Миелин-олигодендроцитарный гликопротеин
| MOG | ||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Идентификаторы | ||||||||||||||||||||||||||||||||||||||||||||||||
| Псевдонимы | MOG , BTN6, BTNL11, MOGIG2, NRCLP7, гликопротеин миелин-олигодендроцитов | |||||||||||||||||||||||||||||||||||||||||||||||
| Внешние идентификаторы | OMIM : 159465 ; MGI : 97435 ; HomoloGene : 111009 ; GeneCards : MOG ; OMA : MOG - orthologs | |||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||
| Викиданные | ||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||




Миелин-олигодендроцитарный гликопротеин ( МОГ ) представляет собой гликопротеин, играет важную роль в миелинизации нервов который, как полагают , центральной нервной системы (ЦНС). У человека этот белок кодируется MOG геном . [5] [6] [7] Предполагается, что он служит необходимой «молекулой адгезии», обеспечивающей структурную целостность миелиновой оболочки, и известно, что он поздно развивается на олигодендроцитах . [8]
Молекулярная функция
[ редактировать ]Хотя основная молекулярная функция MOG еще не известна, его вероятная роль в миелиновой оболочке заключается либо в «завершении и/или поддержании» оболочки. [7] Более конкретно, предполагается, что MOG «необходим» в качестве «молекулы адгезии» на миелиновой оболочке ЦНС для обеспечения структурной целостности миелиновой оболочки. [8] "
MOG у людей «высоко гомологична». кДНК Было показано, что кодирующая область [9] для крыс, мышей и крупного рогатого скота и, следовательно, хорошо сохраняются. Это предполагает «важную биологическую роль этого белка». [7]
Физиология
[ редактировать ]Ген MOG, обнаруженный на хромосоме 6 p21.3-p22, [10] Впервые секвенирован в 1995 году. [3] Это трансмембранный белок, экспрессирующийся на поверхности клеток олигодендроцитов и на внешней поверхности миелиновых оболочек . «MOG — это количественно минорный трансмембранный белок I типа , [11] и обнаруживается исключительно в ЦНС. «Один Ig-домен экспонируется во внеклеточное пространство» [11] и, следовательно, обеспечивает легкий доступ к аутоантителам. и поэтому легко доступен и для аутоантител. [7] [11] MOG «первичный ядерный транскрипт… имеет длину 15 561 нуклеотид». [7] а у человека он имеет восемь экзонов , «разделенных семью интронами ». [7] Интроны «содержат многочисленные повторяющиеся [ sic ] ДНК. [7] "последовательности, среди которых "14 последовательностей Alu в пределах 3-х интронов", [7] и имеют диапазон от 242 до 6484 п.н.
Структура
[ редактировать ]Альтернативно сплайсированная человеческая мРНК гена MOG образует по меньшей мере девять изоформ. [12]
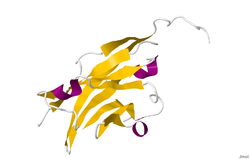
Кристаллическую структуру гликопротеина миелин-олигодендроцитов определяли методом рентгеновской дифракции с разрешением 1,45 ангстрем с использованием белка норвежской крысы. Этот белок имеет длину 139 остатков и является членом суперсемейства иммуноглобулинов. [13] Вторичная структура dssp белка на 6% спиральная и на 43% бета-листная: имеется три коротких спиральных сегмента и десять бета-цепей. [14] Бета-цепи находятся внутри двух антипараллельных бета-листов, которые образуют иммуноглобулиноподобную бета-сэндвич-складку. [15] Некоторые особенности структуры белка позволяют предположить, что MOG играет роль «адгезина в завершении и/или уплотнении миелиновой оболочки». Существует «значительная полоса» электроотрицательного заряда, начинающаяся около N-конца и простирающаяся примерно на половину длины молекулы. Кроме того, было показано, что MOG димеризуется в растворе, а индекс комплементарности формы на границе раздела димеров высок, что позволяет предположить наличие «биологически значимого димера MOG». [16]
Синтез
[ редактировать ]В процессе развития MOG формируется «очень поздно на олигодендроцитах и миелиновой оболочке». [8]
Роль в болезни
[ редактировать ]Невоспалительные демиелинизирующие заболевания
[ редактировать ]Интерес к MOG сосредоточен на его роли в демиелинизирующих заболеваниях . Некоторые из них не являются воспалительными, например, адренолейкодистрофия , болезнь исчезновения белого вещества и умственная отсталость, вызванная краснухой. [17]
Анти-MOG-ассоциированные воспалительные демиелинизирующие заболевания
[ редактировать ]MOG уделяла большую часть лабораторного внимания исследованиям рассеянного склероза. Несколько исследований показали роль антител против MOG в патогенезе рассеянного склероза. [8] [18] хотя большинство из них были написаны до открытия NMO-IgG и спектра заболеваний NMO.
Статус анти-MOG различается в зависимости от того, измеряется ли он с помощью ELISA или с помощью микрочипа ( CBA ). Правильный способ идентифицировать его — с помощью микрочипа, взаимодействия сыворотки пациента с живыми клетками и обнаружения связывающего IgG с помощью вторичного антитела, меченного флуоресцентной меткой. [19]
В моделях животных
[ редактировать ]Животные модели рассеянного склероза, а именно модели экспериментального аутоиммунного энцефаломиелита (EAE) , показали, что «MOG-специфичные модели EAE (разных линий животных) отображают/отражают рассеянный склероз человека». [8] но в основном объясняет часть неврита зрительного нерва. [20] Эти модели с антителами против MOG были тщательно исследованы и считаются единственными антителами, обладающими демиелинизирующей способностью. [8] но опять же, патология EAE ближе к NMO и ADEM, чем к сливной демиелинизации, наблюдаемой при рассеянном склерозе.
Было показано, что антитела против MOG ведут себя аналогично антителам AQP4 на животных моделях. [20] и считаются биомаркером против диагноза рассеянного склероза. [21] [22]
При серонегативном оптиконевромиелите
[ редактировать ]Было обнаружено, что аутоиммунитет против MOG участвует в большинстве AQP4-серонегативных NMO. [23] [24] а также при неврите зрительного нерва и некоторых молниеносных формах ОРЭМ . [25] Антитела MOG при NMOSD варьируются в зависимости от серопозитивного статуса. [26]
В других условиях
[ редактировать ]Наличие аутоантител против MOG было связано со следующими состояниями: [27]
- В большинстве случаев аквапорин-4-серонегативного оптикомиелита: NMO возникает из-за антиMOG-ассоциированного энцефаломиелита . [28]
- Некоторые случаи острого рассеянного энцефаломиелита , особенно рецидивирующего (MDEM) [29] и молниеносные курсы [25]
- Некоторые случаи рассеянного склероза [27]
- изолированный неврит зрительного нерва или поперечный миелит [27]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б с ENSG00000232641, ENSG00000137345, ENSG00000230885, ENSG00000236561, ENSG00000237834, ENSG00000204655, ENSG00000234623 GRCh38: Версия ансамбля 89: 0000234096, ENSG00000232641, ENSG00000137345, ENSG00000230885, ENSG00000236561, ENSG00000237834, ENSG00000204655, ENSG00000234623 – ансамбль , май 2017 год
- ^ Перейти обратно: а б с GRCm38: выпуск Ensembl 89: ENSMUSG00000076439 – Ensembl , май 2017 г.
- ^ «Ссылка на Human PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ «Ссылка на Mouse PubMed:» . Национальный центр биотехнологической информации, Национальная медицинская библиотека США .
- ^ Фам-Динь Д., Делла Гаспера Б., Керлеро де Росбо Н., Даутиньи А. (сентябрь 1995 г.). «Структура человеческого гена гликопротеина миелина / олигодендроцитов и множественных альтернативных сплайсированных изоформ». Геномика . 29 (2): 345–52. дои : 10.1006/geno.1995.9995 . ПМИД 8666381 .
- ^ Фам-Динь Д., Джонс Э.П., Питио Дж., Делла Гаспера Б., Даубас П., Маллет Дж., Ле Паслье Д., Фишер Линдал К., Даутиньи А. (1995). «Физическое картирование гена MOG человека и мыши на дистальном конце области MHC класса Ib». Иммуногенетика . 42 (5): 386–91. дои : 10.1007/bf00179400 . ПМИД 7590972 . S2CID 8310478 .
- ^ Перейти обратно: а б с д и ж г час Рот М.П., Малфрой Л., Оффер С., Севин Дж., Эно Дж., Борот Н., Понтаротти П., Коппин Х. (июль 1995 г.). «Ген миелинового олигодендроцитарного гликопротеина (MOG) человека: полная нуклеотидная последовательность и структурная характеристика». Геномика . 28 (2): 241–50. дои : 10.1006/geno.1995.1137 . ПМИД 8530032 .
- ^ Перейти обратно: а б с д и ж Бергер Т., кафедра неврологии Медицинского университета Инсбрука, интервью С. Гиллули, 24 ноября 2008 г.
- ^ Фам-Дин Д., Аллинквант Б., Руберг М., Делла Гаспера Б., Нуссбаум Дж.Л., Даутиньи А. (декабрь 1994 г.). «Характеристика и экспрессия кДНК, кодирующей гликопротеин миелина / олигодендроцитов человека». Журнал нейрохимии . 63 (6): 2353–6. дои : 10.1046/j.1471-4159.1994.63062353.x . ПМИД 7964757 . S2CID 2788720 .
- ^ Фам-Дин Д., Маттей М.Г., Нуссбаум Дж.Л., Руссель Г., Понтаротти П., Рокель Н., Матер И.Х., Артц К., Линдал К.Ф., Даутиньи А. (сентябрь 1993 г.). «Миелин/олигодендроцитарный гликопротеин является членом подмножества суперсемейства иммуноглобулинов, кодируемого главным комплексом гистосовместимости» . Труды Национальной академии наук Соединенных Штатов Америки . 90 (17): 7990–4. Бибкод : 1993PNAS...90.7990P . дои : 10.1073/pnas.90.17.7990 . ПМК 47273 . ПМИД 8367453 .
- ^ Перейти обратно: а б с Бергер Т., Рейндл М. (август 2007 г.). «Рассеянный склероз: биомаркеры заболевания по патофизиологическим признакам». Журнал неврологических наук . 259 (1–2): 21–6. дои : 10.1016/j.jns.2006.05.070 . ПМИД 17367811 . S2CID 23257594 .
- ^ Бойл Л.Х., Траэрн Дж.А., Плотнек Г., Уорд Р., Троусдейл Дж. (сентябрь 2007 г.). «Вариации сплайсинга в цитоплазматических доменах миелин-олигодендроцитарного гликопротеина влияют на его клеточную локализацию и транспорт» . Журнал нейрохимии . 102 (6): 1853–62. дои : 10.1111/j.1471-4159.2007.04687.x . ПМК 2156149 . ПМИД 17573820 .
- ^ Брейтаупт С., Шубарт А., Зандер Х., Скерра А., Хубер Р., Линингтон С., Джейкоб У. (август 2003 г.). «Структурные данные об антигенности гликопротеина миелиновых олигодендроцитов» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (16): 9446–51. Бибкод : 2003PNAS..100.9446B . дои : 10.1073/pnas.1133443100 . ПМК 170938 . ПМИД 12874380 .
- ^ Кабш В., Сандер С. (декабрь 1983 г.). «Словарь вторичной структуры белков: распознавание образов водородных связей и геометрических особенностей». Биополимеры . 22 (12): 2577–637. дои : 10.1002/bip.360221211 . ПМИД 6667333 . S2CID 29185760 .
- ^ Мурзин А.Г., Бреннер С.Е., Хаббард Т., Чотия С. (апрель 1995 г.). «SCOP: база данных структурной классификации белков для исследования последовательностей и структур». Журнал молекулярной биологии . 247 (4): 536–40. дои : 10.1016/S0022-2836(05)80134-2 . ПМИД 7723011 .
- ^ Клементс К.С., Рид Х.Х., Беддо Т., Тайнан Ф.Е., Перуджини М.А., Джонс Т.Г., Бернард К.С., Россджон Дж. (сентябрь 2003 г.). «Кристаллическая структура миелинового олигодендроцитарного гликопротеина, ключевого аутоантигена при рассеянном склерозе» . Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 11059–64. Бибкод : 2003PNAS..10011059C . дои : 10.1073/pnas.1833158100 . ЧВК 196926 . ПМИД 12960396 .
- ^ Конг Х, Цзян Ю, Тянь П (ноябрь 2011 г.). «Идентификация миелинового олигодендроцитарного гликопротеина как клеточного рецептора вируса краснухи» . Журнал вирусологии . 85 (21): 11038–47. дои : 10.1128/JVI.05398-11 . ПМК 3194935 . ПМИД 21880773 .
- ^ Бергер Т., Рубнер П., Шауцер Ф., Эгг Р., Ульмер Х., Майрингер И., Дилиц Э., Дайзенхаммер Ф., Рейндл М. (июль 2003 г.). «Антимиелиновые антитела как предиктор клинически определенного рассеянного склероза после первого события демиелинизации» . Медицинский журнал Новой Англии . 349 (2): 139–45. doi : 10.1056/NEJMoa022328 . ПМИД 12853586 .
- ^ Ичиро Накашима, Антитела к миелиновым олигодендроцитарным гликопротеинам при демиелинизирующих заболеваниях [1]
- ^ Перейти обратно: а б Кезука Т., Усуи Ю., Ямакава Н., Мацунага Ю., Мацуда Р., Масуда М., Уцуми Х., Танака К., Гото Х. (июнь 2012 г.). «Связь между NMO-антителами и анти-MOG-антителами при неврите зрительного нерва» . Журнал нейроофтальмологии . 32 (2): 107–10. дои : 10.1097/WNO.0b013e31823c9b6c . ПМИД 22157536 . S2CID 46667141 .
- ^ Кетельслегерс И.А., Ван Пелт Д.Э., Брайд С., Нойтбум Р.Ф., Кэтсман-Берреветс К.Э., Хаманн Д., Хинтцен Р.К. (октябрь 2015 г.). «Антитела к MOG противодействуют диагнозу рассеянного склероза в когорте приобретенных демиелинизирующих синдромов». Рассеянный склероз . 21 (12): 1513–20. дои : 10.1177/1352458514566666 . ПМИД 25662345 . S2CID 25321614 .
- ^ Китли Дж., Вудхолл М., Уотерс П., Лейте М.И., Девенни Э., Крейг Дж., Пэлас Дж., Винсент А. (сентябрь 2012 г.). «Антитела к миелин-олигодендроцитарным гликопротеинам у взрослых с фенотипом оптикомиелита». Неврология . 79 (12): 1273–7. дои : 10.1212/WNL.0b013e31826aac4e . ПМИД 22914827 . S2CID 855313 .
- ^ Пребстел А.К., Рудольф Г., Дорнмайр К., Коллонг Н., Шансон Дж.Б., Сандерсон Н.С., Линдберг Р.Л., Каппос Л., де Сезе Дж., Дерфусс Т. (2015). «Антитела к MOG присутствуют у подгруппы пациентов с фенотипом оптиконевромиелита» . Журнал нейровоспаления . 12 (1): 46. дои : 10.1186/s12974-015-0256-1 . ПМЦ 4359547 . ПМИД 25889963 .
- ^ СИНТИЯ МАККЕЛВИ, Пресс-репортаж, Какова роль миелин-олигодендроцитарного гликопротеина в NMO? [2]
- ^ Перейти обратно: а б Ди Паули Ф, Хофтбергер Р, Рейндл М, Бир Р, Ромберг П, Шанда К, Сато Д, Фуджихара К, Лассманн Х, Шмутцхард Э, Бергер Т (декабрь 2015 г.). «Фульминантный демиелинизирующий энцефаломиелит: результаты исследований антител и невропатологии» . Неврология: нейроиммунология и нейровоспаление . 2 (6): е175. дои : 10.1212/NXI.0000000000000175 . ПМЦ 4635550 . ПМИД 26587556 .
- ^ Бергер Т., Рейндл М. (август 2015 г.). «Биомаркеры антител при демиелинизирующих заболеваниях ЦНС – долгий и извилистый путь». Европейский журнал неврологии . 22 (8): 1162–8. дои : 10.1111/ene.12759 . ПМИД 26010364 . S2CID 39301229 .
- ^ Перейти обратно: а б с Рейндл М., Ди Паули Ф., Ростаси К., Бергер Т. (август 2013 г.). «Спектр демиелинизирующих заболеваний, связанных с аутоантителами MOG». Обзоры природы. Неврология . 9 (8): 455–61. дои : 10.1038/nrneurol.2013.118 . ПМИД 23797245 . S2CID 7219279 .
- ^ Спадаро М., Гердес Л.А., Майер М.С., Эртль-Вагнер Б., Лоран С., Крумбхольц М., Брайтаупт С., Хёген Т., Штраубе А., Гизе А., Холфельд Р., Лассманн Х., Мейнл Э., Кюмпфель Т. (март 2015 г.). «Гистопатология и клиническое течение MOG-антитело-ассоциированного энцефаломиелита» . Анналы клинической и трансляционной неврологии . 2 (3): 295–301. дои : 10.1002/acn3.164 . ПМК 4369279 . ПМИД 25815356 .
- ^ Бауманн М, Хеннес Э.М., Шанда К., Каренфорт М., Корнек Б., Зайдл Р., Дипольд К., Лауффер Х., Марквардт И., Страутманис Дж., Сырбе С., Викер С., Хофтбергер Р., Рейндл М., Ростаси К. (2016). «Дети с многофазным рассеянным энцефаломиелитом и антителами к гликопротеину миелин-олигодендроцитов (MOG): расширение спектра заболеваний, положительных на антитела MOG». Рассеянный склероз (Хаундмиллс, Бейзингсток, Англия) . 22 (14): 1821–1829. дои : 10.1177/1352458516631038 . ПМИД 26869530 . S2CID 30428892 .
Внешние ссылки
[ редактировать ]- Онлайн-менделевское наследование у человека (OMIM): 159465
- Обзор всей структурной информации, доступной в PDB для UniProt : Q61885 (мышиный миелин-олигодендроцитарный гликопротеин) на PDBe-KB .
галерея PDB |
|---|
