Верхний холмик
| Верхний холмик | |
|---|---|
 человека Схема верхних холмиков (L) среднего мозга (показаны красным) и прилегающих областей. Верхний холмик окружен красным кольцом и прозрачным красным кружком, обозначающим его местоположение. | |
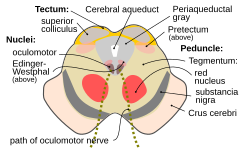 Разрез среднего мозга на уровне верхних холмиков, показывающий путь глазодвигательного нерва. | |
| Подробности | |
| Часть | Крыша |
| Идентификаторы | |
| латинский | верхний холмик |
| МеШ | D013477 |
| Нейроимена | 473 |
| НейроЛекс ID | бирнлекс_1040 |
| ТА98 | A14.1.06.015 |
| ТА2 | 5912 |
| ТД | Х3.11.03.3.01002 |
| ТО | colliculus_by_E5.14.3.3.1.4.4 E5.14.3.3.1.4.4 |
| ФМА | 62403 |
| Анатомические термины нейроанатомии | |
В нейроанатомии верхний бугорок (от латинского «верхний холм») — это структура, лежащая на крыше мозга млекопитающих среднего . [1] У немлекопитающих позвоночных гомологичная структура известна как зрительный покров или зрительная доля . [1] [2] [3] Форма прилагательного tectal обычно используется для обеих структур.
У млекопитающих верхние холмики образуют основной компонент среднего мозга. Представляет собой парную структуру и вместе с парными нижними холмиками образует четверохолмия . Верхний бугорок представляет собой слоистую структуру, рисунок которой аналогичен всем млекопитающим. [4] Слои можно сгруппировать в поверхностные слои ( оптический слой и выше) и более глубокие оставшиеся слои. Нейроны поверхностных слоев получают прямую информацию от сетчатки и реагируют почти исключительно на зрительные стимулы. Многие нейроны в более глубоких слоях также реагируют на другие модальности, а некоторые реагируют на стимулы в нескольких модальностях. [5] Более глубокие слои также содержат популяцию двигательных нейронов, способных активировать движения глаз , а также другие реакции. [6] У других позвоночных число слоев гомологичного зрительного тектума варьирует. [4]
Общая функция тектальной системы — направлять поведенческие реакции в определенные точки телесно-центрированного пространства. Каждый слой содержит топографическую карту окружающего мира в ретинотопических координатах, и активация нейронов в определенной точке карты вызывает ответ, направленный в сторону соответствующей точки пространства. У приматов верхний холмик изучен главным образом с точки зрения его роли в управлении движениями глаз. Визуальный сигнал от сетчатки или «командный» сигнал от коры головного мозга создает «всплеск» активности на тектальной карте, который, если он достаточно силен, вызывает саккадическое движение глаз . Однако даже у приматов верхний холмик также участвует в пространственно-направленных поворотах головы, движениях вытягивания рук и т.д. [7] и сдвиги внимания, не связанные с какими-либо явными движениями. [8] У других видов верхний холмик участвует в широком спектре реакций, включая повороты всего тела у ходячих крыс. У млекопитающих, и особенно у приматов, массивное расширение коры головного мозга уменьшает верхние холмики до гораздо меньшей части всего мозга. Тем не менее, он остается важным с точки зрения функции основного интегрирующего центра движений глаз.
У видов, не являющихся млекопитающими, зрительная оболочка участвует во многих реакциях, включая плавание у рыб, полет у птиц, удары языком по добыче у лягушек и удары клыками у змей. У некоторых видов, в том числе у рыб и птиц, зрительная оболочка, также известная как зрительная доля, является одним из крупнейших компонентов мозга.
Примечание по терминологии: эта статья следует терминологии, установленной в литературе, используя термин «верхний бугорок» при обсуждении млекопитающих и «зрительный покров» при обсуждении конкретных видов, не относящихся к млекопитающим, или позвоночных в целом.
Анатомия
[ редактировать ]

Верхние бугорки представляют собой структуру дорсального отдела среднего мозга и являются частью мезэнцефалической оболочки. Два верхних холмика расположены ниже/каудальнее шишковидной железы и волчанки мозолистого тела . Они перекрываются пульвинарной частью таламуса , а медиальное коленчатое ядро таламуса расположено латеральнее любого из верхних холмиков. [9] Два нижних холмика расположены непосредственно ниже/каудальнее верхних холмов; нижние и верхние холмики известны под общим названием четвероногие тела (лат. « четвероногие тела» ). Верхние бугорки крупнее нижних, хотя нижние бугорки более заметны. [10]
Плечо верхнего холмика (или верхнее плечо ) — это ветвь, которая отходит латерально от верхнего холмика и, переходя к таламусу между пульвинарным и медиальным ядрами коленчатого тела , частично продолжается в возвышение, называемое латеральным коленчатым ядром , а частично в зрительный тракт . [ нужна ссылка ]
Верхний бугорок связан с близлежащей структурой, называемой парабигеминальным ядром , часто называемой его сателлитом. В зрительном тектуме эта близлежащая структура известна как перешеек ядра . [11]
Структура
[ редактировать ]Верхний бугорок представляет собой слоистую синаптическую структуру. [12] Микроструктура верхних холмиков и зрительной крышки различается у разных видов. Как правило, всегда существует четкое различие между поверхностными слоями, которые получают входные данные в основном от зрительной системы и демонстрируют преимущественно зрительные реакции, и более глубокими слоями, которые получают множество типов входных данных и проецируются на многочисленные области мозга, связанные с моторикой. Различие между этими двумя зонами настолько четкое и последовательное, что некоторые анатомы предложили считать их отдельными структурами мозга.
У млекопитающих нейроанатомы условно выделяют семь слоев. [13] Три верхних слоя называются поверхностными :
- Lamina I или SZ , stratum zonale , представляет собой тонкий слой, состоящий из мелких миелинизированных аксонов вместе с маргинальными и горизонтальными клетками.
- Lamina II или SGS , stratum griseum superficiale («поверхностный серый слой»), содержит множество нейронов различной формы и размера.
- Lamina III или SO , stratum opticum («оптический слой»), состоит в основном из аксонов, идущих из зрительного тракта .
Далее идут два промежуточных слоя :
- Пластинка IV или SGI , stratum griseum intermedium («промежуточный серый слой»), является самым толстым слоем и заполнена множеством нейронов разных размеров. Этот слой часто имеет такую же толщину, как и все остальные слои вместе взятые. Его часто подразделяют на «верхнюю» и «нижнюю» части.
- Lamina V или SAI , промежуточный слой альбома («промежуточный белый слой»), состоит в основном из волокон различного происхождения.
Наконец, идут два глубоких слоя :
- Пластинка VI или SGP , stratum griseum profundum («глубокий серый слой»), состоит из рыхло упакованных нейронов и миелинизированных волокон.
- Пластинка VII или SAP , stratum album profundum («глубокий белый слой»), лежащая непосредственно над околоводопроводной серой , целиком состоит из волокон.
Поверхностные слои получают информацию в основном от сетчатки, областей коры головного мозга, связанных со зрением, и двух связанных с тектальной структурой структур, называемых претектумом и парабигеминальным ядром . Вход в сетчатку охватывает всю поверхностную зону и является двусторонним, хотя контрлатеральная часть более обширна. Кортикальный входной сигнал в большей степени поступает из первичной зрительной коры (область 17, V1), вторичной зрительной коры (области 18 и 19 ) и лобных полей глаза . Парабигеминальное ядро играет очень важную роль в функции тектала, которая описана ниже.
В отличие от входных сигналов поверхностных слоев, где преобладает зрение, промежуточные и глубокие слои получают входные данные от очень разнообразного набора сенсорных и моторных структур. Большинство областей коры головного мозга проецируются на эти слои, хотя входные данные из «ассоциативных» областей имеют тенденцию быть более тяжелыми, чем входные данные из первичных сенсорных или двигательных областей. [14] Однако задействованные области коры и сила их относительных проекций различаются у разных видов. [15] Другой важный входной сигнал поступает от черной субстанции , pars reticulata , компонента базальных ганглиев . Эта проекция использует тормозной нейромедиатор ГАМК и, как полагают, оказывает «запирающий» эффект на верхние холмики. Промежуточные и глубокие слои также получают информацию от ядра тройничного нерва спинного мозга , которое передает соматосенсорную информацию от лица, а также от гипоталамуса , неясной зоны , таламуса и нижних холмиков .
Помимо своих отличительных входов, поверхностная и глубокая зоны верхних холмиков также имеют отличительные выходы. Один из наиболее важных выходов поступает в легочную и латеральную промежуточную области таламуса, которые, в свою очередь, проецируются на области коры головного мозга, участвующие в управлении движениями глаз. Имеются также проекции из поверхностной зоны на претектальные ядра, латеральное коленчатое ядро таламуса и парабигеминальное ядро. Проекции из более глубоких слоев более обширны. Существует два крупных нисходящих пути, ведущих к стволу головного мозга и спинному мозгу, а также многочисленные восходящие проекции к различным сенсорным и двигательным центрам, в том числе несколько, которые участвуют в генерации движений глаз.
Оба холма также имеют нисходящие проекции на парамедиальную ретикулярную формацию моста и спинной мозг и, таким образом, могут участвовать в реакциях на стимулы быстрее, чем это позволяет корковая обработка.
Мозаичная структура
[ редактировать ]При детальном рассмотрении колликулярные слои на самом деле не представляют собой гладкие листы, а разделены на сотовое расположение отдельных столбиков. [16] Наиболее четким признаком столбчатой структуры являются холинергические сигналы, исходящие из парабигеминального ядра, окончания которого образуют равномерно расположенные кластеры, простирающиеся сверху вниз от тектума . [17] Некоторые другие нейрохимические маркеры, включая кальретинин, парвальбумин, рецепторы GAP-43 и NMDA, а также связи с многочисленными другими структурами мозга в стволе мозга и промежуточном мозге, также демонстрируют соответствующую неоднородность. [18] Общее количество колонн оценивается примерно в 100. [16] Функциональное значение этой столбчатой архитектуры неясно, но интересно, что недавние данные показали, что холинергические входы являются частью рекуррентной цепи, производящей динамику «победитель получает все» внутри тектума, как более подробно описано ниже.
Все виды, которые были исследованы, включая млекопитающих и немлекопитающих, демонстрируют компартментализацию, но существуют некоторые систематические различия в деталях этого расположения. [17] У видов с полосатой сетчаткой (в основном у видов с латерально расположенными глазами, например кроликов и оленей) компартменты покрывают всю длину СК. Однако у видов с центрально расположенной ямкой компартментализация нарушается в передней (ростральной) части СК. Эта часть SC содержит множество нейронов «фиксации», которые постоянно срабатывают, в то время как глаза остаются фиксированными в постоянном положении.
Функция
[ редактировать ]История исследования зрительного покрова ознаменовалась несколькими крупными сдвигами во взглядах. Примерно до 1970 года в большинстве исследований участвовали немлекопитающие — рыбы, лягушки, птицы — то есть виды, у которых зрительная оболочка является доминирующей структурой, получающей сигналы от глаз. Общее мнение тогда заключалось в том, что зрительная оболочка у этих видов является основным зрительным центром в мозгу немлекопитающих и, как следствие, участвует в широком спектре поведенческих реакций. [19] Однако с 1970-х по 1990-е годы нейронные записи млекопитающих, в основном обезьян, были сосредоточены в первую очередь на роли верхних холмиков в контроле движений глаз. Это направление исследований стало доминировать в литературе до такой степени, что большинство придерживалось мнения, что контроль движения глаз является единственной важной функцией млекопитающих, и эта точка зрения до сих пор отражена во многих современных учебниках.
Однако в конце 1990-х годов эксперименты с использованием животных, головы которых могли свободно двигаться, ясно показали, что SC на самом деле вызывает сдвиги взгляда , обычно состоящие из комбинированных движений головы и глаз, а не движений глаз как таковых . Это открытие возобновило интерес ко всему спектру функций верхних холмиков и привело к изучению мультисенсорной интеграции у различных видов и ситуаций. Тем не менее роль СК в управлении движениями глаз понята гораздо глубже, чем любая другая функция.
Поведенческие исследования показали, что SC не необходим для распознавания объектов, но играет решающую роль в способности направлять поведение на конкретные объекты и может поддерживать эту способность даже при отсутствии коры головного мозга. [20] Таким образом, кошки с серьезным повреждением зрительной коры не могут распознавать объекты, но все же могут следить за движущимися стимулами и ориентироваться на них, хотя и медленнее, чем обычно. Однако если половина SC удалена, кошки будут постоянно кружить в сторону поражения и компульсивно ориентироваться на объекты, расположенные там, но вообще не смогут ориентироваться на объекты, расположенные в противоположном полуполе. Эти дефициты со временем уменьшаются, но никогда не исчезают.
Движения глаз
[ редактировать ]У приматов движения глаз можно разделить на несколько типов: фиксация , при которой взгляд направлен на неподвижный предмет, причем движения глаз лишь компенсируют движения головы; плавное преследование , при котором глаза постоянно движутся, отслеживая движущийся объект; саккады , при которых глаза очень быстро перемещаются из одного места в другое; и вергентность , при которой глаза движутся одновременно в противоположных направлениях для получения или поддержания единого бинокулярного зрения. Во всех этих процессах участвует верхний бугорок, но наиболее интенсивно изучена его роль в саккадах. [21] [22] [23]
Каждый из двух холмиков — по одному на каждой стороне мозга — содержит двумерную карту, представляющую половину поля зрения. Фовеа — область максимальной чувствительности — представлена на переднем крае карты, а периферия — на заднем. Движения глаз вызываются активностью в глубоких слоях СК. Во время фиксации нейроны вблизи переднего края — фовеальной зоны — тонически активны. Во время плавного преследования активируются нейроны на небольшом расстоянии от переднего края, что приводит к небольшим движениям глаз. При саккадах нейроны активируются в области, которая представляет собой точку, к которой будет направлена саккада. Непосредственно перед саккадой активность быстро нарастает в целевом месте и снижается в других частях СК. Кодирование довольно широкое, так что для любой данной саккады профиль активности образует «холм», охватывающий значительную часть колликулярной карты: местоположение пика этого «холма» представляет собой цель саккады. [24]
SC кодирует цель перемещения взгляда, но, похоже, не определяет точные движения, необходимые для достижения этой цели. [25] Разложение смещения взгляда на движения головы и глаз, а также точная траектория движения глаз во время саккады зависят от интеграции колликулярных и неколликулярных сигналов нижестоящими двигательными областями, причем способами, которые еще не до конца понятны. Независимо от того, как движение вызывается или выполняется, СК кодирует его в «ретинотопических» координатах: то есть расположение «холма» СК соответствует фиксированному месту на сетчатке. Это, по-видимому, противоречит наблюдению о том, что стимуляция Одна точка на SC может привести к разным направлениям смещения взгляда в зависимости от исходной ориентации глаза. Однако было показано, что это происходит потому, что расположение стимула на сетчатке является нелинейной функцией местоположения цели, ориентации глаза и угла зрения. сферическая геометрия глаза. [26]
Были некоторые разногласия по поводу того, просто ли SC управляет движениями глаз и оставляет выполнение другим структурам, или же он активно участвует в выполнении саккады. В 1991 г. Муньос и др. на основании собранных ими данных предположили, что во время саккады «холм» активности в СК перемещается постепенно, отражая изменение смещения глаза от целевого местоположения во время саккады. прогрессирует. [27] В настоящее время преобладает мнение, что, хотя «холм» и слегка смещается во время саккады, он не смещается устойчивым и пропорциональным образом, как предсказывает гипотеза «движущегося холма». [28] Однако движущиеся холмы могут играть и другую роль в верхних холмиках; более поздние эксперименты продемонстрировали непрерывно движущийся холм активности зрительной памяти, когда глаза движутся медленно, при этом сохраняется отдельная цель саккады. [29]
Выходной сигнал двигательного сектора СК поступает в набор ядер среднего мозга и ствола мозга, которые преобразуют код «места», используемый СК, в код «скорости», используемый глазодвигательными нейронами. Движения глаз производятся шестью мышцами, расположенными в трех ортогонально расположенных парах. Таким образом, на уровне конечного общего пути движения глаз кодируются по существу в декартовой системе координат.
Хотя СК получает сильный сигнал непосредственно от сетчатки, у приматов он в значительной степени находится под контролем коры головного мозга, которая содержит несколько областей, участвующих в определении движений глаз. [30] Фронтальные поля глаза , часть моторной коры, участвуют в запуске намеренных саккад, а прилегающая область, дополнительные поля глаза, участвуют в организации групп саккад в последовательности. Теменные поля глаза, расположенные дальше в мозгу, участвуют в основном в рефлекторных саккадах, совершаемых в ответ на изменения зрения. Недавние доказательства [31] [32] предполагает, что первичная зрительная кора (V1) управляет рефлекторными движениями глаз, согласно гипотезе значимости V1 , используя восходящую карту значимости зрительного поля, генерируемую в V1 на основе внешних визуальных входов. [33]
SC получает зрительную информацию только в своих поверхностных слоях, тогда как более глубокие слои холмика получают также слуховую и соматосенсорную информацию и связаны со многими сенсомоторными областями мозга. Считается, что холмик в целом помогает ориентировать голову и глаза на что-то видимое и услышанное. [8] [34] [35] [36]
Верхние холмики также получают слуховую информацию от нижних холмиков. Эта слуховая информация объединяется с уже присутствующей визуальной информацией, создавая эффект чревовещания .
отвлекаемость
[ редактировать ]Помимо того, что SC связан с движениями глаз, он, по-видимому, играет важную роль в схеме, лежащей в основе отвлекаемости. Повышенная отвлекаемость возникает при нормальном старении. [37] а также является центральным признаком ряда заболеваний, включая синдром дефицита внимания и гиперактивности (СДВГ). [38] Исследования показали, что поражение СК у ряда видов может привести к повышенной отвлекаемости. [39] [40] а у людей снятие тормозного контроля над СК со стороны префронтальной коры, что приводит к увеличению активности в этой области, также увеличивает отвлекаемость. [41] Исследование на животной модели СДВГ, крысе со спонтанной гипертензией, также показывает изменение колликулярно-зависимого поведения. [42] [43] и физиология. [43] Более того, амфетамин (основной метод лечения СДВГ) также подавляет активность холмиков у здоровых животных. [44]
Другие животные
[ редактировать ]Другие млекопитающие
[ редактировать ]Приматы
[ редактировать ]Обычно считается, что верхний холмик приматов уникален среди млекопитающих , поскольку он не содержит полной карты поля зрения, видимого контралатеральным глазом. Вместо этого, как и зрительная кора и латеральное коленчатое ядро , каждый холмик представляет только контрлатеральную половину поля зрения , вплоть до средней линии, и исключает представительство ипсилатеральной половины. [45] Такая функциональная особенность объясняется отсутствием у приматов анатомических связей между ганглиозными клетками височной половины сетчатки и контралатеральными верхними холмиками. У других млекопитающих ганглиозные клетки сетчатки по всей контралатеральной сетчатке выступают на контралатеральный холмик. Это различие между приматами и не-приматами было одним из ключевых доказательств в поддержку теории летающих приматов, предложенной австралийским нейробиологом Джеком Петтигрю в 1986 году, после того как он обнаружил, что летучие лисицы ( мега-летучие мыши ) напоминают приматов с точки зрения строения тела. анатомические связи между сетчаткой и верхними холмиками. [46]
Кошки
[ редактировать ]У кошек верхний холмик выступает через ретикулярную формацию и взаимодействует с мотонейронами ствола мозга . [47]
Летучие мыши
[ редактировать ]На самом деле летучие мыши не слепы, но для навигации и захвата добычи они гораздо больше зависят от эхолокации, чем от зрения. Они получают информацию об окружающем мире, издавая звуковые сигналы, а затем прислушиваясь к эху. Их мозг высокоспециализирован для этого процесса, и некоторые из этих специализаций проявляются в верхних холмиках. [48] У летучих мышей проекция сетчатки занимает лишь тонкую зону под поверхностью, но имеются обширные входные сигналы из слуховых областей и выходные данные в двигательные области, способные ориентировать уши, голову или тело. Эхо, идущее с разных направлений, активирует нейроны в разных местах холмиков. [49] а активация колликулярных нейронов влияет на чириканье, издаваемое летучими мышами. Таким образом, есть веские основания полагать, что верхний бугорок выполняет те же функции для слухового поведения летучих мышей, которые он выполняет для зрительного поведения других видов.
Летучих мышей обычно делят на две основные группы: Microchiroptera (самые многочисленные и часто встречающиеся по всему миру) и Megachiroptera (фруктовые летучие мыши, обитающие в Азии, Африке и Австралазии). За одним исключением, летучие мыши-мегабаты не обладают эхолокацией и для навигации полагаются на развитое зрение. Зрительные рецептивные поля нейронов верхних холмиков у этих животных образуют точную карту сетчатки , подобную той, что обнаружена у кошек и приматов .
Грызуны
[ редактировать ]Было высказано предположение, что верхние холмики у грызунов опосредуют сенсорно-ориентированный подход и поведение избегания. [50] [51] Исследования с использованием инструментов анализа цепей верхних холмиков мышей выявили несколько важных функций. [12] В серии исследований исследователи определили набор модулей цепи Инь-Ян в верхних холмиках, которые инициируют поведение по захвату добычи и избеганию хищников у мышей. [52] [53] [54] [55] Используя секвенирование одноклеточной РНК , исследователи проанализировали профили экспрессии генов нейронов верхних холмиков и идентифицировали уникальные генетические маркеры этих модулей цепи. [56]
Другие позвоночные
[ редактировать ]Оптический тектум
[ редактировать ]

Зрительная оболочка — это зрительный центр в мозгу немлекопитающих, который развивается из крыльевой пластинки среднего мозга. У этих других позвоночных соединения от зрительной оболочки важны для распознавания и реакции на объекты различного размера, чему способствуют возбуждающие передатчики зрительного нерва, такие как L-глутамат . [58]
Нарушение зрительного восприятия на ранних стадиях развития рыбок данио приводит к изменению тектальной активности. Изменения в тектальной активности привели к неспособности успешно охотиться и ловить добычу. [59] Ингибирующая передача сигналов гипоталамуса глубокому тектальному нейропилю важна в тектальном процессинге у личинок рыбок данио. Тектальный нейропиль содержит структуры, включающие аксоны и дендриты перивентрикулярных нейронов . Нейропиль также содержит ГАМКергические поверхностные тормозные нейроны, расположенные в зрительном слое . [60] Вместо большой коры головного мозга у рыбок данио имеется относительно большая зрительная оболочка, которая, как предполагается, выполняет часть зрительной обработки, которую кора выполняет у млекопитающих. [61]
Недавние исследования повреждений показали, что зрительная оболочка не влияет на двигательные реакции более высокого порядка, такие как оптомоторная реакция или оптокинетическая реакция . [62] но может быть более неотъемлемой частью сигналов более низкого порядка при восприятии движения, например, при идентификации небольших объектов. [63]
Зрительная оболочка — один из фундаментальных компонентов мозга позвоночных , существующий у многих видов. [64] Некоторые аспекты структуры очень последовательны, в том числе структура, состоящая из нескольких слоев, с плотным входным сигналом от зрительных трактов к поверхностным слоям и другим сильным входным сигналом, передающим соматосенсорный вход в более глубокие слои. Другие аспекты сильно варьируются, например, общее количество слоев (от 3 у африканской двоякодышащей рыбы до 15 у золотой рыбки). [65] ), а также число клеток разных типов (от 2 у двоякодышащей рыбы до 27 у домового воробья [65] ).
Покрытие зрительного нерва тесно связано с прилегающей структурой, называемой перешейком ядра , которая вызвала большой интерес, поскольку, очевидно, вносит очень важный вклад в функцию покрышки. [66] (В верхних холмиках подобная структура называется парабигеминальным ядром ). Ядро перешейка разделено на две части, называемые перешейком магноцеллюлярной части (Imc; «часть с большими клетками») и перешейком парвоцеллюлярной части (Ipc); «часть с мелкими клетками»). Связи между тремя областями — зрительным тектумом, Ipc и Imc — топографические. Нейроны поверхностных слоев зрительного покрова проецируются в соответствующие точки Ipc и Imc. Прогнозы на IPC четко сфокусированы, тогда как прогнозы на Imc несколько более размыты. Ipc вызывает четко сфокусированные холинергические проекции как на Imc, так и на покрышку зрительного нерва. В зрительном тектуме холинергические входы от Ipc разветвляются, образуя терминали, которые проходят через всю колонку сверху вниз. Imc, напротив, дает начало ГАМКергическим проекциям на Ipc и зрительный тектум, которые очень широко распространяются в латеральных измерениях, охватывая большую часть ретинотопической карты. Таким образом, цепь tectum-Ipc-Imc заставляет тектальную активность производить рекуррентную обратную связь, которая включает в себя строго сфокусированное возбуждение небольшого столбца соседних тектальных нейронов вместе с глобальным торможением удаленных тектальных нейронов.
Зрительная оболочка участвует во многих реакциях, включая плавание у рыб, полет у птиц, удары языком по направлению к добыче у лягушек и удары клыками у змей. У некоторых видов, в том числе у рыб и птиц, зрительная оболочка, также известная как зрительная доля, является одним из крупнейших компонентов мозга.
У миксины, миноги и акулы это относительно небольшая структура, но у костистых рыб она сильно расширена, в некоторых случаях становясь самой крупной структурой мозга. У земноводных, рептилий и особенно птиц он также является весьма значимым компонентом. [65]
У змей , которые способны улавливать инфракрасное излучение , таких как питоны и гадюки , первоначальный нервный сигнал поступает через тройничный нерв, а не через зрительный тракт . Остальная часть обработки аналогична обработке зрительных ощущений и, таким образом, задействует зрительную покрышку. [67]
Рыба
[ редактировать ]
Минога
[ редактировать ]Минога . широко изучалась, поскольку у нее относительно простой мозг, который, как полагают, во многих отношениях отражает структуру мозга ранних предков позвоночных Вдохновленный новаторской работой Карла Ровайнена, начавшейся в 1960-х годах (см. [68] ), с 1970-х годов Стен Гриллнер и его коллеги из Каролинского института в Стокгольме использовали миногу в качестве модельной системы, чтобы попытаться разработать принципы двигательного контроля у позвоночных, начиная со спинного мозга и поднимаясь вверх в головной мозг. [69] Как и другие системы (см. [70] Если рассматривать эту идею с исторической точки зрения), нейронные цепи в спинном мозге, по-видимому, способны генерировать некоторые основные ритмические двигательные паттерны, лежащие в основе плавания, и что эти цепи находятся под влиянием определенных локомоторных областей в стволе мозга и среднем мозге, которые, в свою очередь, находятся под влиянием высших сил. Структуры головного мозга, включая базальные ганглии и покрышку. В исследовании тектума миноги, опубликованном в 2007 году, [71] они обнаружили, что электрическая стимуляция может вызывать движения глаз, латеральные наклоны или плавательную активность, и что тип, амплитуда и направление движений варьируются в зависимости от места стимуляции в тектуме. Эти результаты были интерпретированы как соответствующие идее о том, что тектум обеспечивает целенаправленное передвижение у миноги, как это показано у других видов.
Птицы
[ редактировать ]
У птиц зрительная оболочка участвует в полете и является одним из крупнейших компонентов мозга. Изучение зрительной обработки птиц позволило лучше понять эту функцию у млекопитающих, включая человека. [72]
См. также
[ редактировать ]Дополнительные изображения
[ редактировать ] Схема, показывающая центральные соединения зрительных нервов и зрительных трактов. (Верхний холмик виден ближе к центру.)
Схема, показывающая центральные соединения зрительных нервов и зрительных трактов. (Верхний холмик виден ближе к центру.) Верхний холмик
Верхний холмик Мозговой ствол. Вид сзади.
Мозговой ствол. Вид сзади.



Примечания
[ редактировать ]- ^ Jump up to: а б Сквайр, Л. (2013). Фундаментальная нейронаука (Четвертое изд.). Академическая пресса. п. 707. ИСБН 9780123858702 .
- ^ Кнудсен, Э.И. (июнь 2011 г.). «Контроль снизу: роль сети среднего мозга в пространственном внимании» . Европейский журнал неврологии . 33 (11): 1961–72. дои : 10.1111/j.1460-9568.2011.07696.x . ПМК 3111946 . ПМИД 21645092 .
- ^ Лю, Цунг-Хан; Цзяо, Чуан-Чин (25 января 2017 г.). «Мозаичная организация контроля структуры тела в зрительной доле кальмаров» . Журнал неврологии . 37 (4): 768–780. doi : 10.1523/JNEUROSCI.0768-16.2016 . ПМК 6597019 . ПМИД 28123014 .
- ^ Jump up to: а б Бассо, Массачусетс; Мэй, П.Дж. (15 сентября 2017 г.). «Схемы действия и познания: взгляд с верхних холмиков» . Ежегодный обзор Vision Science . 3 : 197–226. doi : 10.1146/annurev-vision-102016-061234 . ПМК 5752317 . ПМИД 28617660 .
- ^ Уоллес и др., 1998.
- ^ Ганди и др., 2011.
- ^ Луненбургер и др., 2001.
- ^ Jump up to: а б Kustov & Robinson, 1996
- ^ Синнатамби, Чамми С. (2011). Анатомия Ласта (12-е изд.). Эльзевир Австралия. п. 476. ИСБН 978-0-7295-3752-0 .
- ^ «IX. Неврология. 4б. Средний мозг или средний мозг. Грей, Генри. 1918. Анатомия человеческого тела» . www.bartleby.com . Проверено 10 октября 2019 г.
- ^ Ван, СР (2003). «Ядро перешейка и двойная модуляция рецептивного поля тектальных нейронов немлекопитающих». Обзоры исследований мозга . 41 (1): 13–25. дои : 10.1016/s0165-0173(02)00217-5 . ПМИД 12505645 .
- ^ Jump up to: а б Ито, С; Фельдхайм, Д.А. (2018). «Верхний бугорок мыши: новая модель для изучения формирования и функции цепей» . Границы в нейронных цепях . 12:10 . doi : 10.3389/fncir.2018.00010 . ПМК 5816945 . ПМИД 29487505 .
- ^ Уэрта и Хартинг, 1984.
- ^ Агстер, Кара Л.; Беруэлл, Ребекка Д. (декабрь 2009 г.). «Корковые эфференты периринальной, постринальной и энторинальной коры крысы» . Гиппокамп . 19 (12): 1159–1186. дои : 10.1002/hipo.20578 . ПМК 3066185 . ПМИД 19360714 .
- ^ Клемо Х.Р., Штейн Б.Е. (1984). «Топографическая организация соматосенсорных кортико-тектальных влияний у кошек». Журнал нейрофизиологии . 51 (5): 843–858. дои : 10.1152/jn.1984.51.5.843 . ПМИД 6726314 .
- ^ Jump up to: а б Шавалье и Мана, 2000 г.
- ^ Jump up to: а б Иллинг, 1996 г.
- ^ Мана и Шевалье, 2001 г.
- ^ Райхенталь, Адам; Бен-Тов, Мор; Сегев, Ронен (2018). «Схемы кодирования в оптическом покрове лучника» . Границы в нейронных цепях . 12:18 . doi : 10.3389/fncir.2018.00018 . ПМЦ 5845554 . ПМИД 29559898 .
- ^ Спраг, 1996.
- ^ Бассо, Массачусетс; Мэй, Пи Джей (2017). «Схемы действия и познания: взгляд с верхних холмиков» . Ежегодный обзор Vision Science . 3 : 197–226. doi : 10.1146/annurev-vision-102016-061234 . ПМК 5752317 . ПМИД 28617660 .
- ^ Фурлан, М; Смит, AT; Уокер, Р. (2015). «Активность верхних холмиков человека, связанная с подготовкой и выполнением эндогенных саккад» . Журнал нейрофизиологии . 114 (2): 1048–1058. дои : 10.1152/jn.00825.2014 . ПМЦ 4725108 . ПМИД 26041830 .
- ^ Мощовакис, АК (1996). «Верхние бугорки и контроль движений глаз». Современное мнение в нейробиологии . 6 (6): 811–816. дои : 10.1016/s0959-4388(96)80032-8 . ПМИД 9000018 .
- ^ Соетеджо, Р; Канеко, CRS; Фукс, А.Ф. (2002). «Доказательства против движущегося холма в верхних холмиках во время саккадических движений глаз у обезьяны». Журнал нейрофизиологии . 87 (6): 2778–2789. дои : 10.1152/jn.2002.87.6.2778 . ПМИД 12037180 .
- ^ Спаркс и Ганди, 2003 г.
- ^ Клиер и др., 2001.
- ^ Муньос и др., 1991.
- ^ Соетеджо и др., 2002 г.
- ^ Дэш и др., 2015.
- ^ Пьеро-Дезейлиньи и др., 2003 г.
- ^ Ян, Инь; Чжаопин, Ли; Ли, Ву (09 октября 2018 г.). «Значимость снизу вверх и обучение сверху вниз в первичной зрительной коре обезьян» . Труды Национальной академии наук . 115 (41): 10499–10504. Бибкод : 2018PNAS..11510499Y . дои : 10.1073/pnas.1803854115 . ПМК 6187116 . ПМИД 30254154 .
- ^ Чжан, Силинь; Чжаопин, Ли; Чжоу, Тяньган; Фанг, Фанг (12 января 2012 г.). «Нейронная активность в версии 1 создает восходящую карту значимости» . Нейрон . 73 (1): 183–192. дои : 10.1016/j.neuron.2011.10.035 . ISSN 0896-6273 . ПМИД 22243756 .
- ^ Ли, Чжаопин (1 января 2002 г.). «Карта значимости в первичной зрительной коре» . Тенденции в когнитивных науках . 6 (1): 9–16. дои : 10.1016/S1364-6613(00)01817-9 . ISSN 1364-6613 . ПМИД 11849610 . S2CID 13411369 .
- ^ Клиер и др., 2003.
- ^ Краузлис и др., 2004 г.
- ^ Спаркс, 1999.
- ^ Прендергаст, Массачусетс; Джексон, штат Вашингтон; Терри, А.В.; Килле, Нью-Джерси; Арнерик, СП; Декер, Миссури; Буккафуско, Джей-Джей (1 марта 1998 г.). «Возрастные различия в отвлекаемости и реакции на метилфенидат у обезьян» . Кора головного мозга . 8 (2): 164–172. дои : 10.1093/cercor/8.2.164 . ISSN 1047-3211 . ПМИД 9542895 .
- ^ Дуглас, В. (1983). Развивающая нейропсихиатрия . Нью-Йорк: Гилфорд Пресс. стр. 280–329.
- ^ Гудейл, Массачусетс; Форман, НП; Милнер, AD (1 марта 1978 г.). «Визуальная ориентация у крыс: диссоциация нарушений после кортикальных и колликулярных поражений». Экспериментальное исследование мозга . 31 (3): 445–457. дои : 10.1007/BF00237301 . ISSN 0014-4819 . ПМИД 648607 . S2CID 41871236 .
- ^ Милнер, AD; Форман, НП; Гудейл, Массачусетс (1 января 1978 г.). «Способность различать и отвлекаться после поражений префронтальной коры или верхних бугорков у короткохвостых макак». Нейропсихология . 16 (4): 381–390. дои : 10.1016/0028-3932(78)90062-3 . ПМИД 99682 . S2CID 35425621 .
- ^ Гаймар, Бертран; Франсуа, Шанталь; Плонер, Кристоф Дж.; Конди, Карин; Риво-Пешу, Софи (1 апреля 2003 г.). «Прямой префронтотектальный тракт против отвлекаемости в человеческом мозге». Анналы неврологии . 53 (4): 542–545. дои : 10.1002/ана.10560 . ISSN 1531-8249 . ПМИД 12666125 . S2CID 40340479 .
- ^ Домметт, Элеонора Дж.; Рострон, Клэр Л. (1 ноября 2011 г.). «Аномальное поведение при восстановлении воздуха в модели СДВГ у крыс со спонтанной гипертензией». Экспериментальное исследование мозга . 215 (1): 45–52. дои : 10.1007/s00221-011-2869-7 . ISSN 0014-4819 . ПМИД 21931982 . S2CID 18981985 .
- ^ Jump up to: а б Брейс, ЛР; Краев И.; Рострон, CL; Стюарт, МГ; Овертон, PG; Домметт, Э.Дж. (2015). «Измененная обработка изображений в модели синдрома дефицита внимания и гиперактивности на грызунах» . Нейронаука . 303 : 364–377. doi : 10.1016/j.neuroscience.2015.07.003 . ПМИД 26166731 . S2CID 38148654 .
- ^ Клементс, К.М.; Девоншир, Индиана; Рейнольдс, JNJ; Овертон, PG (2014). «Усиление зрительных реакций верхних холмиков на животной модели синдрома дефицита внимания с гиперактивностью и их подавление d-амфетамином». Нейронаука . 274 : 289–298. doi : 10.1016/j.neuroscience.2014.05.054 . ПМИД 24905438 . S2CID 38787839 .
- ^ Лейн и др., 1973.
- ^ Петтигрю, 1986.
- ^ Прехт, В. (1974). «Тектальные влияния на мотонейроны глаза кошки». Исследования мозга . 20 (1): 27–40. дои : 10.1016/0006-8993(74)90890-7 . ПМИД 4373140 .
- ^ Улановский и Мосс, 2008.
- ^ Валентин и Мосс, 1997.
- ^ Вестби, GWM; Кей, штат Калифорния; Редгрейв, П.; Дин, П.; Баннистер, М. (август 1990 г.). «Выходные пути от опосредующего подхода и избегания верхних холмиков крысы имеют разные сенсорные свойства». Экспериментальное исследование мозга . 81 (3): 626–638. дои : 10.1007/BF02423513 . ПМИД 2226694 . S2CID 22140043 .
- ^ Коэн, доктор юридических наук; Кастро-Аламанкос, Массачусетс (23 июня 2010 г.). «Нейронные корреляты активного избегающего поведения в верхних бугорках» . Журнал неврологии . 30 (25): 8502–8511. doi : 10.1523/JNEUROSCI.1497-10.2010 . ПМЦ 2905738 . ПМИД 20573897 .
- ^ Шан, Цунпин; Лю, Чжихуэй; Чен, Цзыцзюнь; Ши, Инчао; Ван, Цянь; Лю, Су; Ли, Дапенг; Цао, Пэн (26 июня 2015 г.). «Парвальбумин-положительный возбуждающий зрительный путь, вызывающий реакцию страха у мышей» . Наука . 348 (6242): 1472–1477. Бибкод : 2015Sci...348.1472S . дои : 10.1126/science.aaa8694 . ISSN 0036-8075 . ПМИД 26113723 . S2CID 40701489 .
- ^ Шан, Цунпин; Чен, Цзыцзюнь; Лю, Эксюэ; Ли, Ян; Чжан, Цзяцзин; Цюй, Баоле; Ян, Фэй; Чжан, Янин; Лю, Вэйсю; Лю, Чжихуэй; Го, Сяофэй (декабрь 2018 г.). «Дивергентные цепи среднего мозга управляют реакциями бегства и замирания на надвигающиеся стимулы у мышей» . Природные коммуникации . 9 (1): 1232. Бибкод : 2018NatCo...9.1232S . дои : 10.1038/s41467-018-03580-7 . ISSN 2041-1723 . ПМЦ 5964329 . ПМИД 29581428 .
- ^ Шан, Цунпин; Ли, Дапенг; Се, Чэнь, Цзыцзюнь; Ли, Ян, И; Шэнь, Цао, Пэн (июнь 2019 г.) . вызвал хищническую охоту на мышей» . Nature Neuroscience . 22 (6): 909–920. : 10.1038 /s41593-019-0405-4 . ISSN 1097-6256 . PMID 31127260. . S2CID 163165114 doi
- ^ Хуан, Мэйчжу; Чэн, Синьюй; Гу, Хуатинг; Чэнь, Цзыцзюнь; Сунь, Фанмяо (декабрь 2021 г.) . путь регулирует аппетитную локомоцию при хищной охоте у мышей» Nature Communications . 12 (1): 4409. Бибкод : 2021NatCo..12.4409H . doi : 10.1038/s41467-021-24696-3 . ISSN 2041-1723 . PMC 8292483. PMID . 34285209 .
- ^ Се, Чжиюн; Лю, Цзэюань; Чжан, Чанцзян; Гу, Хуатин; Пэй, Цин; Хуан, Мэйчжу (2021-07-28). » в среднем . eLife . 10 e69825 doi : / . eLife.69825 мозге трансформации 10.7554 . : « сенсомоторной Транскриптомное кодирование
- ^ Калтарп С.А., Пира К.У., Мисима Н., Янгдейл Э.Н., Макнил Д.С., Ливнич Б.Х., Оберг К.К. (2007). «Индукция и локализация NOGO-A во время развития мозга кур указывают на его роль, отличную от ингибирования роста нейритов» . Разработчик BMC. Биол. 7 (1): 32. дои : 10.1186/1471-213X-7-32 . ПМЦ 1865376 . ПМИД 17433109 .
- ^ Беарт, Филипп (1976). «Оценка L-глутамата как передатчика, высвобождаемого из окончаний зрительного нерва голубя». Исследования мозга . 110 (1): 99–114. дои : 10.1016/0006-8993(76)90211-0 . ПМИД 6128 . S2CID 24316801 .
- ^ Авитан, Л.; Пуич, З.; Мёльтер, Дж.; Ван Де Полл, М.; Сан, Б.; Тенг, Х.; Амор, Р.; Скотт, ЕК; Гудхилл, Дж.Дж. (2017). «Спонтанная активность в покрышке рыбок данио реорганизуется в процессе развития и находится под влиянием зрительного опыта» . Современная биология . 27 (16): 2407–2419. дои : 10.1016/j.cub.2017.06.056 . ПМИД 28781054 .
- ^ Данн, Тимоти В.; и др. (2016). «Нейральные цепи, лежащие в основе визуально вызванных побегов у личинок данио» . Нейрон . 89 (3): 613–28. дои : 10.1016/j.neuron.2015.12.021 . ПМЦ 4742414 . ПМИД 26804997 .
- ^ Хип, Лос-Анджелес; Ванваллегем, Греция; Томпсон, AW; Фавр-Бюль, я; Рубинштейн-Данлоп, Х; Скотт, ЭК (2018). «Гипоталамические проекции на зрительный покров личинки данио» . Передний Нейроанат . 11 : 135. дои : 10.3389/fnana.2017.00135 . ПМЦ 5777135 . ПМИД 29403362 .
- ^ Розер, Тобиас (2003). «Визуомоторное поведение личинок рыбок данио после лазерной абляции зрительного покрова под контролем GFP» . Журнал неврологии . 23 (9): 3726–3734. doi : 10.1523/JNEUROSCI.23-09-03726.2003 . ПМК 6742205 . ПМИД 12736343 .
- ^ Баркер, Элисон (2015). «Сенсомоторное принятие решений в тектуме данио» . Современная биология . 25 (21): 2804–2814. дои : 10.1016/j.cub.2015.09.055 . ПМИД 26592341 .
- ^ Максимино, 2008 г.
- ^ Jump up to: а б с Норткатт, 2002 г.
- ^ Энрикес, Педро М.; Рахман, Нилой; Джексон, Сэмюэл Э.; Бьянко, Исаак Х. (3 июня 2019 г.). «Ядро перешейка необходимо для поддержания преследования цели во время визуально управляемой ловли добычи» . Современная биология . 29 (11): 1771–1786.е5. дои : 10.1016/j.cub.2019.04.064 . ПМЦ 6557330 . ПМИД 31104935 .
- ^ Хартлайн и др., 1978.
- ^ Ровайнен С. (1979) Нейробиология миног. Физиологические обзоры 59:1007-1077.
- ^ Гриллнер, 2003 г.
- ^ Стюарт Д., Халтборн Х. (2008), Томас Грэм Браун (1882–1965), Андерс Лундберг (1920–) и нейронный контроль ходьбы. Brain Res Rev 59:74-95.
- ^ Сайто и др., 2007 г.
- ^ Уайли, доктор медицинских наук; Гутьеррес-Ибаньес, Ц; Пакан, Дж. М.; Иванюк, А.Н. (декабрь 2009 г.). «Оптическая оболочка птиц: наш путь к пониманию обработки визуальной информации». Канадский журнал экспериментальной психологии . 63 (4): 328–38. дои : 10.1037/a0016826 . ПМИД 20025392 . S2CID 2712427 .
Ссылки
[ редактировать ]- Шевалье, Г; Мана С (2000). «Сотообразная структура промежуточных слоев верхних холмиков крысы, с дополнительными наблюдениями у нескольких других млекопитающих: формирование паттерна AChE». Джей Комп Нейрол . 419 (2): 137–53. doi : 10.1002/(SICI)1096-9861(20000403)419:2<137::AID-CNE1>3.0.CO;2-6 . ПМИД 10722995 . S2CID 26050145 .
- Дэш, С; Ян Х; Ван Х; Кроуфорд Джей Ди (2015). «Непрерывное обновление зрительно-пространственной памяти в верхних холмиках при медленных движениях глаз» . Курр Биол . 25 (3): 267–74. дои : 10.1016/j.cub.2014.11.064 . ПМИД 25601549 .
- Дин, П; Редгрейв П; Вестби Г.В. (1989). «Событие или чрезвычайная ситуация? Две системы реагирования в верхних холмиках млекопитающих». Тенденции нейробиологии . 12 (4): 137–47. дои : 10.1016/0166-2236(89)90052-0 . ПМИД 2470171 . S2CID 25268593 .
- Ганди, Нью-Джерси; Катани ХА (2011). «Двигатели верхних холмиков» . Анну преподобный Neurosci . 34 : 205–231. doi : 10.1146/annurev-neuro-061010-113728 . ПМЦ 3641825 . ПМИД 21456962 .
- Грильнер, С. (2003). «Двигательная инфраструктура: от ионных каналов к нейронным сетям». Обзоры природы Неврология . 4 (7): 573–86. дои : 10.1038/nrn1137 . ПМИД 12838332 . S2CID 4303607 .
- Хартлайн, штат Пенсильвания; Касс Л; Петля МС (1978). «Слияние модальностей в зрительном тектуме: инфракрасная и визуальная интеграция у гремучих змей». Наука . 199 (4334): 1225–9. Бибкод : 1978Sci...199.1225H . дои : 10.1126/science.628839 . ПМИД 628839 .
- Уэрта, МФ; Хартинг Дж. К. (1984). Ванегас Х (ред.). Сравнительная неврология зрительного покрова . Нью-Йорк: Пленум Пресс. стр. 687–773. ISBN 978-0-306-41236-3 .
- Иллинг, РБ (1996). «Глава 2 мозаичная архитектура верхних холмиков». Экстрагеникулостриарные механизмы, лежащие в основе зрительно-ориентированного поведения . Прогресс в исследованиях мозга. Том. 112. стр. 17–34. дои : 10.1016/S0079-6123(08)63318-X . ISBN 9780444823472 . ПМИД 8979818 .
{{cite book}}:|journal=игнорируется ( помогите ) - Кинг, Эй Джей; Шнупп JWH; Карлайл С; Смит А.Л.; Томпсон ID (1996). «Глава 24. Разработка топографически выровненных карт зрительного и слухового пространства в верхних холмах». Экстрагеникулостриарные механизмы, лежащие в основе зрительно-ориентированного поведения . Прогресс в исследованиях мозга. Том. 112. С. 335–350. дои : 10.1016/S0079-6123(08)63340-3 . ISBN 9780444823472 . ПМИД 8979840 .
{{cite book}}:|journal=игнорируется ( помогите ) - Клиер, Э.М.; Ван Х; Кроуфорд Джей Ди (2001). «Верхний холмик кодирует команды взгляда в координатах сетчатки» (PDF) . Нат Нейроски . 4 (6): 627–32. дои : 10.1038/88450 . ПМИД 11369944 . S2CID 4930662 .
- Клиер, Э; Ван Х; Кроуфорд Д. (2003). «Трехмерная координация глаз и головы реализуется ниже верхних холмиков». J Нейрофизиология . 89 (5): 2839–53. CiteSeerX 10.1.1.548.1312 . дои : 10.1152/jn.00763.2002 . ПМИД 12740415 .
- Краузлис, Р; Листон Д; Карелло С. (2004). «Выбор цели и верхний холмик: цели, выбор и гипотезы» . Видение Рез . 44 (12): 1445–51. дои : 10.1016/j.visres.2004.01.005 . ПМИД 15066403 . S2CID 5705421 .
- Кустов А; Робинсон Д. (1996). «Общий нейронный контроль смещения внимания и движений глаз» . Природа . 384 (6604): 74–77. Бибкод : 1996Natur.384...74K . дои : 10.1038/384074a0 . ПМИД 8900281 . S2CID 68917 .
- Лейн, Р.Х.; Оллман Дж.М.; Каас Дж. Х.; Миезин Ф.М. (1973). «Визуотопическая организация верхних холмиков совиной обезьяны ( Aotus trivirgatus ) и кустарникового детеныша ( Galago senegalensis )». Мозговой Рес . 60 (2): 335–49. дои : 10.1016/0006-8993(73)90794-4 . ПМИД 4202853 .
- Луненбургер, Л; Клейзер Р; Стуфорн V; Миллер Л.Е.; Хоффманн КП (2001). «Глава 8 — возможная роль верхних холмиков в координации глаз и рук». Видение: от нейронов к познанию . Прогресс в исследованиях мозга. Том. 134. стр. 109–25. дои : 10.1016/S0079-6123(01)34009-8 . ISBN 9780444505866 . ПМИД 11702538 .
{{cite book}}:|journal=игнорируется ( помогите ) - Мана, С; Шевалье Ж (2001). «Сотовая структура промежуточных слоев верхних холмиков крысы: афферентные и эфферентные связи». Нейронаука . 103 (3): 673–93. дои : 10.1016/S0306-4522(01)00026-4 . ПМИД 11274787 . S2CID 45660874 .
- Максимино, К; Соареш, Дафна (2008). Соарес, Дафна (ред.). «Эволюционные изменения сложности тектума нететрапод: кладистический подход» . ПЛОС ОДИН . 3 (10): е385. Бибкод : 2008PLoSO...3.3582M . дои : 10.1371/journal.pone.0003582 . ПМК 2571994 . ПМИД 18974789 .
- Муньос, ДП; Пелиссон Д; Гиттон Д. (1991). «Движение активности на моторной карте верхних холмиков при смещении взгляда» (PDF) . Наука . 251 (4999): 1358–60. дои : 10.1126/science.2003221 . ПМИД 2003221 .
- Норткатт, Р.Г. (2002). «Понимание эволюции мозга позвоночных» . Интегр Комп Биол . 42 (4): 743–6. дои : 10.1093/icb/42.4.743 . ПМИД 21708771 .
- Петтигрю, доктор медицинских наук (1986). «Летающие приматы? У мегалетучих мышей развитый путь от глаза к среднему мозгу». Наука . 231 (4743): 1304–6. Бибкод : 1986Sci...231.1304P . дои : 10.1126/science.3945827 . ПМИД 3945827 . S2CID 16582493 .
- Пьеро-Дезейлиньи, К; Мюри Р.М.; Плонер Си Джей; Гаймар Б; Риво-Пешу С (2003). «Корковый контроль глазных саккад у человека: модель моторики». Нейронное управление пространственным кодированием и производством действий . Прогресс в исследованиях мозга. Том. 142. стр. 3–17. дои : 10.1016/S0079-6123(03)42003-7 . ISBN 9780444509772 . ПМИД 12693251 .
{{cite book}}:|journal=игнорируется ( помогите ) - Сайто, К; Менар А; Грильнер С (2007). «Тектальный контроль локомоции, рулевого управления и движений глаз у миноги». J Нейрофизиология . 97 (4): 3093–108. дои : 10.1152/jn.00639.2006 . ПМИД 17303814 . S2CID 5711513 .
- Соетеджо, Р; Канеко ЧР; Фукс А.Ф. (2002). «Доказательства против движущегося холма в верхних холмиках во время саккадических движений глаз у обезьяны». J Нейрофизиология . 87 (6): 2778–89. дои : 10.1152/jn.2002.87.6.2778 . ПМИД 12037180 . S2CID 18294502 .
- Спаркс, Д.Л. (1999). «Концептуальные вопросы, связанные с ролью верхних холмиков в контроле взгляда». Современное мнение в нейробиологии . 9 (6): 698–707. дои : 10.1016/S0959-4388(99)00039-2 . ПМИД 10607648 . S2CID 14389002 .
- Спаркс, ДЛ; Ганди, Нью-Джерси (2003). «Одноклеточные сигналы: глазодвигательная перспектива». Нейронное управление пространственным кодированием и производством действий . Прогресс в исследованиях мозга. Том. 142. стр. 35–53. дои : 10.1016/S0079-6123(03)42005-0 . ISBN 9780444509772 . ПМИД 12693253 .
{{cite book}}:|journal=игнорируется ( помогите ) - Спрэг, Дж. М. (1996). «Глава 1. Нейронные механизмы зрительно-ориентировочных реакций». Экстрагеникулостриарные механизмы, лежащие в основе зрительно-ориентированного поведения . Прогресс в исследованиях мозга. Том. 112. стр. 1–15. дои : 10.1016/S0079-6123(08)63317-8 . ISBN 9780444823472 . ПМИД 8979817 .
{{cite book}}:|journal=игнорируется ( помогите ) - Штейн, Бельгия; Клэмман Х.П. (1981). «Контроль движений ушной раковины и сенсомоторный регистр верхних холмиков кошки». Эволюция поведения мозга . 19 (3–4): 180–192. дои : 10.1159/000121641 . ПМИД 7326575 .
- Улановский, Н; Мосс CF (2008). «Что голос летучей мыши сообщает мозгу летучей мыши» . ПНАС . 105 (25): 8491–98. Бибкод : 2008PNAS..105.8491U . дои : 10.1073/pnas.0703550105 . ПМК 2438418 . ПМИД 18562301 .
- Валентин, Д; Мосс CF (1997). «Пространственно избирательные слуховые реакции в верхних холмиках летучей мыши, определяющей эхолокацию» . Дж. Нейроски . 17 (5): 1720–33. doi : 10.1523/JNEUROSCI.17-05-01720.1997 . ПМК 6573370 . ПМИД 9030631 .
- Уоллес, Монтана; Мередит Массачусетс; Штейн Б.Е. (1998). «Мультисенсорная интеграция в верхних холмиках бдительной кошки». J Нейрофизиология . 80 (2): 1006–10. дои : 10.1152/jn.1998.80.2.1006 . ПМИД 9705489 .