хайцин Ван; Минсяо Ван; Бинь Лю (2011). «Полная последовательность митохондриального генома Euphausia pacifica (Malacostraca: Euphausiacea) обнаруживает новый порядок генов и необычные тандемные повторы». Геном . 54 (11): 911–922. дои : 10.1139/g11-053 . ПМИД 22017501 .
- ^ Йохан Эрик Вести Боас (1883). «Исследования взаимоотношений Малакостраки». Морфологический ежегодник (на немецком языке). 8 : 485-579.
- ^ Перейти обратно: а б Роберт Герни (1942). Личинки десятиногих ракообразных (PDF) . Рэйское общество .
- ^ Изабелла Гордон (1955). «Систематическое положение эвфаузиацеи» . Природа . 176 (4489): 934. Бибкод : 1955Natur.176..934G . дои : 10.1038/176934a0 . S2CID 4225121 .
- ^ Триша Спирс, Рональд В. ДеБри, Лоуренс Г. Абеле и Катажина Чодил (2005). Бойко, Кристофер Б. (ред.). «Монофилия перакарид и межпорядковая филогения, выведенная из последовательностей ядерной рибосомальной ДНК малых субъединиц (Crustacea: Malacostraca: Peracarida)» (PDF) . Труды Биологического общества Вашингтона . 118 (1): 117–157. doi : 10.2988/0006-324X(2005)118[117:PMAIPI]2.0.CO;2 . S2CID 85557065 .
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ) - ^ К. Меланд; Э. Уиллассен (2007). «Разобщенность «Мизидацеи» (Рачка)» Молекулярная филогенетика и эволюция . 44 (3): 1083–1104. CiteSeerX 10.1.1.653.5935 . дои : 10.1016/j.ympev.2007.02.009 . ПМИД 17398121 .
- ^ Фредерик Р. Шрам (1986). Ракообразные . Издательство Оксфордского университета . ISBN 978-0-19-503742-5 .
- ^ Джей Джей Торрес; Джей Джей Чилдресс (1985). «Дыхание и химический состав батипелагических эвфаузиид Bentheuphausia amblyops ». Морская биология . 87 (3): 267–272. дои : 10.1007/BF00397804 . S2CID 84486097 .
- ^ Волкер Сигел (2011). « Тисаноэсса Брандт, 1851 год» . ВОРМС . Всемирный реестр морских видов . Проверено 18 июня 2011 г.
- ^ Перейти обратно: а б Д'Амато, МЭ и др. : « Молекулярное датирование и биогеография неритического криля Nyctiphanes. Архивировано 17 марта 2012 г. в Wayback Machine », в журнале Marine Biology vol. 155, нет. 2 , стр. 243–247, август 2008 г.
- ^ Волкер Сигел (2011). В. Сигел (ред.). « Никтифан Сарс, 1883» . Мировая база данных Euphausiacea . Всемирный реестр морских видов . Проверено 18 июня 2011 г.
- ^ Перейти обратно: а б с Дж. Моклин; Л. Р. Фишер (1969). Биология Евфаузиид . Достижения морской биологии. Том. 7. Академическая пресса . ISBN 978-7-7708-3615-2 .
- ^ Перейти обратно: а б с Хайме Гомес-Гутьеррес; Карлос Дж. Робинсон (2005). нерестящихся в мешочках «Эмбриональное, раннее время развития личинок, механизм вылупления и межвыводковый период эвфаузиид Nyctiphanes simplex Hansen», . Журнал исследований планктона . 27 (3): 279–295. дои : 10.1093/plankt/fbi003 .
- ^ С.Н. Джарман; Н.Г. Эллиотт; С. Николь; А. МакМинн (2002). «Генетическая дифференциация антарктического прибрежного криля Euphausia Crystallorophias » . Наследственность . 88 (4): 280–287. дои : 10.1038/sj.hdy.6800041 . ПМИД 11920136 .
- ^ Р. Писец; В. Марин; Дж. Иррибаррен (2000). «Распространение Euphausia mucronata в зоне апвеллинга полуострова Медхильонес, север Чили: влияние слоя минимума кислорода» . Морская наука . 64 (1): 69–77. дои : 10.3989/scimar.2000.64n169 .
- ^ П. Брюггеман. « Эуфаузия кристаллорофиас » . Подводный путеводитель по острову Росс и проливу Мак-Мердо, Антарктида . Калифорнийский университет, Сан-Диего .
- ^ «Криль, Евфаузия суперба » . MarineBio.org . Проверено 25 февраля 2009 г.
- ^ Дж. А. Кирквуд (1984). «Путеводитель по эвфаузиацеям Южного океана». Заметки об исследованиях ANARE . 1 :1–45.
- ^ Бьянки, Даниэле; Мислан, КАС (январь 2016 г.). «Глобальные закономерности суточного времени и скорости вертикальной миграции по акустическим данным» . Лимнология и океанография . 61 (1). дои : 10.1002/lno.10219 .
- ^ А. Сала; М. Аззали; А. Руссо (2002). «Криль моря Росса: распространение, численность и демография Euphausia superba и Euphausia Crystallorophias во время итальянской антарктической экспедиции (январь – февраль 2000 г.)» . Сциентия Марина . 66 (2): 123–133. дои : 10.3989/scimar.2002.66n2123 .
- ^ Г. В. Хози; М. Фукучи; С. Кавагути (2003). «Развитие системы непрерывного наблюдения за планктоном Южного океана» (PDF) . Прогресс в океанографии . 58 (2–4): 263–283. Бибкод : 2003Proce..58..263H . дои : 10.1016/j.pocean.2003.08.007 . [ постоянная мертвая ссылка ]
- ^ Э. Гатен. « Меганиктипанес норвегика » . Университет Лестера . Архивировано из оригинала 1 июля 2009 года . Проверено 25 февраля 2009 г.
- ^ Э. Бринтон (1953). « Thysanopoda spinicauda , новый батипелагический гигантский эвфаузиидный ракообразный, со сравнительными заметками о T. cornuta и T. egregia ». Журнал Вашингтонской академии наук . 43 : 408–412.
- ^ «Эуфаузиацея» . Тасманский институт аквакультуры и рыболовства. Архивировано из оригинала 30 сентября 2009 года . Проверено 6 июня 2010 г.
- ^ О. Шимомура (1995). «Роль двух крайне нестабильных компонентов F и P, участвующих в биолюминесценции эвфаузиидных креветок». Журнал биолюминесценции и хемилюминесценции . 10 (2): 91–101. дои : 10.1002/bio.1170100205 . ПМИД 7676855 .
- ^ Джей Си Данлэп; Дж. У. Гастингс; О. Шимомура (1980). «Кроссреактивность между светоизлучающими системами отдаленно родственных организмов: новый тип светоизлучающих соединений» . Труды Национальной академии наук . 77 (3): 1394–1397. Бибкод : 1980PNAS...77.1394D . дои : 10.1073/pnas.77.3.1394 . JSTOR 8463 . ПМЦ 348501 . ПМИД 16592787 .
- ^ Пи Джей Херринг; Э.А. Виддер (2001). «Биолюминесценция планктона и нектона» . В Дж. Х. Стиле; С.А. Торп; К.К. Турекян (ред.). Энциклопедия наук об океане . Том. 1. Академик Пресс , Сан-Диего. стр. 308–317 . ISBN 978-0-12-227430-5 .
- ^ С.М. Линдсей; М. И. Латц (1999). Экспериментальные доказательства люминесцентного затенения некоторых эвфаузиидных ракообразных . Собрание Американского общества лимнологии и океанографии (ASLO) по водным наукам. Санта Фе.
- ^ Сёнке Йонсен (2005). «Красное и черное: биолюминесценция и окраска животных морских глубин» (PDF) . Интегративная и сравнительная биология . 4 (2): 234–246. дои : 10.1093/icb/45.2.234 . ПМИД 21676767 . S2CID 247718 . Архивировано из оригинала (PDF) 2 октября 2005 г.
- ^ Перейти обратно: а б с д Каван Э.Л., Белчер А., Аткинсон А., Хилл С.Л., Кавагути С., МакКормак С., Мейер Б., Николь С., Ратнараджа Л., Шмидт К. и Стейнберг, ДК (2019) «Значение антарктического криля в биогеохимических циклах». Природные коммуникации , 10 (1): 1–13. два : 10.1038/s41467-019-12668-7 .
 Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0 .
- ^ Г. К. Криппс; А. Аткинсон (2000). «Жирнокислотный состав как показатель плотоядности антарктического криля Euphausia superba ». Канадский журнал рыболовства и водных наук . 57 (С3): 31–37. дои : 10.1139/f00-167 .
- ^ Перейти обратно: а б Олав Сэтер; Тронд Эрлинг Эллингсен; Вигго Мор (1986). «Липиды североатлантического криля» (PDF) . Журнал исследований липидов . 27 (3): 274–285. ПМИД 3734626 .
- ^ М. Дж. Шрамм (10 октября 2007 г.). «Крошечный криль: гиганты в морской пищевой цепи» . Национальная программа морских заповедников NOAA . Проверено 4 июня 2010 г.
- ^ Дж. Вейер (1999). «Изменение течений окрашивает Берингово море в новый оттенок синего» . НОАА Земная обсерватория . Проверено 15 июня 2005 г.
- ^ Р.Д. Бродер; Г.Х. Крузе; П.А. Ливингстон; Г. Уолтерс; Дж. Янелли; Г.Л. Шварцман; М. Степаненко; Т. Уилли-Эчеверрия (1998). Проект отчета Международного семинара FOCI по последним условиям в Беринговом море . НОАА . стр. 22–26.
- ^ Дж. Роуч (17 июля 2003 г.). «Ученые обнаружили загадочного убийцу криля» . Национальные географические новости . Архивировано из оригинала 24 июля 2003 года.
- ^ Х. Гомес-Гутьеррес; В. Т. Петерсон; А. де Робертис; Р. Д. Бродер (2003). «Массовая смертность криля, вызванная паразитоидными инфузориями». Наука . 301 (5631): 339. doi : 10.1126/science.1085164 . ПМИД 12869754 . S2CID 28471713 .
- ^ Джей Ди Шилдс; Х. Гомес-Гутьеррес (1996). « Oculophryxus bicaulis , новый род и вид изопод даджидов, паразитирующих на эвфаузиид Stylocheiron affine Hansen». Международный журнал паразитологии . 26 (3): 261–268. дои : 10.1016/0020-7519(95)00126-3 . ПМИД 8786215 .
- ^ Расти Дорнин (6 июля 1997 г.). «Популяция антарктического криля сокращается» . CNN . Проверено 18 июня 2011 г.
- ^ Доусон, Аманда Л; Кавагути, Со; Кинг, Кэтрин К.; Таунсенд, Кэти А; Кинг, Роберт; Хьюстон, Вильгельмина М; Бенгтсон Нэш, Сьюзан М (2018). «Превращение микропластика в нанопластик посредством фрагментации пищеварения антарктического криля» . Природные коммуникации . 9 (1): 1001. Бибкод : 2018NatCo...9.1001D . дои : 10.1038/s41467-018-03465-9 . ПМК 5843626 . ПМИД 29520086 .
- ^ Доктор медицины Найт (1984). «Изменения в морфогенезе личинок в популяции Euphausia pacifica в заливе Южной Калифорнии с зимы по лето 1977–1978 годов» (PDF) . Отчет CalCOFI . XXV . Архивировано из оригинала (PDF) 3 августа 2019 года . Проверено 5 ноября 2017 г.
- ^ « Эуфаузия суперба » . Информационный бюллетень о видах . Продовольственная и сельскохозяйственная организация . Проверено 4 июня 2010 г.
- ^ Р. М. Росс; Л. Б. Кетин (1986). «Насколько продуктивен антарктический криль?». Бионаука . 36 (4): 264–269. дои : 10.2307/1310217 . JSTOR 1310217 .
- ^ Жанин Кузен-Руди (2000). «Сезонное размножение, множественный нерест и плодовитость северного криля Meganyctiphanes norvegica и антарктического криля Euphausia superba ». Канадский журнал рыболовства и водных наук . 57 (С3): 6–15. дои : 10.1139/f00-165 .
- ^ Х. Гомес-Гутьеррес (2002). «Механизм вылупления и замедленное вылупление икры трех широконерестящихся видов эвфаузиид в лабораторных условиях» . Журнал исследований планктона . 24 (12): 1265–1276. дои : 10.1093/планкт/24.12.1265 .
- ^ Э. Бринтон; доктор медицинских наук Оман; А. В. Таунсенд; доктор медицины Найт; А. Л. Бриджман (2000). Евфаузииды Мирового океана . Серия компакт-дисков Всемирной базы данных биоразнообразия, Springer Verlag . ISBN 978-3-540-14673-5 . Архивировано из оригинала 26 февраля 2012 года . Проверено 4 декабря 2009 г.
- ^ Ф. Бухгольц (2003). «Опыты по физиологии южного и северного криля, Euphausia superba и Meganyctiphanes norvegica с акцентом на линьку и рост – обзор». Морское и пресноводное поведение и физиология . 36 (4): 229–247. Бибкод : 2003MFBP...36..229B . дои : 10.1080/10236240310001623376 . S2CID 85121989 .
- ^ Х.-К. Шин; С. Никол (2002). «Использование зависимости между диаметром глаза и длиной тела для выявления последствий длительного голодания антарктического криля Euphausia superba » . Серия «Прогресс в области морской экологии» . 239 : 157–167. Бибкод : 2002MEPS..239..157S . дои : 10.3354/meps239157 .
- ^ Б. Маринович; М. Мангель (1999). «Криль может сокращаться в результате экологической адаптации к временно неблагоприятным условиям» (PDF) . Экологические письма . 2 : 338–343.
- ^ Дж. Г. Гомес (1995). «Схемы распространения, численность и динамика популяций эвфаузиид Nyctiphanes simplex и Euphausia eximia у западного побережья Нижней Калифорнии, Мексика» (PDF) . Серия «Прогресс в области морской экологии» . 119 : 63–76. Бибкод : 1995MEPS..119...63G . дои : 10.3354/meps119063 .
- ^ У. Килс; П. Маршалл (1995). «Криль, как он плавает и питается – новые знания с помощью новых методов (« Антарктический криль – как он плавает и питается – новые знания с помощью новых методов »)». У И. Гемпеля; Г. Хемпель (ред.). биологии полярных океанов Опыт и . результаты Издательство Фишер . стр. 201–210. ISBN 978-3-334-60950-7 .
- ^ Р. Пайпер (2007). Необыкновенные животные: Энциклопедия любопытных и необычных животных . Гринвуд Пресс . ISBN 978-0-313-33922-6 .
- ^ Гандоми, АХ; Алави, АХ (2012). «Стадо криля: новый алгоритм оптимизации, основанный на биотехнологиях». Коммуникации в нелинейной науке и численном моделировании . 17 (12): 4831–4845. Бибкод : 2012CNSNS..17.4831G . дои : 10.1016/j.cnsns.2012.05.010 .
- ^ Дж. С. Яффе; доктор медицинских наук Оманн; А. де Робертис (1999). «Сонарные оценки уровней дневной активности Euphausia pacifica в заливе Саанич» (PDF) . Канадский журнал рыболовства и водных наук . 56 (11): 2000–2010. doi : 10.1139/cjfas-56-11-2000 . S2CID 228567512 . Архивировано из оригинала (PDF) 20 июля 2011 года.
- ^ Герайнт А. Тарлинг; Магнус Л. Джонсон (2006). «Насыщение вызывает у криля чувство погружения» . Современная биология . 16 (3): 83–84. дои : 10.1016/j.cub.2006.01.044 . ПМИД 16461267 .
- ^ Дэн Ховард (2001). «Криль» (PDF) . У Германа А. Карла; Джон Л. Чин; Эдвард Юбер; Питер Х. Стауффер; Джеймс В. Хендли II (ред.). За Золотыми воротами – океанография, геология, биология и проблемы окружающей среды в заливе Фараллонес . Геологическая служба США . стр. 133–140. Циркуляр 1198 . Проверено 8 октября 2011 г.
- ^ Уишарт, Скай (июль – август 2018 г.). «Эффект криля» . География Новой Зеландии (152): 24.
- ^ Д. Ховард. «Криль в Национальном морском заповеднике Корделл-Бэнк» . Национальное управление океанических и атмосферных исследований . Проверено 15 июня 2005 г.
- ^ Дэвид А. Демер; Стефан Г. Конти (2005). «Новая модель целевой силы указывает на увеличение количества криля в Южном океане» . Журнал морских наук ICES . 62 (1): 25–32. дои : 10.1016/j.icesjms.2004.07.027 .
- ^ У. Килс (1982). «Плавательное поведение, плавательные способности и энергетический баланс антарктического криля Euphausia superba » . Научная серия БИОМАССА 3, Серия исследований БИОМАССЫ : 1–122. Архивировано из оригинала 2 июня 2020 года . Проверено 11 ноября 2017 г.
- ^ С. Николь; Ю. Эндо (1997). «Мировой промысел криля» . Технический документ ФАО по рыболовству . 367 .
- ^ Ратнараджа Л., Боуи А.Р., Ланнузель Д., Майнерс К.М. и Никол С. (2014) «Биогеохимическая роль усатых китов и криля в круговороте питательных веществ в Южном океане». ПЛОС ОДИН , 9 (12): е114067. дои : 10.1371/journal.pone.0114067
- ^ Хопкинс, Т.Л., Эйнли, Д.Г., Торрес, Дж.Дж., Ланкрафт, Т.М., 1993. Трофическая структура в открытых водах краевой ледниковой зоны в районе слияния Уэдделл в Шотландии весной (1983). Полярная биология 13, 389–397.
- ^ Ланкрафт, Т.М., Релсенбихлер, К.Р., Робинсон, Б.Х., Хопкинс, Т.Л., Торрес, Дж.Дж., 2004. Сообщество микронектона и макрозоопланктона с преобладанием криля в проливе Крокер, Антарктида, с оценкой хищничества рыб. Глубоководные исследования II 51, 2247–2260.
- ^ Перейти обратно: а б с Гроссман, Элизабет (14 июля 2015 г.). «Ученые думают, нужно ли защищать криль от чрезмерной охоты со стороны человека» . Международное общественное радио (PRI) . Проверено 1 апреля 2017 г.
- ^ «Промысел криля и устойчивость: антарктический криль (Euphausia superba)» . Комиссия по сохранению морских живых ресурсов Антарктики. 23 апреля 2015 года . Проверено 1 апреля 2017 г.
- ^ Перейти обратно: а б Ширмайер, К. (2010). «Экологи опасаются кризиса антарктического криля» . Природа . 467 (7311): 15. дои : 10.1038/467015a . ПМИД 20811427 .
- ^ Перейти обратно: а б с «Криль – биология, экология и рыболовство» . Комиссия по сохранению морских живых ресурсов Антарктики. 28 апреля 2015 года . Проверено 1 апреля 2017 г.
- ^ Минтурн Дж. Райт (1987). «Право собственности на Антарктиду, ее живые и минеральные ресурсы» . Журнал права и окружающей среды . 4 (2): 49–78.
- ^ С. Николь; Дж. Фостер (2003). «Последние тенденции в промысле антарктического криля» . Водные живые ресурсы . 16 : 42–45. дои : 10.1016/S0990-7440(03)00004-4 .
- ^ Джош, Габбатисс (10 июля 2018 г.). «Индустрия лова криля поддерживает огромный заповедник в Антарктическом океане для защиты пингвинов, тюленей и китов» . Независимый . Проверено 10 июля 2018 г.
- ^ Перейти обратно: а б «Почему криль?» . Юго-западный научный центр рыболовства, Национальное управление океанических и атмосферных исследований США. 22 ноября 2016 года . Проверено 1 апреля 2017 г.
- ^ Чизмен М.А. (22 июля 2011 г.). «Масло криля: ответное письмо Агентства, уведомление GRAS № GRN 000371» . FDA США . Проверено 3 июня 2015 г.
- ^ Омори, М. (1978). «Мировой промысел зоопланктона: обзор». Морская биология . 48 (3): 199–205. дои : 10.1007/BF00397145 . S2CID 86540101 .
- ^ Понгсеткул, Джаксума; Бенджакул, Соттават; Сампавапол, Пеннани; Осако, Кадзуфуми; Фэйтхонг, Нандша (17 сентября 2014 г.). «Химический состав и физические свойства пасты из соленых креветок (Капи) производства Таиланда» . Международные водные исследования . 6 (3): 155–166. дои : 10.1007/s40071-014-0076-4 .
- ^ Абэ, Кенджи; Хашимото, Канехиса (1979). Использование криля в качестве материала для рыбного соуса» . Nippon Suisan Gakkaishi . 45 8): ( 1013–1017 « .
- ^ Оливейра Сантос, Сара; Так, Нильс; Су, Юньсин; Куэнка-Хименес, Франциско; Моралес-Лопес, Оскар; Гомес-Вальдес, П. Антонио; М. Вильгельмус, Моника (13 июня 2023 г.). «Плеобот: модульное роботизированное решение для метахронного плавания» . Научные отчеты . 13 (1): 9574. Бибкод : 2023NatSR..13.9574O . дои : 10.1038/s41598-023-36185-2 . ПМЦ 10264458 . ПМИД 37311777 .
Дальнейшее чтение
[ редактировать ]- Боден, Брайан П.; Джонсон, Мартин В .; Бринтон, Эдвард: «Эуфаузиацея (Рачки) северной части Тихого океана» . Бюллетень Океанографического института Скриппса . Том 6, номер 8, 1955 год.
- Бринтон, Эдвард : «Эуфаузииды вод Юго-Восточной Азии» . Отчет Наги, том 4, часть 5. Ла-Хойя: Калифорнийский университет, Океанографический институт Скриппса, 1975.
- Конвей, главный вице-президент; Уайт, Р.Г.; Хьюг-Дит-Сайлс, Ж.; Галиен, CP; Робинс, Д.Б.: Путеводитель по прибрежному и поверхностному зоопланктону юго-западной части Индийского океана. Архивировано 23 октября 2012 г. в Wayback Machine , Order Euphausiacea , Периодическая публикация Ассоциации морской биологии Соединенного Королевства № 15, Плимут, Великобритания, 2003 г. .
- Эверсон, И. (ред.): Криль: биология, экология и рыболовство . Оксфорд, Блэквелл Сайенс; 2000. ISBN 0-632-05565-0 .
- Хамнер, Уильям М. (май 1984 г.). «Криль — неиспользованный дар моря?». Нэшнл Географик . Том. 165, нет. 5. стр. 626–642. ISSN 0027-9358 . OCLC 643483454 .
- Мочлин, Дж.: Euphausiacea: взрослые особи. Архивировано 15 мая 2011 г. в Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Идентификационные листы взрослого криля со множеством линейных рисунков. PDF- файл, 2 Мб .
- Мочлин, Дж.: Euphausiacea: Личинки. Архивировано 19 апреля 2012 г. в Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Идентификационные листы личиночных стадий криля со множеством линейных рисунков. PDF-файл, 3 Мб.
- Тетт, П.: Биология эвфаузиид , конспекты лекций из курса морской биологии 2003 года в Университете Нейпира .
- Тетт, П.: Биолюминесценция , конспекты лекций из издания того же курса 1999/2000 гг.
Внешние ссылки
[ редактировать ]- Веб-камера крилевого аквариума Австралийского антарктического отдела
- Анимация «Энергия Антарктики» Лизы Робертс

Основные промыслового рыболовства группы видов |
|---|



| Базы данных органов управления : Национальные |
|---|