Дисферлин, также известный как связанный с дистрофией Fer-1-подобный белок, представляет собой белок , который у человека кодируется DYSF геном . [5] Дисферлин связан с восстановлением плазматической мембраны. [6] стабилизация передачи сигналов кальция [7] [8] [9] и развитие системы Т-канальцев мышцы [10] Дефект гена DYSF, расположенного на хромосоме 2р 12-14, приводит к нескольким типам мышечной дистрофии ; включая миопатию Миёши (ММ), поясно- конечностную мышечную дистрофию типа 2В (ЛГМД2В) и дистальную миопатию (ДМ). Снижение или отсутствие дисферлина, называемое дисферлинопатией , обычно становится очевидным на третьем или четвертом десятилетии жизни и характеризуется слабостью и атрофией различных произвольных скелетных мышц . [11] Патогенные мутации, приводящие к дисферлинопатии, могут возникать во всем гене DYSF.
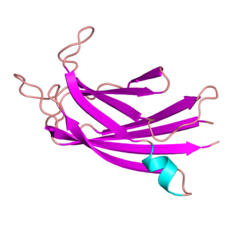
Человеческий белок дисферлин представляет собой трансмембранный белок типа II массой 237 килодальтон. [12] [13] [14] [15] [16] Он содержит большой внутриклеточный цитоплазматический N-концевой домен, крайний С-концевой трансмембранный домен и короткий С-концевой внеклеточный домен. Цитозольный домен дисферлина состоит из семи высококонсервативных доменов C2 (C2A-G), которые консервативны в нескольких белках семейства ферлинов , включая гомолог дисферлина миоферлин . [17] [18] [13] Фактически, домен C2 в любом данном положении более похож на домен C2 в соответствующем положении у других членов семейства ферлинов, чем на соседний домен C2 в том же белке. Это предполагает, что каждый отдельный домен C2 может фактически играть специфическую роль в функции дисферлина, и было показано, что каждый из них необходим для двух ролей дисферлина: стабилизации передачи сигналов кальция и восстановления мембран. [19] Мутации в каждом из этих доменов могут вызывать дисферлинопатию. Была расшифрована кристаллическая структура домена C2A человеческого дисферлина, и показано, что домен C2A меняет конформацию при взаимодействии с ионами кальция. [13] что согласуется с растущим количеством данных, свидетельствующих о том, что домен C2A играет роль в кальций-зависимом связывании липидов. [20] Его способность стабилизировать передачу сигналов кальция в интактном белке дисферлина зависит от его кальцийсвязывающей активности. [21] Помимо доменов C2, дисферлин также содержит домены «FerA» и «DysF». Мутации в обоих FerA [22] и ДисФ [23] может вызвать мышечную дистрофию. Домен DysF имеет интересную структуру: он содержит один домен DysF внутри другого домена DysF в результате дупликации гена; однако функция этого домена в настоящее время неизвестна. [23] Домен FerA консервативен среди всех членов семейства белков ферлин. Домен FerA представляет собой пучок из четырех спиралей и может взаимодействовать с мембраной, обычно кальций-зависимым образом. [22]
Наиболее интенсивно изучаемая роль дисферлина связана с клеточным процессом, называемым восстановлением мембран. Восстановление мембраны является важнейшим механизмом, с помощью которого клетки способны заклеивать серьезные раны на плазматической мембране. Считается, что мышцы особенно склонны к повреждениям мембран, поскольку мышечные клетки передают высокую силу и подвергаются циклам сокращения. Дисферлин высоко экспрессируется в мышцах и гомологичен белкам семейства ферлинов, которые, как полагают, регулируют слияние мембран у самых разных видов и типов клеток. [24] Некоторые данные свидетельствуют о том, что дисферлин может участвовать в восстановлении мембран в мышцах. Во-первых, в мышечных волокнах с дефицитом дисферлина наблюдается накопление везикул (которые имеют решающее значение для восстановления мембран в немышечных типах клеток) вблизи повреждений мембраны, что указывает на то, что дисферлин может потребоваться для слияния репарирующих везикул с плазматической мембраной. Кроме того, мышечные волокна с дефицитом дисферлина поглощают внеклеточные красители в большей степени, чем мышечные волокна дикого типа, после лазерно-индуцированного ранения in vitro. [25] Дисферлин также заметно обогащен при повреждении мембран несколькими дополнительными белками, которые, как полагают, участвуют в повторном уплотнении мембран, включая аннексин и MG53. [26] Как именно дисферлин способствует повторному уплотнению мембран, неясно, но биохимические данные указывают на то, что дисферлин может связывать липиды кальций-зависимым образом, что согласуется с ролью дисферлина в регуляции слияния репарационных везикул с сарколеммой во время восстановления мембраны. [27] Кроме того, визуализация живых клеток мышечных трубочек, экспрессирующих дисферлин-eGFP, указывает на то, что дисферлин локализуется в клеточном компартменте, который реагирует на повреждение образованием больших везикул, содержащих дисферлин, и образование этих везикул может способствовать заживлению ран. [28] Дисферлин также может участвовать в патогенезе болезни Альцгеймера. [29]
Другая хорошо изученная роль дисферлина заключается в стабилизации передачи сигналов кальция, особенно после легкой травмы. Этот подход был основан на двух наблюдениях: мышца, лишенная дисферлина и поврежденная эксцентрическими сокращениями, может восстанавливать свою плазматическую мембрану, или сарколемму, так же эффективно, как и здоровая мышца. [30] и что большая часть дисферлина в здоровых мышцах сосредоточена в поперечных канальцах в местах соединения триад, [31] [32] где регулируется высвобождение кальция. Дестабилизация передачи сигналов в дисферлинопатической мышце может привести к генерации кальциевых волн, [33] которые могут способствовать патологии заболевания. Почти каждое изменение дисферлина, влияющее на восстановление мембран, также дестабилизирует передачу сигналов кальция. [34] предполагая, что эти два вида деятельности тесно связаны. Однако примечательно, что для восстановления мембран необходимы ионы кальция, тогда как ионы кальция способствуют дестабилизации передачи сигналов, когда дисферлин отсутствует или мутирует. [35] Эти парадоксальные результаты еще предстоит согласовать.
Было показано, что дисферлин связывается сам с собой, образуя димеры и, возможно, более крупные олигомеры. [36] Также было показано, что он взаимодействует с кавеолином 3 в скелетных мышцах. [37] Считается, что это взаимодействие удерживает дисферлин внутри плазматической мембраны. [38] Дисферлин также взаимодействует с MG53, и считается, что функциональное взаимодействие между дисферлином, кавеолином-3 и MG53 имеет решающее значение для восстановления мембран в скелетных мышцах. [39]
^ Башир Р., Бриттон С., Страхан Т., Кирс С., Вафиадаки Е., Лако М., Ричард И., Маршан С., Бург Н., Аргов З., Садех М., Махжне И., Маркони Г., Пассос-Буэно М.Р., Морейра Эде С., Затц М. , Бекманн Дж. С., Бушби К. (1998). «Ген, связанный с фактором сперматогенеза Caenorhabditis elegansfer-1, мутирует при мышечной дистрофии конечностей 2B». Нат. Жене . 20 (1): 37–42. дои : 10.1038/1689 . ПМИД 9731527 . S2CID 24234676 .
Башир Р., Страчан Т., Кирс С., Стивенсон А., Махжне И., Маркони Г., Нашеф Л., Бушби К.М. (1994). «Ген аутосомно-рецессивной мышечной дистрофии конечностей картируется на хромосоме 2p». Хм. Мол. Жене . 3 (3): 455–7. дои : 10.1093/hmg/3.3.455 . ПМИД 8012357 .
Лю Дж., Аоки М., Илла И., Ву С., Фардо М., Анджелини С., Серрано С., Уртисбереа Х.А., Хентати Ф., Хамида М.Б., Болега С., Калпер Э.Дж., Амато А.А., Босси К., Олтьен Дж., Бежауи К., МакКенна- Ясек Д., Хослер Б.А., Шурр Э., Арахата К., де Йонг П.Дж., Браун Р.Х. (1998). «Дисферлин, новый ген скелетных мышц, мутирует при миопатии Миёши и мышечной дистрофии пояса конечностей». Нат. Жене . 20 (1): 31–6. дои : 10.1038/1682 . ПМИД 9731526 . S2CID 12018395 .
Башир Р., Бриттон С., Страхан Т., Кирс С., Вафиадаки Е., Лако М., Ричард И., Маршан С., Бург Н., Аргов З., Садех М., Махжне И., Маркони Г., Пассос-Буэно М.Р., Морейра Эде С., Затц М. , Бекманн Дж. С., Бушби К. (1998). «Ген, связанный с фактором сперматогенеза Caenorhabditis elegansfer-1, мутирует при мышечной дистрофии конечностей 2B». Нат. Жене . 20 (1): 37–42. дои : 10.1038/1689 . ПМИД 9731527 . S2CID 24234676 .
Андерсон Л.В., Дэвисон К., Мосс Дж.А., Янг С., Каллен М.Дж., Уолш Дж., Джонсон М.А., Башир Р., Бриттон С., Кирс С., Аргов З., Махжне И., Фужерус Ф., Бекманн Дж.С., Бушби К.М. (1999). «Дисферлин представляет собой белок плазматической мембраны и экспрессируется на ранних этапах развития человека». Хм. Мол. Жене . 8 (5): 855–61. дои : 10.1093/hmg/8.5.855 . ПМИД 10196375 .
Вейлер Т., Башир Р., Андерсон Л.В., Дэвисон К., Мосс Дж.А., Бриттон С., Найлен Е., Кирс С., Вафиадаки Е., Гринберг С.Р., Бушби С.Р., Рогеманн К. (1999). «Идентичная мутация у пациентов с мышечной дистрофией пояса конечностей типа 2B или миопатией Миёши предполагает роль гена (ов)-модификаторов». Хм. Мол. Жене . 8 (5): 871–7. дои : 10.1093/hmg/8.5.871 . ПМИД 10196377 .
Мацуда С., Аоки М., Хаяси Ю.К., Хо М.Ф., Арахата К., Браун Р.Х. (1999). «Дисферлин — это белок, связанный с поверхностной мембраной, который отсутствует при миопатии Миёши». Неврология . 53 (5): 1119–22. дои : 10.1212/wnl.53.5.1119 . ПМИД 10496277 . S2CID 6068681 .
Конфалоньери П., Олива Л., Андреетта Ф., Лоренцони Р., Дасси П., Мариани Э., Моранди Л., Мора М., Корнелио Ф., Мантегацца Р. (2003). «Мышечное воспаление и усиление MHC класса I при мышечной дистрофии с недостатком дисферлина: иммунопатологическое исследование». Дж. Нейроиммунол . 142 (1–2): 130–6. дои : 10.1016/S0165-5728(03) 00255-8 ПМИД 14512171 . S2CID 37727809 .
Кальяни Р., Фортунато Ф, Джорда Р., Родолико К., Бональя МК, Сирони М., Д'Анджело М.Г., Прелле А., Локателли Ф., Тоскано А., Бресолин Н., Коми ГП (2003). «Молекулярный анализ пациентов с ПКМД-2B и ММ: выявление новых мутаций DYSF и возможного эффекта основателя в итальянской популяции». Нервно-мышечная. Разлад . 13 (10): 788–95. дои : 10.1016/S0960-8966(03)00133-0 . ПМИД 14678801 . S2CID 23179310 .
Arc.Ask3.Ru Номер скриншота №: 3cbb96d2e3705eb9fd5243345c640964__1703941680 URL1:https://arc.ask3.ru/arc/aa/3c/64/3cbb96d2e3705eb9fd5243345c640964.html Заголовок, (Title) документа по адресу, URL1: Dysferlin - Wikipedia
Данный printscreen веб страницы (снимок веб страницы, скриншот веб страницы), визуально-программная копия документа расположенного по адресу URL1 и сохраненная в файл, имеет: квалифицированную, усовершенствованную (подтверждены: метки времени, валидность сертификата), открепленную ЭЦП (приложена к данному файлу), что может быть использовано для подтверждения содержания и факта существования документа в этот момент времени. Права на данный скриншот принадлежат администрации Ask3.ru, использование в качестве доказательства только с письменного разрешения правообладателя скриншота. Администрация Ask3.ru не несет ответственности за информацию размещенную на данном скриншоте. Права на прочие зарегистрированные элементы любого права, изображенные на снимках принадлежат их владельцам. Качество перевода предоставляется как есть. Любые претензии, иски не могут быть предъявлены. Если вы не согласны с любым пунктом перечисленным выше, вы не можете использовать данный сайт и информация размещенную на нем (сайте/странице), немедленно покиньте данный сайт. В случае нарушения любого пункта перечисленного выше, штраф 55! (Пятьдесят пять факториал, Денежную единицу (имеющую самостоятельную стоимость) можете выбрать самостоятельно, выплаичвается товарами в течение 7 дней с момента нарушения.)