Мегабат
| Мегабат | |
|---|---|

| |
| Виды летучих мышей разных подсемейств; сверху слева по часовой стрелке: большая коротконосая летучая мышь , индийская летучая лисица , соломенная летучая мышь , египетская фруктовая летучая мышь . | |
| Научная классификация | |
| Домен: | Эукариоты |
| Королевство: | животное |
| Тип: | Хордовые |
| Сорт: | Млекопитающие |
| Заказ: | Рукокрылые |
| Подотряд: | Йинптерохироптера |
| Суперсемейство: | Птероподоидея |
| Семья: | Птероподиды Грей , 1821 г. |
| Типовой род | |
| Птеропус Бриссон , 1762 г.
| |
| Подсемейства | |

| |
| Распространение мегабатов | |
| Синонимы | |
|
Pteropidae (Серый, 1821 г.) [ 1 ] | |
Мегабаты составляют семейство Pteropodidae отряда рукокрылых ( летучие мыши ). Их также называют фруктовыми летучими мышами , фруктовыми летучими мышами Старого Света или — особенно родами Acerodon и Pteropus — летучими лисцами . Они являются единственными представителями надсемейства Pteropodoidea , которое является одним из двух надсемейств подотряда Yinpterochiroptera . Внутренние подразделения Pteropodidae менялись с тех пор, как подсемейства были впервые предложены в 1917 году. Из трех подсемейств в классификации 1917 года сейчас признаны шесть, а также различные трибы . 197 видов По состоянию на 2018 год описано летучих мышей.
Ведущая теория эволюции летучих мышей была определена в первую очередь генетическими данными, поскольку летопись окаменелостей этого семейства является наиболее фрагментированной из всех летучих мышей. Вероятно, они эволюционировали в Австралазии , где общий предок всех ныне живущих птероподидов существовал примерно 31 миллион лет назад. Многие из их линий, вероятно, возникли в Меланезии , а затем со временем распространились по материковой Азии, Средиземноморью и Африке . Сегодня они встречаются в тропических и субтропических районах Евразии, Африки и Океании .
В семейство летучих мышей входят самые крупные виды летучих мышей: особи некоторых видов весят до 1,45 кг (3,2 фунта) и имеют размах крыльев до 1,7 м (5,6 футов). Не все летучие мыши крупного размера; почти треть всех видов весят менее 50 г (1,8 унции). Их можно отличить от других летучих мышей по собачьим мордам, когтистым вторым пальцам и редуцированному уропатагию . У небольшого числа видов есть хвосты. У мегалетучих мышей есть несколько приспособлений для полета, включая быстрое потребление кислорода, способность поддерживать частоту сердечных сокращений более 700 ударов в минуту и большой объем легких.
Большинство летучих мышей ведут ночной или сумеречный образ жизни , хотя некоторые виды активны в дневное время. В период бездействия они ночуют на деревьях или в пещерах. Представители некоторых видов живут поодиночке, а другие образуют колонии численностью до миллиона особей. В период активности они используют полет для перемещения к пищевым ресурсам. За редким исключением, они не способны эхолокировать , вместо этого полагаясь на острое зрение и обоняние, чтобы ориентироваться и находить пищу. Большинство видов преимущественно плодоядны , а некоторые нектароядны . Другие менее распространенные пищевые ресурсы включают листья, пыльцу, ветки и кору.
Они медленно достигают половой зрелости и имеют низкую репродуктивную способность. Большинство видов рожают по одному потомству после четырех-шестимесячной беременности. Такая низкая репродуктивная способность означает, что после сокращения популяции их численность восстанавливается медленно. Четверть всех видов занесена в список находящихся под угрозой исчезновения , в основном из-за разрушения среды обитания и чрезмерной охоты . В некоторых регионах мегалетучие мыши являются популярным источником пищи, что приводит к сокращению популяции и вымиранию. Они также представляют интерес для тех, кто занимается общественным здравоохранением , поскольку являются естественными резервуарами нескольких вирусов, которые могут поражать человека.
Таксономия и эволюция
[ редактировать ]Таксономическая история
[ редактировать ]



| |||
| Внутренние взаимоотношения африканских Pteropodidae основаны на совокупности данных о митохондриальной и ядерной ДНК. были включены по одному виду Pteropodinae, Nyctimeninae и Cynopterinae, которые не встречаются в Африке В качестве внешних групп . [ 2 ] |
Семейство Pteropodidae было впервые описано в 1821 году британским зоологом Джоном Эдвардом Греем . Он назвал семейство «Pteropidae» (в честь рода Pteropus ) и поместил его в ныне несуществующий отряд Fructivorae. [ 3 ] Fructivorae содержала еще одно семейство, ныне несуществующее Cephalotidae, содержащее один род Cephalotes. [ 3 ] (ныне признан синонимом Добсонии ). [ 4 ] Правописание Грея, возможно, было основано на неправильном понимании суффикса « Птеропус ». [ 5 ] Слово « Птеропус » происходит от древнегреческого слова «pterón», что означает «крыло», и «poús», что означает «нога». [ 6 ] Греческое слово pous от Pteropus происходит от корня слова pod- ; следовательно, латинизация Pteropus правильно приводит к образованию префикса « Pteropod- ». [ 7 ] : 230 Французский биолог Шарль Люсьен Бонапарт был первым, кто использовал исправленное написание Pteropodidae в 1838 году. [ 7 ] : 230
В 1875 году зоолог Джордж Эдвард Добсон был первым, кто разделил отряд рукокрылых (летучих мышей) на два подотряда : Megachiroptera (иногда упоминаемый как Macrochiroptera ) и Microchiroptera , которые обычно сокращают до megachiroptera и microbats. [ 8 ] Добсон выбрал эти названия, чтобы намекнуть на различия в размерах тела двух групп: многие летучие мыши, питающиеся фруктами, крупнее летучих мышей, питающихся насекомыми. Pteropodidae были единственным семейством, которое он включил в состав Megachiroptera. [ 5 ] [ 8 ]
Исследование 2001 года показало, что дихотомия летучих мышей и летучих мышей неточно отражает их эволюционные взаимоотношения. Вместо Megachiroptera и Microchiroptera авторы исследования предложили новые подотряды Yinpterochiroptera и Yangochiroptera . [ 9 ] Эта схема классификации впоследствии была проверена несколько раз и по-прежнему широко поддерживается по состоянию на 2019 год. [ 10 ] [ 11 ] [ 12 ] [ 13 ] С 2005 года этот подотряд альтернативно называют «Pteropodiformes». [ 7 ] : 520–521 Yinpterochiroptera содержала виды, ранее входившие в состав Megachiroptera (все из Pteropodidae), а также несколько семейств, ранее входивших в состав Microchiroptera: Megadermatidae , Rhinolophidae , Nycteridae , Craseonycteridae и Rhinopomatidae . [ 9 ] Два надсемейства включают Yinpterochiroptera: Rhinolophoidea, содержащее вышеуказанные семейства, ранее входившие в состав Microchiroptera, и Pteropodoidea, которое содержит только Pteropodidae. [ 14 ]
В 1917 году датский маммолог Кнуд Андерсен разделил Pteropodidae на три подсемейства: Macroglossinae, Pteropinae (исправлено на Pteropodinae ) и Harpyionycterinae. [ 15 ] : 496 Исследование 1995 года показало, что Macroglossinae, определенный ранее, содержащий роды Eonycteris , Notopteris , Macroglossus , Syconycteris , Melonycteris и Megaloglossus , был парафилетическим , то есть подсемейство не объединяло всех потомков общего предка. [ 16 ] : 214 Последующие публикации рассматривают Macroglossini как трибу внутри Pteropodinae, содержащую только Macroglossus и Syconycteris . [ 17 ] [ 18 ] Eonycteris и Melonycteris входят в состав других триб Pteropodinae. [ 2 ] [ 18 ] Megaloglossus был отнесен к трибе Myonycterini подсемейства Rousettinae, а Notopteris имеет неопределенное местонахождение. [ 18 ]
Другие подсемейства и трибы Pteropodidae также претерпели изменения после публикации Андерсена в 1917 году. [ 18 ] В 1997 году птероподиды были разделены на шесть подсемейств и девять триб на основе их морфологии или физических характеристик. [ 18 ] Генетическое исследование 2011 года пришло к выводу, что некоторые из этих подсемейств были парафилетическими и поэтому не отражали точно взаимоотношения между видами летучих мышей. Поддержку получили три из подсемейств, предложенных в 1997 г. на основе морфологии: Cynopterinae, Harpyionycterinae и Nyctimeninae. Остальные три клады , обнаруженные в этом исследовании, включали Macroglossini, Epomophorinae + Rousettini и Pteropodini + Melonycteris . [ 18 ] Генетическое исследование 2016 года, сосредоточенное только на африканских птероподидах (Harpyionycterinae, Rousettinae и Epomophorinae), также поставило под сомнение классификацию 1997 года. Все виды, ранее входившие в Epomophorinae, были перенесены в Rousettinae, который был разделен на дополнительные трибы. Род Eidolon , ранее входивший в трибу Rousettini из Rousettinae, был перенесен в собственное подсемейство Eidolinae . [ 2 ]
В 1984 году было предложено дополнительное подсемейство птероподид, Propottininae, представляющее один вымерший вид, описанный по окаменелости, обнаруженной в Африке, Propottoleakeyi . [ 19 ] В 2018 году окаменелости были повторно исследованы и установлено, что они представляют собой лемура . [ 20 ] По состоянию на 2018 год описано 197 видов летучих мышей. [ 21 ] около трети из них — летучие лисицы рода Pteropus . [ 22 ]
Эволюционная история
[ редактировать ]Ископаемые рекорды и времена расхождения
[ редактировать ]Летопись окаменелостей крылоногих летучих мышей является самой неполной из всех семейств летучих мышей. Хотя плохие данные о скелете рукокрылых, вероятно, объясняются хрупкостью скелетов летучих мышей, у Pteropodidae все еще самый неполный скелет, несмотря на то, что они обычно имеют самые большие и крепкие скелеты. Удивительно также, что Pteropodidae представлены меньше всего, поскольку они были первой крупной группой, которая разошлась. [ 23 ] Несколько факторов могут объяснить, почему было обнаружено так мало окаменелостей крылоногих: тропические регионы, где могут быть найдены их окаменелости, недостаточно отобраны по сравнению с Европой и Северной Америкой; В тропиках условия для окаменения плохие, что может привести к уменьшению количества окаменелостей в целом; и даже когда окаменелости образуются, они могут быть уничтожены последующей геологической деятельностью. [ 24 ] По оценкам, более 98% истории окаменелостей птероподид отсутствует. [ 25 ] Даже без окаменелостей возраст и время дивергенции семейства можно оценить с помощью компьютерной филогенетики . Pteropodidae отделились от надсемейства Rhinolophoidea (которое включает все остальные семейства подотряда Yinpterochiroptera) примерно 58 млн лет назад (миллионы лет назад). [ 25 ] Предок кронной группы Pteropodidae, или всех ныне живущих видов, жил примерно 31 млн лет назад. [ 26 ]
Биогеография
[ редактировать ]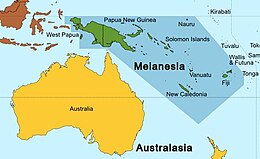
семейство Pteropodidae, вероятно, возникло в Австралазии Судя по биогеографическим реконструкциям, . [ 2 ] Другие биогеографические анализы показали, что Меланезийские острова, включая Новую Гвинею , являются вероятным кандидатом на происхождение большинства подсемейств крупных летучих мышей, за исключением Cynopterinae; [ 18 ] циноптерины, вероятно, возникли на Зондском шельфе . Судя по результатам взвешенного анализа предковой области шести ядерных и митохондриальных генов, [ 26 ] Из этих регионов птероподиды колонизировали другие территории, включая континентальную Азию и Африку. Мегабаты достигли Африки как минимум в четырех различных событиях. Согласно исследованию 2016 года , четыре предложенных вида представлены (1) Scotonycteris , (2) Rousettus , (3) Scotonycterini и (4) «эндемической африканской кладой», в которую входят Stenonycterini, Plerotini, Myonycterini и Epomophorini. Неизвестно, когда летучие мыши достигли Африки, но к позднему миоцену там присутствовало несколько племен (Scotonycterini, Stenonycterini, Plerotini, Myonycterini и Epomophorini) . Как мегалетучие мыши попали в Африку, также неизвестно. Было высказано предположение, что они могли прийти через Ближний Восток до того, как в конце миоцена он стал более засушливым. И наоборот, они могли попасть на континент через сухопутный мост Гомфотерий , который соединял Африку и Аравийский полуостров с Евразией . Предполагается, что род Pteropus (летучие лисицы), который не встречается на материковой части Африки, расселился из Меланезии в результате прыжков по островам. через Индийский океан ; [ 27 ] это менее вероятно для других родов летучих мышей, которые имеют меньшие размеры тела и, следовательно, имеют более ограниченные возможности полета. [ 2 ]
Эхолокация
[ редактировать ]Мегабаты — единственное семейство летучих мышей, неспособное к гортанной эхолокации. Неясно, был ли общий предок всех летучих мышей способен к эхолокации, и, таким образом, эхолокация была потеряна в линии мегалетучих мышей, или несколько линий летучих мышей независимо развили способность к эхолокации (надсемейство Rhinolophoidea и подотряд Yangochiroptera ). Этот неизвестный элемент эволюции летучих мышей был назван «большим вызовом биологии». [ 28 ] Исследование онтогенеза (эмбрионального развития) летучих мышей, проведенное в 2017 году, выявило доказательства того, что эмбрионы мегабатов сначала имеют большую развитую улитку , похожую на эхолокационную улитку летучих мышей, хотя при рождении у них есть маленькая улитка, похожая на неэхолокирующие млекопитающие. Эти данные подтверждают, что гортанная эхолокация однажды возникла у летучих мышей и была потеряна у крылоподид, а не развивалась дважды независимо. [ 29 ] Летучие мыши рода Rousettus способны к примитивной эхолокации посредством щелкания языком. [ 30 ] Некоторые виды — пещерная нектарная летучая мышь ( Eonycteris spelaea ), малая коротконосая фруктовая летучая мышь ( Cynopterus brachyotis ) и длинноязычная фруктовая летучая мышь ( Maroglossus sobrinus ) — как было показано, издают щелчки, похожие на щелчки летучих мышей, выполняющих эхолокацию, используя свои крылья. . [ 31 ]
И эхолокация, и полет — энергетически затратные процессы. [ 32 ] Летучие мыши, использующие эхолокацию, сочетают производство звука с механизмами полета, что позволяет им снизить дополнительную энергетическую нагрузку, связанную с эхолокацией. Вместо того, чтобы создавать давление на поток воздуха для производства звука, летучие мыши, владеющие гортанной эхолокацией, вероятно, используют силу ударов своих крыльев для создания давления в воздухе, сокращая энергетические затраты за счет синхронизации взмахов крыльев и эхолокации. [ 33 ] Утрата эхолокации (или, наоборот, отсутствие ее эволюции) может быть связана с разобщением полета и эхолокации у крупных летучих мышей. [ 34 ] Более крупный средний размер тела летучих мышей по сравнению с летучими мышами, обладающими эхолокацией. [ 35 ] предполагает, что больший размер тела нарушает связь полета и эхолокации и делает эхолокацию слишком энергетически дорогой, чтобы ее можно было сохранить у мегалетучих мышей. [ 34 ]
Список родов
[ редактировать ]



Семейство Pteropodidae делится на шесть подсемейств, представленных 46 родами : [ 2 ] [ 18 ]
Семейство Pteropodidae
- подсемейство Cynopterinae [ 18 ]
- род Aethalops - карликовые фруктовые летучие мыши.
- род Alionycteris
- род Balionycteris
- род Хиронакс
- род Cynopterus - фруктовые летучие мыши с собачьей мордой или фруктовые летучие мыши с коротким носом.
- род Dyacopterus - летучие мыши даяки.
- род Гаплониктерис
- род Лайденс
- род Мегаеропс
- род Отоптерус
- Род Пентетор
- род Ptenochirus - мускусные фруктовые летучие мыши.
- род Сфериас
- род Тооптерус
- подсемейство Eidolinae [ 2 ]
- род Eidolon - летучие мыши соломенного цвета.
- подсемейство Harpyionycterinae [ 2 ]
- род Апротелес
- род Бонея
- род Dobsonia - летучие мыши с голой спиной.
- род Гарпииониктер
- подсемейство Nyctimeninae [ 18 ]
- род Nyctimene - трубконосые летучие мыши.
- род Paranyctimene
- подсемейство Pteropodinae
- род Melonycteris [ 18 ]
- племя Птероподини [ 18 ]
- род Ацеродон
- род Птералопекс
- род Pteropus - летучие лисицы.
- род Styloctenium
- подсемейство Rousettinae
- племя Эониктерини [ 2 ]
- род Eonycteris - летучие мыши-рассветные плодовые летучие мыши.
- племя Эпомофорини [ 2 ] [ 18 ]
- род Epomophorus - фруктовые летучие мыши с погонами.
- род Epomops - летучие мыши с погонами.
- род Hypsignathus
- род Micropteropus - карликовые летучие мыши с погонами.
- род Нанониктерис
- племя неопределенного местонахождения [ 36 ]
- род Pilonycteris
- племя Миониктерини [ 2 ]
- род Megaloglossus
- род Myonycteris - маленькие фруктовые летучие мыши с ошейником.
- племя Плеротини [ 2 ]
- род Плеротес
- племя Русеттини [ 2 ]
- род Rousettus - летучие мыши-розетки.
- племя скотониктерини [ 2 ]
- род Casinycteris
- род Scotonycteris
- племя Стенониктерини [ 2 ]
- род Стенониктерис
- племя Эониктерини [ 2 ]
- Неуверенное сиденье
- род Notopteris - длиннохвостые летучие мыши. [ 18 ]
- род Массаж [ 18 ]
- род Неоптерикс [ 18 ]
- род Десмалопекс [ 18 ]
- род † Turkanycteris [ 37 ]
- племя Макроглоссини [ 18 ]
- род Macroglossus - длинноязычные фруктовые летучие мыши.
- род Syconycteris - цветущие летучие мыши.
Описание
[ редактировать ]Появление
[ редактировать ]
Мегалетучие мыши получили свое название из-за большего веса и размера; самый крупный, большая летучая лисица ( Pteropus neohibernicus ), весит до 1,6 кг (3,5 фунта); [ 38 ] у некоторых представителей Acerodon и Pteropus размах крыльев достигает 1,7 м (5,6 футов). [ 39 ] : 48 Несмотря на то, что размер тела был определяющей характеристикой, которую Добсон использовал для разделения летучих мышей и летучих мышей, не все виды летучих мышей крупнее летучих мышей; Пятнистокрылая плодовая летучая мышь ( Balionycteris maculata ), летучая мышь-гигант, весит всего 14,2 г (0,50 унции). [ 35 ] Летучих лисиц Pteropus и Acerodon часто принимают за образцы всего семейства по размерам тела. На самом деле эти роды являются выдающимися, что создает неправильное представление об истинном размере большинства видов крупных летучих мышей. [ 5 ] Обзор 2004 года показал, что 28% видов летучих мышей весят менее 50 г (1,8 унции). [ 35 ]
Мегабатов можно отличить от микролетучих мышей по внешнему виду по собачьим мордам, наличию когтей на втором пальце (см. Megabat#Postcrania ) и простым ушам. [ 40 ] Простой внешний вид уха частично объясняется отсутствием траги (хрящевых створок, выступающих перед ушным каналом), которые встречаются у многих видов летучих мышей. Летучие мыши рода Nyctimene менее похожи на собак, с более короткими мордами и трубчатыми ноздрями. [ 41 ] Исследование 167 видов летучих мышей, проведенное в 2011 году, показало, что, хотя большинство (63%) имеют мех однородного цвета, в этом семействе наблюдаются и другие узоры. К ним относятся обратная тень у четырех процентов видов, полоса на шее или мантия у пяти процентов видов, полосы у десяти процентов видов и пятна у девятнадцати процентов видов. [ 42 ]
В отличие от летучих мышей, у мегалетучих мышей значительно уменьшена уропатагиум , представляющая собой участок летательной мембраны, проходящий между задними конечностями. [ 43 ] Кроме того, хвост отсутствует или сильно редуцирован, [ 41 ] за исключением видов Notopteris , у которых длинный хвост. [ 44 ] У большинства крыльев мегабата вставляются сбоку (прикрепляются к телу непосредственно по бокам). У видов Dobsonia крылья прикрепляются ближе к позвоночнику, что дало им общее название фруктовых летучих мышей с «голой спиной» или «голой спиной». [ 43 ]
Скелет
[ редактировать ]Череп и зубной ряд
[ редактировать ]
Мегабаты имеют большие орбиты , которые сзади окаймлены хорошо развитыми заглазничными отростками . Заглазничные отростки иногда объединяются, образуя заглазничную перекладину . Морда . простая на вид и не сильно видоизмененная, как у других семейств летучих мышей [ 45 ] Длина морды варьируется у разных родов. хорошо Предчелюстная кость развита и обычно свободна. [ 4 ] это означает, что он не сросся с верхней челюстью ; вместо этого он соединяется с верхней челюстью посредством связок , что делает ее свободно подвижной. [ 46 ] [ 47 ] Предчелюстная кость всегда лишена небной ветви. [ 4 ] У видов с более длинной мордой череп обычно изогнут. У родов с более короткими лицами ( Penthetor , Nyctimene , Dobsonia и Myonycteris ) череп практически не имеет изгиба. [ 48 ]
Количество зубов варьируется у разных видов летучих мышей; общее количество для различных видов колеблется от 24 до 34. Все летучие мыши-гиганты имеют по два или четыре верхних и нижних резца , за исключением плодовой летучей мыши Балмера ( Aproteles bulmerae ), у которой резцы полностью отсутствуют, [ 49 ] и летучая мышь Сан-Томе с воротником ( Myonycteris brachycephala ), у которой есть два верхних и три нижних резца. [ 50 ] Это делает его единственным видом млекопитающих с асимметричной зубной формулой . [ 50 ]
Все виды имеют по два верхних и нижних клыка . Количество премоляров варьируется: по четыре или шесть верхних и нижних премоляров. Первые верхние и нижние коренные зубы всегда присутствуют, а это означает, что у всех летучих мышей есть как минимум четыре коренных зуба. Остальные моляры могут присутствовать, присутствовать, но редуцироваться или отсутствовать. [ 49 ] и гребни уменьшаются, Моляры и премоляры Megabat упрощаются, бугры что приводит к более уплощенной коронке . [ 51 ]
Как и большинство млекопитающих, летучие мыши-гиганты являются дифиодонтами , что означает, что у молодых особей есть набор молочных зубов (молочных зубов), которые выпадают и заменяются постоянными зубами. У большинства видов имеется 20 молочных зубов. Что характерно для млекопитающих, [ 52 ] молочный набор не включает моляры. [ 51 ]
Посткрания
[ редактировать ]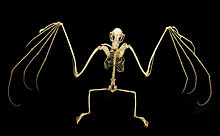
Лопатки . (лопатки) летучих мышей описаны как самые примитивные из всех семейств рукокрылых [ 51 ] Плечо в целом имеет простую конструкцию, но имеет некоторые особенности. Примитивное прикрепление подъязычной мышцы от ключицы (ключицы) к лопатке смещено латерально (больше в сторону тела) — особенность, также наблюдаемая у Phyllostomidae . Плечо также имеет хорошо развитую систему мышечных выступов (узких полосок мышц, которые увеличивают более крупные мышцы), которые закрепляют сухожилие затылочно-полликальной мышцы (мышца у летучих мышей, идущая от основания шеи до основания большого пальца). [ 43 ] к коже. [ 41 ]
В то время как у летучих мышей когти есть только на больших пальцах передних конечностей, у большинства летучих мышей есть также когтистый второй палец; [ 51 ] только у Eonycteris , Dobsonia , Notopteris и Neopteryx отсутствует второй коготь. [ 53 ] Первая цифра самая короткая, а третья цифра самая длинная. Второй палец не способен сгибаться . [ 51 ] Большие пальцы рук летучих мышей длиннее передних конечностей, чем у летучих мышей. [ 43 ]
Задние конечности мегалетучих мышей имеют те же компоненты скелета, что и люди. У большинства видов летучих мышей есть дополнительная структура, называемая калькаром , хрящевая шпора, возникающая из пяточной кости . [ 54 ] Некоторые авторы поочередно называют эту структуру уропатагиальной шпорой, чтобы отличить ее от калькаров летучих мышей, которые устроены иначе. Структура существует для стабилизации уропатагия, позволяя летучим мышам регулировать выпуклость мембраны во время полета. К летучим мышам, лишенным калькара или шпоры, относятся Notopteris , Syconycteris и Harpyionycteris . [ 55 ] Вся нога повернута в бедре по сравнению с нормальной ориентацией млекопитающих, что означает, что колени обращены назад . Все пять пальцев стопы сгибаются в направлении сагиттальной плоскости , при этом ни один палец не может сгибаться в противоположном направлении, как в стопах сидящих птиц. [ 54 ]
Внутренние системы
[ редактировать ]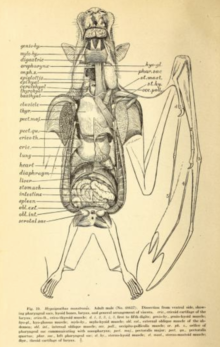
Полет очень энергетически затратен и требует некоторой адаптации сердечно-сосудистой системы . Во время полета летучие мыши могут увеличивать потребление кислорода в двадцать и более раз на продолжительный период времени; люди-спортсмены могут добиться увеличения в двадцать раз максимум за несколько минут. [ 56 ] Исследование, проведенное в 1994 году на летучей мыши соломенного цвета ( Eidolon helvum ) и летучей мыши-молоте ( Hypsignathus monstrosus ), показало, что средний коэффициент дыхательного обмена (вырабатываемый углекислый газ:используемый кислород) составляет примерно 0,78. Среди этих двух видов, седой летучей лисицы ( Pteropus poliocephalus ) и египетской плодовой летучей мыши ( Rousettus aegyptiacus ), максимальная частота сердечных сокращений в полете варьировала от 476 ударов в минуту (сероголовая летучая лисица) до 728 ударов в минуту (египетская летучая лисица). фруктовая летучая мышь). Максимальное число вдохов в минуту колебалось от 163 (сероголовая летучая лисица) до 316 (соломенная летучая мышь). [ 57 ] Кроме того, летучие мыши-гиганты имеют исключительно большой объем легких по сравнению с их размерами. В то время как у наземных млекопитающих, таких как землеройки, объем легких составляет 0,03 см3. 3 на грамм массы тела (0,05 дюйма 3 на унцию массы тела), такие виды, как фруктовая летучая мышь с погонами Уолберга ( Epomophorus wahlbergi ), имеют объем легких в 4,3 раза больше - 0,13 см. 3 за грамм (0,22 дюйма 3 за унцию). [ 56 ]
У мегалетучих мышей пищеварительная система быстрая, время прохождения через кишечник составляет полчаса или меньше. [ 41 ] Пищеварительная система построена на травоядной диете, иногда ограничивающейся мягкими фруктами или нектаром. [ 58 ] Длина пищеварительной системы у травоядных коротка (как и у насекомоядных микрорукокрылых), [ 58 ] поскольку волокнистое содержимое в основном отделяется под действием неба, языка и зубов, а затем выбрасывается. [ 58 ] У многих летучих мышей желудок U-образной формы. нет Четкой разницы между тонкой и толстой кишкой и четко выраженного начала прямой кишки . У них очень высокая плотность кишечных микроворсинок , что создает большую площадь поверхности для всасывания питательных веществ. [ 59 ]
Биология и экология
[ редактировать ]Размер генома
[ редактировать ]Как и все летучие мыши, мегалетучие мыши имеют гораздо меньшие геномы , чем другие млекопитающие. Исследование 43 видов летучих мышей, проведенное в 2009 году, показало, что их геномы варьировались от 1,86 пикограмм (пг, 978 Мбит на пг) у летучей мыши соломенного цвета до 2,51 пг у летучей лисицы Лайла ( Pteropus lylei ). Все значения были намного ниже среднего показателя для млекопитающих (3,5 пг). Геномы мегалетучих мышей даже меньшие, чем у микролетучих мышей: средний вес составляет 2,20 пг по сравнению с 2,58 пг. Было высказано предположение, что это различие может быть связано с тем фактом, что в линии мегалетучих мышей произошел вымирание LINE1 — типа долго рассеянных ядерных элементов . LINE1 составляет 15–20% генома человека и считается наиболее распространенным ядерным элементом с длинными вкраплениями среди млекопитающих. [ 60 ]
Чувства
[ редактировать ]Взгляд
[ редактировать ]
За очень немногими исключениями, летучие мыши-гиганты не обладают эхолокацией и поэтому для навигации полагаются на зрение и обоняние. [ 61 ] У них большие глаза, расположенные на передней части головы. [ 62 ] Они крупнее, чем у общего предка всех летучих мышей, при этом одно исследование предполагает тенденцию увеличения размера глаз среди птероподид. Исследование, в котором изучались глаза 18 видов летучих мышей, показало, что обыкновенная летучая мышь ( Syconycteris australis ) имела самые маленькие глаза диаметром 5,03 мм (0,198 дюйма), а самые большие глаза были у большой летучей лисицы ( Pteropus vampyrus ). Диаметр 12,34 мм (0,486 дюйма). [ 63 ] Ирисы Megabat обычно коричневые, но могут быть красными или оранжевыми, как у Desmalopex , Mirimiri , Pteralopex и некоторых Pteropus . [ 64 ]
При высоких уровнях яркости острота зрения у мегабатов хуже, чем у людей; при низкой яркости он превосходен. [ 62 ] Одно исследование, в котором изучались глаза некоторых видов Rousettus , Epomophorus , Eidolon и Pteropus , показало, что первые три рода обладают Tapetum lucidum , отражающей структурой в глазах, которая улучшает зрение при низких уровнях освещенности, в то время как виды Pteropus этого не делают. [ 61 ] У всех исследованных видов сетчатка имела как палочки, так и колбочки , но только у видов Pteropus были S-колбочки, которые улавливают самые короткие длины волн света; поскольку спектральная настройка опсинов не была различима, неясно, обнаруживают ли S-конусы видов Pteropus синий или ультрафиолетовый свет. Pteropus Летучие мыши двухцветны и имеют два типа колбочек. Остальные три рода из-за отсутствия S-конусов одноцветны и не способны различать цвета. Все роды имели очень высокую плотность палочек, что приводило к высокой чувствительности к свету, что соответствует их ночной активности. У Pteropus и Rousettus измеренная плотность палочек составляла 350 000–800 000 на квадратный миллиметр, что равняется или превосходит других животных, ведущих ночной или сумеречный образ жизни , таких как домашняя мышь , домашняя кошка и домашний кролик . [ 61 ]
Запах
[ редактировать ]
Мегалетучие мыши используют запах, чтобы найти источники пищи, такие как фрукты и нектар. [ 65 ] У них острое обоняние, которое может соперничать с обонянием домашней собаки . [ 66 ] Плодовые летучие мыши с трубчатым носом, такие как восточная трубконосая летучая мышь ( Nyctimene robinsoni ), обладают стереообонянием , то есть они способны отображать и отслеживать шлейфы запахов в трехмерном виде. [ 66 ] Наряду с большинством (или, возможно, всеми) другими видами летучих мышей, матери и потомство мегалетучих мышей также используют запах, чтобы узнавать друг друга, а также для распознавания особей. [ 65 ] увеличены андрогенам чувствительные к сальные железы У летучих лисиц у самцов на плечах , которые они используют для обозначения запахами своей территории, особенно во время брачного сезона. Секреция этих желез различается в зависимости от вида — из 65 химических соединений, выделенных из желез четырех видов, ни одно соединение не обнаружено у всех видов. [ 67 ] Самцы также моются мочой или обмазываются собственной мочой. [ 67 ] [ 68 ]
Вкус
[ редактировать ]Летучие мыши-гиганты обладают геном TAS1R2 , что означает, что они способны определять сладость продуктов. Этот ген присутствует у всех летучих мышей, кроме летучих мышей-вампиров . Как и все другие летучие мыши, летучие мыши-гиганты не могут ощущать вкус умами из-за отсутствия гена TAS1R1 . Было показано , что среди других млекопитающих только у гигантских панд этот ген отсутствует. [ 65 ] У мегалетучих мышей также есть несколько генов TAS2R , что указывает на то, что они чувствуют горечь. [ 69 ]
Размножение и жизненный цикл
[ редактировать ]
Мегабаты, как и все летучие мыши, живут долго относительно своего размера для млекопитающих. Продолжительность жизни некоторых летучих мышей в неволе превышает тридцать лет. [ 53 ] По сравнению с их размерами, летучие мыши-гиганты имеют низкую репродуктивную способность и задержку полового созревания, при этом самки большинства видов не рожают до возраста одного или двух лет. [ 70 ] : 6 Некоторые летучие мыши, по-видимому, способны размножаться в течение всего года, но большинство видов, вероятно, размножаются сезонно . [ 53 ] Спаривание происходит на насесте. [ 71 ] Продолжительность беременности варьируется, [ 72 ] но у большинства видов составляет от четырех до шести месяцев. У разных видов летучих мышей есть репродуктивные приспособления, которые удлиняют период между совокуплением и родами. Некоторые виды, такие как летучая мышь соломенного цвета, имеют репродуктивную адаптацию отсроченной имплантации , что означает, что совокупление происходит в июне или июле, но зигота не имплантируется в стенку матки до тех пор, пока несколько месяцев спустя, в ноябре. [ 70 ] : 6 Карликовая плодовая летучая мышь Фишера ( Haplonycteris fischeri ) с адаптацией к постимплантационной задержке имеет самую большую продолжительность беременности среди всех видов летучих мышей - до 11,5 месяцев. [ 72 ] Постимплантационная задержка означает, что развитие эмбриона приостанавливается на срок до восьми месяцев после имплантации в стенку матки, что является причиной его очень длительной беременности. [ 70 ] : 6 Более короткая продолжительность беременности обнаружена у большой коротконосой летучей мыши ( Cynopterus sphinx ) с периодом в три месяца. [ 73 ]
Размер помета у всех летучих мышей обычно один. [ 70 ] : 6 Немногочисленны находки близнецов у следующих видов: мадагаскарская летучая лисица ( Pteropus rufus ), погонная летучая мышь Добсона ( Epomops dobsoni ), седая летучая лисица, черная летучая лисица ( Pteropus alecto ), очковая летучая лисица ( Pteropus ). конспициллатус ), [ 74 ] большая коротконосая фруктовая летучая мышь, [ 75 ] Летучая мышь Петерса с погонами ( Epomophorus crypturus ), летучая мышь-молот, соломенная летучая мышь, маленькая фруктовая летучая мышь с воротником ( Myonycteris torquata ), египетская фруктовая летучая мышь и розетка Лешено ( Rousettus leschenaultii ). [ 76 ] : 85–87 В случае близнецов оба потомка выживают редко. [ 74 ] Поскольку у мегалетучих мышей, как и у всех летучих мышей, низкая репродуктивная способность, их популяция медленно восстанавливается после сокращения. [ 77 ]
При рождении потомство мегабата составляет в среднем 17,5% послеродового веса их матери. Это наименьшее соотношение потомства к матери для любого семейства летучих мышей; среди всех летучих мышей вес новорожденных составляет 22,3% послеродового веса их матери. Потомство мегалетучих мышей нелегко разделить на традиционные категории альтрициальных (беспомощных при рождении) или преждевременных (способных к рождению). Такие виды, как большая коротконосая летучая мышь, рождаются с открытыми глазами (признак преждевременного развития потомства), тогда как у потомства египетской летучей мыши глаза открываются только через девять дней после рождения (признак альтрициального потомства). [ 78 ]
Как и почти все виды летучих мышей, самцы не помогают самкам в родительской заботе. [ 79 ] Детеныши остаются со своими матерями до тех пор, пока их не отнимут от груди ; продолжительность отлучения от груди варьируется в зависимости от семьи. У мегалетучих мышей, как и у всех летучих мышей, период кормления относительно длительный: потомство будет кормить грудью до тех пор, пока его масса тела не достигнет примерно 71% массы тела взрослой особи по сравнению с 40% массы тела взрослой особи у млекопитающих, не являющихся летучими мышами. [ 80 ] Виды рода Micropteropus отлучают детенышей от груди в возрасте семи-восьми недель, тогда как индийская летучая лисица ( Pteropus medius ) отлучает детенышей только в возрасте пяти месяцев. [ 76 ] Очень необычно, что самцы двух видов летучих мышей, летучей лисицы в маске Бисмарка ( Pteropus capistratus ) и плодовой летучей мыши даяка ( Dyacopterus spadiceus ), были замечены производящими молоко , но никогда не наблюдалось наблюдения за самцами, выкармливающими детенышей. [ 81 ] Неясно, является ли лактация функциональной и самцы действительно выкармливают детенышей, или это результат стресса или недоедания . [ 82 ]
Поведение и социальные системы
[ редактировать ]
Многие виды летучих мышей очень стадны и социальны. Мегалетучие мыши будут издавать звуки, чтобы общаться друг с другом, создавая звуки, описываемые как «звуковые всплески, похожие на трели». [ 83 ] сигналит, [ 84 ] или громкие, похожие на блеяние звонки [ 85 ] в различных родах. По крайней мере, один вид, египетская фруктовая летучая мышь, способен к вокальному обучению , называемому обучением производству голоса, которое определяется как «способность изменять вокализацию в ответ на взаимодействие с представителями своего вида». [ 86 ] [ 87 ] Молодые египетские летучие мыши способны усваивать диалект , слушая своих матерей, а также других особей в своих колониях. Было высказано предположение, что эти диалектные различия могут привести, например, к тому, что люди из разных колоний общаются с разной частотой. [ 88 ] [ 89 ]
Социальное поведение мегабатов включает в себя использование сексуального поведения не только для размножения. Имеющиеся данные свидетельствуют о том, что самки египетских летучих мышей отбирают пищу у самцов в обмен на секс. Тесты на отцовство подтвердили, что самцы, у которых каждая самка добывала еду, имели большую вероятность стать отцом потомства этой самки. [ 90 ] Гомосексуальная фелляция наблюдалась по крайней мере у одного вида — летучей лисицы Бонина ( Pteropus pselaphon ). [ 91 ] [ 92 ] Предполагается, что этот однополый минет способствует образованию колоний самцов-антагонистов в более холодном климате. [ 91 ] [ 92 ]
Летучие мыши-гиганты в основном ведут ночной образ жизни и ведут сумеречный образ жизни , хотя некоторые из них были замечены в полете в течение дня. [ 39 ] Некоторые островные виды и подвиды ведут дневной образ жизни , что предположительно является ответом на отсутствие хищников . Дневные таксоны включают подвид черноухой летучей лисицы ( Pteropus melanotus natalis ), маврикийскую летучую лисицу ( Pteropus niger ), каролинскую летучую лисицу ( Pteropus molossinus ), подвид Pteropus pelagicus ( P. p. insularis ) и Сейшельская летучая мышь ( Pteropus seychellensis ). [ 93 ] : 9
Ночевка
[ редактировать ]В обзоре сорока одного рода летучих мышей за 1992 год отмечено, что двадцать девять родов живут на деревьях. Еще одиннадцать родов гнездятся в пещерах, а остальные шесть родов — в других местах (например, в человеческих постройках, шахтах и расщелинах). Виды, гнездящиеся на деревьях, могут быть одиночными или колониальными , образуя скопления численностью до одного миллиона особей. Пещерные виды образуют скопления от десяти особей до нескольких тысяч. Высококолониальные виды часто демонстрируют преданность ночлегу, а это означает, что их деревья или пещеры могут использоваться в качестве ночлегов в течение многих лет. Одиночные виды или те, которые собираются в меньшие количества, менее преданы своим местам обитания. [ 70 ] : 2
Диета и добыча пищи
[ редактировать ]
Большинство летучих мышей в основном плодоядны . [ 94 ] Члены всей семьи употребляют в пищу разнообразные фрукты почти 188 родов растений. [ 95 ] Некоторые виды также нектароядны , то есть они также пьют нектар из цветов. [ 94 ] В Австралии цветы эвкалипта являются особенно важным источником пищи. [ 41 ] Другие пищевые ресурсы включают листья, побеги, почки, пыльцу, семенные коробочки, сок, шишки, кору и ветки. [ 96 ] Они потрясающие едоки и могут съедать фруктов в 2,5 раза больше собственного веса за ночь. [ 95 ]
Мегалетучие мыши летают к местам ночлега и поиска пищи. Обычно они летают прямо и относительно быстро для летучих мышей; некоторые виды медленнее и обладают большей маневренностью. Виды могут преодолевать 20–50 км (12–31 миль) за ночь. Мигрирующие виды родов Eidolon , Pteropus , Epomophorus , Rousettus , Myonycteris и Nanonycteris могут мигрировать на расстояния до 750 км (470 миль). ниже среднего Большинство мегалетучих мышей имеют соотношение сторон . [ 97 ] Это измерение, связанное с размахом и площадью крыла. [ 97 ] : 348 Нагрузка на крыло, которая измеряет вес относительно площади крыла. [ 97 ] : 348 средний или выше среднего в мегабатах. [ 97 ]
Распространение семян
[ редактировать ]Мегабаты играют важную роль в распространении семян . В результате долгой истории эволюции некоторые растения развили характеристики, совместимые с органами чувств летучих мышей, в том числе плоды с сильным ароматом, яркой окраской и заметно выступающими вдали от листвы. Яркие цвета и расположение плодов могут отражать зависимость летучих мышей от визуальных сигналов и неспособность ориентироваться в беспорядке. В исследовании, в котором изучались плоды более сорока видов инжира, только один вид инжира потреблялся как птицами, так и летучими мышами; большинство видов потребляются тем или иным видом. Инжир, потребляемый птицами, часто бывает красным или оранжевым, а инжир, потребляемый летучими мышами, часто бывает желтым или зеленым. [ 98 ] Большинство семян выводятся из организма вскоре после употребления из-за быстрого прохождения через кишечник, но некоторые семена могут оставаться в кишечнике более двенадцати часов. Это повышает способность летучих мышей распространять семена далеко от родительских деревьев. [ 99 ] Будучи высокомобильными плодоядными животными, летучие мыши-гиганты способны восстанавливать лес между изолированными фрагментами леса, распространяя семена деревьев по обезлесенным ландшафтам. [ 100 ] Эта способность к распространению ограничена растениями с маленькими семенами длиной менее 4 мм (0,16 дюйма), поскольку семена большего размера не попадают в организм. [ 101 ]
Хищники и паразиты
[ редактировать ]
У мегалетучих мышей, особенно у тех, кто живет на островах, мало местных хищников: у таких видов, как маленькая летучая лисица ( Pteropus hypomelanus ), нет известных естественных хищников. [ 102 ] К неместным хищникам летучих лисиц относятся домашние кошки и крысы . Мангровый варан , который является местным хищником для некоторых видов летучих мышей, но интродуцированным хищником для других, оппортунистически охотится на летучих мышей, поскольку он умеет лазать по деревьям. [ 103 ] Другой вид, коричневая древесная змея , может серьезно повлиять на популяции летучих мышей; Будучи неместным хищником Гуама , змея поедает так много потомства, что это свело пополнение популяции марианской плодовой летучей мыши ( Pteropus mariannus ) практически к нулю. В настоящее время остров считается местом обитания марианской летучей мыши, поскольку его популяция зависит от летучих мышей, иммигрирующих с близлежащего острова Рота , которые поддерживают ее, а не от успешного размножения. [ 104 ] Хищники, которые естественным образом симпатизируют летучим мышам, включают рептилий, таких как крокодилы , змеи и крупные ящерицы, а также птиц, таких как соколы , ястребы и совы . [ 70 ] : 5 — Морской крокодил известный хищник летучих мышей, судя по анализу содержимого желудка крокодила в северной Австралии. [ 105 ] Во время экстремальной жары летучим мышам, таким как маленькая рыжая летучая лисица ( Pteropus scapulatus ), приходится охлаждаться и восстанавливать водный баланс, пья воду из водоемов, что делает их уязвимыми для оппортунистического нападения со стороны пресноводных крокодилов . [ 106 ]
Летучие мыши-гиганты являются хозяевами нескольких таксонов паразитов . Известные паразиты включают виды Nycteribiidae и Streblidae («летучие мыши»), [ 107 ] [ 108 ] а также клещи рода Demodex . [ 109 ] Паразиты крови семейства Haemoproteidae и кишечные нематоды Toxocaridae также поражают виды крупных летучих мышей. [ 41 ] [ 110 ]
Ареал и среда обитания
[ редактировать ]Мегабаты широко распространены в тропиках Старого Света , встречаются по всей Африке, Азии, Австралии, а также на всех островах Индийского океана и Океании . [ 18 ] По состоянию на 2013 год в Африке присутствует четырнадцать родов летучих мышей, представляющих двадцать восемь видов. Из этих двадцати восьми видов двадцать четыре встречаются только в тропическом или субтропическом климате. Остальные четыре вида в основном встречаются в тропиках, но их ареалы также охватывают умеренный климат . Что касается типов среды обитания, восемь из них встречаются исключительно или в основном в лесных средах обитания; девять встречаются как в лесах, так и в саваннах ; девять встречаются исключительно или в основном в саваннах; и два найдены на островах. Только один африканский вид, длинноволосая розетка ( Rousettus lanosus ), встречается в основном в горных экосистемах , но ареалы еще тринадцати видов простираются и в горную среду обитания. [ 111 ] : 226
За пределами Юго-Восточной Азии летучие мыши-гиганты имеют относительно небольшое видовое богатство в Азии. Египетская летучая мышь — единственная летучая мышь, ареал которой преимущественно простирается на территорию Палеарктики ; [ 112 ] это и соломенная летучая мышь — единственные виды, встречающиеся на Ближнем Востоке . [ 112 ] [ 113 ] Самая северная часть ареала египетской летучей мыши — северо-восточное Средиземноморье . [ 112 ] В Восточной Азии мегабаты встречаются только в Китае и Японии. В Китае только шесть видов летучих мышей считаются постоянными, а еще семь присутствуют незначительно (на границе ареала), сомнительно (из-за возможной ошибочной идентификации) или как случайные мигранты. [ 114 ] Четыре вида летучих мышей, все Pteropus , встречаются в Японии, но ни одного на ее пяти главных островах. [ 115 ] [ 116 ] [ 117 ] [ 118 ] В Южной Азии разнообразие видов летучих мышей колеблется от двух видов на Мальдивах до тринадцати видов в Индии . [ 119 ] Богатство видов мегабатов в Юго-Восточной Азии составляет всего пять видов в небольшой стране Сингапур и семьдесят шесть видов в Индонезии . [ 119 ] Из девяноста восьми видов летучих мышей, обитающих в Азии, лес является средой обитания для девяноста пяти из них. Другие типы среды обитания включают измененную человеком землю (66 видов), пещеры (23 вида), саванну (7 видов), кустарники (4 вида), скалистые местности (3 вида), луга (2 вида) и пустыни (1 вид). . [ 119 ]
В Австралии обитают пять родов и восемь видов летучих мышей. Это роды Pteropus , Syconycteris , Dobsonia , Nyctimene и Macroglossus . [ 41 ] : 3 Виды Pteropus Австралии встречаются в различных средах обитания, включая мангровых леса с преобладанием зарослей, тропические леса и влажные склерофилловые леса австралийского кустарника. [ 41 ] : 7 Австралийские Pteropus часто встречаются вместе с людьми, поскольку они размещают свои большие колонии в городских районах , особенно в мае и июне, когда наибольшая часть популяций видов Pteropus находится в этих городских колониях. [ 120 ]
В Океании в странах Палау и Тонга наименьшее количество видов летучих мышей - по одному на каждого. В Папуа-Новой Гвинее наибольшее количество видов - тридцать шесть. [ 121 ] Из шестидесяти пяти видов Океании лес является средой обитания пятидесяти восьми. Другие типы среды обитания включают измененную человеком землю (42 вида), пещеры (9 видов), саванну (5 видов), кустарники (3 вида) и скалистые местности (3 вида). [ 121 ] По оценкам, девятнадцать процентов всех видов летучих мышей являются эндемиками одного острова; Из всех семейств летучих мышей только Myzopodidae , включающие два вида, оба эндемики одного острова, имеет более высокий уровень эндемизма одного острова. [ 122 ]
Отношение к людям
[ редактировать ]Еда
[ редактировать ]Мегалетучих мышей убивают и едят как мясо диких животных по всему ареалу обитания. Летучих мышей широко потребляют по всей Азии, а также на островах западной части Индийского океана и Тихого океана, где на виды Pteropus активно охотятся. В континентальной Африке, где не обитают виды Pteropus , соломенная летучая мышь, самая крупная летучая мышь в регионе, является излюбленным объектом охоты. [ 123 ]
На Гуаме употребление марианской летучей мыши подвергает местных жителей воздействию нейротоксина бета-метиламино-L-аланина (БМАА), который впоследствии может привести к нейродегенеративным заболеваниям . BMAA может особенно усиливаться у людей, потребляющих летучих лисиц; летучие лисицы подвергаются воздействию БМАА, поедая плоды саговника . [ 124 ] [ 125 ] [ 126 ]
Как резервуары болезней
[ редактировать ]

Мегалетучие мыши являются резервуарами нескольких вирусов , которые могут поражать человека и вызывать заболевания. Они могут переносить филовирусы , в том числе вирус Эбола (EBOV) и вирус Марбург . [ 127 ] Присутствие вируса Марбург , вызывающего болезнь, вызванную вирусом Марбург , было подтверждено у одного вида — египетской фруктовой летучей мыши. Заболевание встречается редко, однако летальность при вспышке может достигать до 88%. [ 127 ] [ 128 ] Вирус был впервые обнаружен после одновременных вспышек в немецких городах Марбург и Франкфурт, а также в Белграде, Сербия , в 1967 году. [ 128 ] где заболел 31 человек и семеро умерли. [ 129 ] Вспышка была связана с лабораторными работами с верветками из Уганды . [ 128 ] Вирус может передаться от летучей мыши-хозяина человеку (который обычно провел длительное время в шахте или пещере, где обитают египетские летучие мыши); Отсюда он может передаваться от человека к человеку при контакте с инфицированными жидкостями организма, включая кровь и сперму . [ 128 ] Центры США по контролю и профилактике заболеваний перечисляют в общей сложности 601 подтвержденный случай болезни, вызванной вирусом Марбург, с 1967 по 2014 год, из которых 373 человека умерли (62% общей смертности). [ 129 ]
Виды, которые дали положительный результат на наличие EBOV, включают фруктовую летучую мышь Франке с погонами ( Epomops franqueti ), фруктовую летучую мышь с молотком и маленькую фруктовую летучую мышь с воротником. Кроме того, антитела против EBOV были обнаружены у фруктовой летучей мыши соломенного цвета, гамбийской фруктовой летучей мыши с погонами ( Epomophorus gambianus ), карликовой летучей мыши Петерса с погонами ( Micropteropus pusillus ), карликовой летучей мыши Вельдкампа с погонами ( Nanonycteris veldkampii ), розетки Лешено и Египетская фруктовая летучая мышь. [ 127 ] Многое из того, как люди заражаются вирусом Эбола, неизвестно. Ученые предполагают, что люди первоначально заражаются при контакте с инфицированным животным, таким как летучая мышь или примат, не являющийся человеком. [ 130 ] Предполагается, что мегалетучие мыши являются естественным резервуаром вируса Эбола, но это еще не установлено твердо. [ 131 ] Микролетучие мыши также исследуются как резервуар вируса: в 2019 году однажды было обнаружено, что большая длиннопалая летучая мышь ( Miniopterus inflatus ) содержит пятую часть генома вируса (хотя и не дала положительный результат на сам вирус). [ 132 ] Из-за вероятной связи между инфекцией Эбола и «охотой, разделкой и переработкой мяса зараженных животных» несколько стран Западной Африки запретили мясо диких животных (включая летучих мышей) или выпустили предупреждения о нем во время эпидемии 2013–2016 годов ; с тех пор многие запреты были сняты. [ 133 ]
Другими крупными летучими мышами, являющимися переносчиками болезней, являются в первую очередь виды Pteropus . Примечательно, что летучие лисицы могут передавать лиссавирус австралийских летучих мышей , который, наряду с вирусом бешенства , вызывает бешенство . Лиссавирус австралийских летучих мышей был впервые идентифицирован в 1996 году; он очень редко передается человеку. Заражение происходит при укусе или царапине инфицированного животного, но также может произойти и при попадании слюны инфицированного животного на слизистую оболочку или открытую рану . Воздействие крови, мочи или фекалий летучей лисицы не может вызвать заражение лиссавирусом австралийских летучих мышей. было зарегистрировано три случая заражения людей С 1994 года в Квинсленде — каждый случай был смертельным. [ 134 ]
Летучие лисицы также являются резервуарами генипавирусов, таких как вирус Хендра и вирус Нипах . Вирус Хендра был впервые идентифицирован в 1994 году; у людей это случается редко. С 1994 по 2013 год было зарегистрировано семь случаев заражения людей вирусом Хендра, четыре из которых закончились смертельным исходом. Предполагаемый основной путь заражения человека – через контакт с лошадьми , которые контактировали с мочой летучей лисицы . [ 135 ] Задокументированных случаев прямой передачи инфекции между летучими лисами и человеком не зарегистрировано. [ 136 ] По состоянию на 2012 год для лошадей доступна вакцина, снижающая вероятность заражения и передачи инфекции. [ 137 ]
Вирус Нипах был впервые выявлен в 1998 году в Малайзии. С 1998 года произошло несколько вспышек заболевания Нипах в Малайзии, Сингапуре , Индии и Бангладеш, в результате которых погибло более 100 человек. Вспышка 2018 года в Керале, Индия , привела к заражению 19 человек, 17 из которых умерли. [ 138 ] Общий уровень смертности составляет 40–75%. Люди могут заразиться вирусом Нипах при прямом контакте с летучими лисами или их жидкостями, при контакте с промежуточным хозяином , например домашними свиньями , или при контакте с инфицированным человеком. [ 139 ] Исследование индийской летучей лисицы и вируса Нипах, проведенное в 2014 году, показало, что, хотя вспышки вируса Нипах более вероятны в районах, предпочитаемых летучими лисами, «присутствие летучих мышей само по себе не считается фактором риска заражения вирусом Нипах». Скорее, употребление сока финиковой пальмы является важным путем передачи инфекции. Практика сбора сока финиковой пальмы предполагает размещение горшков для сбора на финиковых пальмах. Было замечено, что индийские летучие лисицы слизывают сок, стекающий в горшки, а также испражняются и мочатся вблизи горшков. Таким образом, люди, пьющие пальмовое вино, могут подвергнуться воздействию генипавирусов. Использование бамбуковых юбок в горшках для сбора мусора снижает риск заражения мочой летучих мышей. [ 140 ]
Летучие лисицы также могут передавать несколько несмертельных заболеваний, таких как вирус Менангл. [ 141 ] и вирус Нельсона Бэя . [ 142 ] Эти вирусы редко поражают людей, и зарегистрировано лишь несколько случаев. [ 141 ] [ 142 ] Мегалетучие мыши не подозреваются в том, что они являются переносчиками коронавирусов . [ 143 ]
В культуре
[ редактировать ]
Мегабаты, особенно летучие лисицы, присутствуют в культурах и традициях коренных народов. О них рассказывается в народных историях Австралии и Папуа-Новой Гвинеи. [ 144 ] [ 145 ] Они также были включены в наскальное искусство коренных народов Австралии, о чем свидетельствуют несколько сохранившихся примеров. [ 146 ]
Коренные народы Океании использовали части летучих лисиц в качестве функционального и церемониального оружия. На Соломоновых островах люди делали из своих костей зазубрины для использования в копьях. [ 147 ] В Новой Каледонии церемониальные топоры из нефрита украшались косами из меха летучей лисицы. [ 148 ] Крылья летучей лисицы изображались на боевых щитах народа асматов в Индонезии; они верили, что крылья защищают их воинов. [ 149 ]
Существуют современные и исторические упоминания о побочных продуктах летучей лисицы, используемых в качестве валюты . В Новой Каледонии плетеный мех летучей лисицы когда-то использовался в качестве валюты. [ 147 ] На острове Макира , который является частью Соломоновых островов, коренные народы до сих пор охотятся на летучих лисиц ради зубов, а также ради мяса диких животных. Клыки соединены вместе в ожерелья , которые используются в качестве валюты. [ 150 ] Зубы островной летучей лисицы ( Pteropus tonganus ) особенно ценятся, поскольку они обычно достаточно велики, чтобы в них можно было просверлить отверстия. На летучую лисицу Макира ( Pteropus cognatus ) также охотятся, несмотря на то, что у нее меньшие зубы. Удержание людей от использования зубов летучей лисицы в качестве валюты может нанести вред этому виду, при этом Лавери и Фаси отмечают: «Виды, которые представляют собой важный культурный ресурс, могут быть очень ценны». Акцент на устойчивой охоте на летучих лисиц для сохранения культурной ценности может быть более эффективным, чем поощрение отказа от культурной валюты. Даже если бы на летучих лисиц больше не охотились из-за зубов, их все равно убивали бы ради мяса диких животных; следовательно, сохранение их культурной ценности может способствовать устойчивой охоте. [ 151 ] Лавери заявил: «То, что их зубы имеют такую культурную ценность, является положительным, а не отрицательным. Практику охоты на летучих мышей не обязательно прекращать, ею необходимо управлять устойчиво». [ 150 ]
Сохранение
[ редактировать ]Статус
[ редактировать ]
По состоянию на 2014 год Международный союз охраны природы (МСОП) оценил четверть всех видов летучих мышей как находящихся под угрозой исчезновения , включая виды, перечисленные как находящиеся под угрозой исчезновения , находящиеся под угрозой исчезновения и уязвимые . Мегабаты находятся под серьезной угрозой со стороны людей, поскольку на них охотятся ради еды и медицинских целей. Кроме того, их выбраковывают из-за реального или предполагаемого ущерба сельскому хозяйству, особенно производству фруктов. [ 152 ] По состоянию на 2019 год МСОП провел оценку 187 видов летучих мышей. Распределение статусов следующее: [ 153 ]
- Вымершие: 4 вида (2,1%)
- Находится под угрозой исчезновения: 8 видов (4,3%)
- Под угрозой исчезновения: 16 видов (8,6%)
- Уязвимые: 37 видов (19,8%)
- Находящиеся под угрозой исчезновения : 13 видов (7,0%)
- Наименее опасные : 89 видов (47,6%)
- Недостаточно данных : 20 видов (10,7%)
Факторы, вызывающие спад
[ редактировать ]Антропогенные источники
[ редактировать ]
Мегабатам угрожает разрушение среды обитания людьми. Вырубка лесов в местах их обитания привела к утрате критически важных мест ночевки. Вырубка лесов также приводит к потере пищевых ресурсов, поскольку вырубаются местные плодоносящие деревья. Утрата среды обитания и, как следствие, урбанизация приводят к строительству новых дорог, что облегчает доступ к колониям летучих мышей для чрезмерного вылова. Кроме того, потеря среды обитания в результате вырубки лесов усугубляет естественные угрозы, поскольку фрагментированные леса более восприимчивы к повреждениям от тайфунов . [ 70 ] : 7 Громадным летучим мышам, обитающим в пещерах, угрожает вмешательство человека в места их ночлега. Добыча гуано является источником средств к существованию в некоторых странах, находящихся в их ареале, и приводит людей в пещеры. Пещеры также страдают от добычи полезных ископаемых и пещерного туризма. [ 70 ] : 8
Мегалетучих мышей также убивают люди, намеренно и непреднамеренно. Половина всех видов летучих мышей охотятся ради еды, по сравнению с лишь восемью процентами насекомоядных видов. [ 154 ] в то время как преследования людей, вызванные предполагаемым ущербом урожаю, также являются крупным источником смертности. Документально подтверждено, что некоторые летучие мыши-гиганты предпочитают местные фруктовые деревья фруктовым культурам, но вырубка лесов может сократить их запасы пищи, заставляя их полагаться на фруктовые культуры. [ 70 ] : 8 Их расстреливают, избивают до смерти или отравляют, чтобы сократить их популяцию. Смертность также происходит в результате случайного запутывания в сетке, используемой для того, чтобы летучие мыши не могли есть фрукты. [ 155 ] Кампании по отлову могут значительно сократить популяцию крупных летучих мышей. На Маврикии в период с 2014 по 2016 год было убито более 40 000 маврикийских летучих лисиц, в результате чего популяция этого вида сократилась примерно на 45%. [ 156 ] Мегалетучих мышей также убивают электрическим током. По оценкам, в одном австралийском саду за восемь недель погибло более 21 000 летучих мышей. [ 157 ] Фермеры строят электрифицированные сети над своими фруктовыми деревьями, чтобы уничтожить летучих мышей, прежде чем они смогут съесть свой урожай. Сетки сомнительно эффективны в предотвращении потерь урожая: один фермер, который эксплуатировал такую сетку, подсчитал, что он все равно теряет 100–120 тонн (220 000–260 000 фунтов) фруктов из-за летучих лисиц в год. [ 158 ] Некоторые случаи смерти от поражения электрическим током также являются случайными, например, когда летучие мыши влетают в воздушные линии электропередачи . [ 159 ]
Изменение климата приводит к смертности летучих лисиц и является источником беспокойства по поводу сохранения видов. Волны экстремальной жары в Австралии стали причиной гибели более 30 000 летучих лисиц с 1994 по 2008 год. Самки и молодые летучие мыши наиболее восприимчивы к сильной жаре, что влияет на способность популяции восстанавливаться. [ 160 ] Мегабатам угрожает повышение уровня моря, связанное с изменением климата, поскольку некоторые виды являются эндемичными для низменных атоллов . [ 103 ]
Природные источники
[ редактировать ]Поскольку многие виды являются эндемичными для одного острова, они уязвимы для случайных явлений, таких как тайфуны. Тайфун 1979 года сократил вдвое оставшуюся популяцию летучей лисицы Родригеса ( Pteropusrodricensis ). Тайфуны также приводят к косвенной смертности: поскольку тайфуны уничтожают листву деревьев, они делают летучих мышей более заметными, и, следовательно, людям легче охотиться на них. Пищевые ресурсы для летучих мышей становятся скудными после сильных штормов, и летучие мыши прибегают к более рискованным стратегиям добывания пищи, таким как поедание упавших фруктов с земли. Там они более уязвимы для нападения домашних кошек, собак и свиней. [ 93 ] Поскольку многие виды летучих мышей расположены в тектонически активном огненном кольце , им также угрожают извержения вулканов. Летучие лисицы, в том числе находящаяся под угрозой исчезновения Марианская летучая мышь, [ 118 ] [ 161 ] были почти полностью истреблены на острове Анатахан после серии извержений, начавшихся в 2003 году. [ 162 ]
Ссылки
[ редактировать ]- ^ Перейти обратно: а б Маккенна, MC; Белл, СК (1997). Классификация млекопитающих: выше уровня вида . Издательство Колумбийского университета. п. 296. ИСБН 9780231528535 .
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот Алмейда, Ф.; Джаннини, НП; Симмонс, НБ (2016). «Эволюционная история африканских летучих мышей (Chiroptera: Pteropodidae)» . Acta Chiropterologica . 18 : 73–90. дои : 10.3161/15081109ACC2016.18.1.003 . hdl : 11336/12847 . S2CID 89415407 .
- ^ Перейти обратно: а б Грей, Дж. Э. (1821). «О естественном расположении позвоночных животных» . Лондонский медицинский репозиторий (25): 299.
- ^ Перейти обратно: а б с Миллер, Геррит С. младший (1907). «Семейства и роды летучих мышей» . Бюллетень Национального музея США . 57:63 .
- ^ Перейти обратно: а б с Хатчон, Дж. М.; Кирш, Дж. А. (2006). «Подвижное лицо: деконструкция Microchiroptera и новая классификация современных летучих мышей». Acta Chiropterologica . 8 (1): 1–10. doi : 10.3161/1733-5329(2006)8[1:AMFDTM]2.0.CO;2 . S2CID 85948117 .
- ^ «Определение ПТЕРОПУСА» . Мерриам-Вебстер . Проверено 22 мая 2019 г.
- ^ Перейти обратно: а б с Джексон, С.; Джексон, С.М.; Гровс, К. (2015). Систематика австралийских млекопитающих . Издательство Csiro . ISBN 9781486300136 .
- ^ Перейти обратно: а б Добсон, GE (1875). «Конспект подотрядов, семейств и родов рукокрылых, упорядоченных в соответствии с их естественным родством» . Анналы и журнал естественной истории; Зоология, ботаника и геология . 4. 16 (95).
- ^ Перейти обратно: а б Спрингер, М.С.; Тилинг, ЕС; Мэдсен, О.; Стэнхоуп, MJ; Де Йонг, WW (2001). «Комплексные ископаемые и молекулярные данные реконструируют эхолокацию летучих мышей» . Труды Национальной академии наук . 98 (11): 6241–6246. Бибкод : 2001PNAS...98.6241S . дои : 10.1073/pnas.111551998 . ПМК 33452 . ПМИД 11353869 .
- ^ Лей, М.; Донг, Д. (2016). «Филогеномный анализ субординальных отношений летучих мышей на основе данных транскриптома» . Научные отчеты . 6 (27726): 27726. Бибкод : 2016NatSR...627726L . дои : 10.1038/srep27726 . ПМЦ 4904216 . ПМИД 27291671 .
- ^ Цагкогеорга, Г.; Паркер, Дж.; Ступка, Е.; Коттон, Дж. А.; Росситер, SJ (2013). «Филогеномный анализ выясняет эволюционные отношения летучих мышей» . Современная биология . 23 (22): 2262–2267. дои : 10.1016/j.cub.2013.09.014 . ПМИД 24184098 . S2CID 9133016 .
- ^ Щесняк, М.; Йонеда, М.; Сато, Х.; Макаловская И.; Кьюва, С.; Сугано, С.; Сузуки, Ю.; Макаловский, В.; Кай, К. (2014). «Характеристика митохондриального генома Rousettus leschenaulti». Митохондриальная ДНК . 25 (6): 443–444. дои : 10.3109/19401736.2013.809451 . ПМИД 23815317 . S2CID 207657381 .
- ^ Тилинг, ЕС; Спрингер, М.С.; Мэдсен, О.; Бейтс, П.; О'Брайен, SJ; Мерфи, WJ (2005). «Молекулярная филогения летучих мышей освещает биогеографию и летопись окаменелостей». Наука . 307 (5709): 580–584. Бибкод : 2005Sci...307..580T . дои : 10.1126/science.1105113 . ПМИД 15681385 . S2CID 25912333 .
- ^ Унгар, П. (2010). Зубы млекопитающих: происхождение, эволюция и разнообразие . JHU Пресс . п. 166. ИСБН 9780801899515 .
- ^ Джаннини, НП; Симмонс, НБ (2003). «Филогения рукокрылых летучих мышей (Mammalia: Chiroptera: Pteropodidae) на основе прямого оптимизационного анализа одного ядерного и четырех митохондриальных генов». Кладистика . 19 (6): 496–511. дои : 10.1111/j.1096-0031.2003.tb00385.x . ПМИД 34905855 . S2CID 84696546 .
- ^ Колган, диджей; Фланнери, Т.Ф. (1995). «Филогения индо-западнотихоокеанских мегахироптеров на основе рибосомальной ДНК». Систематическая биология . 44 (2): 209–220. дои : 10.1093/sysbio/44.2.209 .
- ^ Бергманс, В. (1997). «Таксономия и биогеография африканских летучих мышей (Mammalia, Megachiroptera). 5. Роды Lissonycteris Andersen, 1912, Myonycteris Matschie, 1899 и Megaloglossus Pagenstecher, 1885; общие замечания и выводы; приложение: определитель всех видов». Бофортия . 47 (2): 69.
- ^ Перейти обратно: а б с д и ж г час я дж к л м н тот п д р с Алмейда, ФК; Джаннини, НП; Десаль, Р.; Симмонс, НБ (2011). «Эволюционные взаимоотношения летучих мышей Старого Света (Chiroptera, Pteropodidae): еще одна звездная филогения?» . Эволюционная биология BMC . 11 : 281. дои : 10.1186/1471-2148-11-281 . ПМК 3199269 . ПМИД 21961908 .
- ^ Батлер, премьер-министр (1984). «Macroscelidea, Insectivora и Chiroptera из миоцена Восточной Африки» . Палеовертебрата . 14 (3): 175.
- ^ Ганнелл, Г.Ф.; Бойер, DM; Фриша, Арканзас; Наследие, С.; Манти, ФК; Миллер, ER; Саллам, HM ; Симмонс, Северная Каролина; Стивенс, Нью-Джерси; Зайферт, ER (2018). «Ископаемые лемуры из Египта и Кении предполагают африканское происхождение мадагаскарского ай-ай» . Природные коммуникации . 9 (1): 3193. Бибкод : 2018NatCo...9.3193G . дои : 10.1038/s41467-018-05648-w . ПМК 6104046 . ПМИД 30131571 .
- ^ Бургин, Коннор Дж; Колелла, Джоселин П; Кан, Филип Л; Апхэм, Натан С. (2018). «Сколько существует видов млекопитающих?» . Журнал маммологии . 99 (1): 1–14. дои : 10.1093/jmammal/gyx147 . ISSN 0022-2372 . S2CID 90797674 .
- ^ «Таксономия=Птеропус» . Красный список исчезающих видов МСОП . 2019 . Проверено 19 мая 2019 г.
- ^ Браун, Эмили Э.; Кэшмор, Дэниел Д.; Симмонс, Нэнси Б.; Батлер, Ричард Дж. (25 марта 2019 г.). Маннион, Филип (ред.). «Количественная оценка полноты летописи окаменелостей летучих мышей» . Палеонтология . 62 (5). База данных палеобиологии: 757–776. дои : 10.1111/пала.12426 . ISSN 0031-0239 . S2CID 133901426 .
- ^ Эйтинг, Т.П.; Ганнелл, Г.Ф. (2009). «Глобальная полнота летописи окаменелостей летучих мышей». Журнал эволюции млекопитающих . 16 (3): 157. doi : 10.1007/s10914-009-9118-x . S2CID 5923450 .
- ^ Перейти обратно: а б Тилинг, ЕС; Спрингер, М.С.; Мэдсен, О.; Бейтс, П.; О'Брайен, SJ; Мерфи, WJ (2005). «Молекулярная филогения летучих мышей проливает свет на биогеографию и летопись окаменелостей» (PDF) . Наука . 307 (5709): 580–584. Бибкод : 2005Sci...307..580T . дои : 10.1126/science.1105113 . ПМИД 15681385 . S2CID 25912333 .
- ^ Перейти обратно: а б Алмейда, ФК; Джаннини, НП; Десаль, Роб; Симмонс, НБ (2009). «Филогенетические взаимоотношения циноптериновых летучих мышей (Chiroptera: Pteropodidae: Cynopterinae)». Молекулярная филогенетика и эволюция . 53 (3): 772–783. дои : 10.1016/j.ympev.2009.07.035 . hdl : 11336/74530 . ПМИД 19660560 .
- ^ О'Брайен, Дж.; Мариани, К.; Олсон, Л.; Рассел, Алабама; Скажи, Л.; Йодер, AD; Хайден, Ти Джей (2009). «Множественные колонизации западной части Индийского океана летучими мышами Pteropus (Megachiroptera: Pteropodidae): первыми были колонизированы самые дальние острова». Молекулярная филогенетика и эволюция . 51 (2): 294–303. дои : 10.1016/j.ympev.2009.02.010 . ПМИД 19249376 .
- ^ Тилинг ЕС, Джонс Дж., Росситер С.Дж. (2016). «Филогения, гены и слух: последствия для эволюции эхолокации у летучих мышей». В Фентон М.Б., Гриннелл А.Д., Поппер А.Н., Фэй Р.Н. (ред.). Биоакустика летучих мышей . Справочник Спрингера по слуховым исследованиям. Том. 54. Нью-Йорк: Спрингер. стр. 25–54. дои : 10.1007/978-1-4939-3527-7_2 . ISBN 9781493935277 .
- ^ Ван, Чжэ; Чжу, Тэнтэн; Сюэ, Хуэйлин; Фанг, На; Чжан, Цзюньпэн; Чжан, Либяо; Панг, Цзянь; Тилинг, Эмма К.; Чжан, Шуи (2017). «Пренатальное развитие подтверждает единство происхождения гортанной эхолокации у летучих мышей». Экология и эволюция природы . 1 (2): 21. дои : 10.1038/s41559-016-0021 . ПМИД 28812602 . S2CID 29068452 .
- ^ Голландия, РА; Уотерс, Д.А.; Рейнер, Дж. М. (декабрь 2004 г.). «Структура эхолокационного сигнала у летучей мыши-гиганта Rousettus aegyptiacus Geoffroy 1810». Журнал экспериментальной биологии . 207 (Часть 25): 4361–4369. дои : 10.1242/jeb.01288 . ПМИД 15557022 . S2CID 2715542 .
- ^ Бунман, А.; Бумрунгсри, С.; Йовель, Ю. (декабрь 2014 г.). «Нехолокирующие фруктовые летучие мыши производят биосонарные щелчки своими крыльями» . Современная биология . 24 (24): 2962–2967. дои : 10.1016/j.cub.2014.10.077 . ПМИД 25484290 . S2CID 17789233 .
- ^ Спикмен, младший; Рэйси, Пенсильвания (апрель 1991 г.). «Нет затрат на эхолокацию летучих мышей в полете». Природа . 350 (6317): 421–423. Бибкод : 1991Natur.350..421S . дои : 10.1038/350421a0 . ПМИД 2011191 . S2CID 4314715 .
- ^ Ланкастер, Вашингтон; Хенсон, Огайо; Китинг, AW (январь 1995 г.). «Активность дыхательных мышц в отношении вокализации летучих мышей» (PDF) . Журнал экспериментальной биологии . 198 (Часть 1): 175–191. дои : 10.1242/jeb.198.1.175 . ПМИД 7891034 .
- ^ Перейти обратно: а б Альтрингем Джей Ди (2011). «Эхолокация и другие чувства». Летучие мыши: от эволюции к сохранению . Нью-Йорк: Издательство Оксфордского университета . ISBN 9780199207114 .
- ^ Перейти обратно: а б с Хатчон, Дж. М.; Гарланд, Т. младший (2004). «Мегабаты большие?». Журнал эволюции млекопитающих . 11 (3/4): 257–277. дои : 10.1023/B:JOMM.0000047340.25620.89 . S2CID 11528722 .
- ^ Неси, Николас; Цагкогеорга, Грузия; Цанг, Сьюзен М; Николя, Виолен; Лалис, Од; Скэнлон, Аннетт Т; Рисле-Сбарбаро, Силке А; Вианторо, Сигит; Хитч, Алан Т; Жюсте, Хавьер; Пинзари, Коринна А (4 марта 2021 г.). «Изучение филогенетических разногласий разрешает глубокие расколы в быстром распространении летучих мышей Старого Света (Chiroptera: Pteropodidae)» . Систематическая биология . 70 (6): 1077–1089. дои : 10.1093/sysbio/syab013 . ISSN 1063-5157 . ПМЦ 8513763 . ПМИД 33693838 .
- ^ Ганнелл, Грегг Ф.; Манти, Фредрик К. (апрель 2018 г.). «Плиоценовые летучие мыши (Chiroptera) из Канапои, бассейн Туркана, Кения» . Журнал эволюции человека . 140 :4. дои : 10.1016/j.jhevol.2018.01.001 . ISSN 0047-2484 . ПМИД 29628118 . S2CID 206143059 .
- ^ Фланнери, Т. (1995). Млекопитающие юго-западной части Тихого океана и Молуккских островов . Издательство Корнельского университета. п. 271. ИСБН 0801431506 .
- ^ Перейти обратно: а б Новак, Р.М.; Уокер, EP; Кунц, TH; Пирсон, ЭД (1994). Летучие мыши мира Уокера . Джу Пресс. п. 49 . ISBN 9780801849862 .
- ^ Ум, В.; Клейман, Д.Г.; Макдейд, MC (2004). Энциклопедия жизни животных Гржимека Млекопитающие II . Том. 13 (2-е изд.). Гейл . п. 309.
- ^ Перейти обратно: а б с д и ж г час Нельсон, Дж. Э. Фауна Австралии (PDF) (Отчет). Том. 1Б. Департамент окружающей среды и энергетики правительства Австралии .
- ^ Сантана, ЮВ; Циферблат, ТО; Эйтинг, Т.П.; Альфаро, Мэн (2011). «Экология ночлега и эволюция отметин на шерсти летучих мышей» . ПЛОС Один . 6 (10): e25845. Бибкод : 2011PLoSO...625845S . дои : 10.1371/journal.pone.0025845 . ПМК 3185059 . ПМИД 21991371 .
- ^ Перейти обратно: а б с д Холл, Лос-Анджелес; Ричардс, Г. (2000). Летучие лисицы: фруктовые и цветочные летучие мыши Австралии . УНСВ Пресс . ISBN 9780868405612 .
- ^ Инглби, С.; Колган, Д. (2003). «Электрофоретические исследования систематических и биогеографических взаимоотношений фиджийских летучих мышей родов Pteropus, Pteralopex, Chaerephon и Notopteris» . Австралийская маммология . 25:13 . дои : 10.1071/AM03013 .
- ^ Воган, штат Техас; Райан, Дж. М.; Чаплевски, Нью-Джерси (11 декабря 2013 г.). Маммология (6-е изд.). Издательство Джонс и Бартлетт. стр. 100-1 255–256. ISBN 9781284032185 .
- ^ Симмонс, Нэнси Б.; Конвей, Тенли М. (2001). «Филогенетические взаимоотношения летучих мышей-мормоопидов (Chiroptera: Mormoopidae) на основе морфологических данных» . Бюллетень Американского музея естественной истории . 258 : 17. doi : 10.1206/0003-0090(2001)258<0001:PROMBC>2.0.CO;2 . hdl : 2246/1608 . ISSN 0003-0090 . S2CID 83580917 .
- ^ Линденау, Криста (2011). «Летучие мыши среднего плейстоцена (Mammalia: Chiroptera) из пещеры Яримбургаз в Турецкой Фракии (Турция)». E&G – Четвертичный научный журнал . 55 : 127. дои : 10.23689/fidgeo-999 .
- ^ Тейт, GHH (1942). «Результаты экспедиции Арчболда № 48: Pteropodidae (Chiroptera) из коллекций Арчболда» . Бюллетень Американского музея естественной истории . 80 : 332–335.
- ^ Перейти обратно: а б Джаннини, НП; Симмонс, НБ (2007). «Гомология элементов и эволюция зубных формул у рукокрылых летучих мышей (Mammalia: Chiroptera: Pteropodidae)» (PDF) . Американский музей Novitates (3559): 1–27. doi : 10.1206/0003-0082(2007)3559[1:EHATEO]2.0.CO;2 . hdl : 2246/5849 . S2CID 62823404 .
- ^ Перейти обратно: а б Жюсте, Дж.; Ибаньес, К. (1993). «Асимметричная зубная формула у млекопитающего, летучей мыши Myonycteris brachycephala с острова Сан-Томе (Mammalia: Megachiroptera)». Канадский журнал зоологии . 71 (1): 221–224. дои : 10.1139/z93-030 . hdl : 10261/48798 .
- ^ Перейти обратно: а б с д и Воган, Т. (1970). «Глава 3: Скелетная система». В Вимсатте, В. (ред.). Биология летучих мышей . Академическая пресса . стр. 103–136. ISBN 9780323151191 .
- ^ Луо, ZX; Килан-Яворовска, З.; Чифелли, Р.Л. (2004). «Эволюция замены зубов у млекопитающих» (PDF) . Бюллетень Музея естественной истории Карнеги . 2004 (36): 159–176. doi : 10.2992/0145-9058(2004)36[159:EODRIM]2.0.CO;2 . S2CID 5630875 . Архивировано из оригинала (PDF) 23 мая 2019 года.
- ^ Перейти обратно: а б с Новак, Р.М.; Пиллсбери Уокер, Э. (1999). Млекопитающие мира Уокера . Том. 1. ЖУ Пресс. п. 258. ИСБН 9780801857898 .
- ^ Перейти обратно: а б Беннетт, МБ (1993). «Структурные модификации захвата передних и задних конечностей некоторых летучих лисиц (Chiroptera: Pteropodidae)». Журнал зоологии . 229 (2): 237–248. дои : 10.1111/j.1469-7998.1993.tb02633.x .
- ^ Шатт, Вашингтон; Симмонс, НБ (1998). «Морфология и гомология рукокрылых Calca с комментариями к филогенетическим отношениям археоптеропов ». Журнал эволюции млекопитающих . 5 : 2. дои : 10.1023/A:1020566902992 . S2CID 20426664 .
- ^ Перейти обратно: а б Майна, JN; Кинг, А.С. (1984). «Корреляция между структурой и функцией в конструкции легких летучей мыши: морфометрическое исследование» (PDF) . Журнал экспериментальной биологии . 11:44 .
- ^ Карпентер, Р.Э. (1986). «Физиология полета летучих мышей среднего размера (Pteropodidae)» (PDF) . Журнал экспериментальной биологии . 120 : 84–93. дои : 10.1242/jeb.120.1.79 .
- ^ Перейти обратно: а б с Ричардс, GC (1983). «Летучие мыши и их родственники». В Страхане, Р. (ред.). Полная книга млекопитающих Австралии. Национальный фотографический указатель дикой природы Австралии (1-е изд.). Лондон: Ангус и Робертсон. стр. 271–273. ISBN 978-0207144547 .
- ^ Шмидт-Раэса, А., изд. (2017). Сравнительная анатомия желудочно-кишечного тракта у Eutheria II . Вальтер де Грюйтер ГмбХ & Ко КГ . стр. 328–330. ISBN 9783110560671 .
- ^ Смит, JDL; Грегори, ТР (2009). «Размеры генома летучих мышей (Chiroptera: Pteropodidae) чрезвычайно ограничены» . Письма по биологии . 5 (3): 347–351. дои : 10.1098/rsbl.2009.0016 . ПМЦ 2679926 . ПМИД 19324635 .
- ^ Перейти обратно: а б с Мюллер, Б.; Гудман, С.М.; Пейхл, Лео (2007). «Разнообразие конусных фоторецепторов в сетчатке летучих мышей (Megachiroptera)». Мозг, поведение и эволюция . 70 (2): 90–104. дои : 10.1159/000102971 . ПМИД 17522478 . S2CID 29095435 .
- ^ Перейти обратно: а б Грейдон, М.; Георгий, П.; Петтигрю, Дж. (1987). «Зрение летучих лисиц (Chiroptera: Pteropodidae)» . Журнал Австралийского общества млекопитающих . 10 (2): 101–105. дои : 10.1071/AM87021 . S2CID 254716342 .
- ^ Тиагавел, Дж.; Чечетто, К.; Сантана, ЮВ; Якобсен, Л.; Ордер, Э.Дж.; Рэтклифф, Дж. М. (2018). «Возможности слуха и ограничения зрения способствовали развитию эхолокации у летучих мышей» . Природные коммуникации . 9 (1): 98. Бибкод : 2018NatCo...9...98T . дои : 10.1038/s41467-017-02532-x . ПМЦ 5758785 . ПМИД 29311648 .
- ^ Джаннини, НП; Алмейда, ФК; Симмонс, Северная Каролина; Хелген, К.М. (2008). «Систематическое положение Pteropus leucopterus и его влияние на монофилию и взаимоотношения Pteropus (Chiroptera: Pteropodidae)». Acta Chiropterologica . 10 :11–20. дои : 10.3161/150811008X331054 . hdl : 11336/82001 . S2CID 59028595 .
- ^ Перейти обратно: а б с Джонс, Г.; Тилинг, ЕС; Росситер, С.Дж. (2013). «От ультразвука к инфракрасному: молекулярная эволюция и сенсорная биология летучих мышей» . Границы в физиологии . 4 : 117. doi : 10.3389/fphys.2013.00117 . ПМЦ 3667242 . ПМИД 23755015 .
- ^ Перейти обратно: а б Шваб, И.Р. (2005). «Хороидальная ловкость рук» . Британский журнал офтальмологии . 89 (11): 1398. doi : 10.1136/bjo.2005.077966 . ПМК 1772916 . ПМИД 16267906 .
- ^ Перейти обратно: а б Вуд, ВФ; Уолш, А.; Сейджагат, Дж.; Уэлдон, П.Дж. (2005). «Летучие соединения в секретах плечевых желез самцов летучих лисиц, род Pteropus (Pteropodidae, Chiroptera)» . З Натурфорш С. 60 (9–10): 779–784. дои : 10.1515/znc-2005-9-1019 . ПМИД 16320623 . S2CID 1938648 .
- ^ Вагнер, Дж. (2008). «Железистые выделения самцов Pteropus (летучих лисиц): предварительное химическое сравнение видов» . Сборник независимых исследовательских проектов (Isp) .
- ^ Ли, Д.; Чжан, Дж. (2014). «Диета формирует эволюцию генного репертуара рецепторов горького вкуса у позвоночных» . Молекулярная биология и эволюция . 31 (2): 303–309. дои : 10.1093/molbev/mst219 . ПМК 3907052 . ПМИД 24202612 .
- ^ Перейти обратно: а б с д и ж г час я Миклбург, СП; Хатсон, AM; Рэйси, Пенсильвания (1992). Летучие мыши Старого Света: план действий по их сохранению (PDF) (Отчет). Гланд, Швейцария: МСОП .
- ^ Хенгжан, Юпади; Иида, Кейсуке; Дойсабас, Карла Кристин Дж.; Пхичитрасилп, Танмапорн; Омори, Ясусигэ; Хондо, Эйичи (2017). «Суточное поведение и баланс активности летучей лисицы с золотой короной ( Acerodon jubatus ) в лесном заповеднике Субик-Бэй, Филиппины» . Журнал ветеринарной медицины . 79 (10): 1667–1674. дои : 10.1292/jvms.17-0329 . ПМЦ 5658557 . ПМИД 28804092 .
- ^ Перейти обратно: а б Хайдеман, П.Д. (1988). «Время размножения плодовой летучей мыши Haplonycteris fischeri (Pteropodidae): географическое разнообразие и задержка развития». Журнал зоологии . 215 (4): 577–595. дои : 10.1111/j.1469-7998.1988.tb02396.x . hdl : 2027.42/72984 .
- ^ Новак, Р.М.; Пиллсбери Уокер, Э. (1999). Млекопитающие мира Уокера . Том. 1. ЖУ Пресс. п. 287. ИСБН 9780801857898 .
- ^ Перейти обратно: а б Фокс, Саманта; Спенсер, Хью; О'Брайен, Джемма М. (2008). «Анализ близнецов у летучих лисиц (Megachiroptera) выявляет суперфетацию и множественное отцовство». Acta Chiropterologica . 10 (2): 271–278. дои : 10.3161/150811008X414845 . S2CID 83506361 .
- ^ Шринивасан, Массачусетс; Бхат, HR; Гиваргезе, Г. (30 марта 1974 г.). «Наблюдения за репродуктивным циклом Cynopterus sphinx sphinx Vahl, 1797 (Chiroptera: Pteropidae)» Журнал маммологии . 55 (1): 200–202. дои : 10.2307/1379269 . JSTOR 1379269 . ПМИД 4819592 .
- ^ Перейти обратно: а б Дуглас Хейссен, В.; Ван Тиенховен, А.; Ван Тиенховен, А. (1993). Модели размножения млекопитающих Асделла: сборник данных по конкретным видам . Издательство Корнелльского университета . п. 89. ИСБН 9780801417535 .
- ^ Альтрингем, Джон Д.; МакОват, Том; Хаммонд, Люси (2011). Летучие мыши: от эволюции к сохранению (2-е изд.). Оксфорд и Нью-Йорк: Издательство Оксфордского университета. п. хв. ISBN 978-0-19-920711-4 .
- ^ Кунц, TH; Курта, А. (1987). «Размер летучих мышей при рождении и материнские инвестиции во время беременности» (PDF) . Симпозиумы Лондонского зоологического общества . 57 .
- ^ Сафи, К. (2008). «Социальные летучие мыши: взгляд мужчин» . Журнал маммологии . 89 (6): 1342–1350. doi : 10.1644/08-МАММ-S-058.1 . S2CID 85733862 .
- ^ Крайтон, Е.Г.; Крутч, PH, ред. (2000). Репродуктивная биология летучих мышей . Академическая пресса. п. 433 . ISBN 9780080540535 .
- ^ Рэйси, Д.Н.; Пикер, М.; Рэйси, Пенсильвания (2009). «Галакторея – это не лактация». Тенденции в экологии и эволюции . 24 (7): 354–355. дои : 10.1016/j.tree.2009.03.008 . ПМИД 19427057 .
- ^ Кунц, TH; Хоскен, Дэвид Дж (2009). «Мужская лактация: почему, почему бы и нет и нужно ли это?». Тенденции в экологии и эволюции . 24 (2): 80–85. дои : 10.1016/j.tree.2008.09.009 . ПМИД 19100649 .
- ^ Шуман, MC; Гудман, С.М. (2012). «Вокализации малагасийской пещерной летучей мыши Eidolon dupreanum: возможные доказательства зарождающейся эхолокации?». Acta Chiropterologica . 14 (2): 409. дои : 10.3161/150811012X661729 . S2CID 86613252 .
- ^ «Фруктовая летучая мышь с головой молотка» . Журнал БАТС . Том. 34, нет. 1. 2015 . Проверено 30 мая 2019 г.
- ^ Лавлесс, AM; Макби, К. (2017). «Nyctimene robinsoni (Chiroptera: Pteropodidae)» . Виды млекопитающих . 49 (949): 68–75. дои : 10.1093/mspecies/sex007 . S2CID 89828640 .
- ^ Прат, Йосеф; Тауб, Мор; Йовель, Йоси (2015). «Вокальное обучение социального млекопитающего: продемонстрировано экспериментами по изоляции и воспроизведению на летучих мышах» . Достижения науки . 1 (2): e1500019. Бибкод : 2015SciA....1E0019P . дои : 10.1126/sciadv.1500019 . ISSN 2375-2548 . ПМЦ 4643821 . ПМИД 26601149 .
- ^ Верн, Южная Каролина (2017). «Что летучие мыши говорят о речи и языке» . Психономический бюллетень и обзор . 24 (1): 111–117. дои : 10.3758/s13423-016-1060-3 . ПМК 5325843 . ПМИД 27368623 .
- ^ Прат, Йосеф; Азулай, Линдси; Дор, Рой; Йовель, Йоси (2017). «Обучение толпе вокалу вызывает у летучих мышей вокальные диалекты: воспроизведение речи сородичей формирует использование щенками основных частот» . ПЛОС Биология . 15 (10): e2002556. дои : 10.1371/journal.pbio.2002556 . ISSN 1545-7885 . ПМЦ 5663327 . ПМИД 29088225 .
- ^ Циммер, К. (1 января 2018 г.). «Что ссоры летучих мышей говорят нам об обучении вокалу» . Ученый . Проверено 23 мая 2019 г.
- ^ Хартен, Л.; Прат, Ю.; Бен Коэн, С.; Дор, Р.; Йовель, Ю. (2019). «Пища для секса у летучих мышей: самцы-производители размножаются с помощью добывающих самок» . Современная биология . 29 (11): 1895–1900.е3. дои : 10.1016/j.cub.2019.04.066 . ПМИД 31130455 . S2CID 162183551 .
- ^ Перейти обратно: а б Сугита, Н. (2016). «Гомосексуальная фелляция: лизание эрегированного пениса самцами бонинских летучих лисиц Pteropus pselaphon» . ПЛОС ОДИН . 11 (11): e0166024. Бибкод : 2016PLoSO..1166024S . дои : 10.1371/journal.pone.0166024 . ПМК 5100941 . ПМИД 27824953 .
- ^ Перейти обратно: а б Тан, М.; Джонс, Г.; Чжу, Г.; Йе, Дж.; Хонг, Т.; Чжоу, С.; Чжан, С.; Чжан, Л. (2009). «Феллация летучих мышей продлевает время совокупления» . ПЛОС ОДИН . 4 (10): е7595. Бибкод : 2009PLoSO...4.7595T . дои : 10.1371/journal.pone.0007595 . ПМК 2762080 . ПМИД 19862320 .
- ^ Перейти обратно: а б с Пирсон, Эд; Рейни, МЫ (1992). «Биология летучих лисиц рода Pteropus: обзор» . Биологический отчет . 90 (23).
- ^ Перейти обратно: а б Дюмон, скорая помощь; О'Нил, Р. (2004). «Твердость пищи и пищевое поведение летучих мышей Старого Света (Pteropodidae)» . Журнал маммологии . 85 : 8–14. дои : 10.1644/BOS-107 . S2CID 27275791 .
- ^ Перейти обратно: а б Инь, К.; Чжу, Л.; Лю, Д.; Ирвин, DM; Чжан, С.; Пан, Ю. (2016). «Молекулярная эволюция ядерного фактора (эритроидного происхождения 2) - подобного 2 гена Nrf2 у фруктовых летучих мышей Старого Света (Chiroptera: Pteropodidae)» . ПЛОС ОДИН . 11 (1): e0146274. Бибкод : 2016PLoSO..1146274Y . дои : 10.1371/journal.pone.0146274 . ПМК 4703304 . ПМИД 26735303 .
- ^ Суды, SE (1998). «Диетические стратегии летучих мышей Старого Света (Megachiroptera, Pteropodidae): как они получают достаточное количество белка?» . Обзор млекопитающих . 28 (4): 185–194. дои : 10.1046/j.1365-2907.1998.00033.x .
- ^ Перейти обратно: а б с д Норберг, UM и Рейнер, JMV (1987). «Экологическая морфология и полет летучих мышей (Mammalia: Chiroptera): адаптации крыльев, летные характеристики, стратегия кормления и эхолокация». Философские труды Королевского общества Б. 316 (1179): 382–383. Бибкод : 1987РСТБ.316..335Н . дои : 10.1098/rstb.1987.0030 .
- ^ Ходжкисон, Р.; Болдинг, СТ; Зубайд, А.; Кунц, TH (2003). «Фруктовые летучие мыши (Chiroptera: Pteropodidae) как распространители семян и опылители в равнинных тропических лесах Малайзии1». Биотропика . 35 (4): 491–502. дои : 10.1111/j.1744-7429.2003.tb00606.x . S2CID 86327074 .
- ^ Шилтон, Луизиана; Альтрингем, доктор медицинских наук; Комптон, Южная Каролина; Уиттакер, Р.Дж. (1999). «Плодовые летучие мыши Старого Света могут распространять семена на большие расстояния благодаря длительному удержанию жизнеспособных семян в кишечнике» . Труды Лондонского королевского общества. Серия Б: Биологические науки . 266 (1416): 219–223. дои : 10.1098/rspb.1999.0625 . ПМК 1689670 .
- ^ Олексий Р.; Рэйси, Пенсильвания; Джонс, Г. (2015). «GPS-слежение с высоким разрешением показывает выбор среды обитания и возможность распространения семян на большие расстояния мадагаскарскими летучими лисцами Pteropus rufus» . Глобальная экология и охрана природы . 3 : 690. doi : 10.1016/j.gecco.2015.02.012 .
- ^ Корлетт, RT (2017). «Плодоядность и распространение семян позвоночными животными в тропической и субтропической Азии: обновленная информация» . Глобальная экология и охрана природы . 11:13 . дои : 10.1016/j.gecco.2017.04.007 .
- ^ Ридер, DM; Кунц, TH; Видмайер, EP (2004). «Базовый уровень и вызванные стрессом глюкокортикоиды во время размножения у изменчивой летучей лисицы Pteropus hypomelanus (Chiroptera: Pteropodidae)». Журнал экспериментальной зоологии . 301А (8): 682–690. дои : 10.1002/jez.a.58 . ПМИД 15286948 .
- ^ Перейти обратно: а б Буден, Д.; Хелген, КМ; Уайлс, Г. (2013). «Таксономия, распространение и естественная история летучих лисиц (Chiroptera, Pteropodidae) на островах Мортлок и в штате Чуук, Каролинские острова» . ZooKeys (345): 97–135. дои : 10.3897/zookeys.345.5840 . ПМЦ 3817444 . ПМИД 24194666 .
- ^ Эссельстин, Дж.А.; Амар, А.; Янке, Д. (2006). «Влияние охоты после тайфуна на марианских летучих мышей (Pteropus mariannus)». Тихоокеанская наука . 60 (4): 531–532. дои : 10.1353/psc.2006.0027 . S2CID 55543225 .
- ^ Адаме, Мария Фернанда; Джардин, ТД; Фрай, Б.; Вальдес, Д.; Линднер, Г.; Наджи, Дж.; Банн, ЮВ (2018). «Эстуарные крокодилы в тропической прибрежной пойме получают питание за счет наземной добычи» . ПЛОС ОДИН . 13 (6): e0197159. Бибкод : 2018PLoSO..1397159A . дои : 10.1371/journal.pone.0197159 . ПМЦ 5991389 . ПМИД 29874276 .
- ^ Летучие лисицы против пресноводного крокодила (видео). BBC Земля. 10 апреля 2015 г. Архивировано из оригинала 7 ноября 2021 г. . Проверено 22 мая 2019 г.
- ^ Рамасиндразана, Б.; Гудман, С.М.; Гомар, Ю.; Дик, CW; Тортоса, П. (2017). «Скрытое разнообразие летучих мышей Nycteribiidae (Diptera) из малагасийского региона и понимание взаимодействия хозяина и паразита» . Паразиты и переносчики . 10 (1): 630. doi : 10.1186/s13071-017-2582-x . ПМЦ 5747079 . ПМИД 29284533 .
- ^ Раманантсалама, Р.В.; Андрианаримиса, А.; Раселиманана, АП; Гудман, С.М. (2018). «Уровни потребления гематофагов-эктопаразитов во время ухода эндемичной мадагаскарской фруктовой летучей мышью» . Паразиты и переносчики . 11 (1): 330. doi : 10.1186/s13071-018-2918-1 . ПМЦ 5984742 . ПМИД 29859123 .
- ^ Деш, CE (1981). «Новый вид клеща-демодецида (Acari: Prostigmata) из Западной Австралии, паразитирующий на Macroglossus minimus (Chiroptera: Pteropodidae)» (PDF) . Записи Музея Западной Австралии . 9 (1): 41–47.
- ^ Ландау, И.; Чаватт, Дж. М.; Караджян Г.; Шабо, А.; Беверидж, И. (2012). «Гемоспоридии-паразиты летучих мышей с описанием Sprattiella alectogen. Nov., sp. Nov» . Паразит . 19 (2): 137–146. дои : 10.1051/паразит/2012192137 . ПМЦ 3671437 . ПМИД 22550624 .
- ^ Кингдон, Дж.; Хаппольд, Д.; Бутынский Т.; Хоффманн, М.; Хаппольд, М.; Калина, Дж. (2013). Млекопитающие Африки . Том. 4. А&С Черный. ISBN 9781408189962 .
- ^ Перейти обратно: а б с Бенда, Питер; Валло, Питер; Хулва, Пол; Горачек, Иван (2012). «Египетская летучая мышь Rousettus aegyptiacus (Chiroptera: Pteropodidae) в Палеарктике: географическое разнообразие и таксономический статус» . Биология . 67 (6): 1230–1244. дои : 10.2478/s11756-012-0105-y . S2CID 14907114 .
- ^ Миклбург, С.; Хатсон, AM; Бергманс, В.; Фар, Дж.; Рэйси, Пенсильвания (2008). «Эйдолон хелвум» . Красный список исчезающих видов МСОП . 2008 : e.T7084A12824968. doi : 10.2305/IUCN.UK.2008.RLTS.T7084A12824968.en .
- ^ , Цзинь-Шо; Джонс, Гарет; Чжу, Гуан-Цзянь, Шу-И (2010). Чжан 12 103–116 : . /150811010X504626 . S2CID 86402486 10.3161
- ^ Винсено, К. (2017). « Птеропус дасималлус » . Красный список исчезающих видов МСОП . 2017 : e.T18722A22080614. doi : 10.2305/IUCN.UK.2017-2.RLTS.T18722A22080614.en .
- ^ Маэда, К. (2008). «Птеропус лохоэнсис» . Красный список исчезающих видов МСОП . 2008 : e.T18773A8614831. doi : 10.2305/IUCN.UK.2008.RLTS.T18773A8614831.en .
- ^ Винсено, К. (2017). « Птеропус пселафон » . Красный список исчезающих видов МСОП . 2017 : e.T18752A22085351. doi : 10.2305/IUCN.UK.2017-2.RLTS.T18752A22085351.en .
- ^ Перейти обратно: а б Эллисон, А.; Бонаккорсо, Ф.; Хелген К. и Джеймс Р. (2008). « Птеропус Марианнус » . Красный список исчезающих видов МСОП . 2008 : e.T18737A8516291. doi : 10.2305/IUCN.UK.2008.RLTS.T18737A8516291.en .
- ^ Перейти обратно: а б с «Таксономия=Pteropodidae, Регионы суши=Западная и Центральная Азия, Южная и Юго-Восточная Азия» . МСОП . Проверено 24 августа 2019 г.
- ^ Тейт, Джессика; Перотто-Бальдивьесо, Умберто Л.; Маккеун, Адам; Уэсткотт, Дэвид А. (2014). «Летучие лисицы приезжают в город? Урбанизация очковой летучей лисицы (Pteropus conspicillatus) в Австралии» . ПЛОС ОДИН . 9 (10): е109810. Бибкод : 2014PLoSO...9j9810T . дои : 10.1371/journal.pone.0109810 . ПМК 4190360 . ПМИД 25295724 .
- ^ Перейти обратно: а б «Таксономия=Pteropodidae, Сухопутные регионы=Океания» . МСОП . Проверено 24 августа 2019 г.
- ^ Джонс, Кентукки; Миклбург, СП; Сехрест, В.; Уолш, А.Л. «Глобальный обзор сохранения островных летучих мышей: важные проблемы и возможности» . Во Флеминге, TH; Рэйси, Пенсильвания (ред.). Эволюционная экология и охрана островных рукокрылых . Издательство Чикагского университета.
- ^ Миклбург, С.; Уэйлен, К.; Рэйси, П. (2009). «Летучие мыши как мясо диких животных: глобальный обзор» . Орикс . 43 (2): 217. дои : 10.1017/s0030605308000938 . S2CID 85261295 .
- ^ Банак, ЮАР; Марч, С.Дж.; Кокс, Пенсильвания (2006). «Нейротоксичные летучие лисицы как продукт питания для народа чаморро, Марианские острова». Журнал этнофармакологии . 106 (1): 97–104. дои : 10.1016/j.jep.2005.12.032 . ПМИД 16457975 .
- ^ Кокс, П.; Дэвис, Д.; Маш, Д.; Меткалф, Дж. С.; Банак, Ю.А. (2016). «Пищевое воздействие токсинов окружающей среды вызывает нейрофибриллярные клубки и отложения амилоида в мозге» . Труды Королевского общества Б. 283 (3): 1–10. дои : 10.1098/rspb.2015.2397 . ПМЦ 4795023 . ПМИД 26791617 .
- ^ Холткамп, В. (2012). «Новая наука о BMAA: способствуют ли цианобактерии нейродегенеративным заболеваниям?» . Перспективы гигиены окружающей среды . 120 (1823 г.): а110–а116. дои : 10.1289/ehp.120-a110 . ПМЦ 3295368 . ПМИД 22382274 .
- ^ Перейти обратно: а б с Хасанин, А.; Неси, Н.; Марин, Дж.; Каджо, Б.; Пуррут, X.; Лерой, Э.; Гембу, Г.; Мусаба Акава, П.; Нгогуни, К.; Накуне, Э.; Руди, М.; Чикунг, Д.; Понгомбо Шонго, К.; Бонилья, К. (2016). «Сравнительная филогеография африканских летучих мышей (Chiroptera, Pteropodidae) дает новое представление о вспышке болезни, вызванной вирусом Эбола, в Западной Африке, 2014–2016 годы» . Счета Биологии . 339 (11–12): 517–528. дои : 10.1016/j.crvi.2016.09.005 . ПМИД 27746072 . S2CID 44412099 .
- ^ Перейти обратно: а б с д «Марбургская геморрагическая лихорадка» . Всемирная организация здравоохранения . Проверено 30 мая 2019 г.
- ^ Перейти обратно: а б «Хронология вспышек Марбургской геморрагической лихорадки» . Центры по контролю и профилактике заболеваний . 9 октября 2014 года . Проверено 30 мая 2019 г.
- ^ «Эбола (болезнь, вызванная вирусом Эбола): передача» . Центры по контролю и профилактике заболеваний . 17 мая 2019 года . Проверено 30 мая 2019 г.
- ^ «Исследование резервуара Эболы» . Центры по контролю и профилактике заболеваний . 9 июля 2018 года . Проверено 30 мая 2019 г.
- ^ Купфершмидт, К. (24 января 2019 г.). «Этот вид летучих мышей может быть источником эпидемии Эболы, унесшей жизни более 11 000 человек в Западной Африке» . Наука . Проверено 30 мая 2019 г.
- ^ Зон, Х.; Петеш, К. (21 сентября 2016 г.). «После Эболы жители Западной Африки с риском возвращаются к мясу диких животных» . Ассошиэйтед Пресс . Архивировано из оригинала 25 декабря 2019 года . Проверено 30 мая 2019 г.
- ^ «Информационный бюллетень по бешенству и лиссавирусу австралийских летучих мышей» . health.nsw.gov.au . Министерство здравоохранения штата Новый Южный Уэльс, Новый Южный Уэльс, 2015 г., 30 ноября 2015 г. Проверено 14 июня 2018 г.
- ^ «Вирусная болезнь Хендра (ВеВ)» (PDF) . cdc.gov . Министерство здравоохранения и социальных служб США . Проверено 14 июня 2018 г.
- ^ Санчес, Калифорния; Бейкер, МЛ (2016). «Восприятие риска заболеваний и меры безопасности: опрос австралийских реабилитаторов летучей лисицы» . PLOS Забытые тропические болезни . 10 (2): e0004411. дои : 10.1371/journal.pntd.0004411 . ПМЦ 4734781 . ПМИД 26829399 .
- ^ «Вакцина Хендра» . ava.com . Австралийская ветеринарная ассоциация Ltd (AVA). 2018. Архивировано из оригинала 14 июня 2018 года . Проверено 14 июня 2018 г.
- ^ Галланд, А. (12 июня 2018 г.). «Вирус Нипах в Индии «под контролем», но Великобритания и весь мир должны быть готовы к появлению признаков заражения путешественников» . Телеграф . Telegraph Media Group Limited 2018. Архивировано из оригинала 12 января 2022 года . Проверено 14 июня 2018 г.
- ^ «Вирус Нипах» . Всемирная организация здравоохранения . ВОЗ. 30 мая 2018 года . Проверено 14 июня 2018 г.
- ^ Хан, МБ; Эпштейн, Дж. Х.; Герли, ES; Ислам, MS; Лубы, ИП; Дашак, П.; Патц, Дж. А. (2014). «Поведение Pteropus giganteus в условиях ночлега и выбор среды обитания обнаруживают потенциальную связь с эпидемиологией вируса Нипах» . Журнал прикладной экологии . 51 (2): 376–387. дои : 10.1111/1365-2664.12212 . ПМК 4000083 . ПМИД 24778457 .
- ^ Перейти обратно: а б Боуден, TR; Вестенберг, М.; Ван, Л.; Итон, Британская Колумбия; Бойл, Д.Б. (2001). «Молекулярная характеристика вируса Менангл, нового парамиксовируса, поражающего свиней, летучих мышей и людей» . Вирусология . 283 (2): 358–73. дои : 10.1006/виро.2001.0893 . ПМИД 11336561 .
- ^ Перейти обратно: а б Яманака, А.; Ивакири, А.; Ёсикава, Т.; Сакаи, К.; Сингх, Х.; Химедзи, Д.; Кикучи, И.; Уэда, А.; Ямамото, С.; Миура, М.; Сиояма, Ю.; Кавано, К.; Нагайши, Т.; Сайто, М.; Миномо, М.; Ивамото, Н.; Хидака, Ю.; Сохма, Х.; Кобаяши, Т.; Канаи, Ю.; Кавагиси, Т.; Нагата, Н.; Фукуси, С.; Мизутани, Т.; Тани, Х.; Танигучи, С.; Фукума, А.; Симодзима, М.; Коран, И.; и др. (2014). «Завозной случай острой инфекции дыхательных путей, связанной с представителем вида Ортореовирус Нельсона Бэя» . ПЛОС ОДИН 9 (3): е92777. Бибкод : 2014PLoSO... 992777Y дои : 10.1371/journal.pone.0092777 . ПМЦ 3965453 . ПМИД 24667794 .
- ^ Смит, И.; Ван, Л. (2013). «Летучие мыши и их виром: важный источник новых вирусов, способных заражать человека» . Современное мнение в вирусологии . 3 (1): 84–91. дои : 10.1016/j.coviro.2012.11.006 . ПМК 7102720 . ПМИД 23265969 .
- ^ Кэмпбелл-Маклеод, П. (2013). «Наллавилли-Сядь (и слушай): Истории времен снов - устная традиция». В Макдональде, MR (ред.). Традиционное рассказывание историй сегодня: международный справочник . Рутледж. стр. 158–159. ISBN 978-1135917142 .
- ^ Слон, TH, изд. (2001). Тысяча одна ночь Папуа-Новой Гвинеи: Сказки 1972–1985 годов . Том. 1. Масалай Пресс. ISBN 978-0971412712 .
- ^ Пэриш, С.; Ричардс, Г.; Холл, Л. (2012). Естественная история австралийских летучих мышей: работа в ночную смену . Издательство Csiro. ISBN 978-0643103764 .
- ^ Перейти обратно: а б Британский музей. Отдел британских и средневековых древностей и этнографии; Джойс, штат Калифорния; Далтон, О.М. (1910). Справочник по этнографическим коллекциям . Отпечатано по заказу попечителей. п. 125.
- ^ Махрей, Роберт (1899). «Странные виды денег» . Ежемесячный иллюстрированный журнал Хармсворта . 1 : 639–641.
- ^ Вернесс; ХБ (2003). Континуальная энциклопедия местного искусства: мировоззрение, символизм и культура Африки, Океании и Северной Америки . А&С Черный. п. 31. ISBN 978-0826414656 .
- ^ Перейти обратно: а б Чой, Чарльз (16 октября 2017 г.). «В Макире зубы летучей лисицы — это валюта… И это может спасти вид» . Обнаружить . Кальмбах Медиа. Архивировано из оригинала 12 июня 2018 года . Проверено 2 июня 2018 г.
- ^ Лавери, Тайрон Х; Фаси, Джон (2017). «Покупка сквозь зубы: традиционная валюта и сохранение летучих лисиц Pteropus spp. на Соломоновых островах» . Орикс . 53 (3): 1–8. дои : 10.1017/S0030605317001004 . S2CID 90302071 .
- ^ Азиз, SA; Оливал, К.Дж.; Бумрунгсри, С.; Ричардс, GC; Рэйси, Пенсильвания (2016). «Конфликт между крылатоногими летучими мышами и производителями фруктов: виды, законодательство и смягчение последствий». В Фойгте, К.; Кингстон, Т. (ред.). Летучие мыши в антропоцене: сохранение летучих мышей в меняющемся мире . Спрингер. дои : 10.1007/978-3-319-25220-9_13 . ISBN 978-3-319-25220-9 . S2CID 111056244 .
- ^ «Таксономия=Птероподиды» . Красный список исчезающих видов МСОП . 2019 . Проверено 19 мая 2019 г.
- ^ Милденштейн, Т.; Танши, И.; Рэйси, Пенсильвания (2016). «Эксплуатация летучих мышей для добычи мяса диких животных и лекарств». Летучие мыши в антропоцене: сохранение летучих мышей в меняющемся мире . Спрингер. п. 327. дои : 10.1007/978-3-319-25220-9_12 . ISBN 978-3-319-25218-6 . S2CID 130038936 .
- ^ Винсенот, CE; Кояма, Л.; Руссо, Д. (2015). «На грани угрозы? Первый отчет о неожиданных антропогенных факторах сокращения численности летучей лисицы Рюкю (Pteropus dasymallus) в Японии». Биология млекопитающих – Zeitschrift für Säugetierkunde . 80 (4): 273. doi : 10.1016/j.mambio.2015.03.003 .
- ^ Винсенот, CE; Флоренс, ФБВ; Кингстон, Т. (2017). «Можем ли мы защитить островных летучих лисиц?». Наука . 355 (6332): 1368–1370. Бибкод : 2017Sci...355.1368V . дои : 10.1126/science.aam7582 . ПМИД 28360279 . S2CID 206657272 .
- ^ Макилви, AP; Мартин, Л. (2002). «О способности к размножению австралийских летучих лисиц (Pteropus spp., Megachiroptera)». Австралийский зоолог . 32:93 . дои : 10.7882/AZ.2002.008 .
- ^ Мартин, Л. (2011). «Являются ли фрукты, которые вы едите, благоприятными для летучей лисицы? Влияние садовых электросетей на австралийских летучих лисиц (Pteropus spp., Megachiroptera)». Биология и охрана австралийских летучих мышей . Королевское зоологическое общество Нового Южного Уэльса. стр. 380–390. дои : 10.7882/FS.2011.039 . ISBN 978-0-9803272-4-3 .
- ^ Хлопицкий, К. (28 октября 2016 г.). «Электрические провода угрожают летучим лисицам и их детенышам» . «Дейли телеграф» . Проверено 26 июня 2018 г.
- ^ Вельберген, Дж.А.; Клозе, С.М.; Маркус, Н.; Эби, П. (2008). «Изменение климата и влияние экстремальных температур на австралийских летучих лисиц» . Труды Королевского общества B: Биологические науки . 275 (1633): 419–425. дои : 10.1098/rspb.2007.1385 . ПМК 2596826 . ПМИД 18048286 .
- ^ Вальдес, Эрнест В. (2010). Оценка популяции марианской летучей мыши (Pteropus mariannus mariannus) в Анатахане, Саригане, Гугуане, Аламагане, Пагане, Агрихане, Асунсьоне и Мауге (PDF) (отчет). Геологическая служба США. п. 2. Архивировано из оригинала (PDF) 23 марта . Получено 24 августа.
- ^ Флеминг, TH; Рэйси, Пенсильвания, ред. (2010). Островные летучие мыши: эволюция, экология и сохранение . Издательство Чикагского университета . п. 415. ИСБН 9780226253312 .
Внешние ссылки
[ редактировать ] СМИ, связанные с Pteropodidae, на Викискладе?
СМИ, связанные с Pteropodidae, на Викискладе?  Данные, относящиеся к Pteropodidae , в Wikispecies
Данные, относящиеся к Pteropodidae , в Wikispecies